Fear Learning Enhances Prefrontal Cortical Suppression of Auditory Thalamic Inputs to the Amygdala in Adults, but Not Adolescents

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
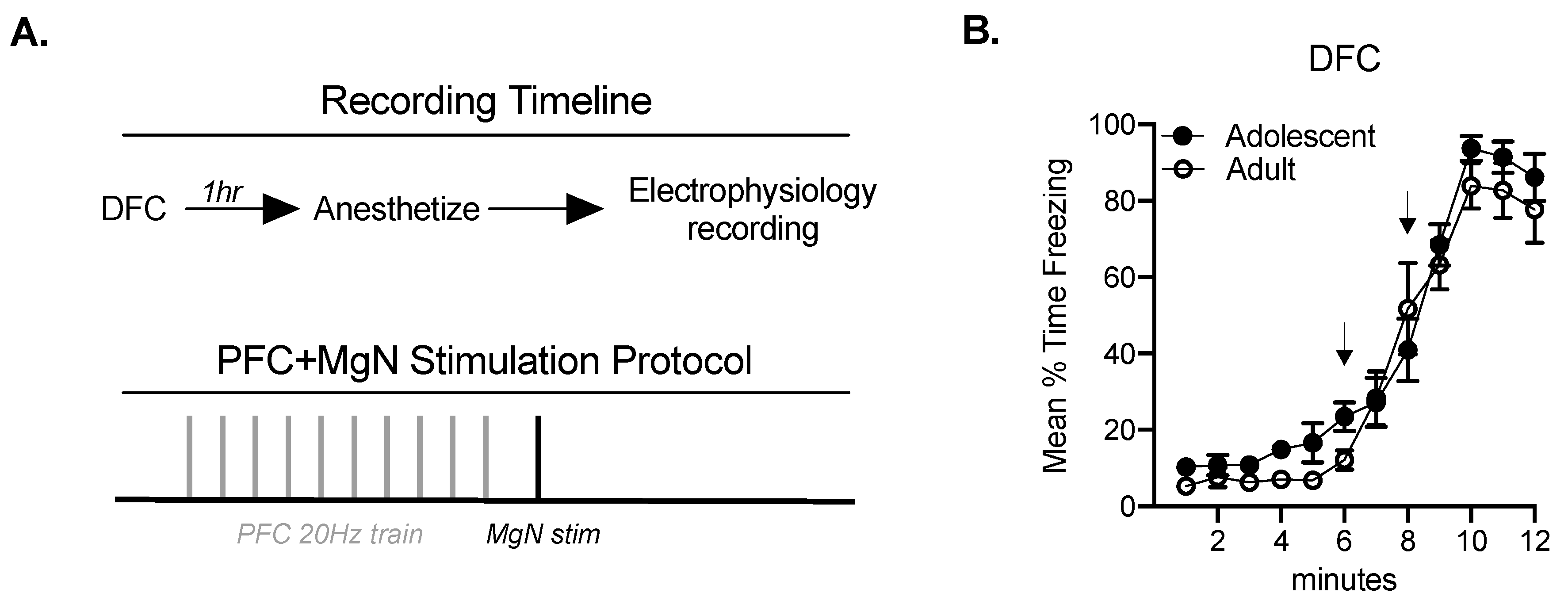
2.1. Fear Conditioning Facilitates BLA Responses to MgN Stimulation in Adults, but Not Adolescents
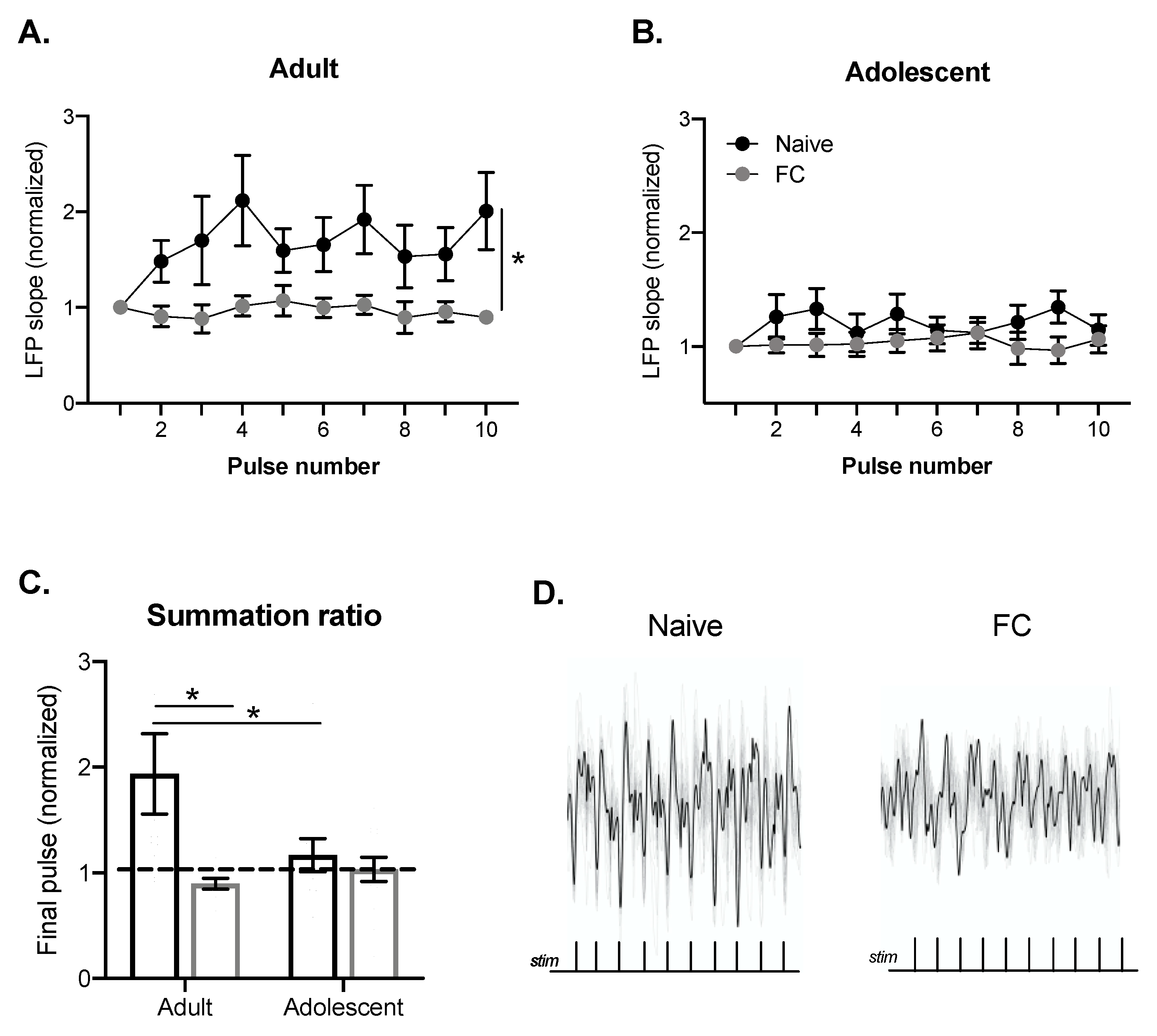
2.2. BLA Summation of LFPs Evoked by PFC Train Stimulation in Adults, but Not Adolescents
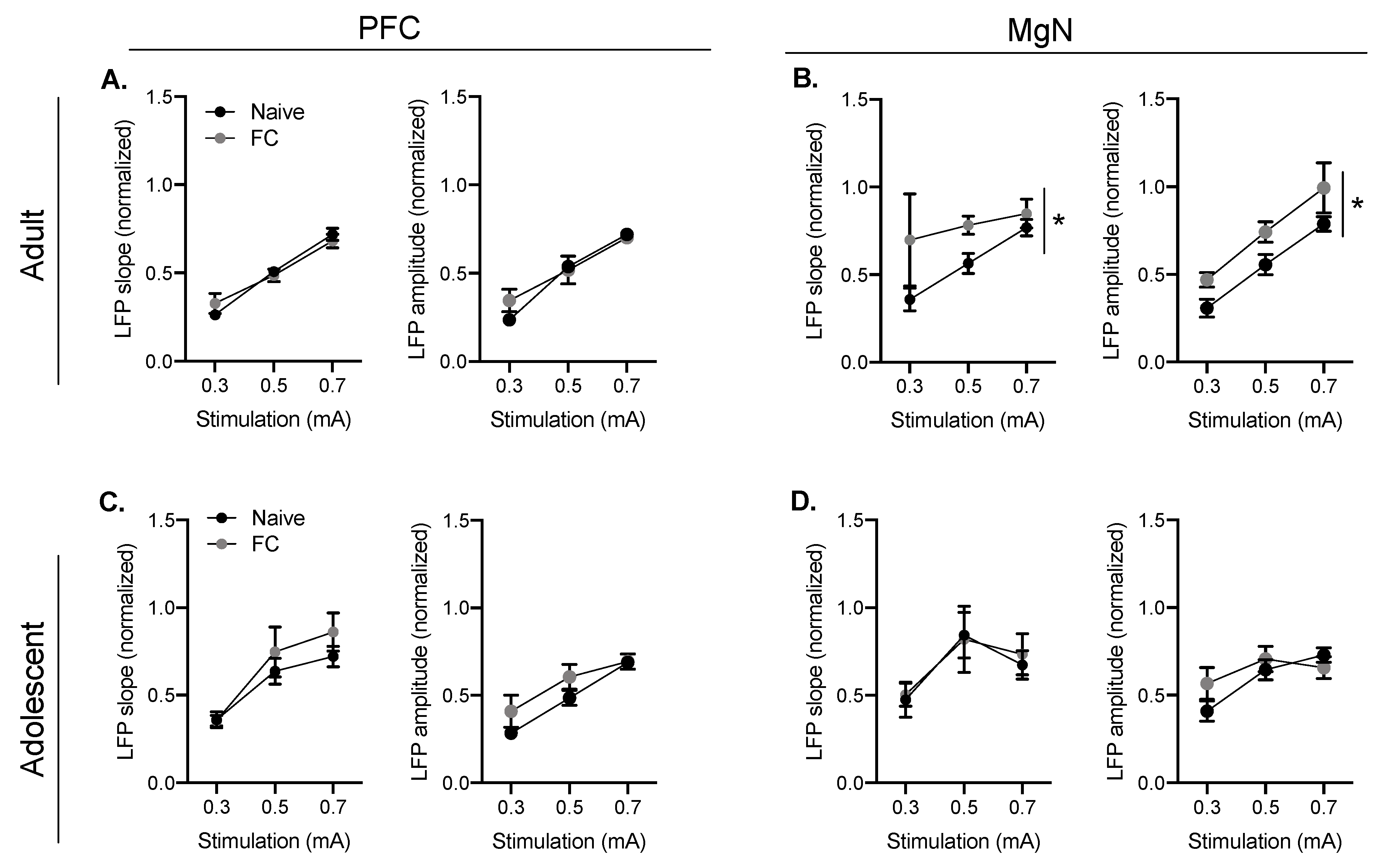
2.3. Developmental Differences in the Suppression of MgN-Evoked BLA LFPs Following Fear Conditioning
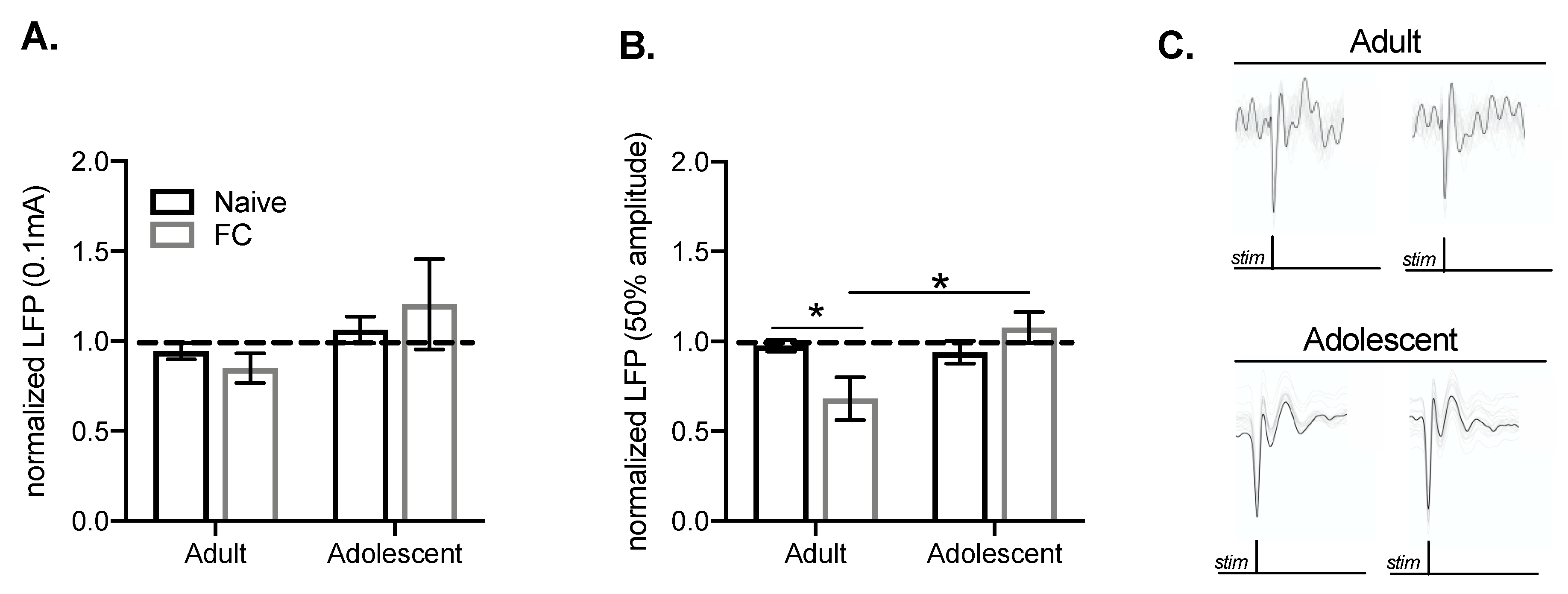
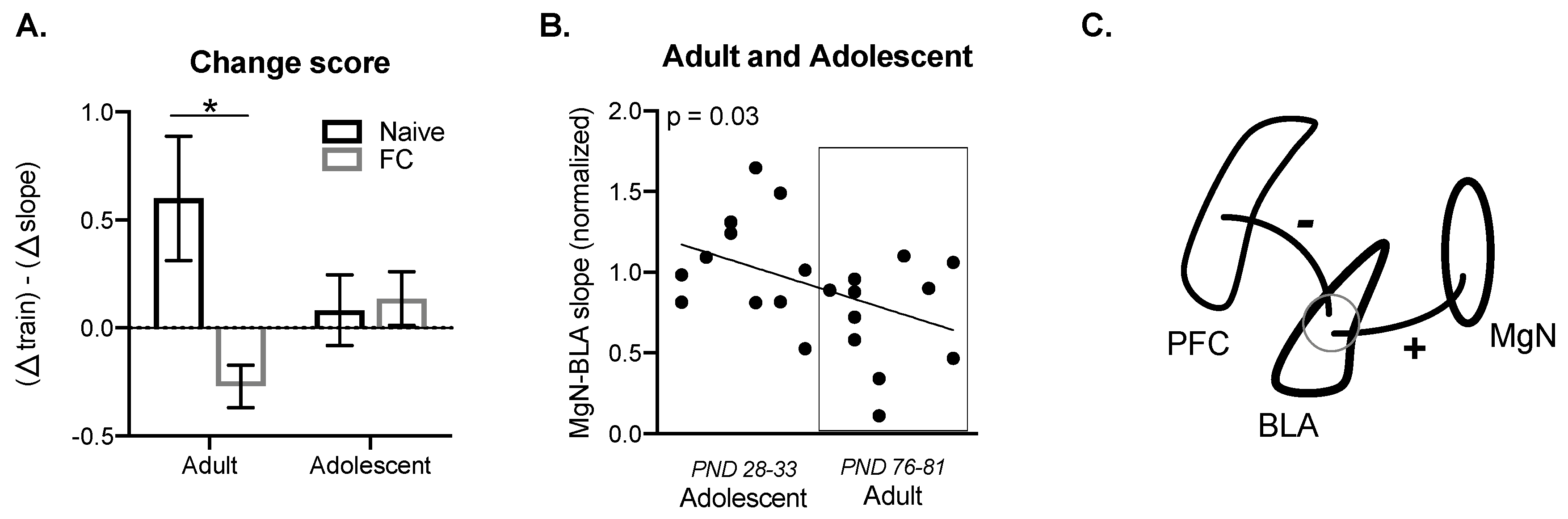
2.4. Prefrontal Cortical Summation is Associated with BLA Suppression of MgN Input
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Fear Conditioning
4.3. In Vivo Extracellular Surgery and Recording
4.4. Local Field Potential (LFP) Recordings
4.5. Histology
4.6. Data Analysis
4.7. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Meyer, H.C.; Lee, F.S. Translating Developmental Neuroscience to Understand Risk for Psychiatric Disorders. Am. J. Psychiatry 2019, 176, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Arp, J.M.; Ter Horst, J.P.; Loi, M.; Blaauwen, J.D.; Bangert, E.; Fernández, G.; Joëls, M.; Oitzl, M.S.; Krugers, H.J. Blocking glucocorticoid receptors at adolescent age prevents enhanced freezing between repeated cue-exposures after conditioned fear in adult mice raised under chronic early life stress. Neurobiol. Learn. Mem. 2016, 133, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Skelly, M.J.; Chappell, A.E.; Carter, E.; Weiner, J. Adolescent social isolation increases anxiety-like behavior and ethanol intake and impairs fear extinction in adulthood: Possible role of disrupted noradrenergic signaling. Neuropharmacology 2015, 97, 149–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Rosenkranz, J.A. Repeated restraint stress enhances cue-elicited conditioned freezing and impairs acquisition of extinction in an age-dependent manner. Behav. Brain Res. 2013, 248, 12–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fanselow, M.S. Conditioned and unconditional components of post-shock freezing. Pavlov. J. Boil. Sci. 1980, 15, 177–182. [Google Scholar]
- Pattwell, S.S.; Duhoux, S.; Hartley, C.A.; Johnson, D.C.; Jing, D.; Elliott, M.D.; Ruberry, E.J.; Powers, A.; Mehta, N.; Yang, R.R.; et al. Altered fear learning across development in both mouse and human. Proc. Natl. Acad. Sci. USA 2012, 109, 16318–16323. [Google Scholar] [CrossRef] [Green Version]
- Gale, G.D.; Anagnostaras, S.; Godsil, B.; Mitchell, S.; Nozawa, T.; Sage, J.R.; Wiltgen, B.; Fanselow, M. Role of the Basolateral Amygdala in the Storage of Fear Memories across the Adult Lifetime of Rats. J. Neurosci. 2004, 24, 3810–3815. [Google Scholar] [CrossRef]
- Goosens, K.A.; Maren, S. Contextual and auditory fear conditioning are mediated by the latera, basal, and central amygdaloid nuclei in rats. Learn. Mem. 2001, 8, 148–155. [Google Scholar] [CrossRef] [Green Version]
- Johansen, J.; Hamanaka, H.; Monfils, M.-H.; Behnia, R.; Deisseroth, K.; Blair, H.; LeDoux, J.E. Optical activation of lateral amygdala pyramidal cells instructs associative fear learning. Proc. Natl. Acad. Sci. USA 2010, 107, 12692–12697. [Google Scholar] [CrossRef] [Green Version]
- Kwapis, J.L.; Jarome, T.J.; Lonergan, M.E.; Helmstetter, F.J. Protein kinase Mzeta maintains fear memory in the amygdala but not in the hippocampus. Behav. Neurosci. 2009, 123, 844–850. [Google Scholar] [CrossRef] [Green Version]
- Kwapis, J.L.; Jarome, T.J.; Schiff, J.C.; Helmstetter, F.J. Memory consolidation in both trace and delay fear conditioning is disrupted by intra-amygdala infusion of the protein synthesis inhibitor anisomycin. Learn. Mem. 2011, 18, 728–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, S.M.; Schafe, G.E.; LeDoux, J.E. Intra-Amygdala Blockade of the NR2B Subunit of the NMDA Receptor Disrupts the Acquisition But Not the Expression of Fear Conditioning. J. Neurosci. 2001, 21, 6889–6896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arruda-Carvalho, M.; Wu, W.-C.; Cummings, K.A.; Clem, R.L. Optogenetic Examination of Prefrontal-Amygdala Synaptic Development. J. Neurosci. 2017, 37, 2976–2985. [Google Scholar] [CrossRef]
- Bosch, D.; Ehrlich, I. Postnatal maturation of GABAergic modulation of sensory inputs onto lateral amygdala principal neurons. J. Physiol. 2015, 593, 4387–4409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, K.D.; Gray, A.R.; Richardson, R. The development of perineuronal nets around parvalbumin gabaergic neurons in the medial prefrontal cortex and basolateral amygdala of rats. Behav. Neurosci. 2017, 131, 289–303. [Google Scholar] [CrossRef]
- Ehrlich, D.; Ryan, S.J.; Hazra, R.; Guo, J.-D.; Rainnie, D.G. Postnatal maturation of GABAergic transmission in the rat basolateral amygdala. J. Neurophysiol. 2013, 110, 926–941. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Rosenkranz, J.A. Effects of Repeated Stress on Age-Dependent GABAergic Regulation of the Lateral Nucleus of the Amygdala. Neuropsychopharmacology 2016, 41, 2309–2323. [Google Scholar] [CrossRef]
- Selleck, R.A.; Zhang, W.; Samberg, H.D.; Padival, M.; Rosenkranz, J.A. Limited prefrontal cortical regulation over the basolateral amygdala in adolescent rats. Sci. Rep. 2018, 8, 17171. [Google Scholar] [CrossRef]
- Moscarello, J.M.; LeDoux, J.E. Active avoidance learning requires prefrontal suppression of amygdala-mediated defensive reactions. J. Neurosci. 2013, 33, 3815–3823. [Google Scholar] [CrossRef]
- Sharpe, M.J.; Killcross, S. Modulation of attention and action in the medial prefrontal cortex of rats. Psychol. Rev. 2018, 125, 822–843. [Google Scholar] [CrossRef]
- Thompson, B.M.; Baratta, M.V.; Biedenkapp, J.; Rudy, J.W.; Watkins, L.R.; Maier, S.F. Activation of the infralimbic cortex in a fear context enhances extinction learning. Learn. Mem. 2010, 17, 591–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidal-Gonzalez, I.; Vidal-Gonzalez, B.; Rauch, S.L.; Quirk, G.J. Microstimulation reveals opposing influences of prelimbic and infralimbic cortex on the expression of conditioned fear. Learn. Mem. 2006, 13, 728–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haddon, J.E.; Killcross, S. Inactivation of the infralimbic prefrontal cortex in rats reduces the influence of inappropriate habitual responding in a response-conflict task. Neuroscience 2011, 199, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Zelikowsky, M.; Bissiere, S.; Hast, T.A.; Bennett, R.Z.; Abdipranoto, A.; Vissel, B.; Fanselow, M. Prefrontal microcircuit underlies contextual learning after hippocampal loss. Proc. Natl. Acad. Sci. USA 2013, 110, 9938–9943. [Google Scholar] [CrossRef] [Green Version]
- Baker, K.D.; Richardson, R. Forming competing fear learning and extinction memories in adolescence makes fear difficult to inhibit. Learn. Mem. 2015, 22, 537–543. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Li, S.; Richardson, R. Immunohistochemical Analyses of Long-Term Extinction of Conditioned Fear in Adolescent Rats. Cereb. Cortex 2010, 21, 530–538. [Google Scholar] [CrossRef] [Green Version]
- Koppensteiner, P.; Galvin, C.; Ninan, I. Lack of experience-dependent intrinsic plasticity in the adolescent infralimbic medial prefrontal cortex. Synapse 2019, 73, e22090. [Google Scholar] [CrossRef]
- Bloodgood, D.W.; Sugam, J.A.; Holmes, A.; Kash, T.L. Fear extinction requires infralimbic cortex projections to the basolateral amygdala. Transl. Psychiatry 2018, 8, 60. [Google Scholar] [CrossRef] [Green Version]
- Vouimba, R.-M.; Maroun, M. Learning-induced changes in mPFC-BLA connections after fear conditioning, extinction, and reistatement of fear. Neuropsychopharmacology 2011, 36, 2276–2285. [Google Scholar] [CrossRef]
- Ashwell, R.; Ito, R. Excitotoxic lesions of the infralimbic, but not prelimbic cortex facilitate reversal of appetitive discriminative context conditioning: The role of the infralimbic cortex in context generalization. Front. Behav. Neurosci. 2014, 8, 63. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.-H.; Deisseroth, K.; Bolshakov, V.Y. Synaptic encoding of fear extinction in mPFC-amygdala circuits. Neuron 2013, 80, 1491–1507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenkranz, J.A.; Grace, A. Cellular Mechanisms of Infralimbic and Prelimbic Prefrontal Cortical Inhibition and Dopaminergic Modulation of Basolateral Amygdala Neurons in Vivo. J. Neurosci. 2002, 22, 324–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenkranz, J.A.; Moore, H.; Grace, A. The Prefrontal Cortex Regulates Lateral Amygdala Neuronal Plasticity and Responses to Previously Conditioned Stimuli. J. Neurosci. 2003, 23, 11054–11064. [Google Scholar] [CrossRef] [PubMed]
- Doron, N.N.; LeDoux, J.E. Organization of projections to the lateral amygdala from auditory and visual areas of the thalamus in the rat. J. Compar.Neurol. 1999, 412, 383–409. [Google Scholar] [CrossRef]
- Parsons, R.G.; Riedner, B.A.; Gafford, G.M.; Helmstetter, F.J. The formation of auditory fear memory requires the synthesis of protein and mRNA in the auditory thalamus. Neurosci. 2006, 141, 1163–1170. [Google Scholar] [CrossRef] [Green Version]
- Clugnet, M.; LeDoux, J. Synaptic plasticity in fear conditioning circuits: Induction of LTP in the lateral nucleus of the amygdala by stimulation of the medial geniculate body. J. Neurosci. 1990, 10, 2818–2824. [Google Scholar] [CrossRef]
- Ferrara, N.C.; Cullen, P.K.; Pullins, S.P.; Rotondo, E.K.; Helmstetter, F.J. Input from the medial geniculate nucleus modulates amygdala encoding of fear memory discrimination. Learn. Mem. 2017, 24, 414–421. [Google Scholar] [CrossRef] [Green Version]
- Bin Kim, W.; Cho, J.-H. Encoding of Discriminative Fear Memory by Input-Specific LTP in the Amygdala. Neuron 2017, 95, 1129–1146. [Google Scholar]
- McKernan, M.G.; Shinnick-Gallagher, P. Fear conditioning induces a lasting potentiation of synaptic currents in vitro. Nature 1997, 390, 607–611. [Google Scholar] [CrossRef]
- Davis, P.; Zaki, Y.; Maguire, J.; Reijmers, L.G. Cellular and oscillatory substrates of fear extinction learning. Nat. Neurosci. 2017, 20, 1624–1633. [Google Scholar] [CrossRef]
- Likhtik, E.; Stujenske, J.M.; Topiwala, M.A.; Harris, A.Z.; Gordon, J.A. Prefrontal entrainment of amygdala activity signals safety in learned fear and innate anxiety. Nat. Neurosci. 2013, 17, 106–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narayanan, V.; Heiming, R.S.; Jansen, F.; Lesting, J.; Sachser, N.; Pape, H.-C.; Seidenbecher, T. Social Defeat: Impact on Fear Extinction and Amygdala-Prefrontal Cortical Theta Synchrony in 5-HTT Deficient Mice. PLoS ONE 2011, 6, e22600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stujenske, J.M.; Likhtik, E.; Topiwala, M.A.; Gordon, J.A. Fear and safety engage competing patterns of theta-gamma coupling in the basolateral amygdala. Neuron. 2014, 83, 919–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrara, N.C.; Mrackova, E.; Loh, M.K.; Padival, M.; Rosenkranz, J.A. Fear Learning Enhances Prefrontal Cortical Suppression of Auditory Thalamic Inputs to the Amygdala in Adults, but Not Adolescents. Int. J. Mol. Sci. 2020, 21, 3008. https://doi.org/10.3390/ijms21083008
Ferrara NC, Mrackova E, Loh MK, Padival M, Rosenkranz JA. Fear Learning Enhances Prefrontal Cortical Suppression of Auditory Thalamic Inputs to the Amygdala in Adults, but Not Adolescents. International Journal of Molecular Sciences. 2020; 21(8):3008. https://doi.org/10.3390/ijms21083008
Chicago/Turabian StyleFerrara, Nicole C., Eliska Mrackova, Maxine K. Loh, Mallika Padival, and J. Amiel Rosenkranz. 2020. "Fear Learning Enhances Prefrontal Cortical Suppression of Auditory Thalamic Inputs to the Amygdala in Adults, but Not Adolescents" International Journal of Molecular Sciences 21, no. 8: 3008. https://doi.org/10.3390/ijms21083008
APA StyleFerrara, N. C., Mrackova, E., Loh, M. K., Padival, M., & Rosenkranz, J. A. (2020). Fear Learning Enhances Prefrontal Cortical Suppression of Auditory Thalamic Inputs to the Amygdala in Adults, but Not Adolescents. International Journal of Molecular Sciences, 21(8), 3008. https://doi.org/10.3390/ijms21083008