Effects of Ferrocenyl 4-(Imino)-1,4-Dihydro-quinolines on Xenopus laevis Prophase I - Arrested Oocytes: Survival and Hormonal-Induced M-Phase Entry

 , ,
, ,
Abstract
:1. Introduction
2. Results
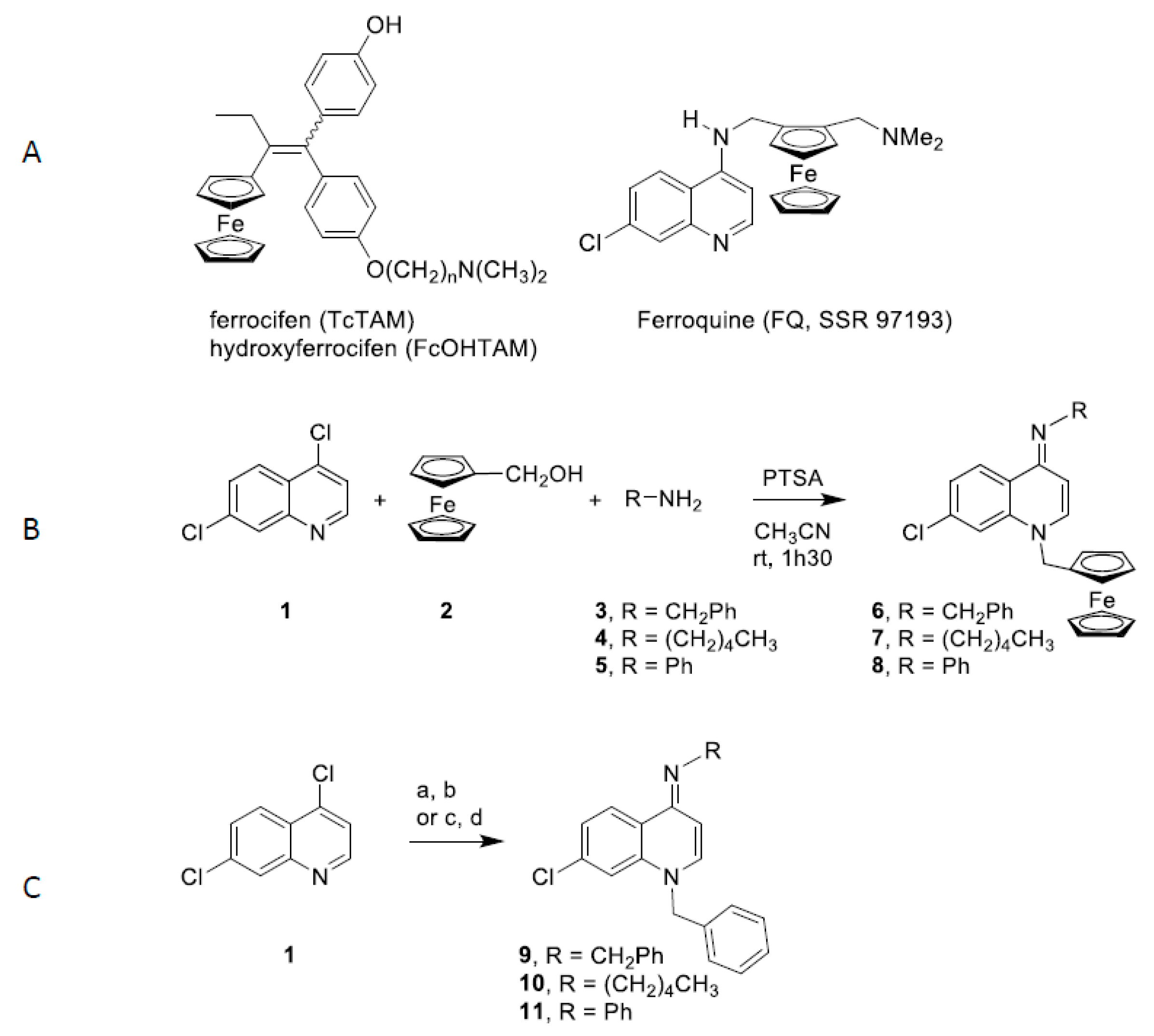
2.1. Synthesis of (imino)-1,4-Dihydroquinoline Derivatives
2.2. Effect of Ferrocenyl 4-(imino)-1,4-Dihydroquinolines on Oocyte Survival
2.3. Effects of 1,4-Dihydroquinolines on Mitotic Cell Division
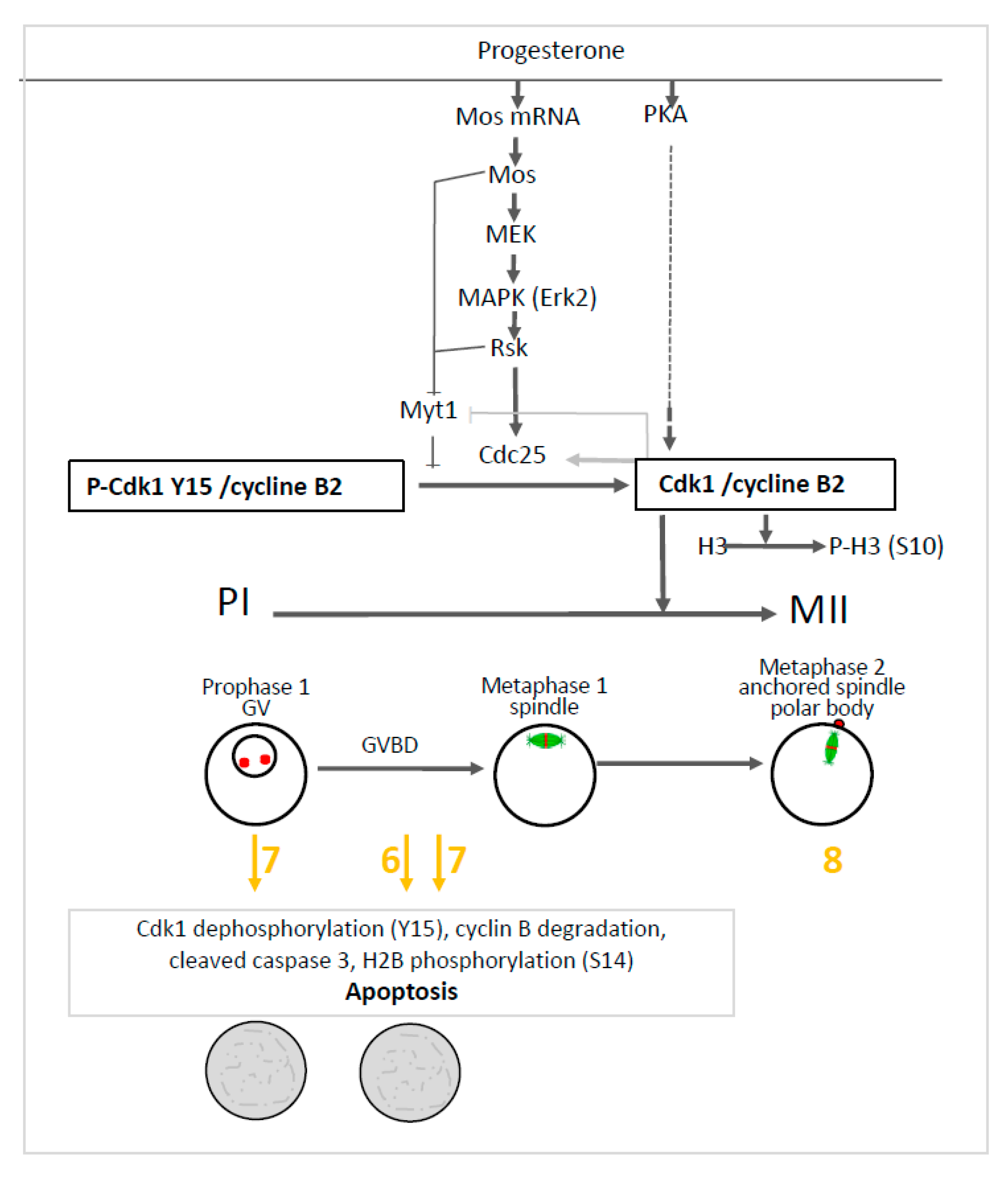
2.4. Effects of 1,4-Dihydroquinolines on Meiosis Resumption
2.5. Effects of 1,4-Dihydroquinolines on the Meiosis Spindle
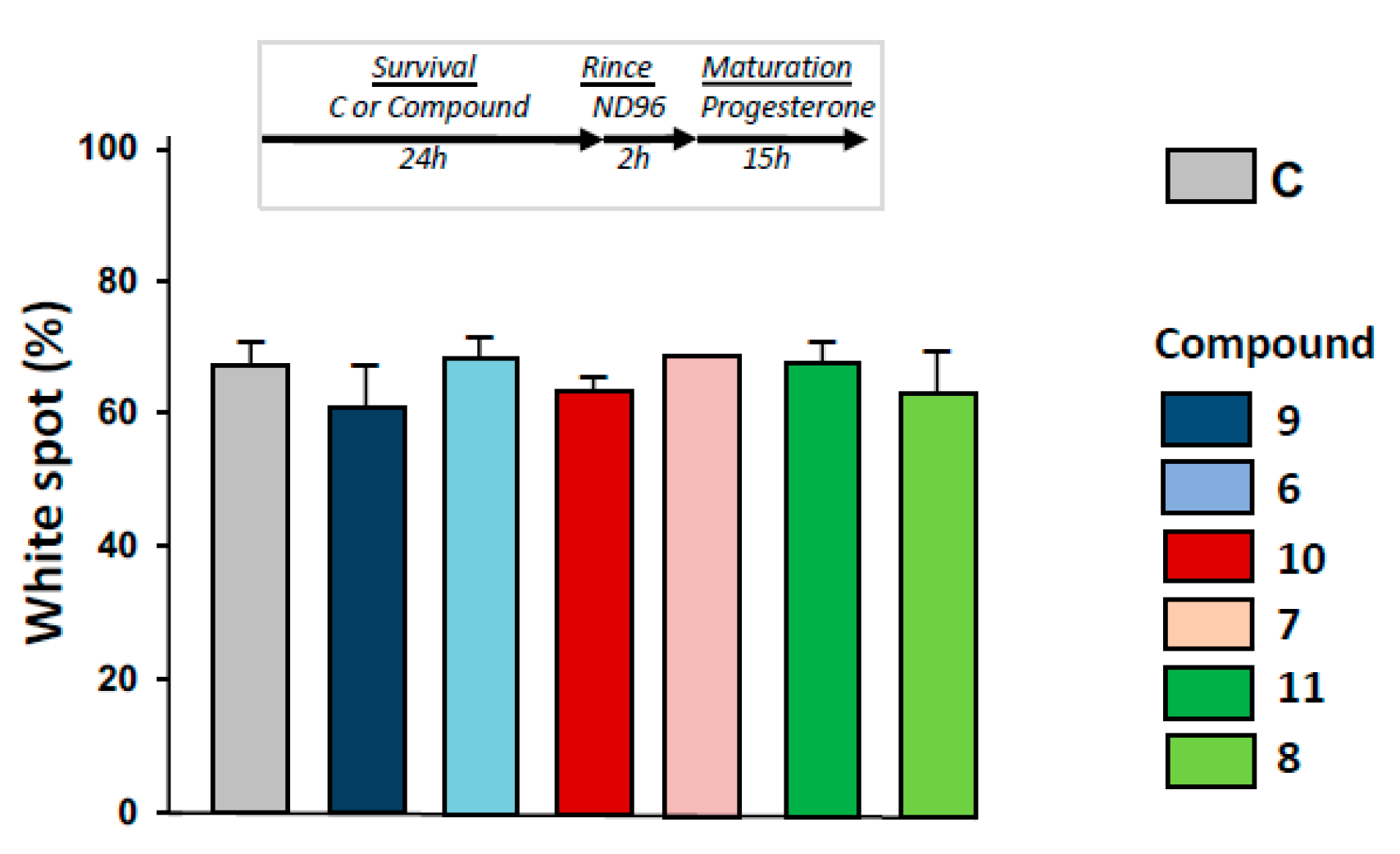
2.6. Maturation Success in 1,4-Dihydroquinolines Pre-Treated Oocytes
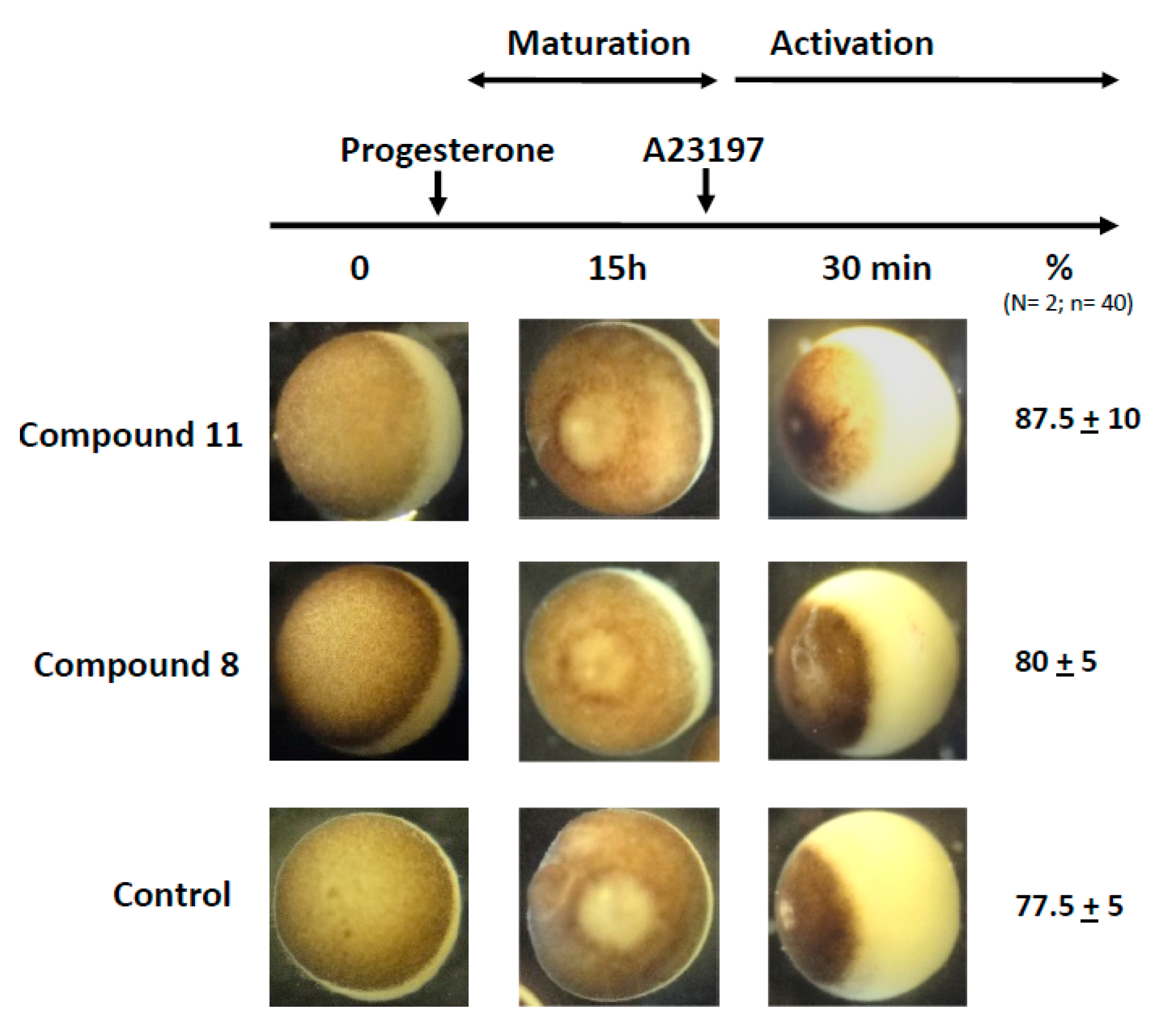
2.7. Effects of 1,4-Dihydroquinolines on Oocyte Parthenogenetic Activation
3. Discussion
4. Materials and Methods
4.1. General Information
4.2. Synthesis of 7-Chloro-1-Ferrocenylmethyl-4-(Benzylimino)-1,4-Dihydroquinoline (6)
4.3. Synthesis of 7-Chloro-1-Ferrocenylmethyl-4-(Pentylimino)-1,4-Dihydroquinoline (7)
4.4. Synthesis of 7-Chloro-1-Ferrocenylmethyl-4-(Phenylylimino)-1,4-Dihydroquinoline (8)
4.5. Synthesis of 1-Benzyl-7-Chloro-4(Benzylimino)-1,4-Dihydroquinoline (9)
4.6. Synthesis of 1-Benzyl-7-Chloro-4(Pentylimino)-1,4-Dihydroquinoline (10)
4.7. Synthesis of 1-Benzyl-7-Chloro-4(Phenylimino)-1,4-Dihydroquinoline (11)
4.8. Reagents and Substances for Biological Assays
4.9. Frog and Oocyte Handling
4.10. 1,4-Dihydroquinoline Exposure, Oocyte Death, GVBD Detection and Oocyte Parthenogenetic Activation
4.11. Electrophoresis and Western Blot Analysis
4.12. Histological Detection of Metaphase II Spindle
4.13. Cell Culture and Antiproliferative Activity Analysis
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| FETAX | Frog Embryo Teratogenesis Assay Xenopus |
| GVBD | Germinal Vesicle BreakDown |
| IC50 | half maximal inhibitory concentration |
| MPF | M-phase Promoting Factor |
| MS222 | Tricaïne Methane Sulfonate |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
References
- Brown, D.D. A tribute to the Xenopus laevis oocyte and egg. J. Biol. Chem. 2004, 279, 45291–45299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrell, J.E., Jr. Xenopus oocyte maturation: New lessons from a good egg. Bioessays 1999, 21, 833–842. [Google Scholar] [CrossRef]
- Schultz, T.W.; Dawson, D.A. Housing and husbandry of Xenopus for oocyte production. Lab. Anim. 2003, 32, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Collins, J.P.; Storfer, A. Global amphibian declines: Sorting the hypotheses. Divers. Distrib. 2003, 9, 89–98. [Google Scholar] [CrossRef]
- Hillman, S.S. Physiological correlates of differential dehydration tolerance in anurans. Copeia 1980, 1980, 125–129. [Google Scholar] [CrossRef]
- ATSM. Annual Book of ASTM Standards; ASTM: West Conshohocken, PA, USA, 1998. [Google Scholar]
- Bodart, J.F.; Gutierrez, D.V.; Nebreda, A.R.; Buckner, B.D.; Resau, J.R.; Duesbery, N.S. Characterization of MPF and MAPK activities during meiotic maturation of Xenopus tropicalis oocytes. Dev. Biol. 2002, 245, 348–361. [Google Scholar] [CrossRef]
- Heikkila, J.J.; Kaldis, A.; Morrow, G.; Tanguay, R.M. The use of the Xenopus oocyte as a model system to analyze the expression and function of eukaryotic heat shock proteins. Biotechnol. Adv. 2007, 25, 385–395. [Google Scholar] [CrossRef]
- Machaca, K. Ca2+ signaling differentiation during oocyte maturation. J. Cell. Physiol. 2007, 213, 331–340. [Google Scholar] [CrossRef]
- Cailliau, K.; Browaeys-Poly, E. A microinjectable biological system, the Xenopus oocyte, as an approach to understanding signal transduction protein function. Methods Mol. Biol. 2009, 518, 43–55. [Google Scholar]
- Weber, W. Ion currents of Xenopus laevis oocytes: State of the art. Biochim. Biophys. Acta 1999, 1421, 213–233. [Google Scholar] [CrossRef] [Green Version]
- Khokha, M.K. Xenopus white papers and resources: Folding functional genomics and genetics into the frog. Genesis 2012, 50, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Marin, M.; Slaby, S.; Marchand, G.; Demuynck, S.; Friscourt, N.; Gelaude, A.; Lemière, S.; Bodart, J.F. Xenopus laevis oocyte maturation is affected by metal chlorides. Toxicol. In Vitro 2015, 5, 1124–1131. [Google Scholar] [CrossRef] [PubMed]
- Nebreda, A.R.; Ferby, I. Regulation of the meiotic cell cycle in oocytes. Curr. Opin. Cell. Biol. 2000, 12, 666–675. [Google Scholar] [CrossRef]
- Ferrell, J.E., Jr.; Wu, M.; Gerhart, J.C.; Martin, G.S. Cell cycle tyrosine phosphorylation of p34cdc2 and a microtubule-associated protein kinase homolog in Xenopus oocytes and eggs. Mol. Cell. Biol. 1991, 11, 1965–1971. [Google Scholar] [CrossRef] [PubMed]
- Baert, F.; Bodart, J.F.; Bocquet-Muchembled, B.; Lescuyer-Rousseau, A.; Vilain, J.P. Xp42(Mpk1) activation is not required for germinal vesicle breakdown but for Raf complete phosphorylation in insulin-stimulated Xenopus oocytes. J. Biol. Chem. 2003, 278, 49714–49720. [Google Scholar] [CrossRef] [Green Version]
- Bodart, J.F.; Baert, F.Y.; Sellier, C.; Duesbery, N.S.; Flament, S.; Vilain, J.P. Differential roles of p39Mos-Xp42Mpk1 cascade proteins on Raf1 phosphorylation and spindle morphogenesis in Xenopus oocytes. Dev. Biol. 2005, 283, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Dupré, A.; Jessus, C.; Ozon, R.; Haccard, O. Mos is not required for the initiation of meiotic maturation in Xenopus oocytes. EMBO J. 2002, 21, 4026–4036. [Google Scholar] [CrossRef] [Green Version]
- Surrey, A.R.; Lesher, G.Y.; Mayer, J.R.; Webb, W.G. Hypotensive agents. 11. The preparation of quaternary salts of some 4-dialkylaminoalkylaminoquinolines. J. Am. Chem. Soc. 1959, 81, 2894–2897. [Google Scholar] [CrossRef]
- Campos Rosa, J.; Galanakis, D.; Piergentili, A.; Bhandari, K.; Ganellin, C.R.; Dunn, P.M.; Jenkinson, D.H. Synthesis, molecular modeling, and pharmacological testing of bis-quinolinium cyclophanes: Potent, non-peptidic blockers of the apamin-sensitive Ca(2+)-activated K(+) channel. J. Med. Chem. 2000, 43, 420–431. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, T.; da Cruz, F.P.; Lafuente-Monasterio, M.J.; Gonçalves, D.; Ressurreição, A.S.; Sitoe, A.R.; Bronze, M.R.; Gu, T.J.; Schneider, G.; Mota, M.M.; et al. Quinolin-4(1H)-imines are potent antiplasmodial drugs targeting the liver stage of malaria. J. Med. Chem. 2013, 56, 4811–4815. [Google Scholar] [CrossRef]
- Patra, M.; Gasser, G. Organometallic compounds an opportunity for chemical biology. Chem. Bio. Chem. 2012, 13, 1232–1252. [Google Scholar] [CrossRef] [PubMed]
- Hartinger, C.G.; Dyson, P.J. Bioorganometallic chemistry—From teaching paradigms to medicinal applications. Chem. Soc. Rev. 2009, 38, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Gasser, G.; Metzler-Nolte, N. The potential of organometallic complexes in medicinal chemistry. Curr. Opin. Chem. Biol. 2012, 16, 84–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasser, G.; Ott, I.; Metzler-Nolte, N. Organometallic anticancer compounds. J. Med. Chem. 2011, 54, 3–25. [Google Scholar] [CrossRef]
- Top, S.; Tang, J.; Vessières, A.; Carrez, D.; Provot, C.; Jaouen, G. Ferrocenyl hydroxytamoxifen: A prototype for a new range of oestradiol receptor site-directed cytotoxics. Chem. Commun. 1996, 8, 955–956. [Google Scholar] [CrossRef]
- Vessières, A.; Top, S.; Beck, W.; Hillard, E.; Jaouen, G. Metal complex SERMs (selective oestrogen receptor modulators). The influence of different metal units on breast cancer cells antiproliferative effects. Dalton Trans. 2006, 529–541. [Google Scholar] [CrossRef]
- Jaouen, G.; Top, S.; Vessières, A.; Leclercq, G.; McGlinchey, M. The first organometallic selective estrogen receptor modulators (SERMs) and their relevance to breast cancer. Curr. Med. Chem. 2004, 11, 2505–2517. [Google Scholar] [CrossRef]
- Kondratskyi, A.; Kondratska, K.; Vanden Abeele, F.; Gordienko, D.; Dubois, C.; Toillon, R.A.; Slomianny, C.; Lemière, S.; Delcourt, P.; Dewailly, E.; et al. Ferroquine, the next generation antimalarial drug, has antitumor activity. Sci. Rep. 2017, 7, 15896–15911. [Google Scholar] [CrossRef] [Green Version]
- Biot, C.; Nosten, F.; Fraisse, L.; Ter-Minassian, D.; Khalife, J.; Dive, D. The antimalarial ferroquine: From bench to clinic. Parasite 2011, 18, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Wambang, N.; Schifano-Faux, N.; Martoriati, A.; Henry, N.; Baldeyrou, B.; Bal-Mahieu, C.; Bousquet, T.; Pellegrini, S.; Meignan, S.; Cailliau, K.; et al. Synthesis, Structure, and Antiproliferative Activity of Ruthenium(II) Arene Complexes of Indenoisoquinoline Derivatives. Organometallics 2016, 35, 2868–2872. [Google Scholar] [CrossRef]
- Wambang, N.; Schifano-Faux, N.; Aillerie, A.; Baldeyrou, B.; Jacquet, C.; Bal-Mahieu, C.; Bousquet, T.; Pellegrini, S.; Ndifon Téké, P.; Meignan, S.; et al. Synthesis and biological activity of ferrocenyl indeno[1,2-c]isoquinolines as topoisomerase II inhibitors. Bioorg. Med. Chem. 2016, 24, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Mwande Maguene, G.; Lekana-Douki, J.B.; Mouray, E.; Bousquet, T.; Grellier, P.; Pellegrini, S.; Toure Ndouo, F.S.; Lebibi, J.; Pélinski, L. Synthesis and in vitro antiplasmodial activity of ferrocenyl aminoquinoline derivatives. Eur. J. Med. Chem. 2015, 90, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, S.; Grad, J.-N.; Bousquet, T.; Pélinski, L. A novel multicomponent reaction: Easy access to ferrocenyl (alkylimino)-1,4-dihydroquinolines. Tetrahedron Lett. 2011, 52, 1742–1744. [Google Scholar] [CrossRef]
- Azuma, T. Distribution of Anticancer Drugs in River Waters and Sediments of the Yodo River Basin, Japan. Appl. Sci. 2018, 8, 2043–2062. [Google Scholar] [CrossRef] [Green Version]
- Heath, E.; Filipic, M.; Kosjek, T.; Isidori, M. Fate and effects of the residues of anticancer drugs in the environment. Environ. Sci. Pollut. Res. 2016, 23, 14687–14691. [Google Scholar] [CrossRef]
- Bazin, M.; Kuhn, C. Use of Quinolinium Salts in Parallel Synthesis for the Preparation of 4-Amino-2-alkyl-1, 2, 3, 4-tetrahydroquinoline. J. Comb. Chem. 2005, 7, 302–308. [Google Scholar] [CrossRef]
- Weaver, B.A. How Taxol/paclitaxel kills cancer cells. Mol. Biol. Cell. 2014, 25, 2677–2681. [Google Scholar] [CrossRef]
- Nassour, C.; Barton, S.J.; Nabhani-Gebara, S.; Saab, Y.; Barker, J. Occutrence of anticancer drugs in the aquatic environment: A systematic review. Environ. Sci. Pollut. Res. Int. 2020, 27, 1339–1347. [Google Scholar] [CrossRef] [Green Version]
- Pereira da Costa Araujo, A.; Mesak, C.; Montalvao, M.F.; Freitas, I.N.; Chagas, T.Q.; Malafaia, G. Anti-cancer drugs in aquatic environment can cause cancer; Insight about mutagenicity in tapoles. Sci. Total. Environ. 2019, 650, 2284–2293. [Google Scholar] [CrossRef]
- Ben Messaoud, N.; Katzarova, I.; López, J.M. Basic Properties of the p38 Signaling Pathway in Response to Hyperosmotic Shock. PLoS ONE 2015, 10, e0135249. [Google Scholar] [CrossRef] [Green Version]
- Ben Messaoud, N.; Yue, J.; Valent, D.; Katzarova, I.; López, J.M. Osmostress-induced apoptosis in Xenopus oocytes: Role of stress protein kinases, calpains and Smac/DIABLO. PLoS ONE 2015, 10, e0124482. [Google Scholar] [CrossRef] [Green Version]
- Börjesson, S.I.; Englund, U.H.; Asif, M.H.; Willander, M.; Elinder, F. Intracellular K+ Concentration Decrease Is Not Obligatory for Apoptosis. J. Biol. Chem. 2011, 286, 39823–39828. [Google Scholar] [CrossRef] [Green Version]
- Du Pasquier, D.; Dupré, A.; Jessus, C. Unfertilized Xenopus eggs die by Bad-dependent apoptosis under the control of Cdk1 and JNK. PLoS ONE 2011, 6, e23672. [Google Scholar] [CrossRef] [Green Version]
- Tokmakov, A.A.; Iguchi, S.; Iwasaki, T.; Fukami, Y. Unfertilized frog eggs die by apoptosis following meiotic exit. BMC Cell Biol. 2011, 12, 56. [Google Scholar] [CrossRef] [Green Version]
- Englund, U.H.; Gertow, J.; Kågedal, K.; Elinder, F.A. Voltage dependent non-inactivating Na+ channel activated during apoptosis in Xenopus oocytes. PLoS ONE 2014, 28, e88381. [Google Scholar] [CrossRef]
- Cheung, W.; Ajiro, K.; Samejima, K.; Kloc, M.; Cheung, P.; Mizzen, C.A.; Beese, R.A.; Etkin, L.D.; Chernoff, J.; Earnshaw, W.C.; et al. Apoptotic phosphorylation of histone H2B is mediated by mammalian sterile twenty kinase. Cell 2003, 16, 507–517. [Google Scholar] [CrossRef] [Green Version]
- Hochegger, H.; Klotzbücher, A.; Kirk, J.; Howell, M.; Le Guellec, K.; Fletcher, K.; Duncan, T.; Sohail, M.; Hunt, T. New B-type cyclin synthesis is required between meiosis I and II during Xenopus oocyte maturation. Development 2001, 128, 3795–3807. [Google Scholar] [PubMed]
- Nishiyama, A.; Tachibana, K.; Igarashi, Y.; Yasuda, H.; Tanahashi, N.; Tanaka, K.; Ohsumi, K.; Kishimoto, T.A. Nonproteolytic function of the proteasome is required for the dissociation of Cdc2 and cyclin B at the end of M phase. Genes Dev. 2000, 14, 2344–2357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, H.; Minshull, J.; Ford, C.; Golsteyn, R.; Poon, R.; Hunt, T. On the synthesis anD destruction of A- and B-type cyclins during oogenesis and meiotic maturation in Xenopus laevis. J. Cell Biol. 1991, 114, 755–765. [Google Scholar] [CrossRef]
- Sentein, P.; Ates, Y. Cytological characteristics and classification of spindle inhibitors according to their effects on segmentation mitoses. Cellule 1978, 72, 265–289. [Google Scholar]
- Schiemann, K.; Finsinger, D.; Zenke, F.; Amendt, C.; Knöchel, T.; Bruge, D.; Buchstaller, H.P.; Emde, U.; Stähle, W.; Anzali, S. The discovery and optimization of hexahydro-2H-pyrano[3, c]quinolines (HHPQs) as potent and selective inhibitors of the mitotic kinesin-5. Bioorg. Med. Chem. Lett. 2010, 20, 1491–1495. [Google Scholar] [CrossRef] [PubMed]
- Gard, D.L. Microtubule organization during maturation of Xenopus oocytes: Assembly and rotation of the meiotic spindles. Dev. Biol. 1992, 151, 516–530. [Google Scholar] [CrossRef]
- Flament, S.; Browaeys, E.; Rodeau, J.L.; Bertout, M.; Vilain, J.P. Xenopus oocyte maturation: Cytoplasm alkalization is involved in germinal vesicle migration. Int. J. Dev. Biol. 1996, 40, 471–476. [Google Scholar] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | IC50 Hela (μM) | IC50 MDA-MB-231 (Μm) |
|---|---|---|
| 9 | 18.28 ± 7.95 | 26.03 ± 2.94 |
| 6 | 6.70 ± 1.11 | 8.06 ± 1.00 |
| 10 | 14.25 ± 7.28 | 11.04 ± 0.29 |
| 7 | 4.91 ± 0.04 | 3.39 ± 1.52 |
| 11 | 42.09 ± 2.87 | 13.61 ± 0.47 |
| 8 | 6.21 ± 1.73 | 7.39 ± 0.19 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marchand, G.; Wambang, N.; Pellegrini, S.; Molinaro, C.; Martoriati, A.; Bousquet, T.; Markey, A.; Lescuyer-Rousseau, A.; Bodart, J.-F.; Cailliau, K.; et al. Effects of Ferrocenyl 4-(Imino)-1,4-Dihydro-quinolines on Xenopus laevis Prophase I - Arrested Oocytes: Survival and Hormonal-Induced M-Phase Entry. Int. J. Mol. Sci. 2020, 21, 3049. https://doi.org/10.3390/ijms21093049
Marchand G, Wambang N, Pellegrini S, Molinaro C, Martoriati A, Bousquet T, Markey A, Lescuyer-Rousseau A, Bodart J-F, Cailliau K, et al. Effects of Ferrocenyl 4-(Imino)-1,4-Dihydro-quinolines on Xenopus laevis Prophase I - Arrested Oocytes: Survival and Hormonal-Induced M-Phase Entry. International Journal of Molecular Sciences. 2020; 21(9):3049. https://doi.org/10.3390/ijms21093049
Chicago/Turabian StyleMarchand, Guillaume, Nathalie Wambang, Sylvain Pellegrini, Caroline Molinaro, Alain Martoriati, Till Bousquet, Angel Markey, Arlette Lescuyer-Rousseau, Jean-François Bodart, Katia Cailliau, and et al. 2020. "Effects of Ferrocenyl 4-(Imino)-1,4-Dihydro-quinolines on Xenopus laevis Prophase I - Arrested Oocytes: Survival and Hormonal-Induced M-Phase Entry" International Journal of Molecular Sciences 21, no. 9: 3049. https://doi.org/10.3390/ijms21093049
APA StyleMarchand, G., Wambang, N., Pellegrini, S., Molinaro, C., Martoriati, A., Bousquet, T., Markey, A., Lescuyer-Rousseau, A., Bodart, J. -F., Cailliau, K., Pelinski, L., & Marin, M. (2020). Effects of Ferrocenyl 4-(Imino)-1,4-Dihydro-quinolines on Xenopus laevis Prophase I - Arrested Oocytes: Survival and Hormonal-Induced M-Phase Entry. International Journal of Molecular Sciences, 21(9), 3049. https://doi.org/10.3390/ijms21093049