Endoplasmic Reticulum Calcium Pumps and Tumor Cell Differentiation

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
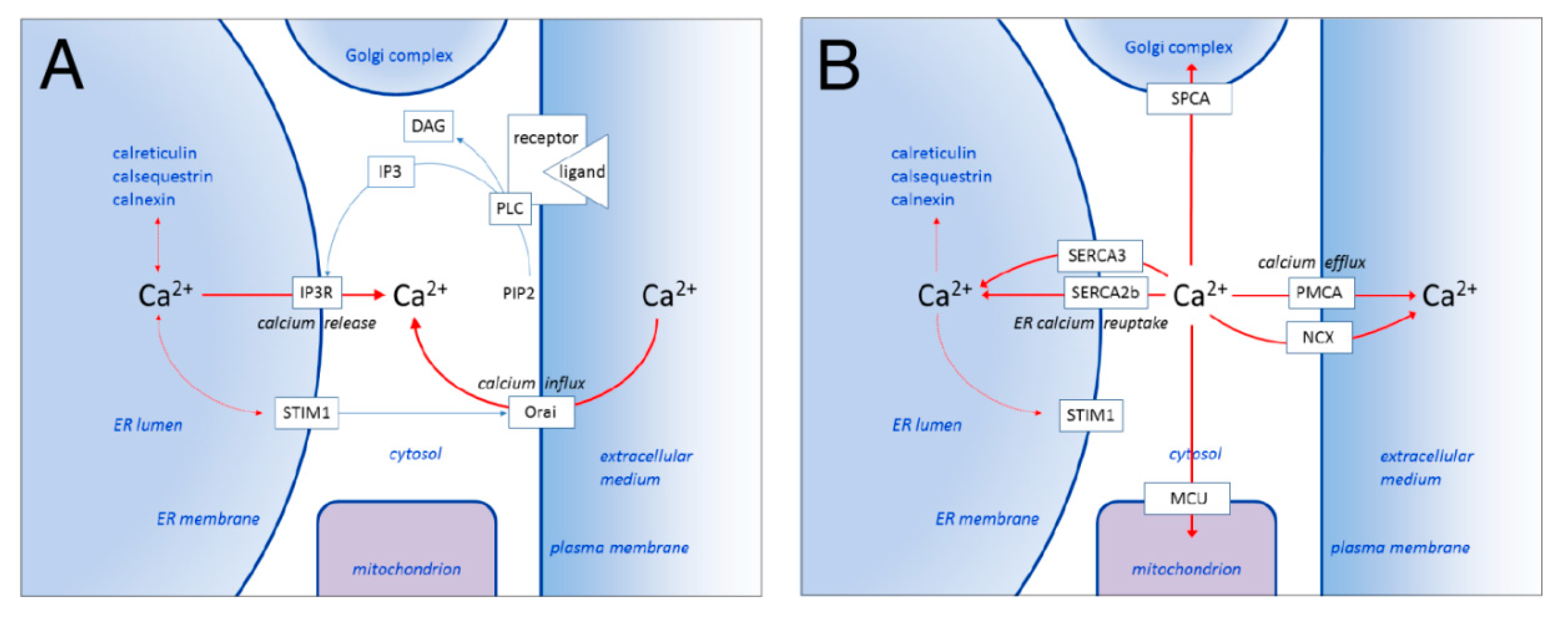
1.1. ER Calcium Homeostasis and SERCA
1.2. SERCA Pharmacology
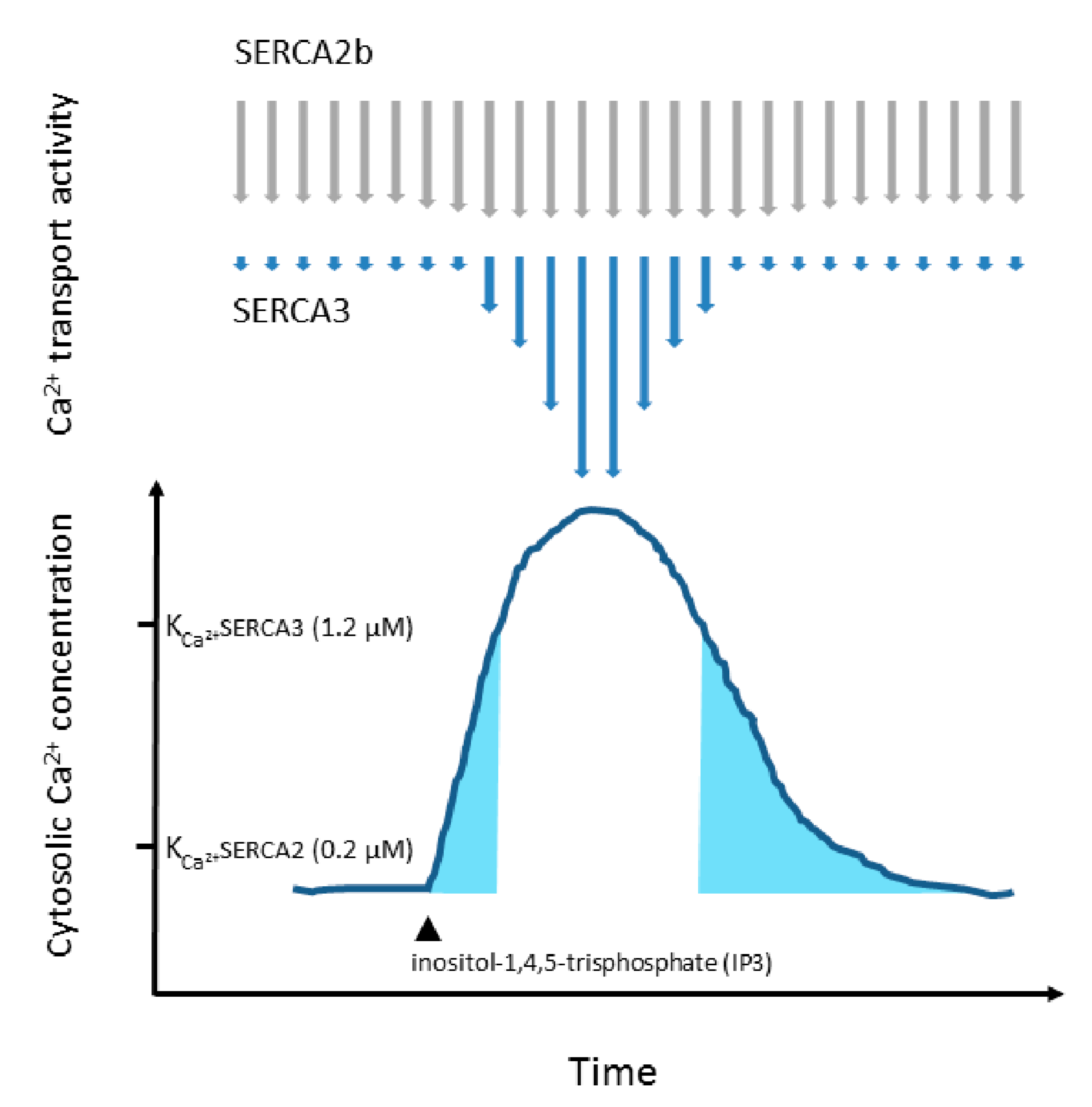
1.3. SERCA Isoenzymes
1.4. Co-Expression of Various SERCA Isoenzymes
2. Loss of SERCA3 Expression in Tumors
2.1. Acute Promyelocytic Leukemia
2.2. Pre-B Cell Acute Lymphoid Leukemia
2.3. Megakaryocytic Differentiation
2.4. Down-Regulation of SERCA3 Expression In Vitro
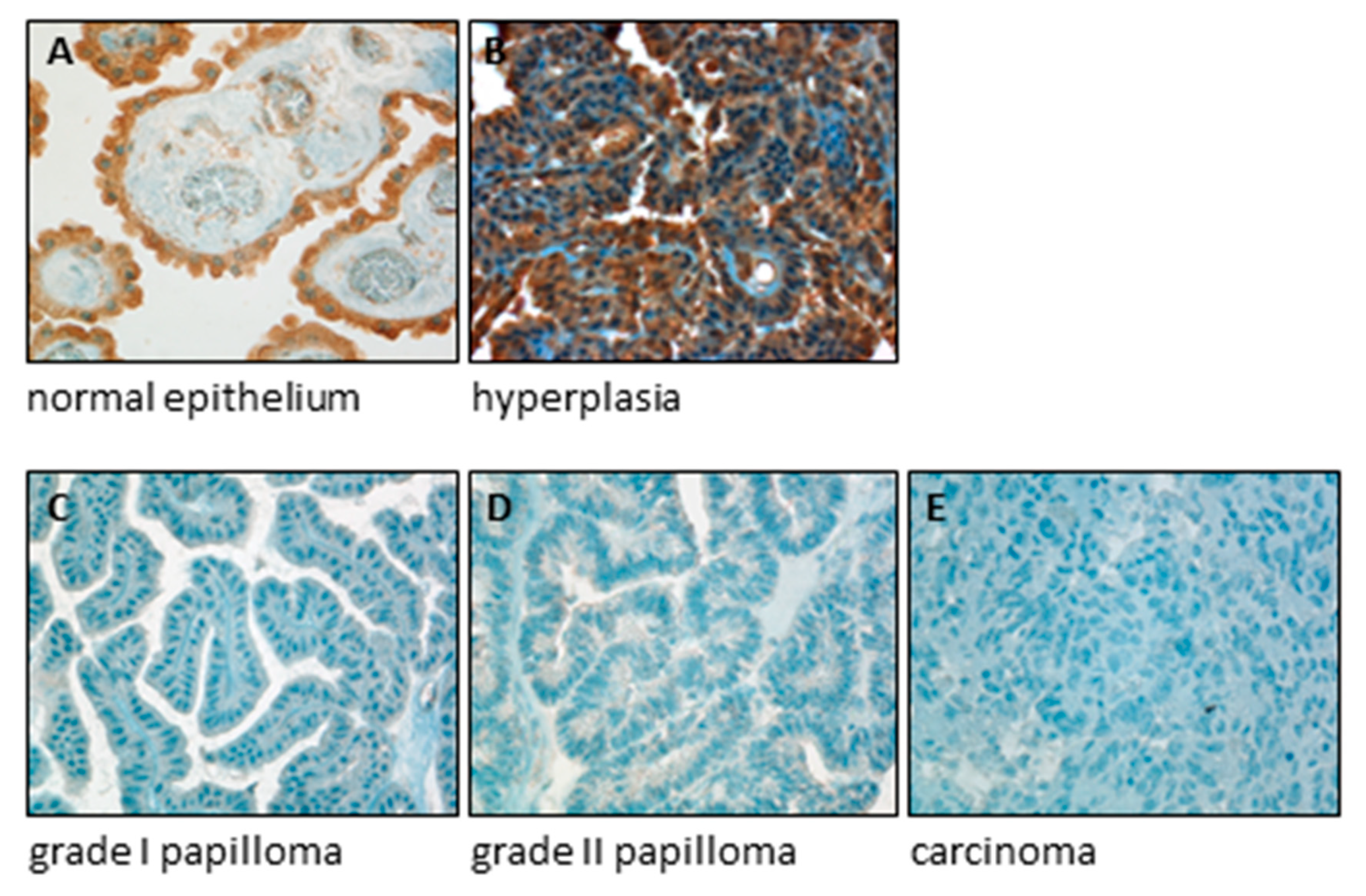
2.5. SERCA3 Loss in Gastrointestinal Carcinomas
2.6. Lung Cancer
2.7. Breast Cancer
2.8. Choroid Plexus
3. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| APL | acute promyelocytic leukemia |
| ATRA | all-trans retinoic acid |
| DAG | diacylglycerol |
| E2A | E2A immunoglobulin enhancer-binding factor E12/E47 |
| EBNA-2 | Epstein-Barr virus nuclear antigen 2 |
| EBV | Epstein-Barr virus |
| ER | endoplasmic reticulum |
| ERK | extracellular signal-regulated kinase |
| IL-2 | interleukin-2 |
| IP3 | inositol 1,4,5-trisphosphate |
| LMP1 | Epstein-Barr virus latent membrane protein 1 |
| MCU | mitochondrial calcium uniporter |
| NCX | sodium/calcium exchanger |
| PBX1 | pre-B-cell leukemia transcription factor 1 |
| PIP2 | phosphatidylinositol 4,5-bisphosphate |
| PLC | phospholipase C |
| PMA | phorbol 12-myristate 13-acetate |
| PMCA | plasma membrane calcium ATPase |
| PML | promyelocytic leukemia protein |
| RAG-1 | recombination activating gene 1 |
| SERCA | sarco/endoplasmic reticulum calcium ATPase |
| SPCA | secretory pathway calcium ATPase |
| STIM | stromal interaction molecule |
| TdT | terminal deoxynucleotidyl transferase |
References
- Berridge, M.J.; Bootman, M.; Roderick, H.L. Calcium signalling: Dynamics, homeostasis and remodelling. Nat. Rev. Mol. Cell Biol. 2003, 4, 517–529. [Google Scholar] [CrossRef] [Green Version]
- Berridge, M.J. The endoplasmic reticulum: A multifunctional signaling organelle. Cell Calcium 2002, 32, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.J. Inositol trisphosphate and calcium signalling mechanisms. Biochim. Biophys. Acta (BBA) Bioenerg. 2009, 1793, 933–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shim, A.H.-R.; Tirado-Lee, L.; Prakriya, M. Structural and Functional Mechanisms of CRAC Channel Regulation. J. Mol. Biol. 2015, 427, 77–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, J.J.; Jardin, I.; Albarrán, L.; Sanchez-Collado, J.; Cantonero, C.; Salido, G.M.; Smani, T.; Rosado, J.A. Molecular Basis and Regulation of Store-Operated Calcium Entry. In Advances in Experimental Medicine and Biology; Springer: Berlin/Heidelberg, Germany, 2020; Volume 1131, pp. 445–469. [Google Scholar]
- Berridge, M.J. Unlocking The Secrets Of Cell Signaling. Annu. Rev. Physiol. 2005, 67, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, A.; Galione, A. The endoplasmic reticulum and junctional membrane communication during calcium signaling. Biochim. Biophys. Acta (BBA) Bioenerg. 2013, 1833, 2542–2559. [Google Scholar] [CrossRef] [Green Version]
- Taylor, C.; Tovey, S.C. IP3 Receptors: Toward Understanding Their Activation. Cold Spring Harb. Perspect. Biol. 2010, 2, a004010. [Google Scholar] [CrossRef]
- Bublitz, M.; Musgaard, M.; Poulsen, H.; Thøgersen, L.; Olesen, C.; Schiøtt, B.; Morth, J.P.; Møller, J.V.; Nissen, P. Ion Pathways in the Sarcoplasmic Reticulum Ca2+-ATPase. J. Biol. Chem. 2013, 288, 10759–10765. [Google Scholar] [CrossRef] [Green Version]
- Vandecaetsbeek, I.; Vangheluwe, P.; Raeymaekers, L.; Wuytack, F.; Vanoevelen, J. The Ca2+ Pumps of the Endoplasmic Reticulum and Golgi Apparatus. Cold Spring Harb. Perspect. Biol. 2011, 3, a004184. [Google Scholar] [CrossRef] [Green Version]
- Wuytack, F.; Raeymaekers, L.; Missiaen, L. Molecular physiology of the SERCA and SPCA pumps. Cell Calcium 2002, 32, 279–305. [Google Scholar] [CrossRef]
- Inesi, G.; Lewis, D.; Ma, H.; Prasad, A.; Toyoshima, C. Concerted Conformational Effects of Ca2+and ATP Are Required for Activation of Sequential Reactions in the Ca Ca2+ATPase (SERCA) Catalytic Cycle. Biochemistry 2006, 45, 13769–13778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inesi, G.; Lewis, D.; Toyoshima, C.; Hirata, A.; De Meis, L. Conformational Fluctuations of the Ca Ca2+-ATPase in the Native Membrane Environment: Effects Of pH, temperature, catalytic substrates, and thapsigargin. J. Biol. Chem. 2007, 283, 1189–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, H.; Lewis, D.; Xu, C.; Inesi, G.; Toyoshima, C. Functional and Structural Roles of Critical Amino Acids within the “N”, “P”, and “A” Domains of the Ca Ca2+ATPase (SERCA) Headpiece. Biochemistry 2005, 44, 8090–8100. [Google Scholar] [CrossRef] [PubMed]
- Tsunekawa, N.; Ogawa, H.; Tsueda, J.; Akiba, T.; Toyoshima, C. Mechanism of the E2 to E1 transition in Ca2+ pump revealed by crystal structures of gating residue mutants. Proc. Natl. Acad. Sci. USA 2018, 115, 12722–12727. [Google Scholar] [CrossRef] [Green Version]
- Møller, J.V.; Olesen, C.; Winther, A.-M.L.; Nissen, P. The sarcoplasmic Ca2+-ATPase: Design of a perfect chemi-osmotic pump. Q. Rev. Biophys. 2010, 43, 501–566. [Google Scholar] [CrossRef]
- Dyla, M.; Kjærgaard, M.; Poulsen, H.; Nissen, P. Structure and Mechanism of P-Type ATPase Ion Pumps. Annu. Rev. Biochem. 2019, 89, 1146. [Google Scholar] [CrossRef]
- Dyla, M.; Hansen, S.B.; Nissen, P.; Kjaergaard, M. Structural dynamics of P-type ATPase ion pumps. Biochem. Soc. Trans. 2019, 47, 1247–1257. [Google Scholar] [CrossRef]
- Brini, M.; Carafoli, E. The Plasma Membrane Ca2+ ATPase and the Plasma Membrane Sodium Calcium Exchanger Cooperate in the Regulation of Cell Calcium. Cold Spring Harb. Perspect. Biol. 2010, 3, a004168. [Google Scholar] [CrossRef]
- Stafford, N.; Wilson, C.; Oceandy, D.; Neyses, L.; Cartwright, E.J. The Plasma Membrane Calcium ATPases and Their Role as Major New Players in Human Disease. Physiol. Rev. 2017, 97, 1089–1125. [Google Scholar] [CrossRef] [Green Version]
- Pizzo, P.; Lissandron, V.; Pozzan, T. The trans-golgi compartment. Commun. Integr. Biol. 2010, 3, 462–464. [Google Scholar] [CrossRef] [Green Version]
- Vanoevelen, J.; Dode, L.; Van Baelen, K.; Fairclough, R.J.; Missiaen, L.; Raeymaekers, L.; Wuytack, F. The Secretory Pathway Ca2+/Mn2+-ATPase 2 Is a Golgi-localized Pump with High Affinity for Ca2+ Ions. J. Biol. Chem. 2005, 280, 22800–22808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belosludtsev, K.N.; Dubinin, M.V.; Belosludtseva, N.V.; Mironova, G.D. Mitochondrial Ca2+ Transport: Mechanisms, Molecular Structures, and Role in Cells. Biochemistry 2019, 84, 593–607. [Google Scholar] [CrossRef] [PubMed]
- Penna, E.; Espino, J.; De Stefani, D.; Rizzuto, R. The MCU complex in cell death. Cell Calcium 2018, 69, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Dupont, G.; Houart, G.; De Koninck, P. Sensitivity of CaM kinase II to the frequency of Ca2+ oscillations: A simple model. Cell Calcium 2003, 34, 485–497. [Google Scholar] [CrossRef]
- Lewis, R.S. Calcium oscillations in T-cells: Mechanisms and consequences for gene expression. Biochem. Soc. Trans. 2003, 31, 925–929. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Li, J.; Zhu, L.; Cai, L.; Xu, Q.; Ling, C.; Su, Y.; Hu, Q. Irregular Ca2+ Oscillations Regulate Transcription via Cumulative Spike Duration and Spike Amplitude. J. Biol. Chem. 2012, 287, 40246–40255. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Song, S.; Pi, Y.; Yu, Y.; She, W.; Ye, H.; Su, Y.; Hu, Q. Cumulated Ca2+ spike duration underlies Ca2+ oscillation frequency-regulated NF B transcriptional activity. J. Cell Sci. 2011, 124, 2591–2601. [Google Scholar] [CrossRef] [Green Version]
- Hu, Q.; Deshpande, S.; Irani, K.; Ziegelstein, R.C. [Ca2+]i Oscillation Frequency Regulates Agonist-stimulated NF-κB Transcriptional Activity. J. Biol. Chem. 1999, 274, 33995–33998. [Google Scholar] [CrossRef] [Green Version]
- Dolmetsch, R.E.; Xu, K.; Lewis, R.S. Calcium oscillations increase the efficiency and specificity of gene expression. Nature 1998, 392, 933–936. [Google Scholar] [CrossRef]
- Marcello, M.; White, M.R.H. Spatial and temporal information coding and noise in the NF-κB system. Biochem. Soc. Trans. 2010, 38, 1247–1250. [Google Scholar] [CrossRef]
- Raguimova, O.N.; Smolin, N.; Bovo, E.; Bhayani, S.; Autry, J.M.; Zima, A.V.; Robia, S.L. Redistribution of SERCA calcium pump conformers during intracellular calcium signaling. J. Biol. Chem. 2018, 293, 10843–10856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cicek, F.A.; Ozgur, E.O.; Ozgur, E.; Uğur, M. The interplay between plasma membrane and endoplasmic reticulum Ca2+ATPases in agonist-induced temporal Ca2+ dynamics. J. Bioenerg. Biomembr. 2014, 46, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Bandara, S.; Malmersjö, S.; Meyer, T. Regulators of Calcium Homeostasis Identified by Inference of Kinetic Model Parameters from Live Single Cells Perturbed by siRNA. Sci. Signal. 2013, 6, ra56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupont, G.; Combettes, L.; Bird, G.S.; Putney, J. Calcium Oscillations. Cold Spring Harb. Perspect. Biol. 2010, 3, a004226. [Google Scholar] [CrossRef] [Green Version]
- Neymotin, S.A.; McDougal, R.A.; Sherif, M.A.; Fall, C.P.; Hines, M.L.; Lytton, W.W. Neuronal calcium wave propagation varies with changes in endoplasmic reticulum parameters: A computer model. Neural Comput. 2015, 27, 898–924. [Google Scholar] [CrossRef] [Green Version]
- Bertram, R.; Arceo, R.C. A Mathematical Study of the Differential Effects of Two SERCA Isoforms on Ca2+ Oscillations in Pancreatic Islets. Bull. Math. Biol. 2008, 70, 1251–1271. [Google Scholar] [CrossRef]
- Bayles, I.; Milcarek, C. Plasma cell formation, secretion, and persistence: The short and the long of it. Crit. Rev. Immunol. 2014, 34, 481–499. [Google Scholar] [CrossRef]
- Yang, P.-C.; Jafri, M.S. Ca2+ signaling in T lymphocytes: The interplay of the endoplasmic reticulum, mitochondria, membrane potential, and CRAC channels on transcription factor activation. Heliyon 2020, 6, e03526. [Google Scholar] [CrossRef]
- Raeymaekers, L.; Vandecaetsbeek, I.; Wuytack, F.; Vangheluwe, P. Modeling Ca2+ Dynamics of Mouse Cardiac Cells Points to a Critical Role of SERCA’s Affinity for Ca2+. Biophys. J. 2011, 100, 1216–1225. [Google Scholar] [CrossRef] [Green Version]
- Juska, A. Dynamics of Calcium Fluxes in Nonexcitable Cells: Mathematical Modeling. J. Membr. Biol. 2006, 211, 89–99. [Google Scholar] [CrossRef]
- Camello, C.; Lomax, R.; Petersen, O.; Tepikin, A. Calcium leak from intracellular stores—The enigma of calcium signalling. Cell Calcium 2002, 32, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Lomax, R.B.; Camello, C.; Van Coppenolle, F.; Petersen, O.H.; Tepikin, A. Basal and physiological Ca2+ leak from the endoplasmic reticulum of pancreatic acinar cells. Second messenger-activated channels and translocons. J. Biol. Chem. 2002, 277, 26479–26485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moenke, G.; Falcke, M.; Thurley, K. Hierarchic Stochastic Modelling Applied to Intracellular Ca2+ Signals. PLoS ONE 2012, 7, e51178. [Google Scholar] [CrossRef]
- Benham, A. Protein Secretion and the Endoplasmic Reticulum. Cold Spring Harb. Perspect. Biol. 2012, 4, a012872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruggiano, A.; Foresti, O.; Carvalho, P. Quality control: ER-associated degradation: Protein quality control and beyond. J. Cell Biol. 2014, 204, 869–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schönthal, A.H. Endoplasmic Reticulum Stress: Its Role in Disease and Novel Prospects for Therapy. Science 2012, 2012, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Kepp, O.; Menger, L.; Vacchelli, E.; Locher, C.; Adjemian, S.; Yamazaki, T.; Martins, I.; Sukkurwala, A.Q.; Michaud, M.; Senovilla, L.; et al. Crosstalk between ER stress and immunogenic cell death. Cytokine Growth Factor Rev. 2013, 24, 311–318. [Google Scholar] [CrossRef]
- Groenendyk, J.; Michalak, M. Endoplasmic reticulum quality control and apoptosis. Acta Biochim. Pol. 2005, 52, 381–395. [Google Scholar] [CrossRef]
- Mandl, J.; Mészáros, T.; Bánhegyi, G.; Csala, M. Minireview: Endoplasmic Reticulum Stress: Control in Protein, Lipid, and Signal Homeostasis. Mol. Endocrinol. 2013, 27, 384–393. [Google Scholar] [CrossRef] [Green Version]
- Son, T.G.; Kim, S.J.; Kim, K.; Kim, M.-S.; Chung, H.Y.; Lee, J. Cytoprotective roles of senescence marker protein 30 against intracellular calcium elevation and oxidative stress. Arch. Pharmacal Res. 2008, 31, 872–877. [Google Scholar] [CrossRef]
- Niu, H.; Qian, L.; Sun, B.; Liu, W.; Wang, F.; Wang, Q.; Ji, X.; Luo, Y.; Nesa, E.U.; Lou, H.; et al. Inactivation of TFEB and NF-κB by marchantin M alleviates the chemotherapy-driven pro-tumorigenic senescent secretion. Acta Pharm. Sin. B 2019, 9, 923–936. [Google Scholar] [CrossRef] [PubMed]
- Ravier, M.; Daro, D.; Roma, L.P.; Jonas, J.-C.; Cheng-Xue, R.; Schuit, F.C.; Gilon, P. Mechanisms of control of the free Ca2+ concentration in the endoplasmic reticulum of mouse pancreatic beta-cells: Interplay with cell metabolism and [Ca2+]c and role of SERCA2b and SERCA3. Diabetes 2011, 60, 2533–2545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thurley, K.; Tovey, S.C.; Moenke, G.; Prince, V.L.; Meena, A.; Thomas, A.P.; Skupin, A.; Taylor, C.; Falcke, M. Reliable Encoding of Stimulus Intensities Within Random Sequences of Intracellular Ca2+ Spikes. Sci. Signal. 2014, 7, ra59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visegrády, A.; Lakos, Z.; Czimbalek, L.; Somogyi, B. Stimulus-dependent control of inositol 1,4,5-trisphosphate-induced Ca(2+) oscillation frequency by the endoplasmic reticulum Ca2+-ATPase. Biophys. J. 2001, 81, 1398–1405. [Google Scholar] [CrossRef] [Green Version]
- Gilon, P.; Arredouani, A.; Gailly, P.; Gromada, J.; Henquin, J.-C. Uptake and Release of Ca2+ by the Endoplasmic Reticulum Contribute to the Oscillations of the Cytosolic Ca2+ Concentration Triggered by Ca2+ Influx in the Electrically Excitable Pancreatic B-cell. J. Biol. Chem. 1999, 274, 20197–20205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerrero-Hernández, A.; Sánchez-Vázquez, V.H.; Martínez-Martínez, E.; Sandoval-Vázquez, L.; Perez-Rosas, N.C.; Lopez-Farias, R.; Dagnino-Acosta, A. Sarco-Endoplasmic Reticulum Calcium Release Model Based on Changes in the Luminal Calcium Content. Adv. Exp. Med. Biol. 2020, 1131, 337–370. [Google Scholar] [CrossRef]
- Quintana, A.; Schwarz, E.C.; Hoth, M. Calcium-dependent activation of T-lymphocytes. Pflüger Archiv Die Gesammte Physiologie des Menschen Thiere 2004, 450, 1–12. [Google Scholar] [CrossRef]
- Smedler, E.; Uhlén, P. Frequency decoding of calcium oscillations. Biochim. Biophys. Acta (BBA) Gen. Subj. 2014, 1840, 964–969. [Google Scholar] [CrossRef] [Green Version]
- Mohis, M.; Edwards, S.; Ryan, S.; Rizvi, F.; Tajik, A.J.; Jahangir, A.; Ross, G.R. Aging-related increase in store-operated Ca2+ influx in human ventricular fibroblasts. Am. J. Physiol. Circ. Physiol. 2018, 315, H83–H91. [Google Scholar] [CrossRef] [Green Version]
- De Loof, A. Calcitox-aging counterbalanced by endogenous farnesol-like sesquiterpenoids: An undervalued evolutionarily ancient key signaling pathway. Commun. Integr. Biol. 2017, 10, e1341024. [Google Scholar] [CrossRef] [Green Version]
- Thastrup, O.; Cullen, P.J.; Drobak, B.K.; Hanley, M.R.; Dawson, A.P. Thapsigargin, a tumor promoter, discharges intracellular Ca2+ stores by specific inhibition of the endoplasmic reticulum Ca2+-ATPase. Proc. Natl. Acad. Sci. USA 1990, 87, 2466–2470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seidler, N.W.; Jona, I.; Vegh, M.; Martonosi, A. Cyclopiazonic acid is a specific inhibitor of the Ca2+-ATPase of sarcoplasmic reticulum. J. Biol. Chem. 1989, 264, 17816–17823. [Google Scholar]
- Paula, S.; Abell, J.; Deye, J.; Elam, C.; Lape, M.; Purnell, J.; Ratliff, R.; Sebastian, K.; Zultowsky, J.; Kempton, R.J. Design, synthesis, and biological evaluation of hydroquinone derivatives as novel inhibitors of the sarco/endoplasmic reticulum calcium ATPase. Bioorg. Med. Chem. 2009, 17, 6613–6619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindner, P.; Christensen, S.B.; Nissen, P.; Møller, J.V.; Engedal, N. Cell death induced by the ER stressor thapsigargin involves death receptor 5, a non-autophagic function of MAP1LC3B, and distinct contributions from unfolded protein response components. Cell Commun. Signal. 2020, 18, 12–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, S.-H.; Zhang, W.; McGrath, B.C.; Zhang, P.; Cavener, U.R. PERK (eIF2α kinase) is required to activate the stress-activated MAPKs and induce the expression of immediate-early genes upon disruption of ER calcium homoeostasis. Biochem. J. 2005, 393, 201–209. [Google Scholar] [CrossRef] [Green Version]
- Pinton, P.; Giorgi, C.; Siviero, R.; Zecchini, E.; Rizzuto, R. Calcium and apoptosis: ER-mitochondria Ca2+ transfer in the control of apoptosis. Oncogene 2008, 27, 6407–6418. [Google Scholar] [CrossRef] [Green Version]
- Szabadkai, G.; Rizzuto, R. Chaperones as Parts of Organelle Networks. Plant Promot. Transcr. Factors 2007, 594, 64–77. [Google Scholar] [CrossRef]
- Chemaly, E.R.; Troncone, L.; Lebeche, D. SERCA control of cell death and survival. Cell Calcium 2017, 69, 46–61. [Google Scholar] [CrossRef]
- Giorgi, C.; Bonora, M.; Sorrentino, G.; Missiroli, S.; Poletti, F.; Suski, J.M.; Ramirez, F.G.; Rizzuto, R.; Di Virgilio, F.; Zito, E.; et al. p53 at the endoplasmic reticulum regulates apoptosis in a Ca2+-dependent manner. Proc. Natl. Acad. Sci. USA 2015, 112, 1779–1784. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.G.; Zhang, H. The ER-localized autophagy protein EPG-3/VMP1 regulates ER contacts with other organelles by modulating ATP2A/SERCA activity. Autophagy 2018, 14, 362–363. [Google Scholar] [CrossRef]
- Short, A.D.; Bian, J.; Ghosh, T.K.; Waldron, R.T.; Rybak, S.L.; Gill, D.L. Intracellular Ca2+ pool content is linked to control of cell growth. Proc. Natl. Acad. Sci. USA 1993, 90, 4986–4990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergner, A.; Kellner, J.; Tufman, A.; Huber, R.M. Endoplasmic reticulum Ca2+-homeostasis is altered in small and non-small cell lung cancer cell lines. J. Exp. Clin. Cancer Res. 2009, 28, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Launay, S.; Giannı, M.; Diomede, L.; Machesky, L.M.; Enouf, J.; Papp, B.; Giannì, M. Enhancement of ATRA-induced cell differentiation by inhibition of calcium accumulation into the endoplasmic reticulum: Cross-talk between RARα and calcium-dependent signaling. Blood 2003, 101, 3220–3228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apáti, Á.; Jánossy, J.; Brózik, A.; Bauer, P.I.; Magócsi, M. Calcium Induces Cell Survival and Proliferation through the Activation of the MAPK Pathway in a Human Hormone-dependent Leukemia Cell Line, TF-1. J. Biol. Chem. 2003, 278, 9235–9243. [Google Scholar] [CrossRef] [Green Version]
- Suisse, A.; Treisman, J. Reduced SERCA Function Preferentially Affects Wnt Signaling by Retaining E-Cadherin in the Endoplasmic Reticulum. Cell Rep. 2019, 26, 322–329. [Google Scholar] [CrossRef] [Green Version]
- Roti, G.; Carlton, A.; Ross, K.N.; Markstein, M.; Pajcini, K.; Su, A.H.; Perrimon, N.; Pear, W.S.; Kung, A.; Blacklow, S.C.; et al. Complementary genomic screens identify SERCA as a therapeutic target in NOTCH1 mutated cancer. Cancer Cell 2013, 23, 390–405. [Google Scholar] [CrossRef] [Green Version]
- Periz, G.; Fortini, M.E. Ca2+-ATPase function is required for intracellular trafficking of the Notch receptor in Drosophila. EMBO J. 1999, 18, 5983–5993. [Google Scholar] [CrossRef] [Green Version]
- De Ford, C.; Heidersdorf, B.; Haun, F.; Murillo, R.; Friedrich, T.; Borner, C.; Merfort, I. The clerodane diterpene casearin J induces apoptosis of T-ALL cells through SERCA inhibition, oxidative stress, and interference with Notch1 signaling. Cell Death Dis. 2016, 7, e2070. [Google Scholar] [CrossRef]
- Bittremieux, M.; Parys, J.B.; Pinton, P.; Bultynck, G. ER functions of oncogenes and tumor suppressors: Modulators of intracellular Ca2+ signaling. Biochim. Biophys. Acta (BBA) Bioenerg. 2016, 1863, 1364–1378. [Google Scholar] [CrossRef]
- Liu, J.; Huang, K.; Cai, G.-Y.; Chen, X.; Yang, J.-R.; Lin, L.-R.; Yang, J.; Huo, B.-G.; Zhan, J.; He, Y.-N. Receptor for advanced glycation end-products promotes premature senescence of proximal tubular epithelial cells via activation of endoplasmic reticulum stress-dependent p21 signaling. Cell. Signal. 2014, 26, 110–121. [Google Scholar] [CrossRef]
- Hakii, H.; Fujiki, H.; Suganuma, M.; Nakayasu, M.; Tahira, T.; Sugimura, T.; Scheuer, P.J.; Christensen, S.B. Thapsigargin, a histamine secretagogue, is a non-12-O-tetradecanolphorbol-13-acetate (TPA) type tumor promoter in two-stage mouse skin carcinogenesis. J. Cancer Res. Clin. Oncol. 1986, 111, 177–181. [Google Scholar] [CrossRef]
- Hoji, A.; Xu, S.; Bilben, H.; Rowe, D.T. Calcium mobilization is responsible for Thapsigargin induced Epstein Barr virus lytic reactivation in in vitro immortalized lymphoblstoid cell lines. Heliyon 2018, 4, e00917. [Google Scholar] [CrossRef] [PubMed]
- Papp, B.; Byrn, R.A. Stimulation of HIV Expression by Intracellular Calcium Pump Inhibition. J. Biol. Chem. 1995, 270, 10278–10283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahalingam, D.; Peguero, J.; Cen, P.; Arora, S.P.; Sarantopoulos, J.; Rowe, J.H.; Allgood, V.; Tubb, B.; Campos, L. A Phase II, Multicenter, Single-Arm Study of Mipsagargin (G-202) as a Second-Line Therapy Following Sorafenib for Adult Patients with Progressive Advanced Hepatocellular Carcinoma. Cancers 2019, 11, 833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahalingam, D.; Wilding, G.; Denmeade, S.; Sarantopoulas, J.; Cosgrove, D.; Cetnar, J.; Azad, N.; Bruce, J.; Kurman, M.; Allgood, V.; et al. Mipsagargin, a novel thapsigargin-based PSMA-activated prodrug: Results of a first-in-man phase I clinical trial in patients with refractory, advanced or metastatic solid tumours. Br. J. Cancer 2016, 114, 986–994. [Google Scholar] [CrossRef]
- Doan, N.T.Q.; Paulsen, E.S.; Sehgal, P.; Møller, J.V.; Nissen, P.; Denmeade, S.R.; Isaacs, J.T.; Dionne, C.A.; Christensen, S.B. Targeting thapsigargin towards tumors. Steroids 2014, 97, 2–7. [Google Scholar] [CrossRef]
- Dubois, C.; Vanden Abeele, F.; Sehgal, P.; Olesen, C.; Junker, S.; Christensen, S.B.; Prevarskaya, N.; Møller, J.V. Differential effects of thapsigargin analogues on apoptosis of prostate cancer cells: complex regulation by intracellular calcium. FEBS J. 2013, 280, 5430–5440. [Google Scholar] [CrossRef]
- White, M.; Johnson, G.; Zhang, W.; Hobrath, J.; Piazza, G.A.; Grimaldi, M. Sulindac sulfide inhibits sarcoendoplasmic reticulum Ca2+ATPase, induces endoplasmic reticulum stress response, and exerts toxicity in glioma cells: Relevant similarities to and important differences from celecoxib. J. Neurosci. Res. 2012, 91, 393–406. [Google Scholar] [CrossRef] [Green Version]
- Fan, L.; Li, A.; Li, W.; Cai, P.; Yang, B.; Zhang, M.; Gu, Y.; Shu, Y.; Sun, Y.; Shen, Y.; et al. Novel role of Sarco/endoplasmic reticulum calcium ATPase 2 in development of colorectal cancer and its regulation by F36, a curcumin analog. Biomed. Pharmacother. 2014, 68, 1141–1148. [Google Scholar] [CrossRef]
- Peterson, J.A.; Crowther, C.M.; Andrus, M.B.; Kenealey, J. Resveratrol derivatives increase cytosolic calcium by inhibiting plasma membrane ATPase and inducing calcium release from the endoplasmic reticulum in prostate cancer cells. Biochem. Biophys. Rep. 2019, 19, 100667. [Google Scholar] [CrossRef]
- Tadini-Buoninsegni, F.; Smeazzetto, S.; Gualdani, R.; Moncelli, M.R. Drug Interactions With the Ca2+-ATPase From Sarco(Endo)Plasmic Reticulum (SERCA). Front. Mol. Biosci. 2018, 5, 36. [Google Scholar] [CrossRef] [PubMed]
- Ji, S.; Lee, J.-Y.; Schrör, J.; Mazumder, A.; Jang, D.M.; Chateauvieux, S.; Schnekenburger, M.; Hong, C.R.; Christov, C.; Kang, H.J.; et al. The dialkyl resorcinol stemphol disrupts calcium homeostasis to trigger programmed immunogenic necrosis in cancer. Cancer Lett. 2018, 416, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Plenge, L.; Soler, F.; Fernández-Belda, F. Ritodrine Inhibition of the Plasma Membrane Ca2+-ATPase from Human Erythrocyte. Arch. Biochem. Biophys. 1998, 357, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Yiallouris, A.; Patrikios, I.; Johnson, E.O.; Sereti, E.; Dimas, K.; De Ford, C.; Fedosova, N.U.; Graier, W.F.; Sokratous, K.; Kyriakou, K.; et al. Annonacin promotes selective cancer cell death via NKA-dependent and SERCA-dependent pathways. Cell Death Dis. 2018, 9, 764. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Wang, Y.; Yu, Y.; Zheng, J.; Chen, J.; Li, S.; Chen, R.; Zhang, C.; Naman, C.B.; Yu, D.; et al. Schekwanglupaside C, a new lupane saponin from Schefflera kwangsiensis, is a potent activator of sarcoplasmic reticulum Ca2+-ATPase. Fitoterapia 2019, 137, 104150. [Google Scholar] [CrossRef] [PubMed]
- Rahate, K.; Bhatt, L.K.; Prabhavalkar, K.S. SERCA stimulation: A potential approach in therapeutics. Chem. Biol. Drug Des. 2019, 95, 5–15. [Google Scholar] [CrossRef]
- Kang, S.; Dahl, R.; Hsieh, W.; Shin, A.; Zsebo, K.M.; Buettner, C.; Hajjar, R.J.; Lebeche, D. Small Molecular Allosteric Activator of the Sarco/Endoplasmic Reticulum Ca2+-ATPase (SERCA) Attenuates Diabetes and Metabolic Disorders. J. Biol. Chem. 2015, 291, 5185–5198. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, R.; Schraml, E.; Leitinger, G.; Letofsky-Papst, I.; Stelzer, I.; Haas, H.S.; Schauenstein, K.; Sadjak, A. α1-adrenergic drugs exhibit affinity to a thapsigargin-sensitive binding site and interfere with the intracellular Ca2+ homeostasis in human erythroleukemia cells. Exp. Cell Res. 2011, 317, 2969–2980. [Google Scholar] [CrossRef]
- Yaguchi, T.; Nagata, T.; Nishizaki, T. 1,2-Dilinoleoyl-sn-glycero-3-phosphoethanolamine ameliorates age-related spatial memory deterioration by preventing neuronal cell death. Behav. Brain Funct. 2010, 6, 52. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, T.; Igarashi, H.; Kawagoe, J.; Amita, M.; Hara, S.; Kurachi, H. Poor Embryo Development in Mouse Oocytes Aged In Vitro Is Associated with Impaired Calcium Homeostasis. Biol. Reprod. 2009, 80, 493–502. [Google Scholar] [CrossRef] [Green Version]
- Ryu, S.J.; An, H.J.; Oh, Y.S.; Choi, H.R.; Ha, M.K.; Park, S.C. On the role of major vault protein in the resistance of senescent human diploid fibroblasts to apoptosis. Cell Death Differ. 2008, 15, 1673–1680. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Adhikary, A.; Chakraborty, S.; Nandi, P.; Mohanty, S.; Chakraborty, S.; Bhattacharjee, P.; Mukherjee, S.; Putatunda, S.; Chakraborty, S.; et al. Nifetepimine, a Dihydropyrimidone, Ensures CD4+ T Cell Survival in a Tumor Microenvironment by Maneuvering Sarco(endo)plasmic Reticulum Ca2+ ATPase (SERCA). J. Biol. Chem. 2012, 287, 32881–32896. [Google Scholar] [CrossRef] [Green Version]
- Bleeker, N.P.; Cornea, R.L.; Thomas, D.D.T.; Xing, C. A Novel SERCA Inhibitor Demonstrates Synergy with Classic SERCA Inhibitors and Targets Multidrug-Resistant AML. Mol. Pharm. 2013, 10, 4358–4366. [Google Scholar] [CrossRef] [Green Version]
- Puzianowska-Kuźnicka, M.; Kuznicki, J. The ER and ageing II: Calcium homeostasis. Ageing Res. Rev. 2009, 8, 160–172. [Google Scholar] [CrossRef]
- Sadafi, F.-Z.; Massai, L.; Bartolommei, G.; Moncelli, M.R.; Messori, L.; Tadini-Buoninsegni, F. Anticancer Ruthenium(III) Complex KP1019 Interferes with ATP-Dependent Ca2+ Translocation by Sarco-Endoplasmic Reticulum Ca2+-ATPase (SERCA). ChemMedChem 2014, 9, 1660–1664. [Google Scholar] [CrossRef]
- Guven, A.; Lin, W.-Y.; Leggett, R.E.; Kogan, B.A.; Levin, R.M.; Mannikarottu, A. Effect of aging on the response of biochemical markers in the rabbit subjected to short-term partial bladder obstruction. Mol. Cell. Biochem. 2007, 306, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Arvanitis, D.; Vafiadaki, E.; Fan, G.-C.; Mitton, B.A.; Gregory, K.N.; Del Monte, F.; Kontrogianni-Konstantopoulos, A.; Sanoudou, D.; Kranias, E.G. Histidine-rich Ca-binding protein interacts with sarcoplasmic reticulum Ca-ATPase. Am. J. Physiol. Circ. Physiol. 2007, 293, H1581–H1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vangheluwe, P.; Tjwa, M.; Bergh, A.V.D.; Louch, W.E.; Beullens, M.; Dode, L.; Carmeliet, P.; Kranias, E.; Herijgers, P.; Sipido, K.R.; et al. A SERCA2 pump with an increased Ca2+ affinity can lead to severe cardiac hypertrophy, stress intolerance and reduced life span. J. Mol. Cell. Cardiol. 2006, 41, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Ying, J.; Jiang, B.; Guo, W.; Adachi, T.; Sharov, V.; Lazar, H.; Menzoian, J.; Knyushko, T.V.; Bigelow, D.; et al. Detection of sequence-specific tyrosine nitration of manganese SOD and SERCA in cardiovascular disease and aging. Am. J. Physiol. Circ. Physiol. 2006, 290, H2220–H2227. [Google Scholar] [CrossRef] [PubMed]
- Ginzburg, L.; Futerman, A.H. Defective calcium homeostasis in the cerebellum in a mouse model of Niemann-Pick A disease. J. Neurochem. 2005, 95, 1619–1628. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Joyner, R.W. Expression of protein phosphatases during postnatal development of rabbit heart. Mol. Cell. Biochem. 2003, 245, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.G.; Zhai, J.; Carr, A.N.; Gerst, M.J.; Lorenz, J.N.; Pollesello, P.; Annila, A.; Hoit, B.D.; Kranias, E.G. Structural and functional implications of the phospholamban hinge domain: Impaired SR Ca2+ uptake as a primary cause of heart failure. Cardiovasc. Res. 2002, 56, 248–259. [Google Scholar] [CrossRef] [Green Version]
- Squier, T.C. Oxidative stress and protein aggregation during biological aging. Exp. Gerontol. 2001, 36, 1539–1550. [Google Scholar] [CrossRef]
- Pottorf, W.J.; Duckles, S.P.; Buchholz, J. Adrenergic nerves compensate for a decline in calcium buffering during ageing. J. Auton. Pharmacol. 2000, 20, 1–13. [Google Scholar] [CrossRef]
- Bublitz, M.; Kjellerup, L.; Cohrt, K.O.; Gordon, S.; Mortensen, A.L.; Clausen, J.D.; Pallin, T.D.; Hansen, J.B.; Fuglsang, A.T.; Dalby-Brown, W.; et al. Tetrahydrocarbazoles are a novel class of potent P-type ATPase inhibitors with antifungal activity. PLoS ONE 2018, 13, e0188620. [Google Scholar] [CrossRef] [Green Version]
- Johnson, A.J.; Hsu, A.-L.; Lin, H.-P.; Song, X.; Chen, C.-S. The cyclo-oxygenase-2 inhibitor celecoxib perturbs intracellular calcium by inhibiting endoplasmic reticulum Ca2+-ATPases: A plausible link with its anti-tumour effect and cardiovascular risks. Biochem. J. 2002, 366, 831–837. [Google Scholar] [CrossRef] [Green Version]
- Lax, A.; Soler, F.; Fernandez-Belda, F. Inhibition of sarcoplasmic reticulum Ca2+-ATPase by miconazole. Am. J. Physiol. Physiol. 2002, 283, C85–C92. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Zhang, M.; Gao, J.; Li, J.; Fan, L.; Xiang, G.; Wang, X.; Wang, X.; Wu, X.; Sun, Y.; et al. Small molecule RL71 targets SERCA2 at a novel site in the treatment of human colorectal cancer. Oncotarget 2015, 6, 37613–37625. [Google Scholar] [CrossRef] [Green Version]
- Ogunbayo, O.A.; Lai, P.; Connolly, T.J.; Michelangeli, F. Tetrabromobisphenol A (TBBPA), induces cell death in TM4 Sertoli cells by modulating Ca2+ transport proteins and causing dysregulation of Ca2+ homeostasis. Toxicol. Vitr. Int. J. Publ. Assoc. BIBRA 2008, 22, 943–952. [Google Scholar] [CrossRef]
- Fan, X.-X.; Yao, X.-J.; Xu, S.W.; Wong, V.K.W.; He, J.-X.; Ding, J.; Xue, W.W.; Mujtaba, T.; Michelangeli, F.; Huang, M.; et al. (Z)3,4,5,4′-trans-tetramethoxystilbene, a new analogue of resveratrol, inhibits gefitinb-resistant non-small cell lung cancer via selectively elevating intracellular calcium level. Sci. Rep. 2015, 5, 16348. [Google Scholar] [CrossRef] [Green Version]
- Muscella, A.; Calabriso, N.; Vetrugno, C.; Fanizzi, F.P.; De Pascali, S.A.; Storelli, C.; Marsigliante, S. The platinum (II) complex [Pt(O,O′-acac)(γ-acac)(DMS)] alters the intracellular calcium homeostasis in MCF-7 breast cancer cells. Biochem. Pharmacol. 2011, 81, 91–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wuytack, F.; Raeymaekers, L.; Smedt, H.; Eggermont, J.A.; Missiaen, L.; Bosch, L.V.D.; Jaegere, S.; Verboomen, H.; Plessers, L.; Casteels, R. Ca2+-Transport ATPases and Their Regulation in Muscle and Brain. Ann. N. Y. Acad. Sci. 1992, 671, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Papp, B.; Enyedi, A.; Kovács, T.; Sarkadi, B.; Wuytack, F.; Thastrup, O.; Gárdos, G.; Bredoux, R.; Levy-Toledano, S.; Enouf, J. Demonstration of two forms of calcium pumps by thapsigargin inhibition and radioimmunoblotting in platelet membrane vesicles. J. Biol. Chem. 1991, 266, 14593–14596. [Google Scholar]
- Papp, B.; Enyedi, A.; Pászty, K.; Kovacs, T.; Sarkadi, B.; Gárdos, G.; Magnier, C.; Wuytack, F.; Enouf, J. Simultaneous presence of two distinct endoplasmic-reticulum-type calcium-pump isoforms in human cells. Characterization by radio-immunoblotting and inhibition by 2,5-di-(t-butyl)-1,4-benzohydroquinone. Biochem. J. 1992, 288, 297–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gélébart, P.; Kovács, T.; Brouland, J.-P.; Van Gorp, R.; Grossmann, J.; Rivard, N.; Panis, Y.; Martin, V.; Bredoux, R.; Enouf, J.; et al. Expression of endomembrane calcium pumps in colon and gastric cancer cells. Induction of SERCA3 expression during differentiation. J. Biol. Chem. 2002, 277, 26310–26320. [Google Scholar] [CrossRef] [Green Version]
- Ho, P.; Bihuniak, J.D.; MacIntyre, A.; Staron, M.; Liu, X.; Amezquita, R.; Tsui, Y.-C.; Cui, G.; Micevic, G.; Perales, J.C.; et al. Phosphoenolpyruvate Is a Metabolic Checkpoint of Anti-tumor T Cell Responses. Cell 2015, 162, 1217–1228. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Felici, J.; Hyroššová, P.; Aragó, M.; Rodríguez-Arévalo, S.; García-Rovés, P.M.; Escolano, C.; Perales, J.C. Phosphoenolpyruvate from Glycolysis and PEPCK Regulate Cancer Cell Fate by Altering Cytosolic Ca2+. Cells 2019, 9, 18. [Google Scholar] [CrossRef] [Green Version]
- Anderson, D.M.; Makarewich, C.A.; Shelton, J.M.; Bezprozvannaya, S.; Bassel-Duby, R.; Olson, E.; Anderson, U.M.; Anderson, K.M. Widespread control of calcium signaling by a family of SERCA-inhibiting micropeptides. Sci. Signal. 2016, 9, ra119. [Google Scholar] [CrossRef] [Green Version]
- Jäger, S.; Cimermancic, P.; Gulbahce, N.; Johnson, J.R.; McGovern, K.E.; Clarke, S.C.; Shales, M.; Mercenne, G.; Pache, L.; Li, K.; et al. Global landscape of HIV–human protein complexes. Nature 2011, 481, 365–370. [Google Scholar] [CrossRef]
- Lai, S.; Li, Y.; Kuang, Y.; Cui, H.; Yang, Y.; Sun, W.; Liu, K.; Chen, N.; Yan, Q.; Wen, L. PKCδ silencing alleviates saturated fatty acid induced ER stress by enhancing SERCA activity. Biosci. Rep. 2017, 37, 37. [Google Scholar] [CrossRef] [Green Version]
- Zarain-Herzberg, A.; García-Rivas, G.; Estrada-Avilés, R. Regulation of SERCA pumps expression in diabetes. Cell Calcium 2014, 56, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Chaabane, C.; Dally, S.; Corvazier, E.; Bredoux, R.; Bobe, R.; Ftouhi, B.; Raies, A.; Enouf, J. Platelet PMCA- and SERCA-type Ca2+-ATPase expression in diabetes: A novel signature of abnormal megakaryocytopoiesis. J. Thromb. Haemost. 2007, 5, 2127–2135. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Fonseca, K.; Massieu, L. Disruption of endoplasmic reticulum calcium stores is involved in neuronal death induced by glycolysis inhibition in cultured hippocampal neurons. J. Neurosci. Res. 2005, 82, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Martínez, M.C.; Martin, S.; Toti, F.; Fressinaud, E.; Dachary-Prigent, J.; Meyer, M.; Freyssinet, J.-M. Significance of Capacitative Ca2+ Entry in the Regulation of Phosphatidylserine Expression at the Surface of Stimulated Cells. Biochemistry 1999, 38, 10092–10098. [Google Scholar] [CrossRef] [PubMed]
- Bittremieux, M.; Bultynck, G. p53 and Ca2+ signaling from the endoplasmic reticulum: Partners in anti-cancer therapies. Oncoscience 2015, 2, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Betzer, C.; Jensen, P.H. Reduced Cytosolic Calcium as an Early Decisive Cellular State in Parkinson’s Disease and Synucleinopathies. Front. Mol. Neurosci. 2018, 12, 819. [Google Scholar] [CrossRef]
- Yoboue, E.D.; Sitia, R.; Simmen, T. Redox crosstalk at endoplasmic reticulum (ER) membrane contact sites (MCS) uses toxic waste to deliver messages. Cell Death Dis. 2018, 9, 331. [Google Scholar] [CrossRef] [Green Version]
- Ghezali, L.A.; Arbabian, A.; Roudot, H.; Brouland, J.-P.; Baran-Marszak, F.; Salvaris, E.; Boyd, A.; Drexler, H.G.; Enyedi, Á.; Letestu, R.; et al. Induction of endoplasmic reticulum calcium pump expression during early leukemic B cell differentiation. J. Exp. Clin. Cancer Res. 2017, 36, 87. [Google Scholar] [CrossRef]
- Arbabian, A.; Brouland, J.-P.; Apáti, A.; Pászty, K.; Hegedüs, L.; Enyedi, Á.; Chomienne, C.; Papp, B. Modulation of endoplasmic reticulum calcium pump expression during lung cancer cell differentiation. FEBS J. 2012, 280, 5408–5418. [Google Scholar] [CrossRef] [Green Version]
- Bobe, R.; Bredoux, R.; Wuytack, F.; Quarck, R.; Kovàcs, T.; Papp, B.; Corvazier, E.; Magnier, C.; Enouf, J. The rat platelet 97-kDa Ca2+ ATPase isoform is the sarcoendoplasmic reticulum Ca2+ATPase 3 protein. J. Biol. Chem. 1994, 269, 1417–1424. [Google Scholar]
- Brouland, J.-P.; Gélébart, P.; Kovács, T.; Enouf, J.; Grossmann, J.; Papp, B. The Loss of Sarco/Endoplasmic Reticulum Calcium Transport ATPase 3 Expression Is an Early Event during the Multistep Process of Colon Carcinogenesis. Am. J. Pathol. 2005, 167, 233–242. [Google Scholar] [CrossRef] [Green Version]
- Dellis, O.; Arbabian, A.; Brouland, J.-P.; Kovács, T.; Rowe, M.; Chomienne, C.; Joab, I.; Papp, B. Modulation of B-cell endoplasmic reticulum calcium homeostasis by Epstein-Barr virus Latent Membrane Protein-1. Mol. Cancer 2009, 8, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enouf, J.; Bobe, R.; Lacabaratz, C.; Bredoux, R.; Corvazier, E.; Kovacs, T.; Papp, B.; Enouf, R.B.J. The platelet Ca2+ transport ATPase system. Platelets 1997, 8, 5–14. [Google Scholar] [CrossRef]
- Gélébart, P.; Martin, V.; Enouf, J.; Papp, B. Identification of a new SERCA2 splice variant regulated during monocytic differentiation. Biochem. Biophys. Res. Commun. 2003, 303, 676–684. [Google Scholar] [CrossRef]
- Aït-Ghezali, L.; Arbabian, A.; Jeibmann, A.; Hasselblatt, M.; Hallaert, G.G.; Broecke, C.V.D.; Gray, F.; Brouland, J.-P.; Varin-Blank, N.; Papp, B. Loss of endoplasmic reticulum calcium pump expression in choroid plexus tumours. Neuropathol. Appl. Neurobiol. 2014, 40, 726–735. [Google Scholar] [CrossRef] [PubMed]
- Launay, S.; Bobe, R.; Lacabaratz, C.; Bredoux, R.; Kovács, T.; Enouf, J.; Papp, B. Modulation of Endoplasmic Reticulum Calcium Pump Expression during T Lymphocyte Activation. J. Biol. Chem. 1997, 272, 10746–10750. [Google Scholar] [CrossRef] [Green Version]
- Launay, S.; Giannì, M.; Kovács, T.; Bredoux, R.; Bruel, A.; Gélébart, P.; Zassadowski, F.; Chomienne, C.; Enouf, J.; Papp, B. Lineage-specific modulation of calcium pump expression during myeloid differentiation. Blood 1999, 93, 4395–4405. [Google Scholar] [CrossRef]
- Martin, V.; Bredoux, R.; Corvazier, E.; Papp, B.; Enouf, J. Platelet Ca2+ ATPases: A plural, species-specific, and multiple hypertension-regulated expression system. Hypertension 2000, 35, 91–102. [Google Scholar] [CrossRef] [Green Version]
- Papp, B.; Brouland, J.-P. Altered Endoplasmic Reticulum Calcium Pump Expression during Breast Tumorigenesis. Breast Cancer Basic Clin. Res. 2011, 5, 163–174. [Google Scholar] [CrossRef]
- Papp, B.; Brouland, J.-P.; Gélébart, P.; Kovács, T.; Chomienne, C. Endoplasmic reticulum calcium transport ATPase expression during differentiation of colon cancer and leukaemia cells. Biochem. Biophys. Res. Commun. 2004, 322, 1223–1236. [Google Scholar] [CrossRef]
- Varadi, A.; Molnár, E.; Östenson, C.-G.; Ashcroft, S.J.H. Isoforms of endoplasmic reticulum Ca2+-ATPase are differentially expressed in normal and diabetic islets of Langerhans. Biochem. J. 1996, 319, 521–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magnier-Gaubil, C.; Herbert, J.M.; Quarck, R.; Papp, B.; Corvazier, E.; Wuytack, F.; Lévy-Tolédano, S.; Enouf, J. Smooth muscle cell cycle and proliferation. Relationship between calcium influx and sarco-endoplasmic reticulum Ca2+ ATPase regulation. J. Biol. Chem. 1996, 271, 27788–27794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, S.; Laude-Lemaire, I.; Kerbiriou-Nabias, D.; Freyssinet, J.-M.; Martinez, M. Relation between Phosphatidylserine Exposure and Store-Operated Ca2+ Entry in Stimulated Cells. Biochem. Biophys. Res. Commun. 2000, 279, 639–645. [Google Scholar] [CrossRef] [PubMed]
- Periasamy, M.; Kalyanasundaram, A. SERCA pump isoforms: Their role in calcium transport and disease. Muscle Nerve 2007, 35, 430–442. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.D.; Lytton, J. Molecular cloning and quantification of sarcoplasmic reticulum Ca2+-ATPase isoforms in rat muscles. Am. J. Physiol. Physiol. 1993, 264, C333–C341. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.D.; Lee, W.S.; Wey, J.; Bungard, D.; Lytton, J. Localization and quantification of endoplasmic reticulum Ca2+-ATPase isoform transcripts. Am. J. Physiol. Physiol. 1995, 269, C775–C784. [Google Scholar] [CrossRef]
- Poch, E.; Leach, S.; Snape, S.; Cacic, T.; MacLennan, D.H.; Lytton, J. Functional characterization of alternatively spliced human SERCA3 transcripts. Am. J. Physiol. Content 1998, 275, C1449–C1458. [Google Scholar] [CrossRef]
- Martin, V.; Bredoux, R.; Corvazier, E.; Van Gorp, R.; Kovàcs, T.; Gélébart, P.; Enouf, J. Three novel sarco/endoplasmic reticulum Ca2+-ATPase (SERCA) 3 isoforms. Expression, regulation, and function of the membranes of the SERCA3 family. J. Biol. Chem. 2002, 277, 24442–24452. [Google Scholar] [CrossRef] [Green Version]
- Dally, S.; Corvazier, E.; Bredoux, R.; Bobe, R.; Enouf, J. Multiple and diverse coexpression, location, and regulation of additional SERCA2 and SERCA3 isoforms in nonfailing and failing human heart. J. Mol. Cell. Cardiol. 2010, 48, 633–644. [Google Scholar] [CrossRef]
- Wuytack, F.; Dode, L.; Baba-Aissa, F.; Raeymaekers, L. The SERCA3-type of organellar Ca2+ pumps. Biosci. Rep. 1995, 15, 299–306. [Google Scholar] [CrossRef]
- Kovács, T.; Felföldi, F.; Papp, B.; Pászty, K.; Bredoux, R.; Enyedi, Á.; Enouf, J. All three splice variants of the human sarco/endoplasmic reticulum Ca2+-ATPase 3 gene are translated to proteins: A study of their co-expression in platelets and lymphoid cells. Biochem. J. 2001, 358, 559. [Google Scholar] [CrossRef] [PubMed]
- Wuytack, F.; Papp, B.; Verboomen, H.; Raeymaekers, L.; Dode, L.; Bobe, R.; Enouf, J.; Bokkala, S.; Authi, K.S.; Casteels, R. A sarco/endoplasmic reticulum Ca2+-ATPase 3-type Ca2+ pump is expressed in platelets, in lymphoid cells, and in mast cells. J. Biol. Chem. 1994, 269, 1410–1416. [Google Scholar] [PubMed]
- Baba-Aïssa, F.; Raeymaekers, L.; Wuytack, F.; Callewaert, G.; Dode, L.; Missiaen, L.; Casteels, R. Purkinje neurons express the SERCA3 isoform of the organellar type Ca2+-transport ATPase. Mol. Brain Res. 1996, 41, 169–174. [Google Scholar] [CrossRef]
- Anger, M.; Samuel, J.-L.; Marotte, F.; Wuytack, F.; Rappaport, L.; Lompré, A.-M. The sarco(endo)plasmic reticulum Ca2+-ATPase mRNA isoform, SERCA 3, is expressed in endothelial and epithelial cells in various organs. FEBS Lett. 1993, 334, 45–48. [Google Scholar] [CrossRef] [Green Version]
- Lytton, J.; Westlin, M.; Burk, S.E.; Shull, G.E.; MacLennan, D.H. Functional comparisons between isoforms of the sarcoplasmic or endoplasmic reticulum family of calcium pumps. J. Biol. Chem. 1992, 267, 14483–14489. [Google Scholar]
- Chandrasekera, P.C.; Kargacin, M.E.; Deans, J.P.; Lytton, J. Determination of apparent calcium affinity for endogenously expressed human sarco(endo)plasmic reticulum calcium-ATPase isoform SERCA3. Am. J. Physiol. Physiol. 2009, 296, C1105–C1114. [Google Scholar] [CrossRef] [Green Version]
- Dode, L.; Vilsen, B.; Van Baelen, K.; Wuytack, F.; Clausen, J.D.; Andersen, J.P. Dissection of the Functional Differences between Sarco(endo)plasmic Reticulum Ca2+-ATPase (SERCA) 1 and 3 Isoforms by Steady-state and Transient Kinetic Analyses. J. Biol. Chem. 2002, 277, 45579–45591. [Google Scholar] [CrossRef] [Green Version]
- Vandecaetsbeek, I.; Trekels, M.; De Maeyer, M.; Ceulemans, H.; Lescrinier, E.; Raeymaekers, L.; Wuytack, F.; Vangheluwe, P. Structural basis for the high Ca2+ affinity of the ubiquitous SERCA2b Ca2+ pump. Proc. Natl. Acad. Sci. USA 2009, 106, 18533–18538. [Google Scholar] [CrossRef] [Green Version]
- Inesi, G.; Kurzmack, M.; Coan, C.; Lewis, D.E. Cooperative calcium binding and ATPase activation in sarcoplasmic reticulum vesicles. J. Biol. Chem. 1980, 255, 3025–3031. [Google Scholar]
- Kekenes-Huskey, P.; Metzger, V.T.; Grant, B.J.; McCammon, J.A. Calcium binding and allosteric signaling mechanisms for the sarcoplasmic reticulum Ca2+ ATPase. Protein Sci. Publ. Protein Soc. 2012, 21, 1429–1443. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Lewis, D.; Strock, C.; Inesi, G.; Nakasako, M.; Nomura, H.; Toyoshima, C. Detailed Characterization of the Cooperative Mechanism of Ca2+-Binding and Catalytic Activation in the Ca2+ Transport (SERCA) ATPase. Biochemistry 2000, 39, 8758–8767. [Google Scholar] [CrossRef] [PubMed]
- Hovnanian, A. Serca pumps and human diseases. Membr. Biogen. 2007, 45, 337–363. [Google Scholar] [CrossRef]
- Prasad, V.; Boivin, G.P.; Miller, M.L.; Liu, L.H.; Erwin, C.R.; Warner, B.W.; Shull, G.E. Haploinsufficiency of Atp2a2, Encoding the Sarco(endo)plasmic Reticulum Ca2+-ATPase Isoform 2 Ca2+ Pump, Predisposes Mice to Squamous Cell Tumors via a Novel Mode of Cancer Susceptibility. Cancer Res. 2005, 65, 8655–8661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasad, V.; Okunade, G.W.; Miller, M.L.; Shull, G.E. Phenotypes of SERCA and PMCA knockout mice. Biochem. Biophys. Res. Commun. 2004, 322, 1192–1203. [Google Scholar] [CrossRef]
- Arredouani, A.; Guiot, Y.; Jonas, J.-C.; Liu, L.H.; Nenquin, M.; Pertusa, J.A.; Rahier, J.; Rolland, J.-F.; Shull, G.E.; Stevens, M.; et al. SERCA3 ablation does not impair insulin secretion but suggests distinct roles of different sarcoendoplasmic reticulum Ca2+ pumps for Ca2+ homeostasis in pancreatic beta-cells. Diabetes 2002, 51, 3245–3253. [Google Scholar] [CrossRef]
- Shull, G.E.; Okunade, G.; Liu, L.H.; Kozel, P.; Periasamy, M.; Lorenz, J.N.; Prasad, V. Physiological functions of plasma membrane and intracellular Ca2+ pumps revealed by analysis of null mutants. Ann. N. Y. Acad. Sci. 2003, 986, 453–460. [Google Scholar] [CrossRef]
- Zhang, X.; Pathak, T.; Yoast, R.; Emrich, S.; Xin, P.; Nwokonko, R.M.; Johnson, M.; Wu, S.; Delierneux, C.; Gueguinou, M.; et al. A calcium/cAMP signaling loop at the ORAI1 mouth drives channel inactivation to shape NFAT induction. Nat. Commun. 2019, 10, 1971. [Google Scholar] [CrossRef]
- Beauvois, M.C.; Arredouani, A.; Jonas, J.-C.; Rolland, J.-F.; Schuit, F.; Henquin, J.-C.; Gilon, P. Atypical Ca2+-induced Ca2+ release from a sarco-endoplasmic reticulum Ca2+-ATPase 3-dependent Ca2+ pool in mouse pancreatic β-cells. J. Physiol. 2004, 559, 141–156. [Google Scholar] [CrossRef] [Green Version]
- Elaïb, Z.; Adam, F.; Berrou, E.; Bordet, J.-C.; Prévost, N.; Bobe, R.; Bryckaert, M.; Rosa, J.-P. Full activation of mouse platelets requires ADP secretion regulated by SERCA3 ATPase–dependent calcium stores. Blood 2016, 128, 1129–1138. [Google Scholar] [CrossRef] [Green Version]
- Arbabian, A.; Brouland, J.-P.; Gélébart, P.; Kovács, T.; Bobe, R.; Enouf, J.; Papp, B. Endoplasmic reticulum calcium pumps and cancer. BioFactors 2011, 37, 139–149. [Google Scholar] [CrossRef]
- Fenaux, P.; Chomienne, C.; Degos, L. Treatment of acute promyelocytic leukaemia. Best Pract. Res. Clin. Haematol. 2001, 14, 153–174. [Google Scholar] [CrossRef] [PubMed]
- Fenaux, P.; Wang, Z.Z.; Degos, L. Treatment of Acute Promyelocytic Leukemia by Retinoids. Mol. Asp. Myeloid Stem Cell Dev. 2007, 313, 101–128. [Google Scholar] [CrossRef]
- Noguera, N.; Catalano, G.; Banella, C.; Divona, M.; Faraoni, I.; Ottone, T.; Arcese, W.; Voso, M.T. Acute Promyelocytic Leukemia: Update on the Mechanisms of Leukemogenesis, Resistance and on Innovative Treatment Strategies. Cancers 2019, 11, 1591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Thé, H.; Dejean, A. Retinoic acid receptor alpha in acute promyelocytic leukaemia. Cancer Surv. 1992, 14, 195–203. [Google Scholar] [PubMed]
- De Thé, H. Differentiation therapy revisited. Nat. Rev. Cancer 2017, 18, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Lavau, C.P.; The, H.; Dejean, A. Retinoic Acid Receptor? in Acute Promyelocytic Leukemia. Ann. N. Y. Acad. Sci. 1993, 684, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Chapekar, M.S.; Hartman, K.D.; Knode, M.C.; Glazer, R.I. Synergistic effect of retinoic acid and calcium ionophore A23187 on differentiation, c-myc expression, and membrane tyrosine kinase activity in human promyelocytic leukemia cell line HL-60. Mol. Pharmacol. 1987, 31, 140–145. [Google Scholar]
- Lacabaratz-Porret, C.; Launay, S.; Corvazier, E.; Bredoux, R.; Papp, B.; Enouf, J. Biogenesis of endoplasmic reticulum proteins involved in Ca2+ signalling during megakaryocytic differentiation: An in vitro study. Biochem. J. 2000, 350 Pt 3, 723–734. [Google Scholar] [CrossRef]
- Bobe, R.; Bredoux, R.; Corvazier, E.; Lacabaratz, C.; Martin, V.; Kovacs, T.; Enouf, J. How many Ca2+ATPase isoforms are expressed in a cell type? A growing family of membrane proteins illustrated by studies in platelets. Platelets 2005, 16, 133–150. [Google Scholar] [CrossRef]
- Bonifacino, J.S.; Merćep, M.; Sussman, J.J.; Klausner, R.D.; Ashwell, J.D. The T-cell antigen receptor: A complex signal-transducing molecule. Princess Takamatsu Symp. 1988, 19, 87–104. [Google Scholar]
- Stankunas, K.; Graef, I.; Neilson, J.R.; Park, S.-H.; Crabtree, G. Signaling through calcium, calcineurin, and NF-AT in lymphocyte activation and development. Cold Spring Harb. Symp. Quant. Biol. 1999, 64, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Im, S.H.; Rao, A. Activation and deactivation of gene expression by Ca2+/calcineurin-NFAT-mediated signaling. Mol. Cells 2004, 18, 1–9. [Google Scholar] [PubMed]
- Trebak, M.; Kinet, J.-P. Calcium signalling in T cells. Nat. Rev. Immunol. 2019, 19, 154–169. [Google Scholar] [CrossRef] [PubMed]
- Cheadle, C.; Fan, J.; Cho-Chung, Y.S.; Werner, T.; Ray, J.; Do, L.; Gorospe, M.; Becker, K.G. Control of gene expression during T cell activation: Alternate regulation of mRNA transcription and mRNA stability. BMC Genom. 2005, 6, 75. [Google Scholar] [CrossRef] [Green Version]
- Banerji, S.S.; Parsons, J.N.; Tocci, M.J. The immunosuppressant FK-506 specifically inhibits mitogen-induced activation of the interleukin-2 promoter and the isolated enhancer elements NFIL-2A and NF-AT1. Mol. Cell. Biol. 1991, 11, 4074–4087. [Google Scholar] [CrossRef] [Green Version]
- Bornkamm, G.W.; Hammerschmidt, W. Molecular virology of Epstein-Barr virus. Philos. Trans. R. Soc. B: Biol. Sci. 2001, 356, 437–459. [Google Scholar] [CrossRef] [Green Version]
- Rowe, D.T. Epstein-Barr virus immortalization and latency. Front. Biosci. 1999, 4, d346. [Google Scholar] [CrossRef] [Green Version]
- Strobl, L.J.; Höfelmayr, H.; Stein, C.; Marschall, G.; Brielmeier, M.; Laux, G.; Bornkamm, G.W.; Zimber-Strobl, U. Both Epstein-Barr Viral Nuclear Antigen 2 (EBNA2) and Activated Notch1 Transactivate Genes by Interacting with the Cellular Protein RBP-Jκ. Immunobiology 1997, 198, 299–306. [Google Scholar] [CrossRef]
- Kieser, A.; Sterz, K.R. The Latent Membrane Protein 1 (LMP1). Mol. Asp. Myeloid Stem Cell Dev. 2015, 391, 119–149. [Google Scholar]
- Farrell, P.J. Epstein-Barr virus immortalizing genes. Trends Microbiol. 1995, 3, 105–109. [Google Scholar] [CrossRef]
- Gou, W.-F.; Niu, Z.-F.; Zhao, S.; Takano, Y.; Zheng, H. Aberrant SERCA3 expression during the colorectal adenoma-adenocarcinoma sequence. Oncol. Rep. 2013, 31, 232–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.-Y.; Gou, W.-F.; Yang, X.; Wang, G.-L.; Takahashi, H.; Yu, M.; Mao, X.-Y.; Takano, Y.; Zheng, H.-C. Aberrant SERCA3 expression is closely linked to pathogenesis, invasion, metastasis, and prognosis of gastric carcinomas. Tumor Biol. 2012, 33, 1845–1854. [Google Scholar] [CrossRef]
- Meneses-Morales, I.; Izquierdo-Torres, E.; Flores-Peredo, L.; Rodríguez, G.; Hernández-Oliveras, A.; Zarain-Herzberg, A. Epigenetic regulation of the human ATP2A3 gene promoter in gastric and colon cancer cell lines. Mol. Carcinog. 2019, 58, 887–897. [Google Scholar] [CrossRef] [PubMed]
- Jen, J.; Powell, S.M.; Papadopoulos, N.; Smith, K.J.; Hamilton, S.R.; Vogelstein, B.; Kinzler, K.W. Molecular determinants of dysplasia in colorectal lesions. Cancer Res. 1994, 54, 5523–5526. [Google Scholar]
- Van De Wetering, M.; Sancho, E.; Verweij, C.; De Lau, W.; Oving, I.; Hurlstone, A.F.L.; Van Der Horn, K.; Batlle, E.; Coudreuse, D.; Haramis, A.-P.; et al. The β-Catenin/TCF-4 Complex Imposes a Crypt Progenitor Phenotype on Colorectal Cancer Cells. Cell 2002, 111, 241–250. [Google Scholar] [CrossRef] [Green Version]
- Flores-Peredo, L.; Rodríguez, G.; Zarain-Herzberg, A. Induction of cell differentiation activates transcription of the Sarco/Endoplasmic Reticulum calcium-ATPase 3 gene (ATP2A3) in gastric and colon cancer cells. Mol. Carcinog. 2016, 56, 735–750. [Google Scholar] [CrossRef]
- Lizák, B.; Csala, M.; Benedetti, A.; Bánhegyi, G. The translocon and the non-specific transport of small molecules in the endoplasmic reticulum (Review). Mol. Membr. Biol. 2008, 25, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Leal, E.; Hernández-Oliveras, A.; Flores-Peredo, L.; Zarain-Herzberg, A.; Santiago-García, J. Histone deacetylase inhibitors promote the expression ofATP2A3 gene in breast cancer cell lines. Mol. Carcinog. 2015, 55, 1477–1485. [Google Scholar] [CrossRef]
- Izquierdo-Torres, E.; Hernández-Oliveras, A.; Meneses-Morales, I.; Rodríguez, G.; Fuentes-García, G.; Zarain-Herzberg, A. Resveratrol up-regulates ATP2A3 gene expression in breast cancer cell lines through epigenetic mechanisms. Int. J. Biochem. Cell Biol. 2019, 113, 37–47. [Google Scholar] [CrossRef]
- Azeez, J.M.; Vini, R.; Remadevi, V.; Surendran, A.; Jaleel, A.; Kumar, T.R.S.; Sreeja, S.; Ravindran, V.; Sreeharshan, S. VDAC1 and SERCA3 Mediate Progesterone-Triggered Ca2+ Signaling in Breast Cancer Cells. J. Proteome Res. 2017, 17, 698–709. [Google Scholar] [CrossRef]
- Shapey, J.; Toma, A.; Saeed, S. Physiology of cerebrospinal fluid circulation. Curr. Opin. Otolaryngol. Head Neck Surg. 2019, 27, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.; Ruland, V.; Kordes, U.; Hartung, S.; Capper, D.; Pietsch, T.; Gerß, J.; Wolff, J.E.A.; Paulus, W.; Hasselblatt, M. Pediatric atypical choroid plexus papilloma reconsidered: Increased mitotic activity is prognostic only in older children. Acta Neuropathol. 2015, 129, 925–927. [Google Scholar] [CrossRef] [PubMed]
- Wrede, B.; Hasselblatt, M.; Peters, O.; Thall, P.F.; Kutluk, T.; Moghrabi, A.; Mahajan, A.; Rutkowski, S.; Diez, B.; Wang, X.; et al. Atypical choroid plexus papilloma: Clinical experience in the CPT-SIOP-2000 study. J. Neuro-Oncol. 2009, 95, 383–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeibmann, A.; Wrede, B.; Peters, O.; Wolff, J.E.; Paulus, W.; Hasselblatt, M. Malignant progression in choroid plexus papillomas. J. Neurosurg. Pediatr. 2007, 107, 199–202. [Google Scholar] [CrossRef]
- Jeibmann, A.; Hasselblatt, M.; Gerss, J.; Wrede, B.; Egensperger, R.; Beschorner, R.; Hans, V.H.J.; Rickert, C.H.; Wolff, J.E.; Paulus, W. Prognostic Implications of Atypical Histologic Features in Choroid Plexus Papilloma. J. Neuropathol. Exp. Neurol. 2006, 65, 1069–1073. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, Y.; Matsushita, H.; Plese, J.P.; Marino, R., Jr. Hydrocephalus due to diffuse villous hyperplasia of the choroid plexus. Case report and review of the literature. Pediatr. Neurosurg. 2004, 40, 32–36. [Google Scholar] [CrossRef]
- Hallaert, G.G.; Vanhauwaert, D.J.; Logghe, K.; Broecke, C.V.D.; Baert, E.; Van Roost, D.; Caemaert, J. Endoscopic coagulation of choroid plexus hyperplasia. J. Neurosurg. Pediatr. 2012, 9, 169–177. [Google Scholar] [CrossRef]
- Ribiczey, P.; Tordai, A.; Andrikovics, H.; Filoteo, A.G.; Penniston, J.T.; Enouf, J.; Enyedi, A.; Papp, B.; Kovács, T. Isoform-specific up-regulation of plasma membrane Ca2+ATPase expression during colon and gastric cancer cell differentiation. Cell Calcium 2007, 42, 590–605. [Google Scholar] [CrossRef] [Green Version]
- Aung, C.S.; Kruger, W.A.; Poronnik, P.; Roberts-Thomson, S.; Monteith, G. Plasma membrane Ca2+-ATPase expression during colon cancer cell line differentiation. Biochem. Biophys. Res. Commun. 2007, 355, 932–936. [Google Scholar] [CrossRef]
- Ribiczey, P.; Papp, B.; Homolya, L.; Enyedi, Á.; Kovács, T. Selective upregulation of the expression of plasma membrane calcium ATPase isoforms upon differentiation and 1,25(OH)2D3-vitamin treatment of colon cancer cells. Biochem. Biophys. Res. Commun. 2015, 464, 189–194. [Google Scholar] [CrossRef]
- Curry, M.; Roberts-Thomson, S.; Monteith, G. PMCA2 silencing potentiates MDA-MB-231 breast cancer cell death initiated with the Bcl-2 inhibitor ABT-263. Biochem. Biophys. Res. Commun. 2016, 478, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, J.; VanHouten, J.N.; Dann, P.; Kim, W.; Sullivan, C.; Yu, H.; Liotta, L.; Espina, V.; Stern, D.F.; Friedman, P.A.; et al. PMCA2 regulates HER2 protein kinase localization and signaling and promotes HER2-mediated breast cancer. Proc. Natl. Acad. Sci. USA 2016, 113, E282–E290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varga, K.; Pászty, K.; Padanyi, R.; Hegedüs, L.; Brouland, J.-P.; Papp, B.; Enyedi, Á. Histone deacetylase inhibitor- and PMA-induced upregulation of PMCA4b enhances Ca2+ clearance from MCF-7 breast cancer cells. Cell Calcium 2014, 55, 78–92. [Google Scholar] [CrossRef] [PubMed]
- Roberts-Thomson, S.J.; Curry, M.C.; Monteith, G. Plasma membrane calcium pumps and their emerging roles in cancer. World J. Biol. Chem. 2010, 1, 248–253. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.J.; Roberts-Thomson, S.; Monteith, G. Plasma membrane calcium-ATPase 2 and 4 in human breast cancer cell lines. Biochem. Biophys. Res. Commun. 2005, 337, 779–783. [Google Scholar] [CrossRef]
- Varga, K.; Hollósi, A.; Pászty, K.; Hegedűs, L.; Szakács, G.; Tímár, J.; Papp, B.; Enyedi, Á.; Padányi, R. Expression of calcium pumps is differentially regulated by histone deacetylase inhibitors and estrogen receptor alpha in breast cancer cells. BMC Cancer 2018, 18, 1029. [Google Scholar] [CrossRef] [Green Version]
- VanHouten, J.; Sullivan, C.; Bazinet, C.; Ryoo, T.; Camp, R.; Rimm, D.L.; Chung, G.; Wysolmerski, J. PMCA2 regulates apoptosis during mammary gland involution and predicts outcome in breast cancer. Proc. Natl. Acad. Sci. USA 2010, 107, 11405–11410. [Google Scholar] [CrossRef] [Green Version]
- Reinhardt, T.; Lippolis, J. Mammary gland involution is associated with rapid down regulation of major mammary Ca2+-ATPases. Biochem. Biophys. Res. Commun. 2009, 378, 99–102. [Google Scholar] [CrossRef]
- Faddy, H.; Smart, C.E.; Xu, R.; Lee, G.Y.; Kenny, P.A.; Feng, M.; Rao, R.; Brown, M.; Bissell, M.J.; Roberts-Thomson, S.; et al. Localization of plasma membrane and secretory calcium pumps in the mammary gland. Biochem. Biophys. Res. Commun. 2008, 369, 977–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinhardt, T.; Lippolis, J.; Shull, G.E.; Horst, R.L. Null Mutation in the Gene Encoding Plasma Membrane Ca2+-ATPase Isoform 2 Impairs Calcium Transport into Milk. J. Biol. Chem. 2004, 279, 42369–42373. [Google Scholar] [CrossRef] [Green Version]
- Tagliavacca, L.; Anelli, T.; Fagioli, C.; Mezghrani, A.; Ruffato, E.; Sitia, R.; Sitia, L.R. The Making of a Professional Secretory Cell: Architectural and Functional Changes in the ER during B Lymphocyte Plasma Cell Differentiation. Biol. Chem. 2003, 384, 1273–1277. [Google Scholar] [CrossRef] [PubMed]
- Reinhardt, T.; Horst, R.L. Ca2+-ATPases and their expression in the mammary gland of pregnant and lactating rats. Am. J. Physiol. Content 1999, 276, C796–C802. [Google Scholar] [CrossRef] [PubMed]
- Hegedũs, L.; Garay, T.; Molnar, E.; Varga, K.; Bilecz, Á.; Török, S.; Padányi, R.; Pászty, K.; Wolf, M.; Grusch, M.; et al. The plasma membrane Ca2+ pump PMCA 4b inhibits the migratory and metastatic activity of BRAF mutant melanoma cells. Int. J. Cancer 2016, 140, 2758–2770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegedűs, L.; Padányi, R.; Molnár, J.; Pászty, K.; Varga, K.; Kenessey, I.; Sárközy, E.; Wolf, M.; Grusch, M.; Hegyi, Z.; et al. Histone Deacetylase Inhibitor Treatment Increases the Expression of the Plasma Membrane Ca2+ Pump PMCA4b and Inhibits the Migration of Melanoma Cells Independent of ERK. Front. Oncol. 2017, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aulestia, F.J.; Redondo, P.C.; Rodríguez-García, A.; Rosado, J.A.; Salido, G.M.; Alonso, M.T.; García-Sancho, J. Two distinct calcium pools in the endoplasmic reticulum of HEK-293T cells. Biochem. J. 2011, 435, 227–235. [Google Scholar] [CrossRef] [Green Version]
- Papp, B.; Pászty, K.; Kovacs, T.; Sarkadi, B.; Gárdos, G.; Enouf, J.; Enyedi, A. Characterization of the inositol trisphosphate-sensitive and insensitive calcium stores by selective inhibition of the endoplasmic reticulum-type calcium pump isoforms in isolated platelet membrane vesicles. Cell Calcium 1993, 14, 531–538. [Google Scholar] [CrossRef]
- Papp, S.; Dziak, E.; Michalak, M.; Opas, M. Is all of the endoplasmic reticulum created equal? The effects of the heterogeneous distribution of endoplasmic reticulum Ca2+-handling proteins. J. Cell Biol. 2003, 160, 475–479. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.G.; Xu, X.; Zeng, W.; Diaz, J.; Kuo, T.H.; Wuytack, F.; Racymaekers, L.; Muallem, S. Polarized expression of Ca2+ pumps in pancreatic and salivary gland cells. Role in initiation and propagation of [Ca2+]i waves. J. Biol. Chem. 1997, 272, 15771–15776. [Google Scholar] [CrossRef] [Green Version]
- Bird, G.S.; Putney, J. Capacitative calcium entry supports calcium oscillations in human embryonic kidney cells. J. Physiol. 2004, 562, 697–706. [Google Scholar] [CrossRef]
- Hudmon, A.; Schulman, H. Structure–function of the multifunctional Ca2+/calmodulin-dependent protein kinase II. Biochem. J. 2002, 364, 593–611. [Google Scholar] [CrossRef]
- Fisher, W.G.; Yang, P.-C.; Medikonduri, R.K.; Jafri, M.S. NFAT and NFκB Activation in T Lymphocytes: A Model of Differential Activation of Gene Expression. Ann. Biomed. Eng. 2006, 34, 1712–1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerrero-Hernández, A.; Dagnino-Acosta, A.; Verkhratsky, A. An intelligent sarco-endoplasmic reticulum Ca2+ store: Release and leak channels have differential access to a concealed Ca2+ pool. Cell Calcium 2010, 48, 143–149. [Google Scholar] [CrossRef] [PubMed]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papp, B.; Launay, S.; Gélébart, P.; Arbabian, A.; Enyedi, A.; Brouland, J.-P.; Carosella, E.D.; Adle-Biassette, H. Endoplasmic Reticulum Calcium Pumps and Tumor Cell Differentiation. Int. J. Mol. Sci. 2020, 21, 3351. https://doi.org/10.3390/ijms21093351
Papp B, Launay S, Gélébart P, Arbabian A, Enyedi A, Brouland J-P, Carosella ED, Adle-Biassette H. Endoplasmic Reticulum Calcium Pumps and Tumor Cell Differentiation. International Journal of Molecular Sciences. 2020; 21(9):3351. https://doi.org/10.3390/ijms21093351
Chicago/Turabian StylePapp, Bela, Sophie Launay, Pascal Gélébart, Atousa Arbabian, Agnes Enyedi, Jean-Philippe Brouland, Edgardo D. Carosella, and Homa Adle-Biassette. 2020. "Endoplasmic Reticulum Calcium Pumps and Tumor Cell Differentiation" International Journal of Molecular Sciences 21, no. 9: 3351. https://doi.org/10.3390/ijms21093351
APA StylePapp, B., Launay, S., Gélébart, P., Arbabian, A., Enyedi, A., Brouland, J. -P., Carosella, E. D., & Adle-Biassette, H. (2020). Endoplasmic Reticulum Calcium Pumps and Tumor Cell Differentiation. International Journal of Molecular Sciences, 21(9), 3351. https://doi.org/10.3390/ijms21093351