Ruthenium, Not Carbon Monoxide, Inhibits the Procoagulant Activity of Atheris, Echis, and Pseudonaja Venoms


Abstract
:1. Introduction
2. Results
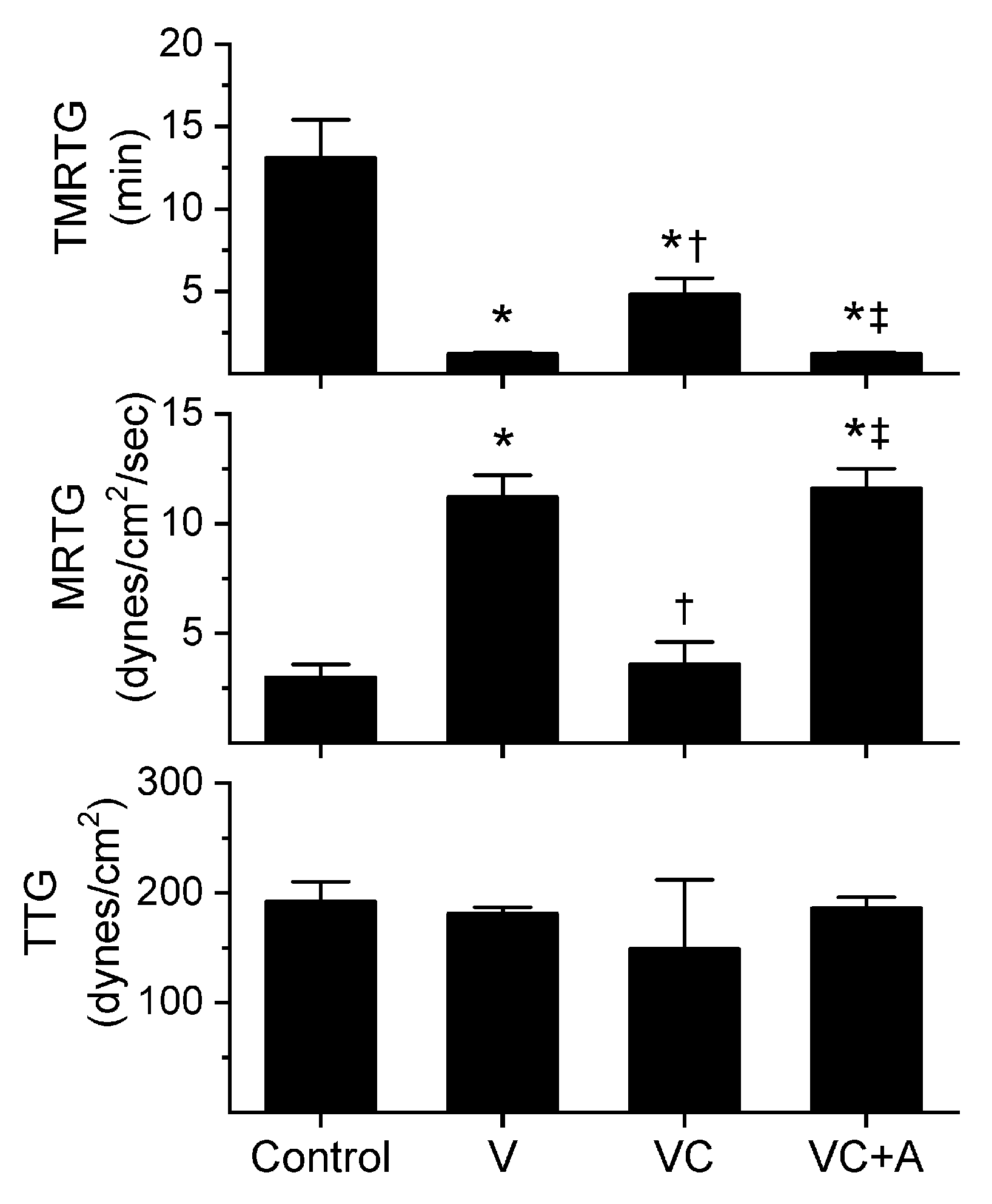
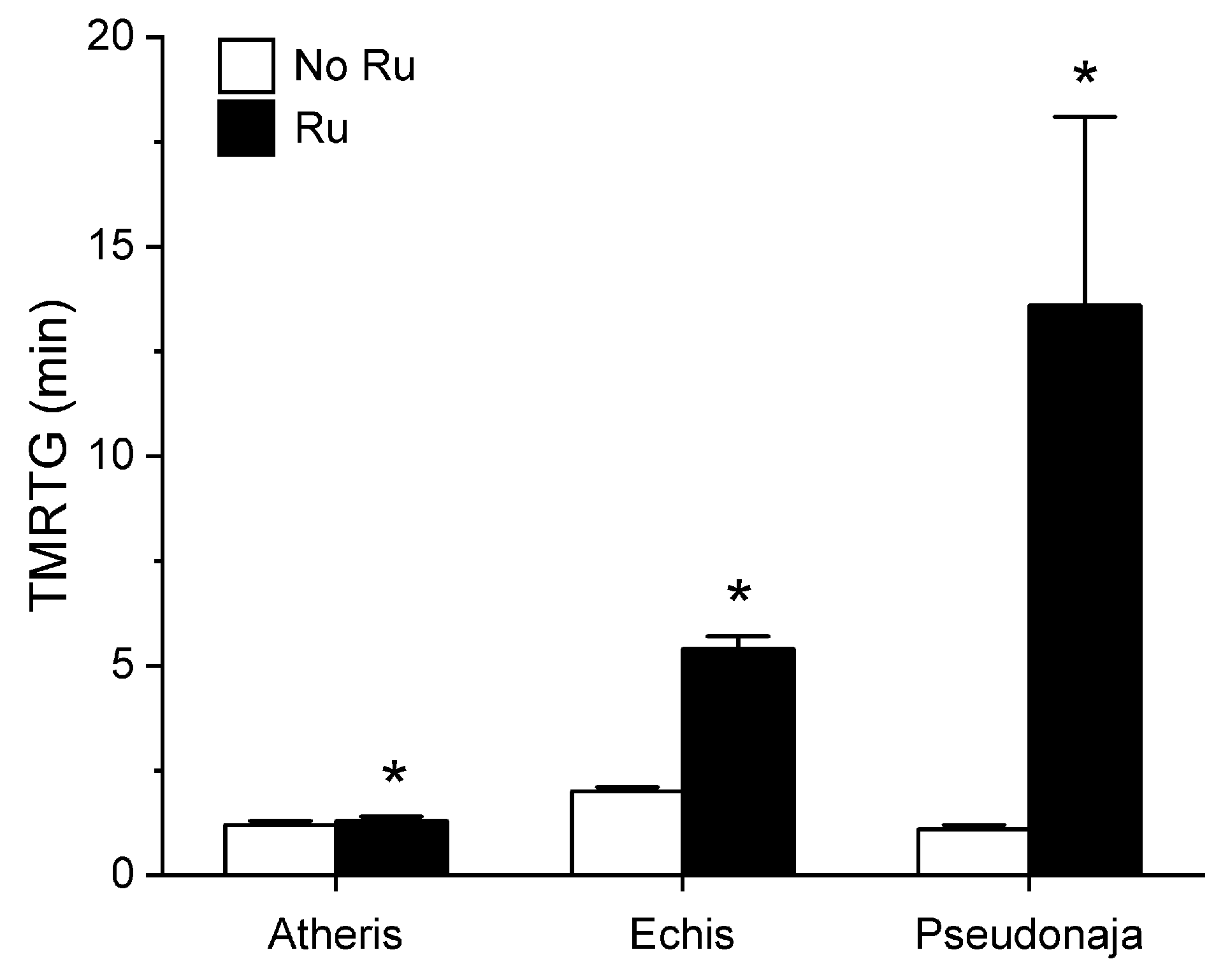
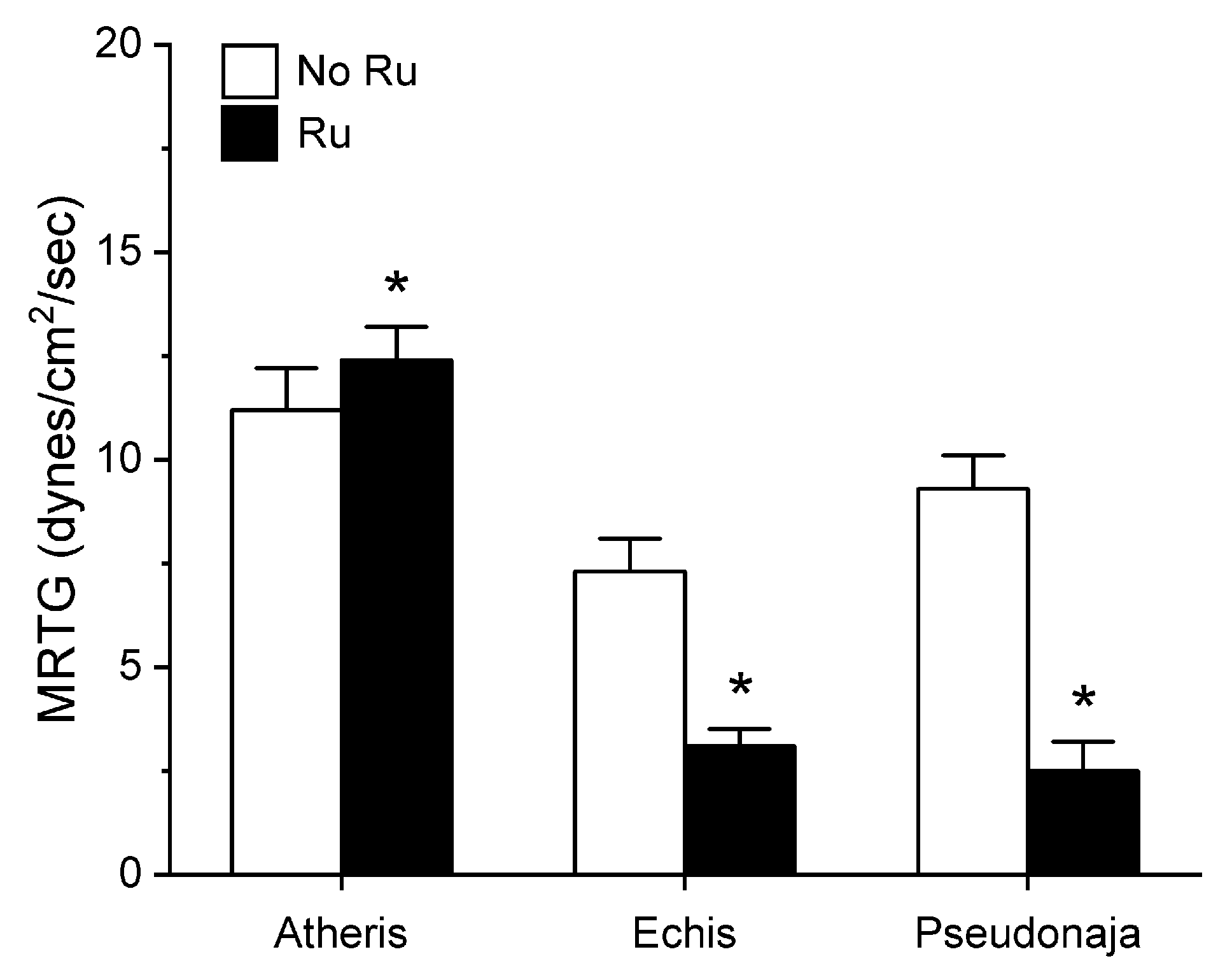
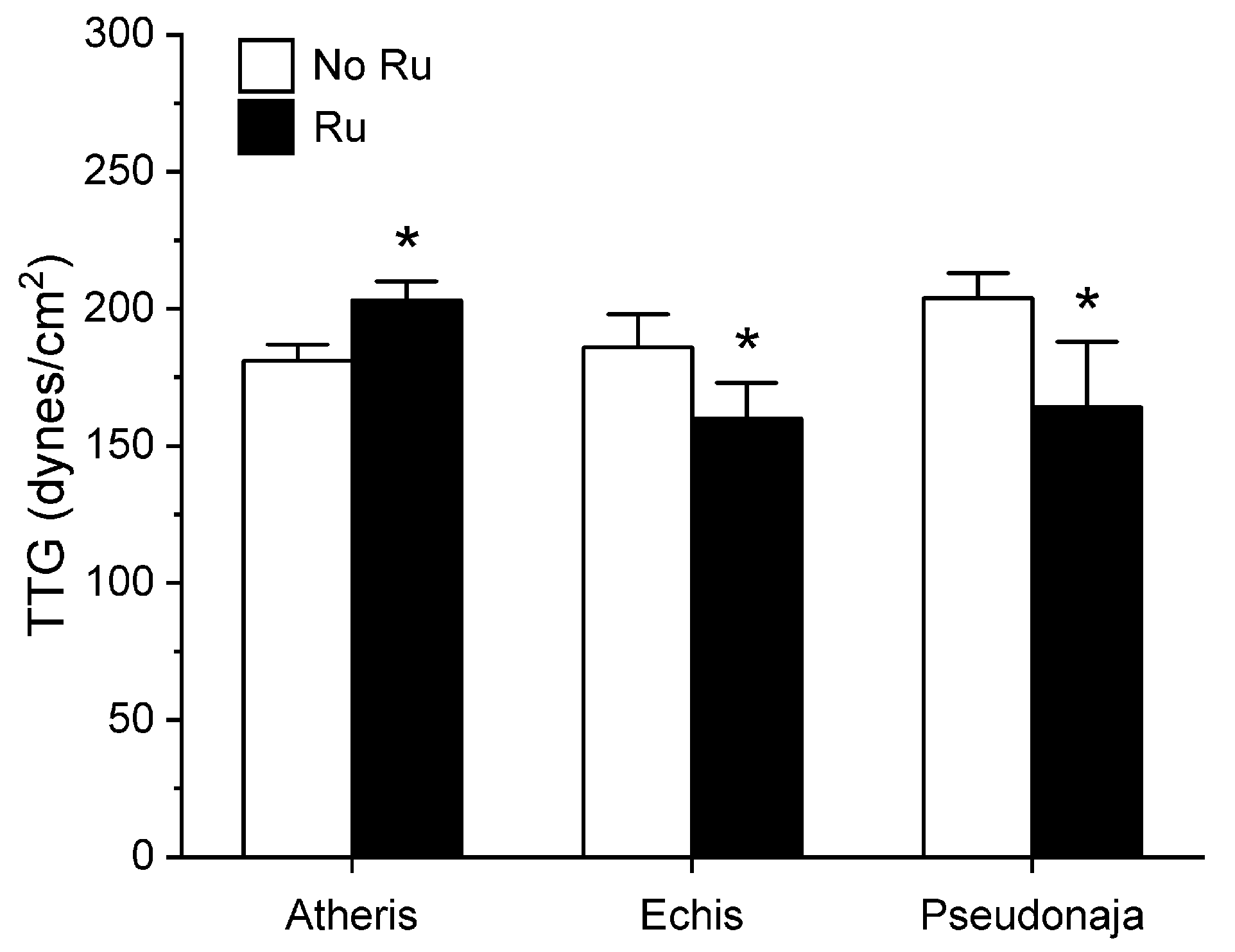
2.1. Assessment of the CO-Independent, Ru-Dependent Inhibition of CORM-2 on Procoagulant Activity of A. nitschei, E. leucogaster, and P. textilis Venoms Assessed with Thrombelastography
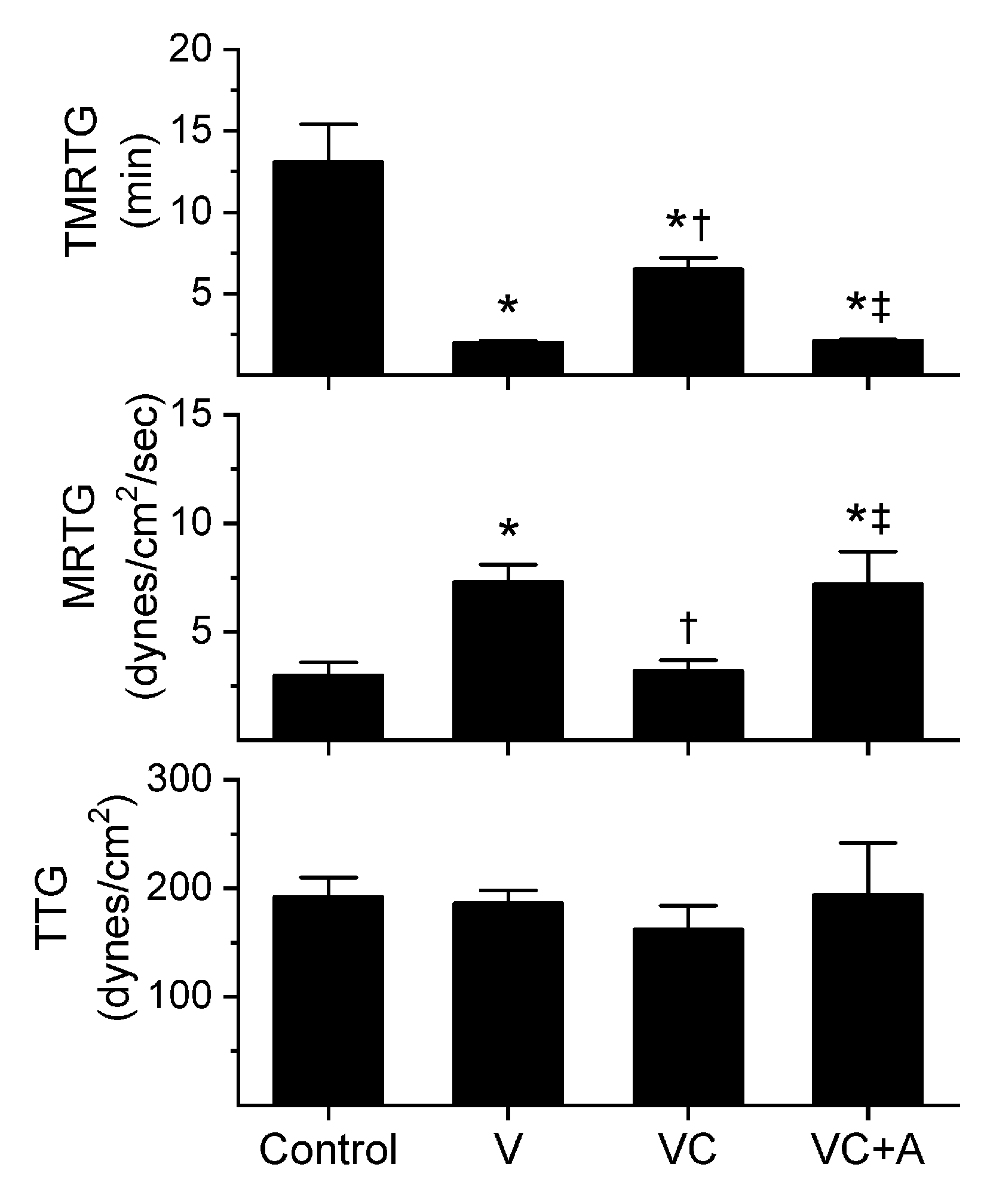
2.2. Assessment of the Effects of RuCl3 on Human Plasmatic Coagulation—Roles of Concentration and Vehicle
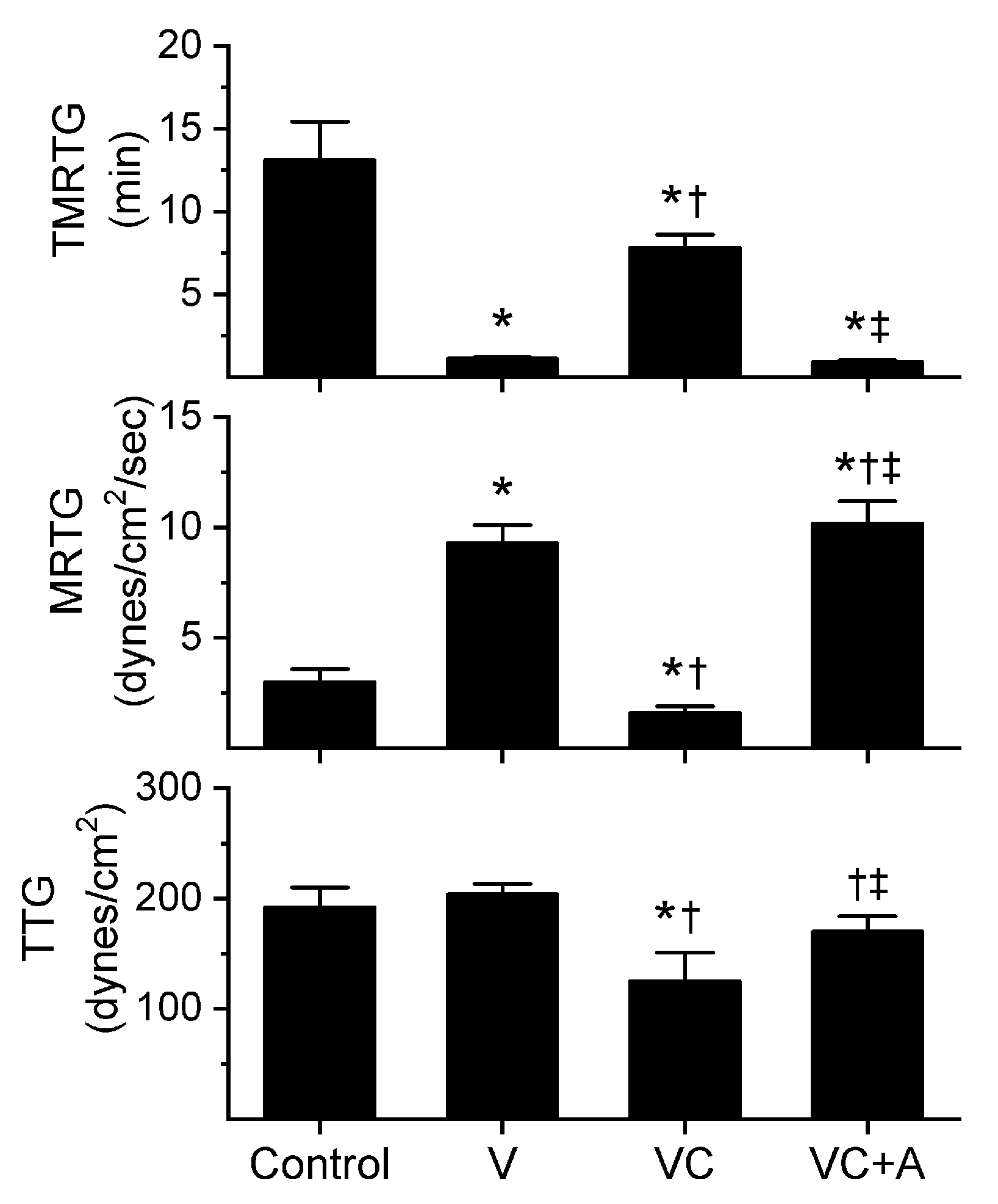
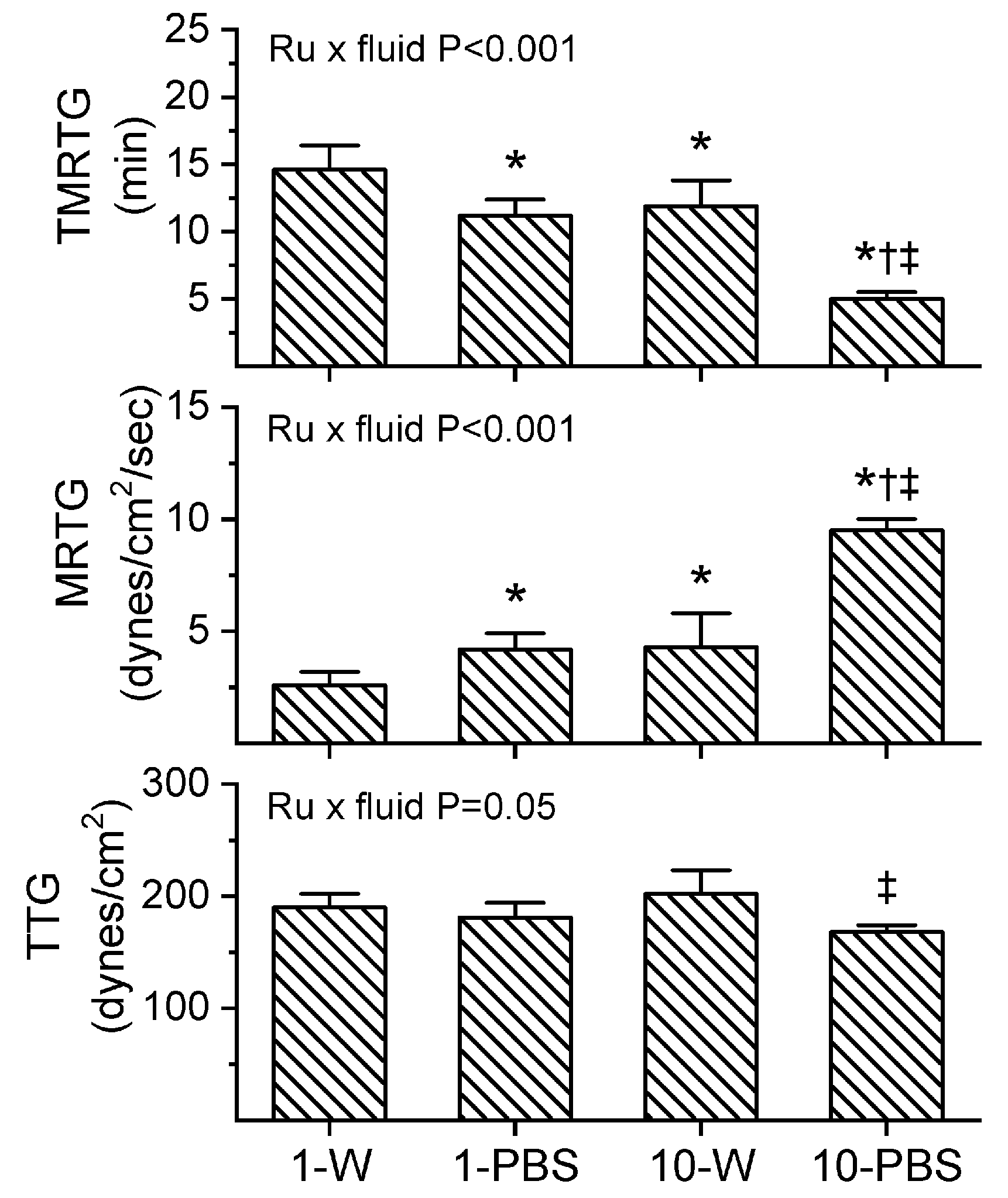
2.3. Assessment of RuCl3-Dependent Modulation of CORM-2 on the Procoagulant Activity of A. nitschei, E. leucogaster, and P. textilis Venoms Assessed with Thrombelastography
3. Discussion
4. Materials and Methods
4.1. Chemicals and Human Plasma
4.2. Thrombelastographic Analyses
4.3. CORM-2 Addition Experiments
4.4. RuCl3 Addition Experiments
4.5. Graphics and Statistical Analyses
Funding
Conflicts of Interest
Abbreviations
| CO | carbon monoxide |
| CORM | carbon monoxide releasing molecule; can have Ru, or other metal centers |
| CORM-2 | tricarbonyldichlororuthenium (II) dimer; Ru-based CORM |
| Cu | copper |
| DMSO | dimethyl sulfoxide |
| Fe | iron |
| Hg | mercury |
| MRTG | maximum rate of thrombus generation (dynes/cm2/s) |
| Pb | lead |
| PBS | phosphate buffered saline, calcium-free, pH = 7.1–7.4 |
| PLA2 | phospholipase A2 |
| Ru | ruthenium |
| RuCl3 | ruthenium chloride |
| SD | standard deviation |
| SVMP | snake venom metalloproteinase |
| SVSP | snake venom serine protease |
| TMRTG | time to maximum rate of thrombus generation (minutes) |
| TTG | total thrombus generation (dynes/cm2) |
References
- Nielsen, V.G.; Cerruti, M.A.; Valencia, O.M.; Amos, Q. Decreased snake venom metalloproteinase effects via inhibition of enzyme and modification of fibrinogen. Biometals 2016, 29, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, V.G.; Losada, P.A. Direct Inhibitory Effects of Carbon Monoxide on Six Venoms Containing Fibrinogenolytic Metalloproteinases. Basic Clin. Pharmacol. Toxicol. 2017, 120, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, V.G.; Frank, N. The kallikrein-like activity of Heloderma venom is inhibited by carbon monoxide. J. Thromb. Thrombolysis 2019, 47, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, V.G.; Bazzell, C.M. Carbon monoxide releasing molecule-2 inhibition of snake venom thrombin-like activity: Novel biochemical “brake”? J. Thromb. Thrombolysis 2017, 43, 203–208. [Google Scholar] [CrossRef]
- Nielsen, V.G.; Matika, R.W. Effects of iron and carbon monoxide on Lachesis muta muta venom-mediated degradation of plasmatic coagulation. Hum. Exp. Toxicol. 2017, 36, 727–733. [Google Scholar] [CrossRef]
- Nielsen, V.G.; Frank, N.; Matika, R.W. Effects of Heme Modulation on Ovophis and Trimeresurus Venom Activity in Human Plasma. Toxins 2018, 10, 322. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, V.G.; Frank, N.; Matika, R.W. Carbon monoxide inhibits hemotoxic activity of Elapidae venoms: Potential role of heme. Biometals 2018, 31, 51–59. [Google Scholar] [CrossRef]
- Nielsen, V.G.; Frank, N. Differential heme-mediated modulation of Deinagkistrodon, Dispholidus, Protobothrops and Pseudonaja hemotoxic venom activity in human plasma. Biometals 2018, 31, 951–959. [Google Scholar] [CrossRef]
- Nielsen, V.G.; Frank, N.; Afshar, S. De Novo Assessment and Review of Pan-American Pit Viper Anticoagulant and Procoagulant Venom Activities via Kinetomic Analyses. Toxins 2019, 11, 94. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, V.G.; Frank, N. Role of heme modulation in inhibition of Atheris, Atractaspis, Causus, Cerastes, Echis, and Macrovipera hemotoxic venom activity. Hum Exp. Toxicol. 2019, 38, 216–226. [Google Scholar] [CrossRef]
- Nielsen, V.G. Carbon monoxide inhibits the anticoagulant activity of Mojave rattlesnake venoms type A and B. J. Thromb. Thrombolysis 2019, 48, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, V.G.; Frank, N.; Turchioe, B.J. Anticoagulant activity of krait, coral snake, and cobra neurotoxic venoms with diverse proteomes are inhibited by carbon monoxide. Blood Coagul. Fibrinolysis 2019, 30, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, V.G. Carbon monoxide inhibits the anticoagulant activity of phospholipase A2 purified from Crotalus adamanteus venom. J. Thromb. Thrombolysis 2019, 47, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Gessner, G.; Sahoo, N.; Swain, S.M.; Hirth, G.; Schönherr, R.; Mede, R.; Westerhausen, M.; Brewitz, H.H.; Heimer, P.; Imhof, D.; et al. CO-independent modification of K+ channels by tricarbonyldichlororuthenium(II) dimer (CORM-2). Eur. J. Pharmacol. 2017, 815, 33–41. [Google Scholar] [CrossRef]
- Nielsen, V.G. The anticoagulant effect of Apis mellifera phospholipase A2 is inhibited by CORM-2 via a carbon monoxide-independent mechanism. J. Thromb. Thrombolysis 2020, 49, 100–107. [Google Scholar] [CrossRef]
- Nielsen, V.G.; Wagner, M.T.; Frank, N. Mechanisms Responsible for the Anticoagulant Properties of Neurotoxic Dendroaspis Venoms: A Viscoelastic Analysis. Int. J. Mol. Sci. 2020, 21, 2082. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Chen, X.; König, E.; Zhou, M.; Wang, L.; Chen, T.; Shaw, C. Comparative Profiling of Three Atheris Snake Venoms: A. squamigera, A. nitschei and A. chlorechis. Protein J. 2018, 37, 353–360. [Google Scholar] [CrossRef] [Green Version]
- Patra, A.; Kalita, B.; Chanda, A.; Mukherjee, A.K. Proteomics and antivenomics of Echis carinatus carinatus venom: Correlation with pharmacological properties and pathophysiology of envenomation. Sci. Rep. 2017, 7, 17119. [Google Scholar] [CrossRef] [Green Version]
- Yamada, D.; Morita, T. CA-1 method, a novel assay for quantification of normal prothrombin using a Ca2+ -dependent prothrombin activator, carinactivase-1. Thromb Res. 1999, 94, 221–226. [Google Scholar] [CrossRef]
- Chen, Y.L.; Tsai, I.H. Functional and sequence characterization of coagulation factor IX/factor X-binding protein from the venom of Echis carinatus leucogaster. Biochemistry 1996, 35, 5264–52671. [Google Scholar] [CrossRef]
- Viala, V.L.; Hildebrand, D.; Trusch, M.; Fucase, T.M.; Sciani, J.M.; Pimenta, D.C.; Arni, R.K.; Schlüter, H.; Betzel, C.; Mirtschin, P.; et al. Venomics of the Australian eastern brown snake (Pseudonaja textilis): Detection of new venom proteins and splicing variants. Toxicon 2015, 107 Pt B, 252–265. [Google Scholar] [CrossRef] [Green Version]
- Messori, L.; Orioli, P.; Vullo, D.; Alessio, E.; Iengo, E. A spectroscopic study of the reaction of NAMI, a novel ruthenium(III)anti-neoplastic complex, with bovine serum albumin. Eur. J. Biochem. 2000, 267, 1206–1213. [Google Scholar] [CrossRef] [PubMed]
- Messori, L.; Gonzales Vilchez, F.; Vilaplana, R.; Piccioli, F.; Alessio, E.; Keppler, B. Binding of Antitumor Ruthenium(III) Complexes to Plasma Proteins. Met. Based Drugs 2000, 7, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, V.G.; Cohen, B.M.; Cohen, E. Effects of coagulation factor deficiency on plasma coagulation kinetics determined via thrombelastography: Critical roles of fibrinogen and factors II, VII, X and XII. Acta Anaesthesiol. Scand. 2005, 49, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, V.G.; Kirklin, J.K.; Hoogendoorn, H.; Ellis, T.C.; Holman, W.L. Thrombelastographic method to quantify the contribution of factor XIII to coagulation kinetics. Blood Coagul. Fibrinolysis 2007, 18, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Motterlini, R.; Clark, J.E.; Foresti, R.; Sarathchandra, P.; Mann, B.E.; Green, C.J. Carbon monoxide-releasing molecules: Characterization of biochemical and vascular activities. Circ. Res. 2002, 90, E17–E24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meloun, B.; Morávek, L.; Kostka, V. Complete amino acid sequence of human serum albumin. FEBS Lett. 1975, 58, 134–137. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, V.G.; Pretorius, E. Iron and carbon monoxide enhance coagulation and attenuate fibrinolysis by different mechanisms. Blood Coagul. Fibrinolysis 2014, 25, 695–702. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, V.G.; Ward, T.; Ford, P.M. Effects of cupric chloride on coagulation in human plasma: Role of fibrinogen. J. Thromb. Thrombolysis 2018, 46, 359–364. [Google Scholar] [CrossRef]
- Nielsen, V.G. Lethal concentrations of mercury or lead do not affect coagulation kinetics in human plasma. J. Thromb. Thrombolysis 2019, 48, 697–698. [Google Scholar] [CrossRef]
- Nielsen, V.G.; Kirklin, J.K.; George, J.F. Carbon monoxide releasing molecule-2 increases the velocity of thrombus growth and strength in human plasma. Blood Coagul. Fibrinolysis 2009, 20, 377–380. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, V.G.; Kirklin, J.K.; George, J.F. Carbon monoxide-releasing molecule-2 decreases fibrinolysis in human plasma. Blood Coagul. Fibrinolysis 2009, 20, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Ellis, T.C.; Nielsen, V.G.; Marques, M.B.; Kirklin, J.K. Thrombelastographic measures of clot propagation: A comparison of alpha with the maximum rate of thrombus generation. Blood Coagul. Fibrinolysis 2007, 18, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, V.G. Crotalus atrox Venom Exposed to Carbon Monoxide Has Decreased Fibrinogenolytic Activity in Vivo in Rabbits. Basic Clin. Pharmacol. Toxicol. 2018, 122, 82–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Common Name | Proteome | CORM-2/iRM Inhibition |
|---|---|---|---|
| Atheris nitschei [16] | Great Lakes Bush Viper | SVSP, SVMP, PLA2 | Yes/No |
| Echis leucogaster [17,18,19] | White-Bellied Carpet Viper | SVSP, SVMP, PLA2 | Yes/No |
| Pseudonaja textilis [20] | Eastern Brown Snake | SVSP, SVMP, PLA2 | Yes/No |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nielsen, V.G. Ruthenium, Not Carbon Monoxide, Inhibits the Procoagulant Activity of Atheris, Echis, and Pseudonaja Venoms. Int. J. Mol. Sci. 2020, 21, 2970. https://doi.org/10.3390/ijms21082970
Nielsen VG. Ruthenium, Not Carbon Monoxide, Inhibits the Procoagulant Activity of Atheris, Echis, and Pseudonaja Venoms. International Journal of Molecular Sciences. 2020; 21(8):2970. https://doi.org/10.3390/ijms21082970
Chicago/Turabian StyleNielsen, Vance G. 2020. "Ruthenium, Not Carbon Monoxide, Inhibits the Procoagulant Activity of Atheris, Echis, and Pseudonaja Venoms" International Journal of Molecular Sciences 21, no. 8: 2970. https://doi.org/10.3390/ijms21082970
APA StyleNielsen, V. G. (2020). Ruthenium, Not Carbon Monoxide, Inhibits the Procoagulant Activity of Atheris, Echis, and Pseudonaja Venoms. International Journal of Molecular Sciences, 21(8), 2970. https://doi.org/10.3390/ijms21082970