The Serine Protease Autotransporters TagB, TagC, and Sha from Extraintestinal Pathogenic Escherichia coli Are Internalized by Human Bladder Epithelial Cells and Cause Actin Cytoskeletal Disruption

 , ,
, ,  ,
,  and
and 
Abstract
:
1. Introduction
2. Results
2.1. Processing and Secretion of TagB, TagC, and Sha Is Independent of the Serine Protease Motif
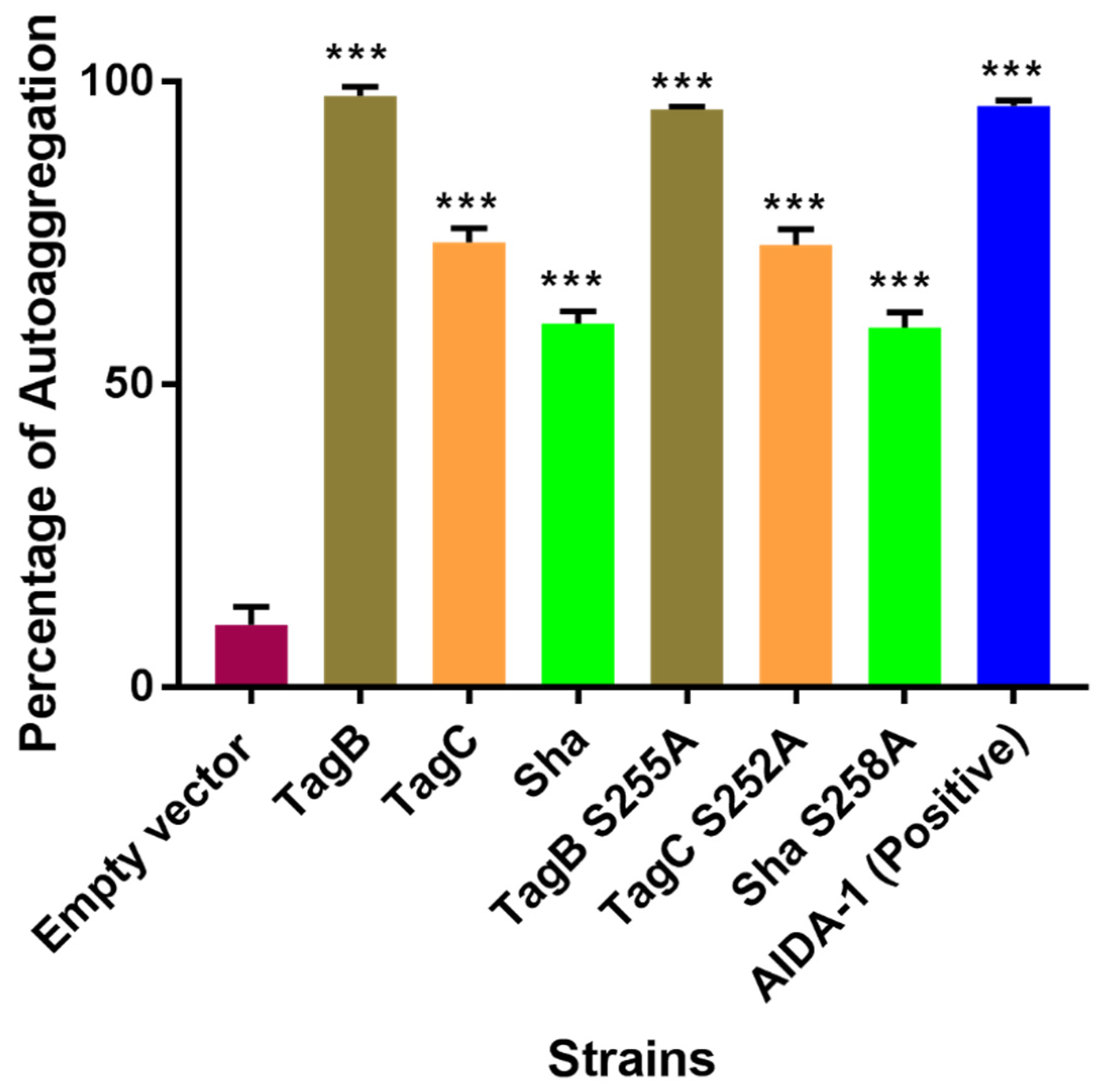
2.2. The Serine Catalytic Motif of SPATEs Is Not Required for Autoaggregation or Hemagglutination Activity
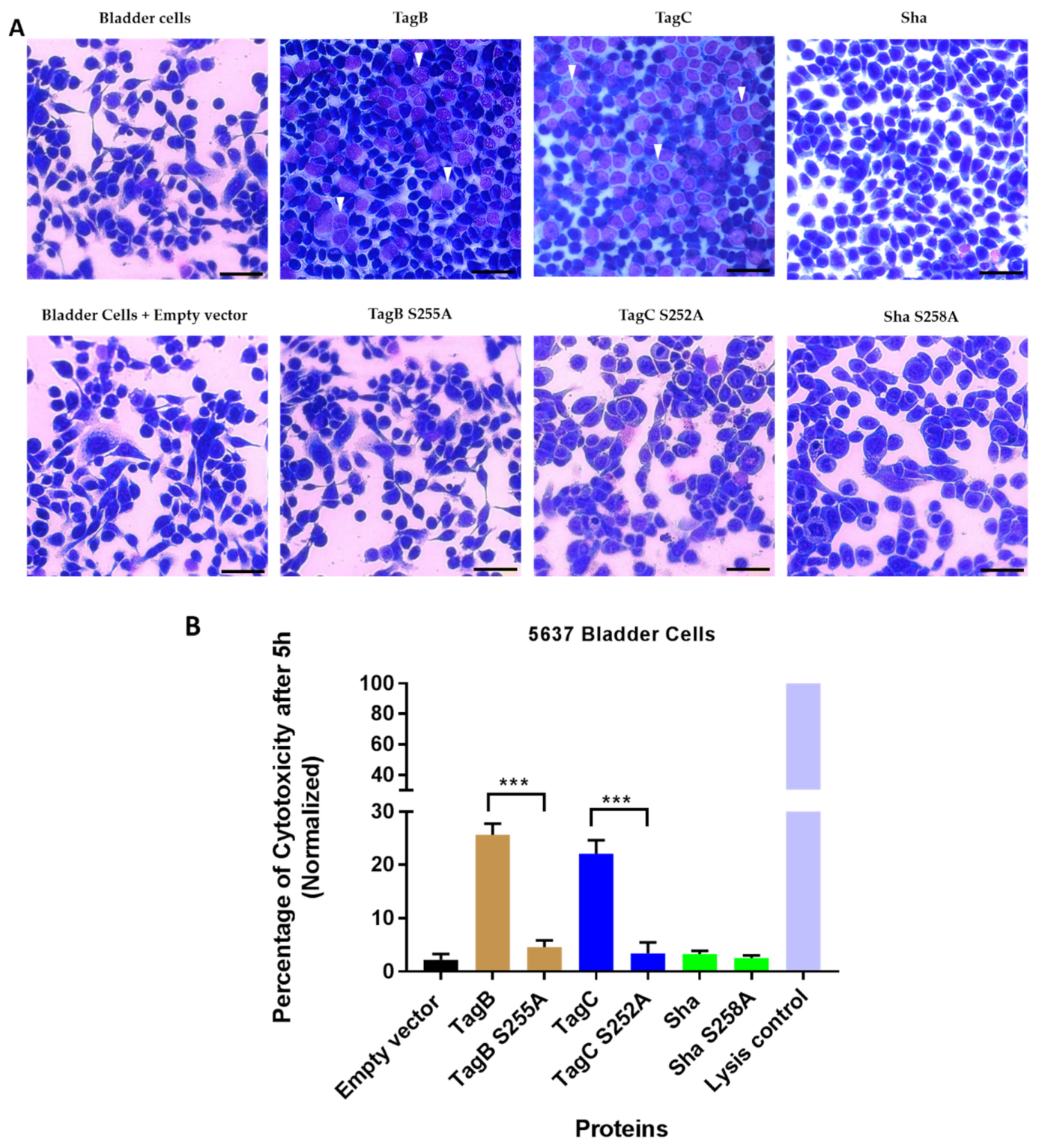
2.3. Cytopathic Effect of TagB and TagC Requires the Serine Protease Motif
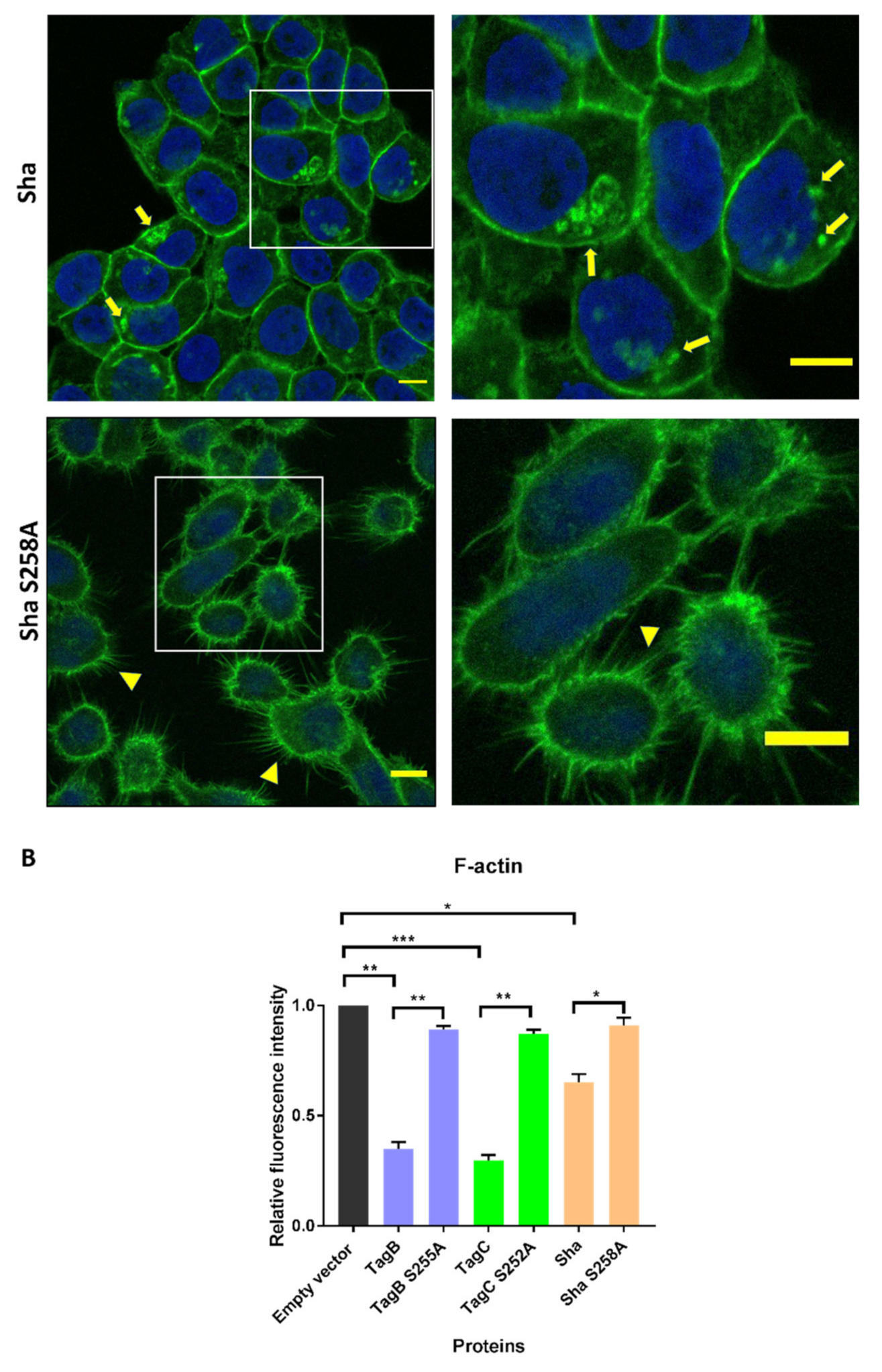
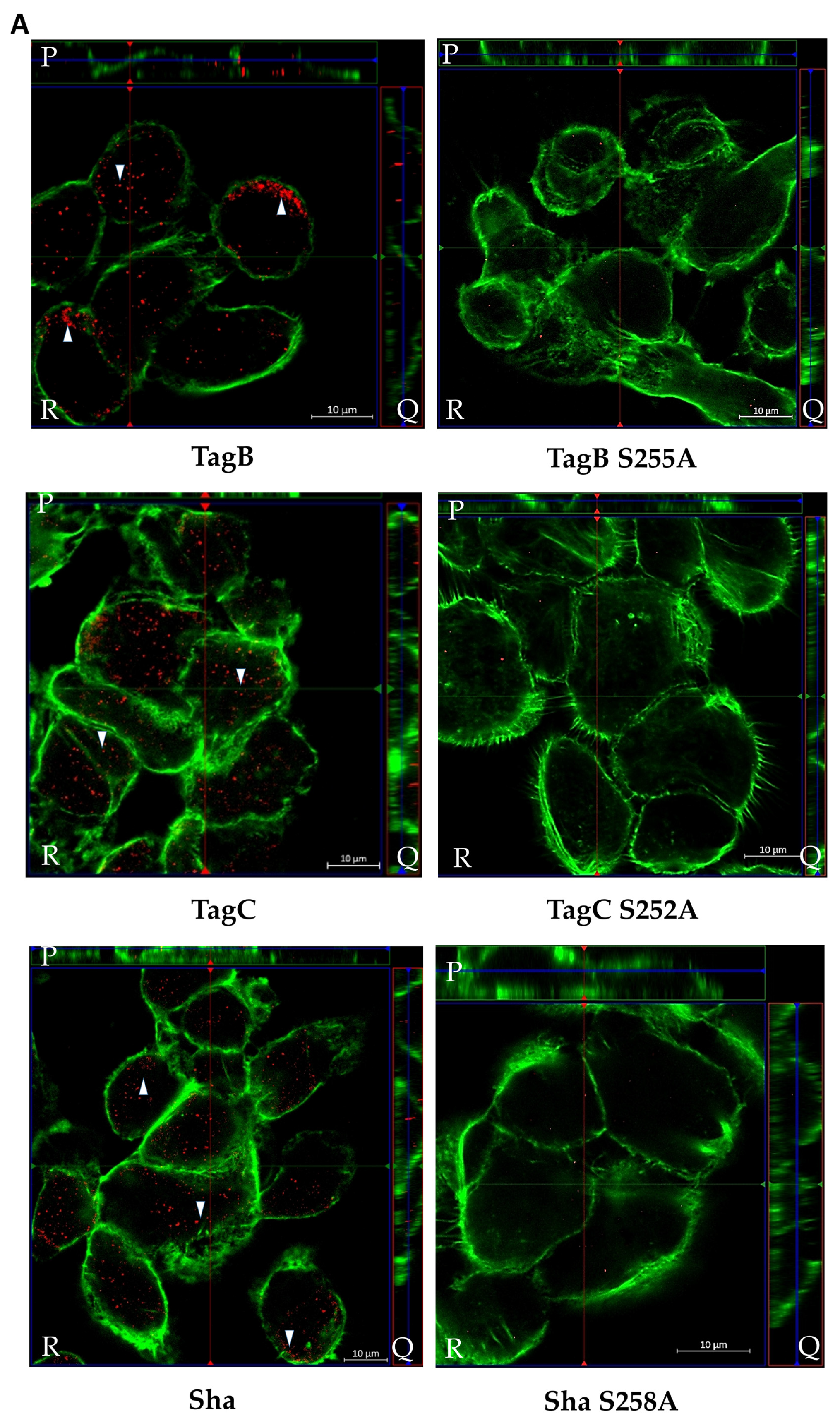
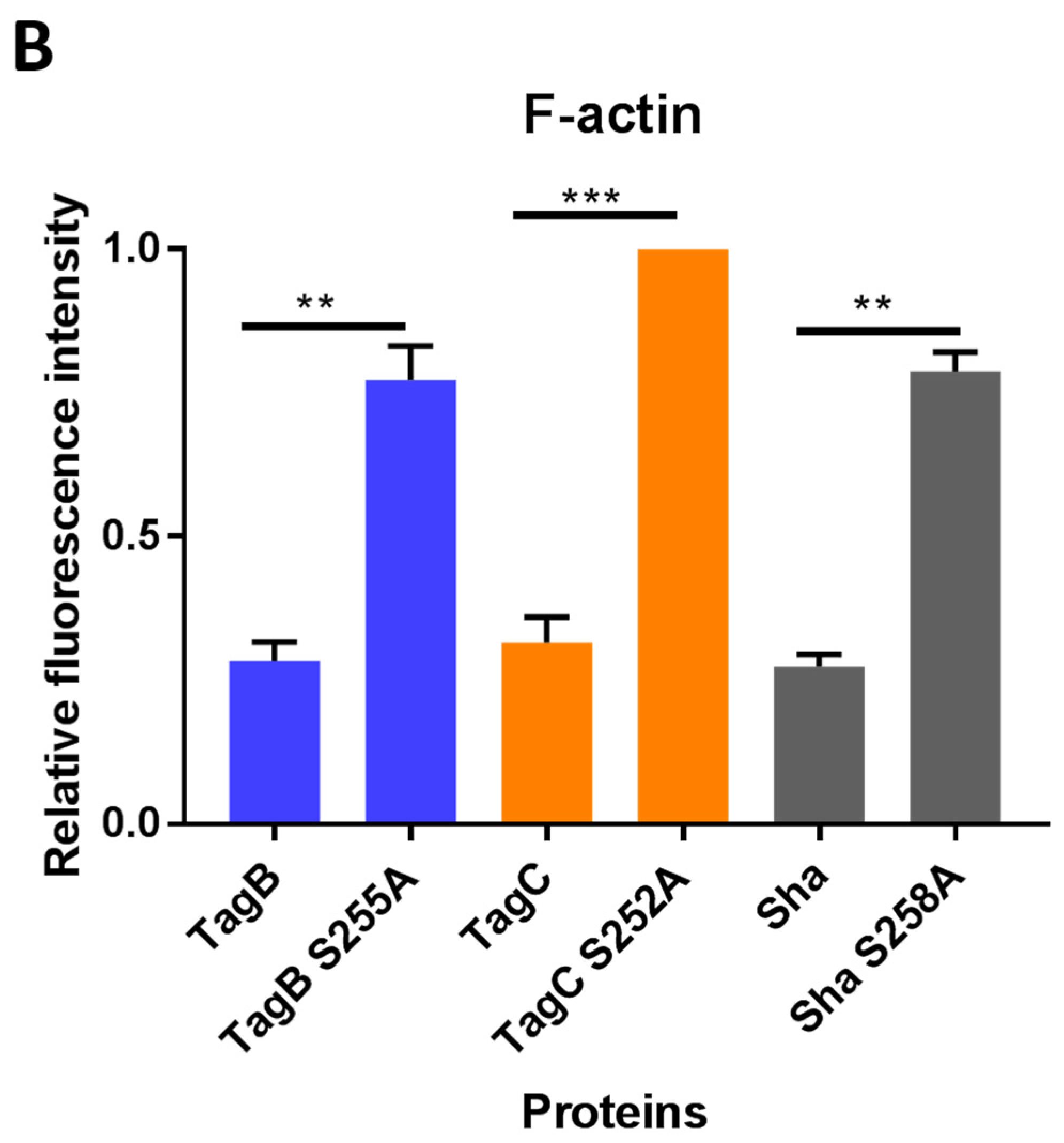
2.4. Exposure to TagB, TagC, or Sha Alters Actin Distribution in Bladder Epithelial Cells
2.5. SPATE Entry into Bladder Epithelial Cells Is Dependent on the Serine Protease Active Site
2.6. Sha Exhibits Serine Protease-Dependent Mucinase Activity
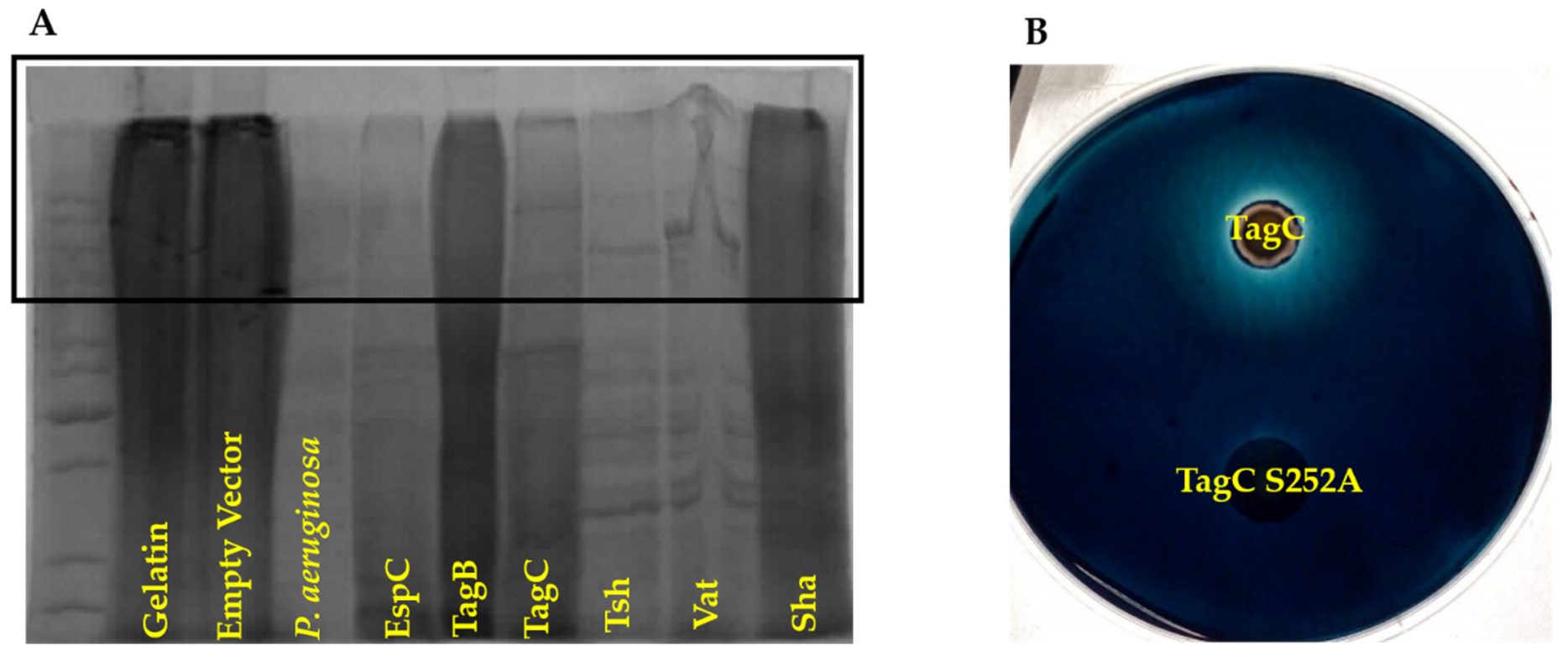
2.7. TagC Exhibits Serine Protease-Dependent Gelatinase Activity
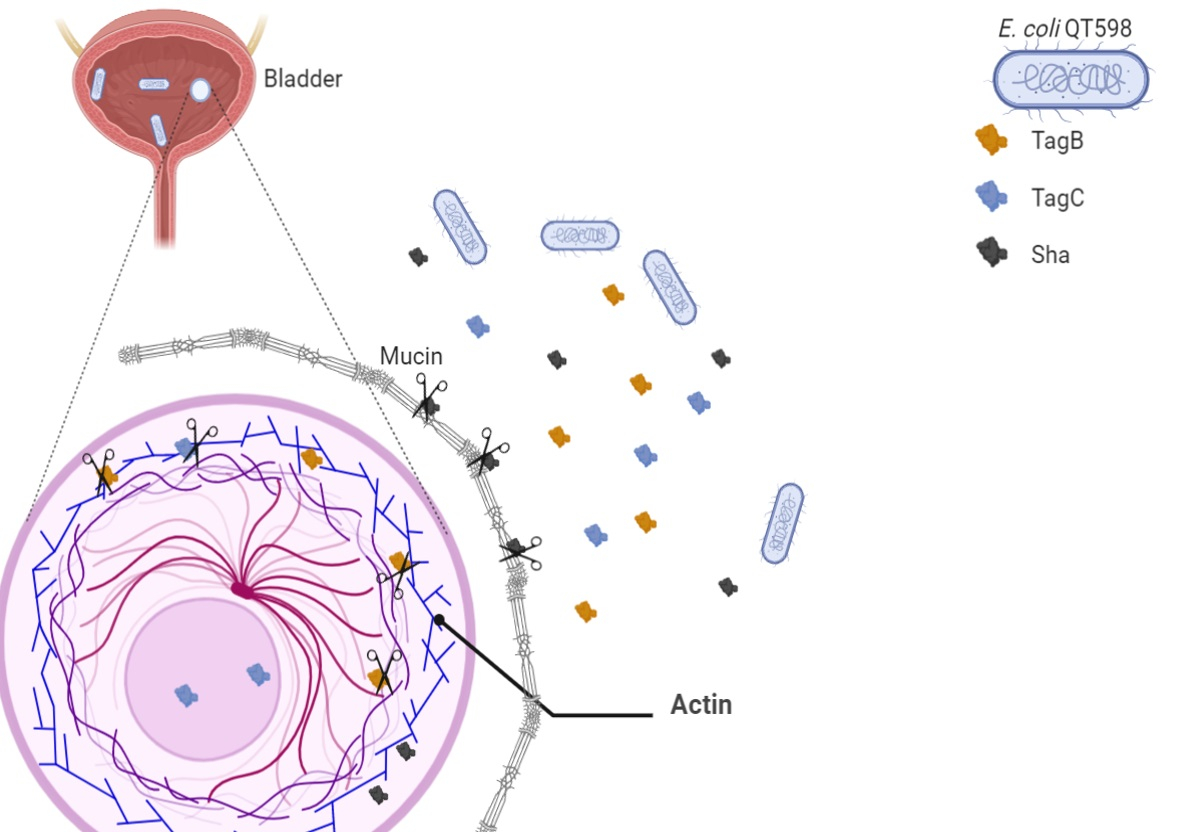
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Bacterial Strains, Plasmids, and Growth Conditions
4.3. Site-Directed Mutagenesis
4.4. Recombinant Protein and Antibody Preparation
4.5. Autoaggregation and Hemagglutination Tests
4.6. Epithelial Cell Culture
4.7. Electron Microscopy
4.8. Cleavage of Protein Substrates
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AIDA-I | Adhesin involved in diffuse adherence |
| ANOVA | Analysis of variance |
| APEC | Avian pathogenic E. coli |
| CNF1 | Cytotoxic necrotizing factor 1 |
| DAPI | 4′,6-Diamidino-2-phenylindole |
| EAEC | Enteroaggregative E. coli |
| EPEC | Enteropathogenic E. coli |
| ExPEC | Extra-intestinal pathogenic E. coli |
| LDH | Lactate dehydrogenase |
| LEE | Locus of enterocyte effacement |
| LPS | Lipopolysaccharides |
| OD | Optical density |
| PAS | Periodic acid–Schiff |
| PMSF | Phenylmethylsulfonyl fluoride |
| SDS-PAGE | Sodium dodecyl sulfate-polyacrylamide gel electrophoresis |
| SPATEs | Serine protease autotransporters of Enterobacteriaceae |
| TEM | Transmission electron microscopy |
| UPEC | Uropathogenic Escherichia coli |
| UTIs | Urinary tract infections |
References
- Foxman, B.; Brown, P. Epidemiology of Urinary Tract Infections: Transmission and Risk Factors, Incidence, and Costs. Infect. Dis. Clin. North Am. 2003, 17, 227–241. [Google Scholar] [CrossRef]
- Foxman, B. Urinary Tract Infection Syndromes: Occurrence, Recurrence, Bacteriology, Risk Factors, and Disease Burden. Infect. Dis. Clin. North Am. 2014, 28, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Flores-Mireles, A.L.; Walker, J.N.; Caparon, M.; Hultgren, S.J. Urinary Tract Infections: Epidemiology, Mechanisms of Infection and Treatment Options. Nature Rev. Microbiol. 2015, 13, 269–284. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, V.P.; Hannan, T.J.; Nielsen, H.V.; Hultgren, S.J. Drug and Vaccine Development for the Treatment and Prevention of Urinary Tract Infections. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Lewis, S.A. Everything You Wanted to Know About the Bladder Epithelium but Were Afraid to Ask. Am. J. Physiol. Ren. Physiol. 2000, 278, F867–F874. [Google Scholar] [CrossRef]
- Keane, W.F.; Welch, R.; Gekker, G.; Peterson, P.K. Mechanism of Escherichia coli Alpha-Hemolysin-Induced Injury to Isolated Renal Tubular Cells. Am. J. Pathol. 1987, 126, 350. [Google Scholar]
- Russo, T.A.; Davidson, B.A.; Genagon, S.A.; Warholic, N.M.; MacDonald, U.; Pawlicki, P.D.; Beanan, J.; Olson, M.R.; Holm, B.A.; Knight, P.R., 3rd. E. coli Virulence Factor Hemolysin Induces Neutrophil Apoptosis and Necrosis/Lysis in Vitro and Necrosis/Lysis and Lung Injury in a Rat Pneumonia Model. Am. J. Physiol. Lung Cell. Mol. Physiol. 2005, 289, L207–L216. [Google Scholar] [CrossRef]
- Smith, Y.C.; Rasmussen, S.B.; Grande, K.K.; Conran, R.M.; O’Brien, A.D. Hemolysin of Uropathogenic Escherichia coli Evokes Extensive Shedding of the Uroepithelium and Hemorrhage in Bladder Tissue within the First 24 Hours after Intraurethral Inoculation of Mice. Infect. Immun. 2008, 76, 2978–2990. [Google Scholar] [CrossRef] [Green Version]
- Dhakal, B.K.; Mulvey, M.A. The Upec Pore-Forming Toxin A-Hemolysin Triggers Proteolysis of Host Proteins to Disrupt Cell Adhesion, Inflammatory, and Survival Pathways. Cell Host Microbe 2012, 11, 58–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiles, T.J.; Mulvey, M.A. The Rtx Pore-Forming Toxin A-Hemolysin of Uropathogenic Escherichia coli: Progress and Perspectives. Future Microbiol. 2013, 8, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Ristow, L.C.; Welch, R.A. Hemolysin of Uropathogenic Escherichia coli: A Cloak or a Dagger? Biochim. Biophys. Acta Biomembr. 2016, 1858, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Visvikis, O.; Boyer, L.; Torrino, S.; Doye, A.; Lemonnier, M.; Lorès, P.; Rolando, M.; Flatau, G.; Mettouchi, A.; Bouvard, D. Escherichia coli Producing Cnf1 Toxin Hijacks Tollip to Trigger Rac1-Dependent Cell Invasion. Traffic 2011, 12, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Mills, M.; Meysick, K.C.; O’Brien, A.D. Cytotoxic Necrotizing Factor Type 1 of Uropathogenic Escherichia coli Kills Cultured Human Uroepithelial 5637 Cells by an Apoptotic Mechanism. Infect. Immun. 2000, 68, 5869–5880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guyer, D.M.; Radulovic, S.; Jones, F.-E.; Mobley, H.L.T. Sat, the Secreted Autotransporter Toxin of Uropathogenic Escherichia coli, Is a Vacuolating Cytotoxin for Bladder and Kidney Epithelial Cells. Infect. Immun. 2002, 70, 4539–4546. [Google Scholar] [CrossRef] [Green Version]
- Maroncle, N.M.; Sivick, K.E.; Brady, R.; Stokes, F.-E.; Mobley, H.L.T. Protease Activity, Secretion, Cell Entry, Cytotoxicity, and Cellular Targets of Secreted Autotransporter Toxin of Uropathogenic Escherichia coli. Infect. Immun. 2006, 74, 6124–6134. [Google Scholar] [CrossRef] [Green Version]
- Heimer, S.R.; Rasko, D.A.; Lockatell, C.V.; Johnson, D.E.; Mobley, H.L.T. Autotransporter Genes Pic and Tsh Are Associated with Escherichia coli Strains That Cause Acute Pyelonephritis and Are Expressed During Urinary Tract Infection. Infect. Immun. 2004, 72, 593–597. [Google Scholar] [CrossRef] [Green Version]
- Carter, P.; Wells, J.A. Dissecting the Catalytic Triad of a Serine Protease. Nature 1988, 332, 564. [Google Scholar] [CrossRef]
- Gutierrez-Jimenez, J.; Arciniega, I.; Navarro-García, F. The Serine Protease Motif of Pic Mediates a Dose-Dependent Mucolytic Activity after Binding to Sugar Constituents of the Mucin Substrate. Microb. Pathog. 2008, 45, 115–123. [Google Scholar] [CrossRef]
- Navarro-Garcia, F.; Gutierrez-Jimenez, J.; Garcia-Tovar, C.; Castro, L.A.; Salazar-Gonzalez, H.; Cordova, V. Pic, an Autotransporter Protein Secreted by Different Pathogens in the Enterobacteriaceae Family, Is a Potent Mucus Secretagogue. Infect. Immun. 2010, 78, 4101–4109. [Google Scholar] [CrossRef] [Green Version]
- Habouria, H.; Pokharel, P.; Maris, S.; Garénaux, A.; Bessaiah, H.; Houle, S.; Veyrier, F.J.; Guyomard-Rabenirina, S.; Talarmin, A.; Dozois, C.M. Three New Serine-Protease Autotransporters of Enterobacteriaceae (SPATEs) from Extra-Intestinal Pathogenic Escherichia coli and Combined Role of Spates for Cytotoxicity and Colonization of the Mouse Kidney. Virulence 2019, 10, 568–587. [Google Scholar] [CrossRef] [Green Version]
- Benz, I.; Alexander Schmidt, M. Aida-I, the Adhesin Involved in Diffuse Adherence of the Diarrhoeagenic Escherichia coli Strain 2787 (O126: H27), Is Synthesized Via a Precursor Molecule. Mol. Microbiol. 1992, 6, 1539–1546. [Google Scholar] [CrossRef] [PubMed]
- Abràmoff, M.D.; Magalhães, P.J.; Ram, S.J. Image Processing with Imagej. Biophotonics Int. 2004, 11, 36–42. [Google Scholar]
- Henderson, I.R.; Czeczulin, J.; Eslava, C.; Noriega, F.; Nataro, J.P. Characterization of Pic, a Secreted Protease Ofshigella Flexneri and Enteroaggregative Escherichia coli. Infect. Immun. 1999, 67, 5587–5596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickett, M.J.; Greenwood, J.R.; Harvey, S.M. Tests for Detecting Degradation of Gelatin: Comparison of Five Methods. J. Clin. Microbiol. 1991, 29, 2322–2325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xicohtencatl-Cortes, J.; Saldaña, Z.; Deng, W.; Castañeda, E.; Freer, E.; Tarr, P.I.; Finlay, B.B.; Puente, J.L.; Girón, J.A. Bacterial Macroscopic Rope-Like Fibers with Cytopathic and Adhesive Properties. J. Biol. Chem. 2010, 285, 32336–32342. [Google Scholar] [CrossRef] [Green Version]
- Vigil, P.D.; Wiles, T.J.; Engstrom, M.D.; Prasov, L.; Mulvey, M.A.; Mobley, H.L.T. The Repeat-in-Toxin Family Member Tosa Mediates Adherence of Uropathogenic Escherichia coli and Survival During Bacteremia. Infect. Immun. 2012, 80, 493–505. [Google Scholar] [CrossRef] [Green Version]
- Rippere-Lampe, K.E.; O’Brien, A.D.; Conran, R.; Lockman, H.A. Mutation of the Gene Encoding Cytotoxic Necrotizing Factor Type 1 (Cnf 1) Attenuates the Virulence of Uropathogenic Escherichia coli. Infect. Immun. 2001, 69, 3954–3964. [Google Scholar] [CrossRef] [Green Version]
- Davis, J.M.; Rasmussen, S.B.; O’Brien, A.D. Cytotoxic Necrotizing Factor Type 1 Production by Uropathogenic Escherichia coli Modulates Polymorphonuclear Leukocyte Function. Infect. Immun. 2005, 73, 5301–5310. [Google Scholar] [CrossRef] [Green Version]
- Pokharel, P.; Habouria, H.; Bessaiah, H.; Dozois, C.M. Serine Protease Autotransporters of the Enterobacteriaceae (SPATEs): Out and About and Chopping It Up. Microorganisms 2019, 7, 594. [Google Scholar] [CrossRef] [Green Version]
- Martinez, J.J.; Mulvey, M.A.; Schilling, J.D.; Pinkner, J.S.; Hultgren, S.J. Type 1 Pilus-Mediated Bacterial Invasion of Bladder Epithelial Cells. EMBO J. 2000, 19, 2803–2812. [Google Scholar] [CrossRef]
- Mulvey, M.A.; Schilling, J.D.; Hultgren, S.J. Establishment of a Persistent Escherichia coli Reservoir During the Acute Phase of a Bladder Infection. Infect. Immun. 2001, 69, 4572–4579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eslava, C.; Navarro-García, F.; Czeczulin, J.R.; Henderson, I.R.; Cravioto, A.; Nataro, J.P. Pet, an Autotransporter Enterotoxin from Enteroaggregative Escherichia coli. Infect. Immune. 1998, 66, 3155–3163. [Google Scholar] [CrossRef] [Green Version]
- Brunder, W.; Schmidt, H.; Karch, H. Espp, a Novel Extracellular Serine Protease of Enterohaemorrhagic Escherichia coli O157: H7 Cleaves Human Coagulation Factor, V. Mol. Microbiol. 1997, 24, 767–778. [Google Scholar] [CrossRef]
- Dozois, C.M.; Dho-Moulin, M.; Brée, A.; Fairbrother, J.M.; Desautels, C.; Curtiss, R. Relationship between the Tsh Autotransporter and Pathogenicity of Avian Escherichia coli and Localization and Analysis of the Tsh Genetic Region. Infect. Immun. 2000, 68, 4145–4154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, M.; Kenny, B.; Stein, M.A.; Finlay, B.B. Characterization of Espc, a 110-Kilodalton Protein Secreted by Enteropathogenic Escherichia coli Which Is Homologous to Members of the Immunoglobulin a Protease-Like Family of Secreted Proteins. J. Bacteriol. 1996, 178, 6546–6554. [Google Scholar] [CrossRef] [Green Version]
- Dutta, P.R.; Cappello, R.; Navarro-García, F.; Nataro, J.P. Functional Comparison of Serine Protease Autotransporters of Enterobacteriaceae. Infect. Immun. 2002, 70, 7105–7113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, D.P.J.; Wooldridge, K.G.; Ala’Aldeen, D.A.A. Autotransported Serine Protease a of Neisseria Meningitidis: An Immunogenic, Surface-Exposed Outer Membrane, and Secreted Protein. Infect. Immun. 2002, 70, 4447–4461. [Google Scholar] [CrossRef] [Green Version]
- Jonsson, P.; Wadström, T. Cell Surface Hydrophobicity Ofstaphylococcus Aureus Measured by the Salt Aggregation Test (Sat). Curr. Microbiol. 1984, 10, 203–209. [Google Scholar] [CrossRef]
- Hendrixson, D.R.; De La Morena, M.L.; Stathopoulos, C.; St Geme, J.W., 3rd. Structural Determinants of Processing and Secretion of the Haemophilus Influenzae Hap Protein. Mol. Microbiol. 1997, 26, 505–518. [Google Scholar] [CrossRef]
- Hendrixson, D.R.; St Geme, J.W., 3rd. The Haemophilus Influenzae Hap Serine Protease Promotes Adherence and Microcolony Formation, Potentiated by a Soluble Host Protein. Mol. Cell 1998, 2, 841–850. [Google Scholar] [CrossRef]
- Kostakioti, M.; Stathopoulos, C. Functional Analysis of the Tsh Autotransporter from an Avian Pathogenic Escherichia coli Strain. Infect. Immun. 2004, 72, 5548–5554. [Google Scholar] [CrossRef] [Green Version]
- Brockmeyer, J.; Spelten, S.; Kuczius, T.; Bielaszewska, M.; Karch, H. Structure and Function Relationship of the Autotransport and Proteolytic Activity of Espp from Shiga Toxin-Producing Escherichia coli. PLoS ONE 2009, 4, e6100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro-Garcia, F.; Serapio-Palacios, A.; Vidal, J.E.; Salazar, M.I.; Tapia-Pastrana, G. Espc Promotes Epithelial Cell Detachment by Enteropathogenic Escherichia coli Via Sequential Cleavages of a Cytoskeletal Protein and Then Focal Adhesion Proteins. Infect. Immun. 2014, 82, 2255–2265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruenheid, S.; Finlay, B.B. Microbial Pathogenesis and Cytoskeletal Function. Nature 2003, 422, 775. [Google Scholar] [CrossRef]
- Rottner, K.; Stradal, T.E.B.; Wehland, J. Bacteria-Host-Cell Interactions at the Plasma Membrane: Stories on Actin Cytoskeleton Subversion. Dev. Cell 2005, 9, 3–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, J.J.; Hultgren, S.J. Requirement of Rho-Family Gtpases in the Invasion of Type 1-Piliated Uropathogenic Escherichia coli. Cell. Microbiol. 2002, 4, 19–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eto, D.S.; Sundsbak, J.L.; Mulvey, M.A. Actin-Gated Intracellular Growth and Resurgence of Uropathogenic Escherichia coli. Cell. Microbiol. 2006, 8, 704–717. [Google Scholar] [CrossRef]
- Balish, M.J.; Jensen, J.; Uehling, D.T. Bladder Mucin: A Scanning Electron Microscopy Study in Experimental Cystitis. J. Urol. 1982, 128, 1060–1063. [Google Scholar] [CrossRef]
- Parsons, C.L.; Greenspan, C.; Moore, S.W.; Grant, S. Mulholland. Role of Surface Mucin in Primary Antibacterial Defense of Bladder. Urology 1977, 9, 48–52. [Google Scholar] [CrossRef]
- Lehker, M.W.; Sweeney, D. Trichomonad Invasion of the Mucous Layer Requires Adhesins, Mucinases, and Motility. Sex. Transm. Infect. 1999, 75, 231–238. [Google Scholar] [CrossRef] [Green Version]
- Mantle, M.; Husar, S.D. Binding of Yersinia Enterocolitica to Purified, Native Small Intestinal Mucins from Rabbits and Humans Involves Interactions with the Mucin Carbohydrate Moiety. Infect. Immun. 1994, 62, 1219–1227. [Google Scholar] [CrossRef] [Green Version]
- Finkelstein, R.A.; Boesman-Finkelstein, M.; Holt, P. Vibrio Cholerae Hemagglutinin/Lectin/Protease Hydrolyzes Fibronectin and Ovomucin: Fm Burnet Revisited. Proc. Natl. Acad. Sci. USA 1983, 80, 1092–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhullar, K.; Zarepour, M.; Yu, H.; Yang, H.; Croxen, M.; Stahl, M.; Finlay, B.B.; Turvey, S.E.; Vallance, B.A. The Serine Protease Autotransporter Pic Modulates Citrobacter Rodentium Pathogenesis and Its Innate Recognition by the Host. Infect. Immun. 2015, 83, 2636–2650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, R.K.T.; Gaziri, L.C.J.; Vidotto, M.C. Functional Activities of the Tsh Protein from Avian Pathogenic Escherichia coli (APEC) Strains. J. Vet. Sci. 2010, 11, 315–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricard-Blum, S. The Collagen Family. Cold Spring Harbor Perspect. Biol. 2011, 3, a004978. [Google Scholar] [CrossRef] [Green Version]
- Navarro-García, F.; Canizalez-Roman, A.; Burlingame, K.E.; Teter, K.; Vidal, J.E. Pet, a Non-Ab Toxin, Is Transported and Translocated into Epithelial Cells by a Retrograde Trafficking Pathway. Infect. Immun. 2007, 75, 2101–2109. [Google Scholar]
- Nava-Acosta, R.; Navarro-Garcia, F. Cytokeratin 8 Is an Epithelial Cell Receptor for Pet, a Cytotoxic Serine Protease Autotransporter of Enterobacteriaceae. MBio 2013, 4, e00838-13. [Google Scholar] [CrossRef] [Green Version]
- Navarro-García, F.; Sears, C.; Eslava, C.; Cravioto, A.; Nataro, J.P. Cytoskeletal Effects Induced by Pet, the Serine Protease Enterotoxin of Enteroaggregative Escherichia coli. Infect. Immun. 1999, 67, 2184–2192. [Google Scholar]
- Dutta, P.R.; Sui, B.Q.; Nataro, J.P. Structure-Function Analysis of the Enteroaggregative Escherichia coli Plasmid-Encoded Toxin Autotransporter Using Scanning Linker Mutagenesis. J. Biol. Chem. 2003, 278, 39912–39920. [Google Scholar] [CrossRef] [Green Version]
- Vidal, J.E.; Navarro-García, F. Espc Translocation into Epithelial Cells by Enteropathogenic Escherichia coli Requires a Concerted Participation of Type V and Iii Secretion Systems. Cell. Microbiol. 2008, 10, 1975–1986. [Google Scholar] [CrossRef]
- Marc, D.; Dho-Moulin, M. Analysis of the Fim Cluster of an Avian O2 Strain of Escherichia coli: Serogroup-Specific Sites within Fima and Nucleotide Sequence of Fimi. J. Med Microbiol. 1996, 44, 444–452. [Google Scholar] [CrossRef] [Green Version]
- Woodall, L.D.; Russell, P.W.; Harris, S.L.; Orndorff, P.E. Rapid, Synchronous, and Stable Induction of Type 1 Piliation in Escherichia coli by Using a Chromosomal Lacuv5 Promoter. J. Bacteriol. 1993, 175, 2770–2778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charbonneau, M.-È.; Berthiaume, F.; Mourez, M. Proteolytic Processing Is Not Essential for Multiple Functions of the Escherichia coli Autotransporter Adhesin Involved in Diffuse Adherence (Aida-I). J. Bacteriol. 2006, 188, 8504–8512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakazawa, M.; Mukumoto, M.; Miyatake, K. Production and Purification of Polyclonal Antibodies. Methods Mol. Biol. (Clifton, N.J.). 2016. [Google Scholar] [CrossRef]
- Mellies, J.L.; Navarro-Garcia, F.; Okeke, I.; Frederickson, J.; Nataro, J.P.; Kaper, J.B. Espc Pathogenicity Island of Enteropathogenic Escherichia coli Encodes an Enterotoxin. Infect. Immun. 2001, 69, 315–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parham, N.J.; Srinivasan, U.; Desvaux, M.; Foxman, B.; Marrs, C.F.; Henderson, I.R. Picu, a Second Serine Protease Autotransporter of Uropathogenic Escherichia coli. FEMS Microbiol. Lett. 2004, 230, 73–83. [Google Scholar] [CrossRef]
- Provence, D.L.; Curtiss, R., 3rd. Role of Crl in Avian Pathogenic Escherichia coli: A Knockout Mutation of Crl Does Not Affect Hemagglutination Activity, Fibronectin Binding, or Curli Production. Infect. Immun. 1992, 60, 4460–4467. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Characteristic(s) | Reference |
|---|---|---|
| QT598 | APEC O1: K1, serum resistant | [61] |
| ORN172 | thr-1 leuB thi-1Δ (argF-lac)U169 xyl-7 ara-13 mtl-2 gal-6 rpsL tonA2 supE44Δ (fimBEACDFGH)::Km pilG1 | [62] |
| BL21 | fhuA2 [lon] ompT gal [dcm] ΔhsdS | New England Biolabs |
| QT1603 | HB101 with AIDA-1 operon | [63] |
| QT4767 | ORN172/pIJ553 (Expressing sha) | [20] |
| QT5194 | BL21/pIJ548 (Expressing tagB) | |
| QT5195 | BL21/pIJ549 (Expressing tagC) | |
| QT5197 | BL21/pIJ550 (Expressing espC) | |
| QT5198 | ORN172/pIJ548 (Expressing tagB) | |
| QT5199 | ORN172/pIJ549 (Expressing tagC) | |
| QT5431 | BL21/pIJ551 (Expressing vat) | |
| QT5432 | BL21/pIJ552 (Expressing tsh) | |
| QT5433 | BL21/pIJ553 (Expressing sha) | |
| QT5437 | BL21 + pIJ554 (Expressing tagB S255A) | This study |
| QT5438 | BL21 + pIJ555 (Expressing tagC S252A) | This study |
| QT5439 | BL21 + pIJ556 (Expressing sha S258A) | This study |
| QT5598 | ORN172 + pIJ554 (Expressing tagB S255A) | This study |
| QT5599 | ORN172 + pIJ555 (Expressing tagC S252A) | This study |
| QT5600 | ORN172 + pIJ556 (Expressing sha S258A) | This study |
| QT3046 | Pseudomonas aeruginosa PA14 | Eric Déziel, INRS |
| Plasmids | ||
| pBCsk+ | Cloning vector; Cmr | Stratagene, La Jolla, CA |
| pIJ548 | pBCsk+::tagB | [20] |
| pIJ549 | pBCsk+::tagC | |
| pIJ551 | pBCsk+::vat | |
| pIJ552 | pBCsk+::tsh | |
| pIJ553 | pBCsk+::sha | |
| pIJ554 | pBCsk+::tagB S255A | This study |
| pIJ555 | pBCsk+::tagC S252A | This study |
| pIJ556 | pBCsk+::sha S258A | This study |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pokharel, P.; Díaz, J.M.; Bessaiah, H.; Houle, S.; Guerrero-Barrera, A.L.; Dozois, C.M. The Serine Protease Autotransporters TagB, TagC, and Sha from Extraintestinal Pathogenic Escherichia coli Are Internalized by Human Bladder Epithelial Cells and Cause Actin Cytoskeletal Disruption. Int. J. Mol. Sci. 2020, 21, 3047. https://doi.org/10.3390/ijms21093047
Pokharel P, Díaz JM, Bessaiah H, Houle S, Guerrero-Barrera AL, Dozois CM. The Serine Protease Autotransporters TagB, TagC, and Sha from Extraintestinal Pathogenic Escherichia coli Are Internalized by Human Bladder Epithelial Cells and Cause Actin Cytoskeletal Disruption. International Journal of Molecular Sciences. 2020; 21(9):3047. https://doi.org/10.3390/ijms21093047
Chicago/Turabian StylePokharel, Pravil, Juan Manuel Díaz, Hicham Bessaiah, Sébastien Houle, Alma Lilián Guerrero-Barrera, and Charles M. Dozois. 2020. "The Serine Protease Autotransporters TagB, TagC, and Sha from Extraintestinal Pathogenic Escherichia coli Are Internalized by Human Bladder Epithelial Cells and Cause Actin Cytoskeletal Disruption" International Journal of Molecular Sciences 21, no. 9: 3047. https://doi.org/10.3390/ijms21093047
APA StylePokharel, P., Díaz, J. M., Bessaiah, H., Houle, S., Guerrero-Barrera, A. L., & Dozois, C. M. (2020). The Serine Protease Autotransporters TagB, TagC, and Sha from Extraintestinal Pathogenic Escherichia coli Are Internalized by Human Bladder Epithelial Cells and Cause Actin Cytoskeletal Disruption. International Journal of Molecular Sciences, 21(9), 3047. https://doi.org/10.3390/ijms21093047