Insights on the Functions and Ecophysiological Relevance of the Diverse Carbonic Anhydrases in Microalgae


Abstract
:1. Introduction
2. The Need for CO2-Concentrating Mechanisms in Microalgae
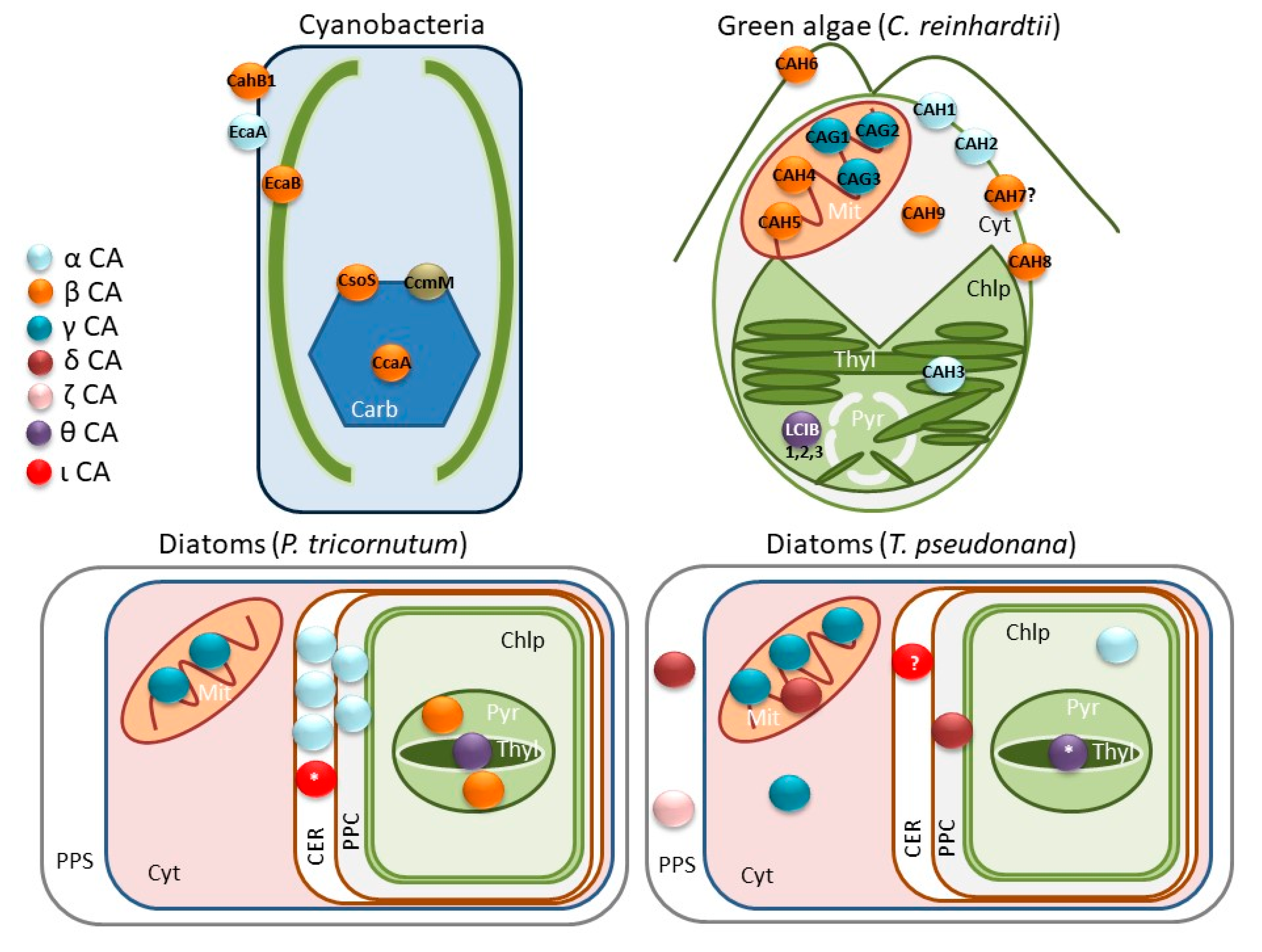
3. Diversity of Carbonic Anhydrases in Microalgae and Their Importance in the CCM
4. Regulation of CA Expression
5. Other Roles of CAs in Microalgae
6. Microalgal CAs and the Ecology of Aquatic Environments
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Nonoyama, T.; Kazamia, E.; Nawaly, H.; Gao, X.; Tsuji, Y.; Matsuda, Y.; Bowler, C.; Tanaka, T.; Dorrell, R. Metabolic Innovations Underpinning the Origin and Diversification of the Diatom Chloroplast. Biomolecules 2019, 9, 322. [Google Scholar] [CrossRef] [PubMed]
- Soo, R.M.; Hemp, J.; Parks, D.H.; Fischer, W.W.; Hugenholtz, P. On the origins of oxygenic photosynthesis and aerobic respiration in Cyanobacteria. Science 2017, 1440, 1436–1440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, H.S.; Hackett, J.D.; Ciniglia, C.; Pinto, G.; Bhattacharya, D. A molecular timeline for the origin of photosynthetic eukaryotes. Mol. Biol. Evol. 2004, 21, 809–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorrell, R.G.; Bowler, C. Secondary Plastids of Stramenopiles. In Advances in Botanical Research, 1st ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2017. [Google Scholar] [CrossRef]
- Keeling, P.J. The endosymbiotic origin, diversification and fate of plastids. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 729–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhola, V.; Swalaha, F.; Kumar, R.R.; Singh, M.; Bux, F. Overview of the potential of microalgae for CO2 sequestration. Int. J. Environ. Sci. Technol. 2014, 11, 2103–2118. [Google Scholar] [CrossRef] [Green Version]
- Raven, J.A.; Giordano, M.; Beardall, J.; Maberly, S.C. Algal evolution in relation to atmospheric mechanisms and carbon oxidation cycles. Philos. Trans. R. Soc. 2012, 367, 493–507. [Google Scholar] [CrossRef] [Green Version]
- Raven, A. Photosynthetic and non-Photosynthetic roles of carbonic anhydrase in algae and cyanobacteria. Phycologia 1995, 34, 93–101. [Google Scholar] [CrossRef]
- Lionetto, M.G.; Caricato, R.; Giordano, M.E.; Schettino, T. The complex relationship between metals and carbonic anhydrase: New insights and perspectives. Int. J. Mol. Sci. 2016, 17, 127. [Google Scholar] [CrossRef] [Green Version]
- DiMario, R.J.; Machingura, M.C.; Waldrop, G.L.; Moroney, J.V. The many types of carbonic anhydrases in photosynthetic organisms. Plant Sci. 2018, 268, 11–17. [Google Scholar] [CrossRef]
- Kikutani, S.; Nakajima, K.; Nagasato, C.; Tsuji, Y.; Miyatake, A.; Matsuda, Y. Thylakoid luminal θ-Carbonic anhydrase critical for growth and photosynthesis in the marine diatom Phaeodactylum tricornutum. Proc. Natl. Acad. Sci. USA 2016, 113, 9828–9833. [Google Scholar] [CrossRef] [Green Version]
- Jensen, E.L.; Clement, R.; Kosta, A.; Maberly, S.C.; Gontero, B. A new widespread subclass of carbonic anhydrase in marine phytoplankton. ISME J. 2019, 13, 2094–2106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nocentini, A.; Supuran, C.T. Carbonic anhydrases: An overview. In Carbonic Anhydrases; Elsevier Inc.: Amsterdam, The Netherlands, 2019. [Google Scholar] [CrossRef]
- Supuran, C.T. Carbonic Anhydrases and Metabolism. Metabolites 2018, 8, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morel, F.M.M.; Lam, P.J.; Saito, M.A. Trace Metal Substitution in Marine Phytoplankton. Annu. Rev. Earth Planet. Sci. 2020, 48, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Del Prete, S.; Vullo, D.; Fisher, G.M.; Andrews, K.T.; Poulsen, S.A.; Capasso, C.; Supuran, C.T. Discovery of a new family of carbonic anhydrases in the malaria pathogen Plasmodium falciparum-The η-Carbonic anhydrases. Bioorganic Med. Chem. Lett. 2014, 24, 4389–4396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, T.W.; Saito, M.A.; George, G.N.; Pickering, I.J.; Prince, R.C.; Morel, F.M.M. A cadmium enzyme from a marine diatom. Nature 2005, 435, 42. [Google Scholar] [CrossRef]
- DiMario, R.J.; Clayton, H.; Mukherjee, A.; Ludwig, M.; Moroney, J.V. Plant Carbonic Anhydrases: Structures, Locations, Evolution, and Physiological Roles. Mol. Plant. 2017, 10, 30–46. [Google Scholar] [CrossRef] [Green Version]
- Whittington, D.A.; Waheed, A.; Ulmasov, B.; Shah, G.N.; Grubb, J.H.; Sly, W.S.; Christianson, D.W. Crystal structure of the dimeric extracellular domain of human carbonic anhydrase XII, a bitopic membrane protein overexpressed in certain cancer tumor cells. Proc. Natl. Acad. Sci. USA 2002, 98, 9545–9550. [Google Scholar] [CrossRef] [Green Version]
- Supuran, C.T.; Capasso, C. An overview of the bacterial carbonic anhydrases. Metabolites 2017, 7, 56. [Google Scholar] [CrossRef] [Green Version]
- Yee, D.; Morel, F.M.M. In vivo substitution in carbonic of zinc by cobalt of a marine anhydrase diatom. Limnol. Oceanogr. 2011, 41, 573–577. [Google Scholar] [CrossRef]
- McGinn, P.J.; Morel, F.M.M. Expression and regulation of carbonic anhydrases in the marine diatom Thalassiosira pseudonana and in natural phytoplankton assemblages from Great Bay, New Jersey. Physiol. Plant. 2008, 133, 78–91. [Google Scholar] [CrossRef]
- Xu, Y.; Feng, L.; Jeffrey, P.D.; Shi, Y.; Morel, F.M.M.M. Structure and metal exchange in the cadmium carbonic anhydrase of marine diatoms. Nature 2008, 452, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Sun, J.; Wunder, T.; Tang, D.; Cousins, A.B.; Sze, S.K.; Mueller-Cajar, O.; Gao, Y.-G. Structural insights into the LCIB protein family reveals a new group of β-Carbonic anhydrases. Proc. Natl. Acad. Sci. USA 2016, 113, 14716–14721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Lee, K.; Lee, H.; Kim, M. The effect of seawater CO2 concentration on growth of a natural phytoplankton assemblage in a controlled mesocosm experiment. Limnol. Oceanogr. 2006, 51, 1629–1636. [Google Scholar] [CrossRef] [Green Version]
- Gattuso, J.; Gao, K.; Lee, K.; Rost, B.; Schulz, K.G. Approaches and tools to manipulate the carbonate chemistry. In Guid. to Best Pract. Ocean Acidif. Res. Data Report; Riebesell, U., Fabry, V., Hansson, L., Gattuso, J., Eds.; Publications Office ofthe European Union: Luxembourg, 2010; pp. 41–52. [Google Scholar]
- Gao, K.; Campbell, D.A. Photophysiological responses of marine diatoms to elevated CO2 and decreased pH: A review. Funct. Plant Biol. 2014, 41, 449–459. [Google Scholar] [CrossRef]
- Badger, M.R.; Andrews, T.J.; Whitney, S.M.; Ludwig, M.; Yellowlees, D.C.; Leggat, W.; Price, G.D. The diversity and coevolution of Rubisco, plastids, pyrenoids, and chloroplast-Based CO2-Concentrating mechanisms in algae. Can. J. Bot. 1998, 76, 1052–1071. [Google Scholar] [CrossRef]
- Young, J.N.; Heureux, A.M.C.; Sharwood, R.E.; Rickaby, R.E.M.; Morel, F.M.M.; Whitney, S.M. Large variation in the Rubisco kinetics of diatoms reveals diversity among their carbon-Concentrating mechanisms. J. Exp. Bot. 2016, 67, 3445–3456. [Google Scholar] [CrossRef] [Green Version]
- Heureux, A.M.C.; Young, J.N.; Whitney, S.M.; Eason-hubbard, M.R.; Lee, R.B.Y.; Sharwood, R.E.; Rickaby, R.E.M. The role of Rubisco kinetics and pyrenoid morphology in shaping the CCM of haptophyte microalgae. J. Exp. Bot. 2017, 68, 3959–3969. [Google Scholar] [CrossRef]
- Husic, D.W.; Husic, H.D.; Tolbert, N.E.; Black, C.C. The oxidative photosynthetic carbon cycle or C2 cycle. CRC Crit. Rev. Plant Sci. 1987, 5, 45–100. [Google Scholar] [CrossRef]
- Maberly, S.C.; Gontero, B. Ecological imperatives for aquatic carbon dioxide-Concentrating mechanisms. J. Exp. Bot. 2017, 68, 3797–3814. [Google Scholar] [CrossRef]
- Raven, J.A.; Ball, L.A.; Beardall, J.; Giordano, M.; Maberly, S.C. Algae lacking CO2 concentrating mechanisms. Can. J. Bot. 2011, 83, 879–890. [Google Scholar] [CrossRef]
- Matsuda, Y.; Nakajima, K.; Tachibana, M. Recent progresses on the genetic basis of the regulation of CO2 acquisition systems in response to CO2 concentration. Photosynth. Res. 2011, 109, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Hopkinson, B.M.; Dupont, C.L.; Allen, A.E.; Morel, F.M.M. Efficiency of the CO2-Concentrating mechanism of diatoms. Proc. Natl. Acad. Sci. USA 2011, 108, 3830–3837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giordano, M.; Beardall, J.; Raven, J.A. CO2 concentrating mechanisms in algae: Mechanisms, Environmental Modulation, and Evolution. Annu. Rev. Plant Biol. 2005, 56, 99–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moroney, J.V.; Somanchi, A. How Do Algae Concentrate CO2 to Increase the Efficiency of Photosynthetic Carbon Fixation? Plant Physiol. 1999, 119, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Moroney, J.V.; Ynalvez, R.A. Proposed carbon dioxide concentrating mechanism in Chlamydomonas reinhardtii. Eukaryot. Cell 2007, 6, 1251–1259. [Google Scholar] [CrossRef] [Green Version]
- Badger, M.R.; Price, G.D. CO2 concentrating mechanisms in cyanobacteria: Molecular components, their diversity and evolution. J. Exp. Bot. 2003, 54, 609–622. [Google Scholar] [CrossRef]
- Hopkinson, B.M.; Dupont, C.L.; Matsuda, Y. The physiology and genetics of CO2 concentrating mechanisms in model diatoms. Curr. Opin. Plant Biol. 2016, 31, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Bowes, G.; Rao, S.K.; Estavillo, G.M.; Reiskind, J.B. C4 mechanisms in aquatic angiosperms: Comparisons with terrestrial C4 systems. Funct. Plant Biol. 2002, 29, 379–392. [Google Scholar] [CrossRef]
- Zhang, Y.; Yin, L.; Jiang, H.-S.; Li, W.; Gontero, B.; Maberly, S.C. Biochemical and biophysical CO2 concentrating mechanisms in two species of freshwater macrophyte within the genus Ottelia (Hydrocharitaceae). Photosynth. Res. 2014, 121, 285–297. [Google Scholar] [CrossRef] [Green Version]
- Shao, H.; Gontero, B.; Maberly, S.C.; Jiang, H.S.; Cao, Y.; Li, W.; Huang, W.M. Responses of Ottelia alismoides, an aquatic plant with three CCMs, to variable CO2 and light. J. Exp. Bot. 2017, 68, 3985–3995. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Fan, X.; Zhang, X.; Xu, D.; Mou, S.; Cao, S.; Zheng, Z.; Miao, J.; Ye, N. Evidence of Coexistence of C3 and C4 Photosynthetic Pathways in a Green-Tide-Forming Alga, Ulva prolifera. PLoS ONE 2012, 7, e37438. [Google Scholar] [CrossRef] [PubMed]
- Roberts, K.; Granum, E.; Leegood, R.C.; Raven, J.A. C3 and C4 Pathways of Photosynthetic Carbon Assimilation in Marine Diatoms Are under Genetic, Not Environmental, Control. Plant Physiol. 2007, 145, 230–235. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, R.; Kikutani, S.; Mahardika, A.; Matsuda, Y. Localization of enzymes relating to C4 organic acid metabolisms in the marine diatom, Thalassiosira pseudonana. Photosynth. Res. 2014, 121, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Clement, R.; Jensen, E.; Prioretti, L.; Maberly, S.C.; Gontero, B. Diversity of CO2 concentrating mechanisms and responses to CO2 concentration in marine and freshwater diatoms. J. Exp. Bot. 2017, 68, 3925–3935. [Google Scholar] [CrossRef] [PubMed]
- Ewe, D.; Tachibana, M.; Kikutani, S.; Gruber, A.; Río Bártulos, C.; Konert, G.; Kaplan, A.; Matsuda, Y.; Kroth, P.G. The intracellular distribution of inorganic carbon fixing enzymes does not support the presence of a C4 pathway in the diatom Phaeodactylum tricornutum. Photosynth. Res. 2018, 137, 263–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clement, R.; Dimnet, L.; Maberly, S.C.; Gontero, B. The nature of the CO2-Concentrating mechanisms in a marine diatom, Thalassiosira pseudonana. New Phytol. 2016, 209, 1417–1427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haimovich-dayan, M.; Garfinkel, N.; Ewe, D.; Marcus, Y.; Gruber, A.; Wagner, H.; Kroth, P.G.; Kaplan, A. The role of C4 metabolism in the marine diatom Phaeodactylum tricornutum. New Phytol. 2013, 197, 177–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spalding, M.H. Microalgal carbon-Dioxide-Concentrating mechanisms: Chlamydomonas inorganic carbon transporters. J. Exp. Bot. 2007, 59, 1463–1473. [Google Scholar] [CrossRef]
- Price, G.D. Inorganic carbon transporters of the cyanobacterial CO2 concentrating mechanism. Photosynth. Res. 2011, 109, 47–57. [Google Scholar] [CrossRef]
- Klanchui, A.; Cheevadhanarak, S.; Prommeenate, P.; Meechai, A. Exploring Components of the CO2-Concentrating Mechanism in Alkaliphilic Cyanobacteria Through Genome-Based Analysis. Comput. Struct. Biotechnol. J. 2017, 15, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Poschenrieder, C.; Fern, A.; Ter, J.; Barcel, J. Transport and Use of Bicarbonate in Plants: Current Knowledge and Challenges Ahead. Int. J. Mol. Sci. 2018, 19, 1352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omata, T.; Price, G.D.; Badger, M.R.; Okamura, M.; Gohta, S.; Ogawa, T. Identification of an ATP-Binding cassette transporter involved in bicarbonate uptake in the cyanobacterium Synechococcus sp. Strain PCC 7942. Proc. Natl. Acad. Sci. USA 1999, 96, 13571–13576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mcginn, P.J.; Price, G.D.; Maleszka, R.; Badger, M.R. Inorganic Carbon Limitation and Light Control the Expression of Transcripts Related to the CO2-Concentrating Mechanism in the Cyanobacterium Synechocystis sp. Strain PCC6803 1. Plant Physiol. 2003, 132, 218–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaczmarski, J.A.; Hong, N.; Mukherjee, B.; Wey, L.T.; Rourke, L.; Fo, B.; Peat, T.S.; Price, G.D.; Jackson, C.J. Structural Basis for the Allosteric Regulation of the SbtA Bicarbonate Transporter by the PII-Like Protein, SbtB, from Cyanobium sp. PCC7001. Biochemistry 2019, 58, 5030–5039. [Google Scholar] [CrossRef]
- Price, G.D.; Woodger, F.J.; Badger, M.R.; Howitt, S.M.; Tucker, L. Identification of a SulP-Type bicarbonate transporter in marine cyanobacteria. Proc. Natl. Acad. Sci. USA 2004, 101, 18228–18332. [Google Scholar] [CrossRef] [Green Version]
- Badger, M.R.; Price, G.D.; Long, B.M.; Woodger, F.J. The environmental plasticity and ecological genomics of the cyanobacterial CO2 concentrating mechanism. J. Exp. Bot. 2006, 57, 249–265. [Google Scholar] [CrossRef]
- Shibata, M.; Ohkawa, H.; Kaneko, T.; Fukuzawa, H.; Tabata, S.; Kaplan, A.; Ogawa, T. Distinct constitutive and low-CO2 -induced CO2 uptake systems in cyanobacteria: Genes involved and their phylogenetic relationship with homologous genes in other organisms. Proc. Natl. Acad. Sci. USA 2001, 98, 2–7. [Google Scholar] [CrossRef] [Green Version]
- Sandrini, G.; Tann, R.P.; Schuurmans, J.M.; Van Beusekom, S.A.M.; Matthijs, H.C.P.; Huisman, J. Diel Variation in Gene Expression of the CO2-Concentrating Mechanism during a Harmful Cyanobacterial Bloom. Front. Microbiol. 2016, 7, 1–16. [Google Scholar] [CrossRef]
- Machingura, M.C.; Bajsa-hirschel, J.; Laborde, S.M.; Schwartzenburg, J.B.; Mukherjee, B.; Mukherjee, A.; Pollock, S.V.; Förster, B.; Price, G.D.; Moroney, J.V. Identification and characterization of a solute carrier, CIA8, involved in inorganic carbon acclimation in Chlamydomonas reinhardtii. J. Exp. Bot. 2017, 68, 3879–3890. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Spalding, M.H. Acclimation to very low CO2: Contribution of limiting CO2 inducible proteins, LCIB and LCIA, to onorganic carbon uptake in Chlamydomonas reinhardtii. Plant Physiol. 2014, 166, 2040–2050. [Google Scholar] [CrossRef] [Green Version]
- Pollock, S.V.; Prout, D.L.; Godfrey, A.C.; Lemaire, S.D.; Moroney, J.V. The Chlamydomonas reinhardtii proteins Ccp1 and Ccp2 are required for long-term growth, but are not necessary for efficient photosynthesis, in a low-CO2 environment. Plant Mol. Biol. 2004, 56, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, K.; Tanaka, A.; Matsuda, Y. SLC4 family transporters in a marine diatom directly pump bicarbonate from seawater. Proc. Natl. Acad. Sci. USA 2013, 110, 1767–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, Y.; Hopkinson, B.M.; Nakajima, K.; Dupont, C.L.; Tsuji, Y. Mechanisms of carbon dioxide acquisition and CO2 sensing in marine diatoms: A gateway to carbon metabolism. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meldrum, N.U.; Roughton, F.J.W. The state of carbon dioxide in blood. J. Physiol. 1933, 80, 143–170. [Google Scholar] [CrossRef] [Green Version]
- Chegwidden, W.R.; Carter, N.D. Introduction to the carbonic anhydrases. In The Carbonic Anhydrases; Chegwidden, W.R., Carter, N.D., Edwards, Y.H., Eds.; Birkhäuser: Basel, Switzerland, 2000; pp. 13–28. [Google Scholar] [CrossRef]
- Forster, R.E. Remarks on the discovery of carbonic anhydrase. In The Carbonic Anhydrases; Chegwidden, W.R., Carter, N.D., Edwards, Y.H., Eds.; Birkhäuser: Basel, Switzerland, 2000; pp. 1–11. [Google Scholar] [CrossRef]
- Lomelino, C.L.; Andring, J.T.; McKenna, R. Crystallography and Its Impact on Carbonic Anhydrase Research. Int. J. Med. Chem. 2018, 2018, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Cuesta-Seijo, J.A.; Borchert, M.S.; Navarro-Poulsen, J.C.; Schnorr, K.M.; Mortensen, S.B.; Lo Leggio, L. Structure of a dimeric fungal α-Type carbonic anhydrase. FEBS Lett. 2011, 585, 1042–1048. [Google Scholar] [CrossRef] [Green Version]
- Ishida, S.; Muto, S.; Miyachi, S. Structural analysis of periplasmic carbonic anhydrase 1 of Chlamydomonas reinhardtii. Eur. J. Biochem. 1993, 214, 9–16. [Google Scholar] [CrossRef]
- Moroney, J.V.; Ma, Y.; Frey, W.D. The carbonic anhydrase isoforms of Chlamydomonas reinhardtii: Intracellular location, expression, and physiological roles. Photosynth. Res. 2011, 109, 133–149. [Google Scholar] [CrossRef]
- Aspatwar, A.; Haapanen, S.; Parkkila, S. An update on the metabolic roles of carbonic anhydrases in the model alga Chlamydomonas reinhardtii. Metabolites 2018, 8, 22. [Google Scholar] [CrossRef] [Green Version]
- So, A.K.-C.; Espie, G.S. Cyanobacterial carbonic anhydrases. Can. J. Bot. 2005, 83, 721–734. [Google Scholar] [CrossRef]
- Kupriyanova, E.V.; Sinetova, M.A.; Bedbenov, V.S.; Pronina, N.A.; Los, D.A. Putative extracellular α-Class carbonic anhydrase, EcaA, of Synechococcus elongatus PCC 7942 is an active enzyme: A sequel to an old story. Metabolites 2018, 164, 576–586. [Google Scholar] [CrossRef] [PubMed]
- Gee, C.W.; Niyogi, K.K. The carbonic anhydrase CAH1 is an essential component of the carbon-Concentrating mechanism in Nannochloropsis oceanica. Proc. Natl. Acad. Sci. USA 2017, 114, 4537–4542. [Google Scholar] [CrossRef] [Green Version]
- Neish, A.C. Studies on Chloroplasts: Factors Affecting Their Flocculation and the Calculation of the Chloroplast Content of Leaf Tissue From Chemical Analysis. Biochem. J. 1938, 33, 293–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimber, M.S.; Pai, E.F. The active site architecture of Pisum sativum β-Carbonic anhydrase is a mirror image of that of α-Carbonic anhydrases. EMBO J. 2000, 19, 1407–1418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cronk, J.D.; Endrizzi, J.A.; Cronk, M.R.; Neill, J.W.O. Crystal structure of E. coli β-Carbonic anhydrase, an enzyme with an unusual pH-Dependent activity. Protein Sci. 2001, 10, 911–922. [Google Scholar] [CrossRef] [PubMed]
- Covarrubias, A.S.; Bergfors, T.; Jones, T.A.; Högbom, M. Structural mechanics of the pH-Dependent activity of β-Carbonic anhydrase from Mycobacterium tuberculosis. J. Biol. Chem. 2006, 281, 4993–4999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatch, M.D.; Burnell, J.N. Carbonic Anhydrase Activity in Leaves and Its Role in the First Step of C4 Photosynthesis. Plant Physiol. 1990, 93, 825–828. [Google Scholar] [CrossRef] [Green Version]
- Fabre, N.; Reiter, I.M.; Becuwe-linka, N.; Genty, B.; Rumeau, D. Characterization and expression analysis of genes encoding α and β carbonic anhydrases in Arabidopsis. Plant Cell Environ. 2007, 30, 617–629. [Google Scholar] [CrossRef]
- DiMario, R.J.; Quebedeaux, J.C.; Longstreth, D.J.; Dassanayake, M.; Hartman, M.M.; Moroney, J.V. The Cytoplasmic Carbonic Anhydrases β CA2 and β CA4 Are Required for Optimal Plant Growth at Low CO2. Plant Physiol. 2016, 171, 280–293. [Google Scholar] [CrossRef] [Green Version]
- Kerfeld, C.A.; Melnicki, M.R. Assembly, function and evolution of cyanobacterial carboxysomes. Curr. Opin. Plant Biol. 2016, 31, 66–75. [Google Scholar] [CrossRef] [Green Version]
- So, A.K.-C.; Espie, G.S.; Williams, E.B.; Shively, J.M.; Heinhorst, S.; Cannon, G.C. A novel evolutionary lineage of carbonic anhydrase (epsilon class) is a component of the carboxysome shell. J. Bacteriol. 2004, 186, 623–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawaya, M.R.; Cannon, G.C.; Heinhorst, S.; Tanaka, S.; Williams, E.B.; Yeates, T.O.; Kerfeld, C.A. The structure of β-Carbonic anhydrase from the carboxysomal shell reveals a distinct subclass with one active site for the price of two. J. Biol. Chem. 2006, 281, 7546–7555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alber, B.E.; Ferry, J.G. A carbonic anhydrase from the archaeon Methanosarcina thermophila. Proc. Natl. Acad. Sci. USA 2006, 91, 6909–6913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacAuley, S.R.; Zimmerman, S.A.; Apolinario, E.E.; Evilia, C.; Hou, Y.M.; Ferry, J.G.; Sowers, K.R. The archetype γ-Class carbonic anhydrase (cam) contains iron when synthesized in vivo. Biochemistry 2009, 48, 817–819. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Xin, Y.; Wang, Q.; Yang, J.; Hu, H.; Xu, J. RNAi-Based targeted gene knockdown in the model oleaginous microalgae Nannochloropsis oceanica. Plant J. 2017, 89, 1236–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, S.B.; Lane, T.W.; Morel, F.M.M. Carbonic anhydrase in the marine diatom Thalassiosira weissflogii (Bacillariophyceae). J. Phycol. 1997, 33, 845–850. [Google Scholar] [CrossRef]
- Armbrust, E.V.; Berges, J.A.; Bowler, C.; Green, B.R.; Martinez, D.; Putnam, N.H.; Zhou, S.; Allen, A.E.; Apt, K.E.; Bechner, M.; et al. The genome of the diatom Thalassiosira pseudonana: Ecology, evolution, and metabolism. Science 2004, 306, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Bowler, C.; Allen, A.E.; Badger, J.H.; Grimwood, J.; Jabbari, K.; Kuo, A.; Maheswari, U.; Martens, C.; Maumus, F.; Otillar, R.P.; et al. The Phaeodactylum genome reveals the evolutionary history of diatom genomes. Nature 2008, 456, 239–244. [Google Scholar] [CrossRef]
- Samukawa, M.; Shen, C.; Hopkinson, B.M.; Matsuda, Y. Localization of putative carbonic anhydrases in the marine diatom, Thalassiosira pseudonana. Photosynth. Res. 2014, 121, 235–249. [Google Scholar] [CrossRef]
- Tachibana, M.; Allen, A.E.; Kikutani, S.; Endo, Y.; Bowler, C.; Matsuda, Y. Localization of putative carbonic anhydrases in two marine diatoms, Phaeodactylum tricornutum and Thalassiosira pseudonana. Photosynth. Res. 2011, 109, 205–221. [Google Scholar] [CrossRef]
- Alterio, V.; Langella, E.; De Simone, G.; Monti, S.M. Cadmium-Containing carbonic anhydrase CDCA1 in marine diatom Thalassiosira weissflogii. Mar. Drugs. 2015, 13, 1688–1697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christianson, D.W.; Cox, J.D. Catalysis by metal-Activated hydroxide in zinc and manganese metalloenzymes. Annu. Rev. Biochem. 1999, 68, 33–57. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Song, B.; Morel, F.M.M. Diversity of the cadmium-Containing carbonic anhydrase in marine diatoms and natural waters. Environ. Microbiol. 2007, 9, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, Y.; Nakajima, K.; Matsuda, Y. Molecular aspects of the biophysical CO2-Concentrating mechanism and its regulation in marine diatoms. J. Exp. Bot. 2017, 68, 3763–3772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamano, T.; Tsujikawa, T.; Hatano, K.; Ozawa, S.I.; Takahashi, Y.; Fukuzawa, H. Light and low-CO2-Dependent LCIBLCIC complex localization in the chloroplast supports the carbon-Concentrating mechanism in Chlamydomonas reinhardtii. Plant Cell Physiol. 2010, 51, 1453–1468. [Google Scholar] [CrossRef] [Green Version]
- Clement, R.; Lignon, S.; Mansuelle, P.; Jensen, E.; Pophillat, M.; Lebrun, R.; Denis, Y.; Puppo, C.; Maberly, S.C.; Gontero, B. Responses of the marine diatom Thalassiosira pseudonana to changes in CO2 concentration: A proteomic approach. Sci. Rep. 2017, 7, 42333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valenzuela, J.J.; López García De Lomana, A.; Lee, A.; Armbrust, E.V.; Orellana, M.V.M.V.; Baliga, N.S. Ocean acidification conditions increase resilience of marine diatoms. Nat. Commun. 2018, 9, 2328. [Google Scholar] [CrossRef]
- Blanco-Rivero, A.; Shutova, T.; Román, M.J.; Villarejo, A.; Martinez, F. Phosphorylation controls the localization and activation of the lumenal carbonic anhydrase in Chlamydomonas reinhardtii. PLoS ONE 2012, 7, e49063. [Google Scholar] [CrossRef] [Green Version]
- Ynalvez, R.A.; Xiao, Y.; Ayana, S.; Cunnusamy, K.; Moroney, J. Identification and characterization of two closely related beta-carbonic anhydrase from Chlamydomonas reinhardtii. Physiol. Plant. 2008, 133, 15–26. [Google Scholar] [CrossRef]
- Miura, K.; Yamano, T.; Yoshioka, S.; Kohinata, T.; Inoue, Y. Expression Profiling-Based Identification of CO2-Responsive Genes Regulated by CCM1 Controlling a Carbon-Concentrating Mechanism in Chlamydomonas reinhardtii. Plant Physiol. 2004, 135, 1595–1607. [Google Scholar] [CrossRef] [Green Version]
- Yoshioka, S.; Taniguchi, F.; Miura, K.; Inoue, T.; Yamano, T.; Fukuzawa, H. The Novel Myb Transcription Factor LCR1 Regulates the CO2-Responsive Gene Cah1, Encoding a Periplasmic Carbonic Anhydrase in Chlamydomonas reinhardtii. Plant Cell 2004, 16, 1466–1477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, J.N.; Morel, F.M.M. Biological oceanography: The CO2 switch in diatoms. Nat. Clim. Chang. 2015, 5, 1–2. [Google Scholar] [CrossRef]
- Ohno, N.; Inoue, T.; Yamashiki, R.; Nakajima, K.; Kitahara, Y.; Ishibashi, M.; Matsuda, Y. CO2-cAMP-Responsive cis-Elements Targeted by a Transcription Factor with CREB/ATF-Like Basic Zipper Domain in the Marine Diatom. Plant Physiol. 2012, 158, 499–513. [Google Scholar] [CrossRef] [Green Version]
- Soto, A.R.; Zheng, H.; Shoemaker, D.; Rodriguez, J.; Read, B.A.; Wahlund, T.M. Identification and preliminary characterization of two cDNAs encoding unique carbonic anhydrases from the marine alga Emiliania huxleyi. Appl. Environ. Microbiol. 2006, 72, 5500–5511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lapointe, M.; MacKenzie, T.D.B.; Morse, D. An External δ-Carbonic Anhydrase in a Free-Living Marine Dinoflagellate May Circumvent Diffusion-Limited Carbon Acquisition. Plant Physiol. 2008, 147, 1427–1436. [Google Scholar] [CrossRef] [Green Version]
- Lane, T.W.; Morel, F.M.M. Regulation of Carbonic Anhydrase Expression by Zinc, Cobalt, and Carbon Dioxide in the Marine Diatom Thalassiosira weissflogii. Plant Physiol. 2000, 123, 345–352. [Google Scholar] [CrossRef] [Green Version]
- Kikutani, S.; Tanaka, R.; Yamazaki, Y.; Hara, S.; Hisabori, T.; Kroth, P.G.; Matsuda, Y.; Yamazaki, Y.; Hara, S.; Kroth, P.G.; et al. Redox regulation of carbonic anhydrases via thioredoxin in chloroplast of the marine diatom Phaeodactylum tricornutum. J. Biol. Chem. 2012, 287, 20689–20700. [Google Scholar] [CrossRef] [Green Version]
- Rouhier, N.; Villarejo, A.; Srivastava, M.; Gelhaye, E.; Keech, O.; Droux, M.; Finkemeier, I.; Samuelsson, G.; Dietz, K.J.; Jacquot, J.-P.; et al. Identification of Plant Glutaredoxin Targets. Antioxid. Redox Signal. 2005, 7, 919–929. [Google Scholar] [CrossRef]
- Harada, H.; Matsuda, Y. Identification and characterization of a new carbonic anhydrase in the marine diatom Phaeodactylum tricornutum. Can. J. Bot. 2005, 916, 909–916. [Google Scholar] [CrossRef]
- Harada, H.; Nakatsuma, D.; Ishida, M.; Matsuda, Y.; Harada, H.; Nakatsuma, D.; Ishida, M.; Matsuda, Y. Regulation of the Expression of Intracellular β-Carbonic Anhydrase in Response to CO2 and Light in the Marine Diatom Phaeodactylum tricornutum. Plant Physiol. 2017, 139, 1041–1050. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhuang, S.; Wu, Y.; Ren, H.; Chen, F.; Lin, X.; Wang, K.; Beardall, J.; Gao, K. Ocean Acidification modulates expression of genes and physiological performance of a marine diatom. PLoS ONE 2017, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Nienaber, L.; Cave-freeman, E.; Cross, M.; Mason, L.; Bailey, U.; Amani, P.; Davis, R.A.; Taylor, P.; Hofmann, A. Chemical probing suggests redox-Regulation of the carbonic anhydrase activity of mycobacterial Rv1284. FEBS J. 2015, 282, 2708–2721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geers, C.; Gros, G. Carbon Dioxide Transport and Carbonic Anhydrase in Blood and Muscle. Physiol. Rev. 2000, 80, 681–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fegler, J. Function of Carbonic Anhydrase in Blood. Nature 1944, 153, 137–138. [Google Scholar] [CrossRef]
- Henry, R.P. The Role of Carbonic Anhydrase in Blood Ion and Acid-Base regulation. Amer. Zool. 1984, 251, 241–251. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.; Rappel, W.; Occhipinti, R.; Ries, A.; Böhmer, M.; You, L.; Xiao, C.; Engineer, C.B.; Boron, W.F.; Schroeder, J.I. Distinct Cellular Locations of Carbonic Anhydrases Mediate Carbon Dioxide Control of Stomatal Movements. Plant Physiol. 2015, 169, 1168–1178. [Google Scholar] [CrossRef]
- Kolbe, A.R.; Brutnell, T.P.; Cousins, A.B.; Studer, A.J. Carbonic Anhydrase Mutants in Zea mays Have Altered Stomatal Responses to Environmental Signals. Plant Physiol. 2018, 177, 980–989. [Google Scholar] [CrossRef] [Green Version]
- Hoang, C.V.; Chapman, K.D. Biochemical and Molecular Inhibition of Plastidial Carbonic Anhydrase Reduces the Incorporation of Acetate into Lipids in Cotton Embryos and Tobacco Cell Suspensions and Leaves. Plant Physiol. 2002, 128, 1417–1427. [Google Scholar] [CrossRef] [Green Version]
- Kavroulakis, N.; Flemetakis, E.; Aivalakis, G.; Katinakis, P. Carbon Metabolism in Developing Soybean Root Nodules: The Role of Carbonic Anhydrase. Mol. Plant Microbe Interact. 2000, 13, 14–22. [Google Scholar] [CrossRef]
- Sinetova, M.A.; Kupriyanova, E.V.; Markelova, A.G.; Allakhverdiev, S.I.; Pronina, N.A. Identification and functional role of the carbonic anhydrase Cah3 in thylakoid membranes of pyrenoid of Chlamydomonas reinhardtii. Biochim. Biophys. Acta Bioenerg. 2012, 1817, 1248–1255. [Google Scholar] [CrossRef] [Green Version]
- Sato, N.; Sonoike, K.; Tsuzuki, M.; Kawaguchi, A. Photosynthetic characteristics of a mutant of Chlamydomonas reinhardtii impaired in fatty acid desaturation in chloroplasts. Biochim. Biophys. Acta Bioenerg. 1996, 1274, 112–118. [Google Scholar] [CrossRef] [Green Version]
- Mackinder, L.C.M.; Chen, C.; Leib, R.D.; Patena, W.; Blum, S.R.; Rodman, M.; Ramundo, S.; Adams, C.M.; Jonikas, M.C. A Spatial Interactome Reveals the Anatomy of the Algal CO2 Concentrating Mechanism. Cell 2017, 171, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Choi, H., II; Hwan, Y.; Kwak, S. Quantitative analysis of the chemotaxis of a green alga, Chlamydomonas reinhardtii, to bicarbonate using diffusion-Based microfluidic device. Biomicrofluidics 2016, 10, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Clegg, M.R.; Maberly, S.C.; Jones, R.I. Chemosensory behavioural response of freshwater phytoplanktonic flagellates. Plant Cell Environ. 2004, 27, 123–135. [Google Scholar] [CrossRef]
- Giordano, M.; Norici, A.; Forssen, M.; Eriksson, M.; Raven, J.A.; Marche, P.; Bianche, V.B. An Anaplerotic Role for Mitochondrial Carbonic Anhydrase in Chlamydomonas reinhardtii. Plant Physiol. 2003, 132, 2126–2134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kroth, P.G.; Chiovitti, A.; Gruber, A.; Martin-Jezequel, V.; Mock, T.; Parker, M.S.; Stanley, M.S.; Kaplan, A.; Caron, L.; Weber, T.; et al. A model for carbohydrate metabolism in the diatom Phaeodactylum tricornutum deduced from comparative whole genome analysis. PLoS ONE 2008, 3, e1426. [Google Scholar] [CrossRef] [Green Version]
- Sunda, W.G.; Huntsman, S.A. Relationships among growth rate, cellular manganese concentrations and manganese transport kinetics in estuarine and oceanic species of the diatom Thalassiosira. J. Phycol. 1986, 22, 259–270. [Google Scholar] [CrossRef]
- Sunda, W.G.; Huntsman, S.A. Cobalt and zinc interreplacement in marine phytoplankton: Biological and geochemical implications. Limnol. Oceanogr. 1995, 40, 1404–1417. [Google Scholar] [CrossRef]
- Wolfe-Simon, F.; Starovoytov, V.; Reinfelder, J.R.; Schofield, O.; Falkowski, P.G. Localization and role of manganese superoxide dismutase in a marine diatom. Plant Physiol. 2006, 142, 1701–1709. [Google Scholar] [CrossRef] [Green Version]
- Bondarava, N.; Un, S.; Krieger-Liszkay, A. Manganese binding to the 23 kDa extrinsic protein of Photosystem II. Biochim. Biophys. Acta Bioenerg. 2007, 1767, 583–588. [Google Scholar] [CrossRef] [Green Version]
- Sunda, W.G.; Huntsman, S.A. Interactive effects of external manganese, the toxic metals copper and zinc, and light in controlling cellular manganese and growth in a coastal diatom. Limnol. Oceanogr. 1998, 43, 1467–1475. [Google Scholar] [CrossRef]
- Heindl, J.E.; Hibbing, M.E.; Xu, J.; Natarajan, R.; Buechlein, A.M.; Fuqua, C. Discrete Responses to Limitation for Iron and Manganese in Agrobacterium tumefaciens: Influence on Attachment and Biofilm Formation. J. Bacteriol. 2016, 198, 816–829. [Google Scholar] [CrossRef] [Green Version]
- Do, H.; Makthal, N.; Chandrangsu, P.; Olsen, R.J.; Helmann, J.D.; Musser, J.M.; Kumaraswami, M. Metal sensing and regulation of adaptive responses to manganese limitation by MtsR is critical for group A streptococcus virulence. Nucleic Acids Res. 2019, 1, 1–18. [Google Scholar] [CrossRef]
- Los, D.A.; Zorina, A.; Sinetova, M.; Kryazhov, S.; Mironov, K.; Zinchenko, V.V. Stress sensors and signal transducers in cyanobacteria. Sensors 2010, 10, 2386–2415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogawa, T.; Bao, D.H.; Katoh, H.; Shibata, M.; Pakrasi, H.B.; Bhattacharyya-Pakrasi, M. A two-component signal transduction pathway regulates manganese homeostasis in Synechocystis 6803, a photosynthetic organism. J. Biol. Chem. 2002, 277, 28981–28986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, M.D.; Kropat, J.; Tottey, S.; Del Campo, J.A.; Merchant, S.S. Manganese Deficiency in Chlamydomonas Results in Loss of Photosystem II and MnSOD Function, Sensitivity to Peroxides, and Secondary Phosphorus and Iron Deficiency. Plant Physiol. 2006, 143, 263–277. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| CA Class | Metal Cofactor | Organism(s) | References |
|---|---|---|---|
| α-CA | Zn2+ | Mammals, plants, algae, prokaryotes | [18,19] |
| β-CA | Zn2+ | Plants, algae, bacteria | [18] |
| γ-CA | Zn2+, Fe2+, Co2+ | Prokaryotes, plants, fungi, algae | [20] |
| δ-CA | Zn2+, Co2+ | Marine phytoplankton | [21,22] |
| ζ-CA | Cd2+, Zn2+ | Diatoms | [23] |
| η-CA | Zn2+ | Plasmodium sp | [16] |
| θ-CA | Zn2+ | Diatoms, green algae | [11,24] |
| ι-CA | Mn2+ | Marine phytoplankton | [1,12] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jensen, E.L.; Maberly, S.C.; Gontero, B. Insights on the Functions and Ecophysiological Relevance of the Diverse Carbonic Anhydrases in Microalgae. Int. J. Mol. Sci. 2020, 21, 2922. https://doi.org/10.3390/ijms21082922
Jensen EL, Maberly SC, Gontero B. Insights on the Functions and Ecophysiological Relevance of the Diverse Carbonic Anhydrases in Microalgae. International Journal of Molecular Sciences. 2020; 21(8):2922. https://doi.org/10.3390/ijms21082922
Chicago/Turabian StyleJensen, Erik L., Stephen C. Maberly, and Brigitte Gontero. 2020. "Insights on the Functions and Ecophysiological Relevance of the Diverse Carbonic Anhydrases in Microalgae" International Journal of Molecular Sciences 21, no. 8: 2922. https://doi.org/10.3390/ijms21082922
APA StyleJensen, E. L., Maberly, S. C., & Gontero, B. (2020). Insights on the Functions and Ecophysiological Relevance of the Diverse Carbonic Anhydrases in Microalgae. International Journal of Molecular Sciences, 21(8), 2922. https://doi.org/10.3390/ijms21082922