Mechanisms of Reciprocal Regulation of Gonadotropin-Releasing Hormone (GnRH)-Producing and Immune Systems: The Role of GnRH, Cytokines and Their Receptors in Early Ontogenesis in Normal and Pathological Conditions


Abstract
:1. Introduction
2. Effects of Different Signal Molecules on the Normal Development of the GnRH System
2.1. Adhesion Molecules
2.2. Guidance-Cue Molecules
2.3. Cytokines
2.4. Neurotransmitters
2.5. Transcription Factors
3. Development of the GnRH System in Different Pathological States
4. Effects of GnRH on the Development and Functioning of the Immune System
5. Approaches to Preventing or Reversing Disorders Induced by Acute Inflammation in Early Ontogenesis
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| CNS | Central nervous system |
| ED | Embryonic day |
| ERα | Estrogen receptor alfa |
| FSH | Follicle-stimulating hormone |
| GABA | Gamma-aminobutyric acid |
| GM-CSF | Granulocyte macrophage colony-stimulating factor |
| GnRH | Gonadotropin-releasing hormone |
| HGF/SF | Hepatocyte growth factor/scatter factor |
| HPG | Hypothalamic-pituitary-gonadal (system) |
| IFNγ | Interferon gamma |
| IgG | Immunoglobulin G |
| IL | Interleukin |
| KISS1 | Kisspeptin |
| LH | Luteinizing hormone |
| LIF | Leukemia inhibitory factor |
| LPS | Lipopolysaccharide |
| MCP-1 | Monocyte chemoattractant protein-1 |
| SDF-1 | Stromal cell-derived factor 1 and one of its receptors CXCR4 |
| SEMA | Semaphorin |
| TNF | Tumor necrosis factor (alpha and beta) |
References
- Tomaszewska-Zaremba, D.; Herman, A. The role of immunological system in the regulation of gonadoliberin and gonadotropin secretion. Reprod. Biol. 2009, 9, 11–23. [Google Scholar] [CrossRef]
- Zakharova, L.A.; Izvolskaia, M.S. Interactions between the reproductive and immune systems during ontogenesis: The role of GnRH, sex steroids and immunomediators. In Sex Steroids; Kahn, S.M., Ed.; InTech: Rijeka, Croatia; London, UK, 2012; pp. 341–370. [Google Scholar]
- Quintanar, J.L.; Guzmán-Soto, I. Hypothalamic neurohormones and immune responses. Front. Integr. Neurosci. 2013, 7, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segner, H.; Verburg-van Kemenade, B.M.L.; Chadzinska, M. The immunomodulatory role of the hypothalamus-pituitary-gonad axis: Proximate mechanism for reproduction-immune trade offs? Dev. Comp. Immunol. 2017, 66, 43–60. [Google Scholar] [CrossRef] [PubMed]
- Stamatiades, G.A.; Kaiser, U.B. Gonadotropin regulation by pulsatile GnRH: Signaling and gene expression. Mol Cell Endocrinol. 2018, 463, 131–141. [Google Scholar] [CrossRef]
- Melnikova, V.I.; Lifantseva, N.V.; Voronova, S.N.; Zakharova, L.A. Gonadotropin-Releasing Hormone in regulation of thymic development in rats: Profile of thymic cytokines. Int. J. Mol. Sci. 2019, 20, 4033. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Ren, X.; Zeng, Y.; Zhou, Y.; Song, T.; Cao, X.; Du, X.; Meng, F.; Tan, Y.; Liu, Y.; et al. Physiological interactions between the hypothalamic-pituitary-gonadal axis and spleen in rams actively immunized against GnRH. Int. Immunopharmacol. 2016, 38, 275–283. [Google Scholar] [CrossRef]
- Clarkson, J.; Han, S.Y.; Piet, R.; McLennan, T.; Kane, G.M.; Ng, J.; Porteous, R.W.; Kim, J.S.; Colledge, W.H.; Iremonger, K.J.; et al. Definition of the hypothalamic GnRH pulse generator in mice. Proc. Natl. Acad. Sci. USA 2017, 114, E10216–E10223. [Google Scholar] [CrossRef] [Green Version]
- Zakharova, L.A. Plasticity of neuroendocrine-immune interactions during ontogeny: Role of perinatal programming in pathogenesis of inflammation and stress-related diseases in adults. Recent Pat. Endocr. Metab. Immune Drug Discov. 2009, 3, 11–27. [Google Scholar] [CrossRef]
- Fowden, A.L.; Forhead, A.J. Endocrine mechanisms of intrauterine programming. Reproduction 2004, 127, 515–526. [Google Scholar] [CrossRef]
- Zakharova, L.A.; Ermilova, I.Y.; Melnikova, V.I.; Malyukova, I.V.; Adamskaya, E.I. Hypothalamic control of the cell-mediated immunity and of the Luteinizing Hormone Releasing Hormone level in thymus and peripheral blood of rat fetuses. Neuroimmunomodulation 2005, 12, 85–91. [Google Scholar] [CrossRef]
- Ugrumov, M.V.; Sapronova, A.Y.; Melnikova, V.I.; Proshlyakova, E.V.; Adamskaya, E.I.; Lavrentieva, A.V.; Nasirova, D.I.; Babichev, V.N. Brain is an important source of GnRH in general circulation in the rat during prenatal and early postnatal ontogenesis. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2005, 141, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Pronina, T.; Ugrumov, M.; Calas, A.; Seif, I.; Tramu, G. Influence of monoamines on differentiating gonadotropin-releasing hormone neurones in foetal mice. J. Neuroendocrinol. 2003, 15, 925–932. [Google Scholar] [CrossRef] [PubMed]
- Giacobini, P.; Messina, A.; Morello, F.; Ferraris, N.; Corso, S.; Penachioni, J.; Giordano, S.; Tamagnone, L.; Fasolo, A. Semaphorin 4D regulates gonadotropin hormone-releasing hormone-1 neuronal migration through PlexinB1-Met complex. J. Cell Biol. 2008, 183, 555–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oleari, R.; Lettieri, A.; Paganoni, A.; Zanieri, L.; Cariboni, A. Semaphorin signaling in GnRH neurons: From development to disease. Neuroendocrinology 2019, 109, 193–199. [Google Scholar] [CrossRef]
- Casoni, F.; Hutchins, B.I.; Donohue, D.; Fornaro, M.; Condie, B.G.; Wray, S. SDF and GABA interact to regulate axophilic migraton of GnRH neurons. J. Cell. Sci. 2012, 25, 5015–5025. [Google Scholar] [CrossRef] [Green Version]
- Herbison, A.E. The Gonadotropin-Releasing Hormone pulse generator. Endocrinology 2018, 159, 3723–3736. [Google Scholar] [CrossRef] [Green Version]
- Uenoyama, Y.; Inoue, N.; Kei-Maeda, I.; Tsukamura, H. The roles of kisspeptin in the mechanism underlying reproductive functions in mammals. J. Reprod. Dev. 2018, 64, 469–476. [Google Scholar] [CrossRef] [Green Version]
- Tena-Sempere, M. GPR54 and kisspeptin in reproduction. Hum. Reprod. Update 2006, 12, 631–639. [Google Scholar] [CrossRef] [Green Version]
- Dhillo, W.S.; Murphy, K.G.; Bloom, S.R. The neuroendocrine physiology of kisspeptin in the human. Rev. Endocrinol. Metab. Disord. 2007, 8, 41–46. [Google Scholar] [CrossRef]
- Han, X.; Zhou, Y.; Zeng, Y.; Sui, F.; Liu, Y.; Tan, Y.; Cao, X.; Du, X.; Meng, F.; Zeng, X. Effects of active immunization against GnRH versus surgical castration on hypothalamic-pituitary function in boars. Theriogenology 2017, 97, 89–97. [Google Scholar] [CrossRef]
- Wakabayashi, Y.; Yamamura, T.; Sakamoto, K.; Mori, Y.; Okamura, H. Electrophysiological and Morphological Evidence for Synchronized GnRH Pulse Generator Activity among Kisspeptin/neurokinin B/dynorphin A (KNDy) Neurons in Goats. J. Reprod. Dev. 2013, 59, 40–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comninos, A.N.; Anastasovska, J.; Sahuri-Arisoylu, M.; Li, X.; Li, S.; Hu, M.; Jayasena, C.N.; Ghatei, M.A.; Bloom, S.R.; Matthews, P.M.; et al. Kisspeptin signaling in the amygdala modulates reproductive hormone secretion. Brain Struct. Funct. 2016, 221, 2035–2047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hrabovszky, E.; Ciofi, P.; Vida, B.; Horvath, M.; Keller, E.; Caraty, A.; Bloom, S.; Ghatei, M.; Dhillo, W.; Liposits, Z.; et al. The kisspeptin system of the human hypothalamus: Sexual dimorphism and relationship with gonadotropin-releasing hormone and neurokinin B neurons. Eur. J. Neurosci. 2010, 11, 1984–1998. [Google Scholar] [CrossRef] [PubMed]
- Yip, S.H.; Boehm, U.; Herbison, A.E.; and Campbell, R.E. Conditional viral tract tracing delineates the projections of the distinct kisspeptin neuron populations to gonadotropin-releasing hormone (GnRH) neurons in the mouse. Endocrinololy 2015, 156, 2582–2594. [Google Scholar] [CrossRef] [Green Version]
- Lehman, M.N.; Coolen, L.M.; Goodman, R.L. Minireview: Kisspeptin/neurokinin B/dynorphin (KNDy) cells of the arcuate nucleus: A central node in the control of gonadotropin-releasing hormone secretion. Endocrinology 2010, 151, 3479–3489. [Google Scholar] [CrossRef]
- Moore, A.M.; Coolen, L.M.; Porter, D.T.; Goodman, R.L.; Lehman, M.N. KNDy cells revisited. Endocrinology 2018, 159, 3219–3234. [Google Scholar] [CrossRef] [Green Version]
- Morale, M.C.; Batticane, N.; Bartoloni, G.; Guarcello, V.; Farinella, Z.; Galasso, M.G.; Marchetti, B. Blocade of central and peripheral luteinizing hormone-releasing hormone (LHRH) receptors in neonatal rats with a potent LHRH-antagonist inhibits the morphofunctional development of the thymus and maturation of the cell-mediated and humoral immune responses. Endocrinology 1991, 128, 1073–1085. [Google Scholar] [CrossRef]
- Dubois, E.A.; Zandbergen, M.A.; Peute, J.; Goos, H.J. Evolutionary development of three gonadotropin releasing hormone (GnRH) systems in vertebrates. Brain Res. Bull. 2002, 57, 413–418. [Google Scholar] [CrossRef]
- Millar, R.P. GnRHs and GnRH Receptors. Anim. Reprod. Sci. 2005, 88, 5–28. [Google Scholar] [CrossRef]
- Wu, H.M.; Wang, H.S.; Huang, H.Y.; Soong, Y.K.; MacCalman, C.D.; Leung, P.C. GnRH signaling in intrauterine tissues. Reproduction 2009, 137, 769–777. [Google Scholar] [CrossRef] [Green Version]
- Millar, R.P.; Lu, Z.L.; Pawson, A.J.; Flanagan, C.A.; Morgan, K.; Maudsley, S.R. Gonadotropin-releasing hormone receptors. Endocr. Rev. 2004, 25, 235–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramakrishnappa, N.; Rajamahendran, R.; Lin, Y.M.; Leung, P. C: GnRH in non-hypothalamic reproductive tissues. Anim. Reprod. Sci. 2005, 88, 95–113. [Google Scholar] [CrossRef] [PubMed]
- Walters, K.; Wegorzewska, I.N.; Chin, Y.P.; Parikh, M.G.; Wu, T.J. Luteinizing hormone-releasing hormone I (LHRH-I) and its metabolite in peripheral tissues. Exp. Biol. Med. 2008, 233, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Tanriverdi, F.; Silveira, L.F.G.; MacColl, G.S.; Bouloux, P.M.G. The hypothalamic-pituitary-gonadal axis: Immune function and autoimmunity. J. Endocrinol. 2003, 176, 293–304. [Google Scholar] [CrossRef] [Green Version]
- Bornstein, S.R.; Rutkowski, H.; Vrezas, I. Cytokines and steroidogenesis. Mol. Cell. Endocrinol. 2004, 215, 135–141. [Google Scholar] [CrossRef]
- Sharova, V.S.; Izvolskaia, M.S.; Zakharova, L.A. Lipopolysaccharide-induced maternal inflammation affects the gonadotropin-releasing hormone neuron development in fetal mice. Neuroimmunomodulation 2015, 22, 222–232. [Google Scholar] [CrossRef]
- Turnbull, A.V.; Rivier, C.L. Regulation of the hypothalamic-pituitary-adrenal axis by cytokines: Actions and mechanisms of action. Physiol. Rev. 1999, 79, 1–71. [Google Scholar] [CrossRef] [Green Version]
- Barabás, K.; Szabó-Meleg, E.; Ábrahám, I.M. Effect of inflammation on female Gonadotropin-Releasing Hormone (GnRH) neurons: Mechanisms and consequences. Int. J. Mol. Sci. 2020, 21, 529. [Google Scholar] [CrossRef] [Green Version]
- Zuckerman, L.; Weiner, I. Maternal immune activation leads to behavioral and pharmacological changes in the adult offspring. J. Psychiatr. Res. 2005, 39, 311–323. [Google Scholar] [CrossRef]
- Liverman, C.S.; Kaftan, H.A.; Cui, L.; Hersperger, S.G.; Taboada, E.; Klein, R.M.; Berman, N.E. Altered expression of pro-inflammatory and developmental genes in the fetal brain in a mouse model of maternal infection. Neurosci. Lett. 2006, 399, 220–225. [Google Scholar] [CrossRef]
- Wang, X.; Hagberg, H.; Zhu, C.; Jacobsson, B.; Mallard, C. Effects of intrauterine inflammation on the developing mouse brain. Brain Res. 2007, 1144, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yan, J.Y.; Lo, Y.K.; Carvey, P.M.; Ling, Z. Dopaminergic and serotoninergic deficiencies in young adult rats prenatally exposed to the bacterial lipopolysaccharide. Brain Res. 2009, 1265, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Kirsten, T.B.; Chaves, G.P.; Taricano, M.; Martins, D.O.; Flório, J.C.; Britto, L.R.; Torrão, A.S.; Palermo-Neto, J.; Bernardi, M.M. Prenatal LPS exposure reduces olfactory perception in neonatal and adult rats. Physiol. Behav. 2011, 104, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Clark, L.F.; Kodadek, T. The Immune System and Neuroinflammation as Potential Sources of Blood-Based Biomarkers for Alzheimer’s Disease, Parkinson’s Disease, and Huntington’s Disease. ACS Chem. Neurosci. 2016, 7, 520–527. [Google Scholar] [CrossRef]
- Ardalan, M.; Chumak, T.; Vexler, Z.; Mallard, C. Sex-Dependent Effects of Perinatal Inflammation on the Brain: Implication for Neuro-Psychiatric Disorders. Int. J. Mol. Sci. 2019, 20, 2270. [Google Scholar] [CrossRef] [Green Version]
- Izvolskaia, M.S.; Sharova, V.S.; Zakharova, L.A. Perinatal Inflammation Reprograms Neuroendocrine, Immune, and Reproductive Functions: Profile of Cytokine Biomarkers. Inflammation 2020, 43, 1175–1183. [Google Scholar] [CrossRef]
- Straley, M.E.; Togher, K.L.; Nolan, A.M.; Kenny, L.C.; O’Keeffe, G.W. LPS alters placental inflammatory and endocrine mediators and inhibits fetal neurite growth in affected offspring during late gestation. Placenta 2014, 35, 533–538. [Google Scholar] [CrossRef]
- Wang, H.; Yang, L.L.; Hu, Y.F.; Wang, B.W.; Huang, Y.Y.; Zhang, C.; Chen, Y.H.; Xu, D.X. Maternal LPS exposure during pregnancy impairs testicular development, steroidogenesis and spermatogenesis in male offspring. PLoS ONE 2014, 9, e106786. [Google Scholar] [CrossRef]
- Izvolskaia, M.S.; Tillet, Y.; Sharova, V.S.; Voronova, S.N.; Zakharova, L.A. Disruptions in the hypothalamic-pituitary-gonadal axis in rat offspring following prenatal maternal exposure to lipopolysaccharide. Stress 2016, 19, 198–205. [Google Scholar] [CrossRef]
- Izvolskaia, M.S.; Sharova, V.S.; Ignatiuk, V.M.; Voronova, S.N.; Zakharova, L.A. Abolition of prenatal lipopolysaccharide-induced reproductive disorders in rat male offspring by fulvestrant. Andrologia 2019, 51, e13204. [Google Scholar] [CrossRef]
- Ignatiuk, V.M.; Izvolskaya, M.S.; Sharova, V.S.; Voronova, S.N.; Zakharova, L.A. Disruptions in the reproductive system of female rats after prenatal lipopolysaccharide-induced immunological stress: Role of sex steroids. Stress 2019, 22, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Lysenko, L.A. Back to anthropology: What does it mean to development studies? Beacon. J. Stud. Ideol. Ment. Dimens. 2019, 2, 020000000. Available online: https://hdl.handle.net/20.500.12656/thebeacon.2.020000000 (accessed on 22 December 2020).
- Sassin, W. Deja Vue. Beacon. J. Stud. Ideol. Ment. Dimens. 2019, 2, 020210216. Available online: https://hdl.handle.net/20.500.12656/thebeacon.2.020210216 (accessed on 22 December 2020).
- Sassin, W. Er-Schöpfung der Schöpfung, oder Eine neue Kulturstufe in der Entwicklung des homo. Beacon. J. Stud. Ideol. Ment. Dimens. 2019, 2, 020510203. Available online: https://hdl.handle.net/20.500.12656/thebeacon.2.020510203. (accessed on 22 December 2020). (In German).
- Sassin, W. Die Grenzen der Ökonomie: Globalisierung—Vom Füllhorn zum Giftbecher? Eur. Crossrd. 2020, 1, 010410216. Available online: https://hdl.handle.net/20.500.12656/eurcrossrd.1.010410216. (accessed on 22 December 2020). (In German).
- Vitkovic, L.; Konsman, J.P.; Bockaert, J.; Dantzer, R.; Homburger, V.; Jacque, C. Cytokine signals propagate through the brain. Mol. Psychiatry 2000, 5, 604–615. [Google Scholar] [CrossRef] [Green Version]
- Prinz, M.; Priller, J. The role of peripheral immune cells in the CNS in steady state and disease. Nat. Neurosci. 2017, 20, 136–144. [Google Scholar] [CrossRef]
- McCann, S.M.; Kimura, M.; Karanth, S.; Yu, W.H.; Mastronardi, C.A.; Rettori, V. The mechanism of action of cytokines to control the release of hypothalamic and pituitary hormones in infection. Ann. N. Y. Acad. Sci. 2000, 917, 4–18. [Google Scholar] [CrossRef]
- Igaz, P.; Salvi, R.; Rey, J.P.; Glauser, M.; Pralong, F.P.; Gaillard, R.C. Effects of cytokines on gonadotropin-releasing hormone (GnRH) gene expression in primary hypothalamic neurons and in GnRH neurons immortalized conditionally. Endocrinology. 2006, 147, 1037–1043. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, K.; Tobet, S.A.; Crandall, J.E.; Jimenez, T.P.; Schwarting, G.A. The migration of luteinising hormone-releasing hormone neurons in the developing rat is associated with a transient, caudal projection of the vomeronasal nerve. J. Neurosci. 1995, 15, 7769–7777. [Google Scholar] [CrossRef]
- Parkash, J.; Cimino, I.; Ferraris, N.; Casoni, F.; Wray, S.; Cappy, H.; Prevot, V.; Giacobini, P. Suppression of β1-Integrin in gonadotropin-releasing hormone cells disrupts migration and axonal extension resulting in severe reproductive alterations. J. Neurosci. 2012, 32, 16992–17002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, S.; Yamazaki, C.; Masumoto, K.H.; Nagano, M.; Naito, M.; Soga, T.; Hiyama, H.; Matsumoto, M.; Takasaki, J.; Kamohara, M.; et al. Abnormal development of the olfactory bulb and reproductive system in mice lacking prokineticin receptor PKR2. Proc. Natl. Acad. Sci. USA 2006, 103, 4140–4145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, K.; Rutishauser, U.; Crandall, J.E.; Schwarting, G.A. Polysialic acid facilitates migration of luteinezing hormone-releasing hormone neurons on vomeronasal nerve. J. Neurosci. 1999, 19, 794–801. [Google Scholar] [CrossRef] [Green Version]
- Battista, D.; Rutishauser, U. Removal of polysialic acid triggers dispersion of subventricularly derived neuroblasts into surrounding CNS tissues. J. Neurosci. 2010, 30, 3995–4003. [Google Scholar] [CrossRef] [PubMed]
- Huilgol, D.; Tole, S. Cell migration in the developing rodent olfactory system. Cell. Mol. Life Sci. 2016, 73, 2467–2490. [Google Scholar] [CrossRef] [Green Version]
- Tobet, S.A.; Crandall, J.E.; Schawrting, G.A. Relationship of migrating luteinezing hormone-releasing hormone neurons to unique olfactory system of glycoconjugates in embryonic rats. Dev. Biol. 1993, 155, 471–482. [Google Scholar] [CrossRef]
- Fueshko, S.; Wray, S. LHRH cells migrate on periferin fibres in embryonic olfactory explant cultures: An in vitro model for neurotrophic migration. Dev. Biol. 1994, 166, 331–348. [Google Scholar] [CrossRef]
- Rasterkamp, R.J.; Ruitenberg, M.J.; Verhaagen, J. Semaphorins and their receptors in olfactory axon guidance. Cell Mol. Biol. 2006, 45, 763–779. [Google Scholar]
- Lettieri, A.; Oleari, R.; Gimmelli, J.; ANDRe, V.; Cariboni, A. The role of semaphorin signaling in the etiology of hypogonadotropic hypogonadism. Minerva Endocrinol. 2016, 41, 266–278. [Google Scholar]
- Känsäkosk, J.; Fagerholm, R.; Laitinen, E.M.; Vaaralahti, K.; Hackman, P.; Pitteloud, N.; Raivio, T.; Tommiska, J. Mutation screening of SEMA3A and SEMA7A in patients with congenital hypogonadotropic hypogonadism. Pediatr. Res. 2014, 75, 641–644. [Google Scholar] [CrossRef]
- Marcos, S.; Monnier, C.; Rovira, X.; Fouveaut, C.; Pitteloud, N.; Ango, F.; Dodé, C.; Hardelin, J.-P. Defective signaling through plexin-A1 compromises the development of the peripheral olfactory system and neuroendocrine reproductive axis in mice. Hum. Mol. Genet. 2017, 26, 2006–2017. [Google Scholar] [CrossRef] [PubMed]
- Cariboni, A.; Andrews, W.D.; Memi, F.; Ypsilanti, A.R.; Zelina, P.; Chedotal, A.; Parnavelas, J.G. Slit2 and Robo3 modulate the migration of GnRH-secreting neurons. Development 2012, 139, 3326–3331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taroc, E.Z.M.; Lin, J.M.; Tulloch, A.J.; Jaworski, A.; Forni, P.E. GnRH-1 neural migration from the nose to the brain is independent from Slit2, Robo3 and NELL2 signaling. Front. Cell Neurosci. 2019, 13, 70. [Google Scholar] [CrossRef] [Green Version]
- Bless, E.P.; Walker, H.J.; Yu, K.W.; Knoll, J.G.; Moenter, S.M.; Schwarting, G.A.; Tobet, S.A. Live view of gonadotropin-releasing hormone containing neuron migration. Endocrinology 2005, 146, 463–468. [Google Scholar] [CrossRef]
- Cariboni, A.; Rakic, S.; Liapi, A.; Maggi, R.; Goffinet, A.; Parnavelas, J.G. Reelin provides an inhibitory signal in the migration of gonadotropin-releasing hormone neurons. Development 2005, 132, 4709–4718. [Google Scholar] [CrossRef] [Green Version]
- Shu, T.; Valentino, K.M.; Seaman, C.; Cooper, H.M.; Richards, L.J. Expression of the netrin-1 receptor, deleted in colorectal cancer (DCC), is largely confined to projecting neurons in the developing forebrain. J. Comp. Neurol. 2000, 406, 201–212. [Google Scholar] [CrossRef]
- Schwarting, G.A.; Raitcheva, D.; Bless, E.P.; Ackerman, S.L.; Tobet, S. Netrin 1-mediated chemoattraction regulates the migratory pathway of LHRH neurons. Eur. J. Neurosci. 2004, 19, 11–20. [Google Scholar] [CrossRef]
- Giacobini, P.; Messina, A.; Wray, S.; Giampietro, C.; Crepaldi, T.; Carmeliet, P.; Fasolo, A. Hepatocyte growth factor acts as a motogen and guidance signal for gonadotropin hormone-releasing hormone-1 neuronal migration. J. Neurosci. 2007, 10, 431–445. [Google Scholar] [CrossRef] [Green Version]
- Toba, Y.; Tiong, J.D.; Ma, Q.; Wray, S. CXCR4/SDF-1 system modulates development of GnRH-1 neurons and the olfactory system. Dev. Neurobiol. 2008, 68, 487–503. [Google Scholar] [CrossRef]
- Kawaguchi, N.; Zhang, T.T.; Nakanishi, T. Involvement of CXCR4 in normal and abnormal development. Cells 2019, 8, 185. [Google Scholar] [CrossRef] [Green Version]
- Dozio, E.; Ruscica, M.; Galliera, E.; Corsi, M.M.; Magni, P. Leptin, ciliary neurotrophic factor, leukemia inhibitory factor and interleukin-6: Class I cytokines involved in the neuroendocrine regulation of the reproductive function. Curr. Protein Pept. Sci. 2009, 10, 577–584. [Google Scholar] [CrossRef]
- Magni, P.; Dozio, E.; Ruscica, M.; Watanobe, H.; Cariboni, A.; Zaninetti, R.; Motta, M.; Maggi, R. Leukemia inhibitory factor induces the chemomigration of immortalized gonadotropin-releasing hormone neurons through the independent activation of the Janus kinase/signal transducer and activator of transcription 3, mitogen-activated protein kinase/extracellularly regulated kinase 1/2, and phosphatidylinositol 3-kinase/Akt signaling pathways. Mol. Endocrinol. 2007, 21, 1163–1174. [Google Scholar] [CrossRef] [Green Version]
- Chattopadhyay, N.; Jeong, K.H.; Yano, S.; Huang, S.; Pang, J.L.; Ren, X.; Terwilliger, E.; Kaiser, U.B.; Vassilev, P.M.; Pollak, M.R.; et al. Calcium receptor stimulates chemotaxis and secretion of MCP-1 in GnRH neurons in vitro: Potential impact on reduced GnRH neuron population in CaR-null mice. Am. J. Physiol. Endocrinol. Metab. 2007, 292, 523–532. [Google Scholar] [CrossRef] [Green Version]
- Eddie, S.L.; Childs, A.J.; Jabbour, H.N.; Anderson, R.A. Developmentally regulated IL6-type cytokines signal to germ cells in the human fetal ovary. Mol. Hum. Reprod. 2012, 18, 88–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, L.; Gearing, D.P.; White, L.S.; Compton, D.L.; Schooley, K.; Donovan, P.J. Role of leukemia inhibitory factor and its receptor in mouse primordial germ cell growth. Development 1994, 120, 3145–3153. [Google Scholar]
- Simamura, E.; Shimada, H.; Higashi, N.; Uchishiba, M.; Otani, H.; Hatta, T. Maternal leukemia inhibitory factor (LIF) promotes fetal neurogenesis via LIF-ACTH-LIF signaling relay pathway. Endocrinology 2010, 151, 1853–1862. [Google Scholar] [CrossRef]
- Dahlgren, J.; Samuelsson, A.M.; Jansson, T.; Holmäng, A. Interleukin-6 in the maternal circulation reaches the rat fetus in mid-gestation. Pediatr. Res. 2006, 60, 147–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Wolfe, A. Signaling of cytokines is important in regulation of GnRH neurons. Mol. Neurobiol. 2012, 45, 119–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bless, E.P.; Westaway, W.A.; Schwarting, G.A.; Tobet, S.A. Effects of gamma-aminobutyric acid (A) receptor manipulation on migrating gonadotropin-releasing hormone neurons through the entire migratory route in vivo and in vitro. Endocrinology 2000, 141, 1254–1262. [Google Scholar] [CrossRef]
- Liu, X.; Porteous, R.; Herbison, A.E. Dynamics of GnRH neuron ionotropic GABA and glutamate synaptic receptors are unchanged during estrogen positive and negative feedback in female mice. eNeuro 2017, 4, 29109970. [Google Scholar] [CrossRef] [Green Version]
- Vastagh, C.; Schwirtlich, M.; Kwakowsky, A.; Erdélyi, F.; Margolis, F.L.; Yanagawa, Y.; Katarova, Z.; Szabó, G. The spatiotemporal segregation of GAD forms defines distinct GABA signaling functions in the developing mouse olfactory system and provides novel insights into the origin and migration of GnRH neurons. Dev. Neurobiol. 2015, 75, 249–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izvolskaia, M.; Duittoz, A.H.; Tillet, Y.; Ugrumov, M.V. The influence of catecholamine on the migration of gonadotropin-releasing hormone-producing neurons in the rat foetuses. Brain Struct. Funct. 2009, 938, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Melnikova, V.I.; Afanasyeva, M.A.; Voronova, S.N.; Zakharova, L.A. The effect of catecholamine deficit on the development of the immune system in rats. Dokl. Biol. Sci. 2012, 443, 68–70. [Google Scholar] [CrossRef]
- Lifantseva, N.V.; Koneeva, T.O.; Voronova, S.N.; Zakharova, L.A.; Melnikova, V.I. The inhibition of dopamine synthesis in fetuses changes the pattern of T-lymphocyte maturation in the thymus of adult rats. Dokl. Biochem. Biophys. 2016, 470, 342–344. [Google Scholar] [CrossRef]
- Forni, P.E.; Taylor-Burds, C.; Melvin, V.S.; Williams, T.; Wray, S. Neural crest and ectodermal cells intermix in the nasal placode to give rise to GnRH-1 neurons, sensory neurons, and olfactory ensheathing cells. J. Neurosci. 2011, 31, 6915–6927. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.Y.; Cheng, A.M.S.; Zhang, Y.; Zhu, Y.T.; He, H.; Mahabole, M.; Tseng, S.C.G. Pax 6 controls neural crest potential of limbal niche cells to support self-renewal of limbal epithelial stem cells. Sci. Rep. 2019, 9, 9763. [Google Scholar] [CrossRef]
- Ikeda, K.; Watanabe, Y.; Ohto, H.; Kawakami, K. Molecular interaction and synergistic activation of a promoter by Six, Eya, and Dach proteins mediated through CREB binding protein. Mol. Cell. Biol. 2002, 22, 6759–6766. [Google Scholar] [CrossRef] [Green Version]
- Abitua, P.B.; Gainous, T.B.; Kaczmarczyk, A.N.; Winchell, C.J.; Hudson, C.; Kamata, K.; Nakagawa, M.; Tsuda, M.; Kusakabe, T.G.; Levine, M. The pre-vertebrate origins of neurogenic placodes. Nature 2015, 524, 462–465. [Google Scholar] [CrossRef]
- Larder, R.; Clark, D.D.; Miller, N.L.G.; Mellon, P.M. Hypothalamic dysregulation and infertility in mice lacking the homeodomain protein Six 6. J. Neurosci. 2011, 31, 426–438. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, K.; Ookawara, S.; Sato, S.; Ando, Z.; Kageyama, R.; Kawakami, K. Six1 is essential for early neurogenesis in the development of olfactory epithelium. Dev. Biol. 2007, 311, 53–68. [Google Scholar] [CrossRef] [Green Version]
- Simeone, A.; Acampora, D. The role of Otx2 in organizing the anterior patterning in mouse. Int. J. Dev.Biol. 2001, 45, 337–345. [Google Scholar] [PubMed]
- Hoffmann, H.M.; Larder, R.; Lee, J.S.; Hu, R.J.; Trang, C.; Devries, B.M.; Clark, D.D.; Mellon, P.L. Differential CRE Expression in Lhrh-cre and GnRH-cre Alleles and the Impact on Fertility in Otx2-Flox Mice. Neuroendocrinology 2019, 108, 328–342. [Google Scholar] [CrossRef] [PubMed]
- Quaynor, S.D.; Bosley, M.E.; Duckworth, C.G.; Porter, K.R.; Kim, S.H.; Kim, H.G.; Chorich, L.P.; Sullivan, M.E.; Choi, J.H.; Cameron, R.S.; et al. Targeted next generation sequencing approach identifies eighteen new candidate genes in normosmic hypogonadotropic hypogonadism and Kallmann syndrome. Mol Cell Endocrinol. 2016, 437, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Taroc, E.Z.M.; Naik, A.S.; Lin, J.M.; Peterson, N.B.; Keefe, D.L., Jr.; Genis, L.; Fuchs, G.; Balasubramanian, R.; Forni, P.E. Gli3 Regulates Vomeronasal Neurogenesis, Olfactory Ensheathing Cell Formation, and GnRH-1 Neuronal Migration. J. Neurosci. 2020, 40, 311–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corradi, A.; Croci, L.; Broccoli, V.; Zecchini, S.; Previtali, S.; Wurst, W.; Amadio, S.; Maggi, R.; Quattrini, A.; Consalez, G.G. Hypogonadotropic hypogonadism and peripheral neuropathy in Ebf2-null mice. Development 2003, 130, 401–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trarbach, E.B.; Baptista, M.T.; Garmes, H.M.; Hackel, C. Molecular analysis of KAL-1, GnRH-R, NELF and EBF2 genes in a series of Kallmann syndrome and normosmic hypogonadotropic hypogonadism patients. J. Endocrinol. 2005, 187, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Cogliati, T.; Delgado-Romero, P.; Norwitz, E.R.; Guduric-Fuchs, J.; Kaiser, U.B.; Wray, S.; Kirsch, I.R. Pubertal impairment in Nhlh2 null mice is associated with hypothalamic and pituitary deficiencies. Mol. Endocrinol. 2007, 21, 3013–3027. [Google Scholar] [CrossRef]
- Schmid, T.; Boehm, U.; Braun, T. GnRH neurogenesis depends on embryonic pheromone receptor expression. Mol. Cell. Endocrinol. 2020, 518, 111030. [Google Scholar] [CrossRef]
- Taglialatela, P.; Soria, J.M.; Caironi, V.; Moiana, A.; Bertuzzi, S. Compromised generation of GABAergic interneurons in the brains of Vax1−/− mice. Development 2004, 131, 4239–4249. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, H.M.; Mellon, P.L. A small population of hypothalamic neurons govern fertility: The critical role of VAX1 in GnRH neuron development and fertility maintenance. Neurosci. Commun. (Houst.) 2016, 2, e1373. [Google Scholar]
- Cau, E.; Casarosa, S.; Guillemot, F. Mash1 and Ngn1 control distinct steps of determination and differentiation in the olfactory sensory neuron lineage. Development 2002, 129, 1871–1880. [Google Scholar] [PubMed]
- Kramer, P.K.; Guerrero, G.; Krishnamurthy, R.; Mitchel, P.J.; Wray, S. Ectopic expression of LHRH and periferin in the respiratory epithelium of mice lacing transcriptional factor AP-2. Mech. Dev. 2000, 94, 79–94. [Google Scholar] [CrossRef]
- Leclerc, G.M.; Bose, S.K.; Boockfor, F.R. Specific GATA-binding elements in the GnRH promoter are required for gene expression pulse activity: Role of GATA-4 and GATA-5 in this intermittent process. Neuroendocrinology 2008, 88, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, R.; Crowley, W.F., Jr. Isolated GnRH deficiency: A disease model serving as a unique prism into the systems biology of the GnRH neuronal network. Mol. Cell. Endocrinol. 2011, 346, 4–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.Q.; Li, X.F.; Ye, B.; Popat, N.; Milligan, S.R.; Lightman, S.L.; O’Byrne, K.T. Neonatal programming by immunological challenge: Effects on ovarian function in the adult rat. Reproduction 2011, 41, 241–248. [Google Scholar] [CrossRef] [Green Version]
- Sharova, V.S.; Izvol’skaia, M.S.; Voronova, S.N.; Zakharova, L.A. Effect of bacterial endotoxin on migration of gonadotropin-releasing, hormone producing neurons in rat embryogenesis. Ontogenez 2011, 42, 439–446. [Google Scholar] [CrossRef]
- Cadman, S.M.; Kim, S.H.; Hu, Y.; González-Martínez, D.; Bouloux, P.M. Molecular pathogenesis of Kallmann’s syndrome. Horm. Res. 2007, 67, 231–242. [Google Scholar] [CrossRef] [Green Version]
- Gianetti, E.; Tusset, C.; Noel, S.D.; Au, M.G.; Dwyer, A.A.; Hughes, V.A.; Crowley, W.F.; Kaiser, U.B.; Latronico, A.C.; Seminara, S.B. TAC3/TACR3 mutations reveal preferential activation of gonadotropin-releasing hormone release by neurokinin B in neonatal life followed by reversal in adulthood. J. Clin. Endocrinol. Metab. 2010, 95, 2857–2867. [Google Scholar] [CrossRef] [Green Version]
- Shaw, N.D.; Seminara, S.B.; Welt, C.K.; Au, M.G.; Plummer, L.; Hughes, V.A.; Dwyer, A.A.; Martin, K.A.; Quinton, R.; Mericq, V.; et al. Expanding the phenotype and genotype of female GnRH deficiency. J. Clin. Endocrinol. Metab. 2011, 96, 566–576. [Google Scholar] [CrossRef]
- Drummond, E.S.; Martins, R.N.; Handelsman, D.J.; Harvey, A.R. Altered expression of Alzheimer’s disease-related proteins in male hypogonadal mice. Endocrinology 2012, 153, 2789–2799. [Google Scholar] [CrossRef] [Green Version]
- Lillycrop, K.A.; Slater-Jefferies, J.L.; Hanson, M.A.; Godfrey, K.M.; Jackson, A.A.; Burdge, G.C. Induction of altered epigenetic regulation of the hepatic glucocorticoid receptor in the offspring of rats fed a protein-restricted diet during pregnancy suggests that reduced DNA methyltransferase-1 expression is involved in impaired DNA methylation and changes in histone modifications. Br. J. Nutr. 2007, 97, 1064–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aiken, C.E.; Ozann, S.E. Sex differences in developmental programming models. Reproduction 2013, 145, R1–R13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cameron, N.M.; Shahrokh, D.; Del Corpo, A.; Dhir, S.K.; Szyf, M.; Champagne, F.A.; Meaney, M.J. Epigenetic programming of phenotypic variations in reproductive strategies in the rat through maternal care. J. Neuroendocrinol. 2008, 20, 795–801. [Google Scholar] [CrossRef] [PubMed]
- Champagne, F.A.; Curley, J.P. Maternal regulation of estrogen receptor alpha methylation. Curr. Opin. Pharmacol. 2008, 8, 735–739. [Google Scholar] [CrossRef] [Green Version]
- Tomikawa, J.; Uenoyama, Y.; Ozawa, M.; Fukanuma, T.; Takase, K.; Goto, T.; Okamura, H.; Maeda, K.; Tsukamura, H. Epigenetic regulation of Kiss1 gene expression mediating estrogen-positive feedback action in the mouse brain. Proc. Natl. Acad. Sci. USA 2012, 109, E1294–E1301. [Google Scholar] [CrossRef] [Green Version]
- Knox, A.M.; Li, X.F.; Kinsey-Jones, J.S.; Wilkinson, E.S.; Wu, X.Q.; Cheng, Y.S.; Milligan, S.R.; Lightman, S.L.; O’Byrne, K.T. Neonatal lipopolysaccharide exposure delays puberty and alters hypothalamic Kiss1 and Kiss1r mRNA expression in the female rat. J. Neuroendocrinol. 2009, 21, 683–692. [Google Scholar] [CrossRef] [Green Version]
- Bryan, E.R.; Kim, J.; Beagley, K.W.; Carey, A.J. Testicular inflammation and infertility: Could chlamydial infections be contributing? Am. J. Reprod. Immunol. 2020, 84, e13286. [Google Scholar] [CrossRef]
- Cai, Z.; Pan, Z.L.; Pang, Y.; Evans, O.B.; Rhodes, P.G. Cytokine induction in fetal rat brains and brain injury in neonatal rats after maternal lipopolysaccharide administration. Pediatr. Res. 2000, 47, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Gilstrap, L.C.; Ramin, S.M. Urinary tract infections during pregnancy. Obstet. Gynecol. Clin. N. Am. 2001, 28, 581–591. [Google Scholar] [CrossRef]
- Hagberg, H.; Gressens, P.; Mallard, C. Inflammation during fetal and neonatal life: Implications for neurologic and neuropsychiatric disease in children and adults. Ann. Neurol. 2012, 71, 444–457. [Google Scholar] [CrossRef]
- Wang, H.L.; Pei, D.E.; Yang, R.D.; Wan, C.L.; Ye, Y.M.; Peng, S.S.; Zeng, Q.Q.; Yu, Y. Prenatal maternal vaginal inflammation increases anxiety and alters HPA axis signalling in adult male mice. Int. J. Dev. Neurosci. 2019, 75, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Herman, A.P.; Tomaszewska-Zaremba, D. Effect of endotoxin on the expression of GnRH and GnRHR genes in the hypothalamus and anterior pituitary gland of anestrous ewes. Anim. Reprod. Sci. 2010, 120, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Watanobe, H.; Hayakawa, Y. Hypothalamic Interleukin-1 and Tumor Necrosis Factor, but not Interleukin-6, mediate the endotoxin-induced suppression of the reproductive axis in rats. Endocrinology 2003, 144, 4868–4875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurewicz, A.; Matysiak, M.; Tybor, K.; Kilianek, L.; Raine, C.S.; Selmaj, K. Tumour necrosis factor-induced death of adult human oligodendrocytes is mediated by apoptosis inducing factor. Brain 2005, 128, 2675–2688. [Google Scholar] [CrossRef] [PubMed]
- Kothari, N.; Bogra, J.; Abbas, H.; Kohli, M.; Malik, A.; Kothari, D.; Srivastav, S.; Singh, P.K. Tumor necrosis factor gene polymorphism results in high TNF level in sepsis and septic shock. Cytokine 2013, 61, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Ginsberg, Y.; Khatib, N.; Weiner, Z.; Beloosesky, R. Maternal inflammation, fetal brain implications and suggested neuroprotection: A summary of 10 years of research in animal models. Rambam Maimonides Med. J. 2017, 8, e0028. [Google Scholar] [CrossRef]
- Kalra, P.S.; Edwards, T.G.; Xu, B.; Jain, M.; Kalra, S.P. The anti-gonadotropic effects of cytokines: The role of neuropeptides. Domes. Anim. Endocrinol. 1998, 15, 321–332. [Google Scholar] [CrossRef]
- Nouel, D.; Burt, M.; Zhang, Y.; Harvey, L.; Boksa, P. Prenatal exposure to bacterial endotoxin reduces the number of GAD67- and reelin-immunoreactive neurons in the hippocampus of rat offspring. Eur. Neuropsychopharmacol. 2012, 22, 300–307. [Google Scholar] [CrossRef]
- Harvey, L.; Boksa, P. A stereological comparison of GAD67 and reelin expression in the hippocampal stratum oriens of offspring from two mouse models of maternal inflammation during pregnancy. Neuropharmacology 2012, 62, 1767–1776. [Google Scholar] [CrossRef]
- Morale, M.C.; Gallo, F.; Tirolo, C.; L’Episcopo, F.; Gennuso, F.; Testa, N.; Caniglia, S.; Spina-Purrello, V.; Avola, R.; Scoto, G.M.; et al. The reproductive system at the neuroendocrine-immune interface: Focus on LHRH, estrogens and growth factors in LHRH neuron-glial interactions. Domest. Anim. Endocrinol. 2003, 25, 21–46. [Google Scholar] [CrossRef]
- Coe, C.L.; Lubach, G.R. Prenatal origins of individual variation in behavior and immunity. Neurosci. Biobehav. Rev. 2005, 29, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Van den Bergh, B.R.H.; van den Heuvel, M.I.; Lahti, M.; Braeken, M.; de Rooij, S.R.; Entringer, S.; Hoyer, D.; Roseboom, T.; Räikkönen, K.; King, S.; et al. Prenatal developmental origins of behavior and mental health: The influence of maternal stress in pregnancy. Neurosci. Biobehav. Rev. 2020, 117, 26–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakharova, L.A.; Malyukova, I.V.; Proshlyakova, E.V.; Sapronova, A.Y.; Ugrumov, M.V. Hypothalamo-pituitary control of the cell-mediated immunity in rat embryos: Role of LHRH in regulation of lymphocyte proliferation. J. Reprod. Immunol. 2000, 47, 17–32. [Google Scholar] [CrossRef]
- Mann, D.R.; Akinbami, M.A.; Lunn, S.F.; Fraser, H.M.; Gould, K.G.; Ansari, A.A. Endocrine-immune interaction: Alteractions in immune function resulting from neonatal treatment with a GnRH antagonist and seasonality in male primates. Am. J. Reprod. Immunol. 2000, 44, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Dygalo, N.N.; Shemenkova, T.V.; Kalinina, T.S.; Shishkina, G.T. A critical point of male gonad development: Neuroendocrine correlates of accelerated testicular growth in rats during early life. PLoS ONE 2014, 9, e93007. [Google Scholar] [CrossRef]
- Rao, L.V.; Cleveland, R.P.; Kimmel, R.J.; Ataya, K.M. Hematopoietic stem cell antigen-1 (Sca-1) expression in different lymphoid tissues of female mice treated with GnRH agonist. Am. J. Reprod. Immunol. 1995, 34, 257–266. [Google Scholar] [CrossRef]
- Ullewar, M.P.; Umathe, S.N. Gonadotropin-releasing hormone agonist selectively augments thymopoiesis and prevents cell apoptosis in LPS induced thymic atrophy model independent of gonadal steroids. Int. Immunopharmacol. 2014, 23, 46–53. [Google Scholar] [CrossRef]
- Jacobson, J.D.; Ansari, M.A.; Mansfield, M.E.; McArthur, C.P.; Clement, L. T Gonadotropin-releasing hormone increases CD4 T-lymphocyte numbers in an animal model of immunodeficiency. J. Allergy Clin. Immunol. 1999, 104, 653–658. [Google Scholar] [CrossRef]
- Yarilin, A.A.; Belyakov, I.M. Cytokines in the thymus: Production and biological effects. Curr. Med. Chem. 2004, 11, 447–464. [Google Scholar] [CrossRef]
- Beutler, B.A. The role of tumor necrosis factor in health and disease. J. Rheumatol. Suppl. 1999, 57, 16–21. [Google Scholar]
- Metcalf, D. The unsolved enigmas of leukemia inhibitory factor. Stem Cells. 2003, 21, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Batticane, N.; Morale, M.C.; Gallo, F.; Farinella, Z.; Marchetti, B. Luteinizing hormone-releasing hormone signaling at the lymphocyte involves stimulation of interleukin-2 receptor expression. Endocrinology 1991, 129, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Tanriverdi, F.; Gonzalez-Martinez, D.; Hu, Y.; Kelestimur, F.P.; Bouloux, M.G. GnRH-I and GnRH-II have differential modulatory effects on human peripheral blood mononuclear cell proliferation and interleukin-2 receptor γ-chain mRNA expression in healthy males. Clin. Exp. Immunol. 2005, 142, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Tanriverdi, F.; Gonzalez-Martinez, D.; Silveira, L.F.; Hu, Y.; Maccoll, G.S.; Travers, P.; Bouloux, M.G. Expression of gonadotropin-releasing hormone type-I (GnRH-I) and type-II (GnRH-II) in human peripheral blood mononuclear cells (PMBCs) and regulation of B-lymphoblastoid cell proliferation by GnRH-I and GnRH-II. Exp. Clin. Endocrinol. Diabetes 2004, 112, 587–594. [Google Scholar] [CrossRef]
- Montgomery, R.A.; Dallman, M.J. Analysis of cytokine gene expression during fetal thymic ontogeny using the polymerase chain reaction. J. Immunol. 1991, 147, 554–560. [Google Scholar]
- Rouiller-Fabre, V.; Levacher, C.; Pairault, C.; Racine, C.; Moreau, E.; Olaso, R.; Livera, G.; Migrenne, S.; Delbes, G.; Habert, R. Development of the foetal and neonatal testis. Andrologia 2003, 35, 79–83. [Google Scholar] [CrossRef]
- Azad, N.; LaPaglia, N.; Agrawal, L.; Steiner, J.; Uddin, S.; Williams, D.W.; Lawrence, A.M.; Emanuele, N.V. The role of gonadectomy and testosterone replacement on thymic luteinizing hormone-releasing hormone production. J. Endocrinol. 1998, 158, 229–235. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, J.D.; Ansari, M.A. Immunomodulatory actions of gonadal steroids may be mediated by gonadotropin-releasing hormone. Endocrinology 2004, 145, 330–336. [Google Scholar] [CrossRef] [Green Version]
- Zandman-Goddard, G.; Peeva, E.; Shoenfeld, Y. Gender and autoimmunity. Autoimmun. Rev. 2007, 6, 366–372. [Google Scholar] [CrossRef]
- Jacobson, J.D. Gonadotropin-releasing hormone and G proteins: Potential roles in autoimmunity. Ann. N. Y. Acad. Sci. 2000, 917, 809–818. [Google Scholar] [CrossRef]
- Andersson, A.M.; Jørgensen, N.; Main, K.M.; Toppari, J.; Rajpert-De Meyts, E.; Leffers, H.; Skakkebaek, N.E. Adverse trends in male reproductive health: We may have reached a crucial ‘tip- ping point’. Int. J. Androl. 2008, 31, 74–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glaser, A.P.; Schaeffer, A.J. Urinary Tract Infection and Bacteriuria in Pregnancy. Urol. Clin. N. Am. 2015, 42, 547–560. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.M.; Crowther, C.A.; Wilkinson, D.; Bain, E. Magnesium sulphate for women at term for neuroprotection of the fetus. Cochrane Database Syst. Rev. 2013, 2, CD009395. [Google Scholar] [CrossRef] [PubMed]
- Beloosesky, R.; Khatib, N.; Ginsberg, Y.; Anabosy, S.; Shalom-Paz, E.; Dahis, M.; Ross, M.G.; Weiner, Z. Maternal magnesium sulfate fetal neuroprotective effects to the fetus: Inhibition of neuronal nitric oxide synthase and nuclear factor kappa-light-chain-enhancer of activated B cells activation inthom. Am. J. Obstet. Gynecol. 2016, 215, 382e1–382e6. [Google Scholar] [CrossRef]
- Sharabi, H.; Khatib, N.; Ginsberg, Y.; Weiner, Z.; Ross, M.G.; Tamar, B.K.; Efrat, S.; Mordechai, H.; Beloosesky, R. Therapeutic N-Acetyl-Cysteine (Nac) Following Initiation of Maternal Inflammation Attenuates Long-Term Offspring Cerebral Injury, as Evident in Magnetic Resonance Imaging (MRI). Neuroscience 2019, 403, 118–124. [Google Scholar] [CrossRef]
- Lappas, M.; Permezel, M.; Rice, G.E. N-acetyl-cysteine inhibits phospholipid metabolism, proinflammatory cytokine release, protease activity, and nuclear factor-kappaB deoxyribonucleic acid-binding activity in human fetal membranes in vitro. J. Clin. Endocrinol. Metab. 2003, 88, 1723–1729. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Hu, Y.F.; Hao, J.H.; Chen, Y.H.; Su, P.Y.; Wang, Y.; Yu, Z.; Fu, L.; Xu, Y.Y.; Zhang, C.; et al. Maternal zinc deficiency during pregnancy elevates the risks of fetal growth restriction: A population-based birth cohort study. Sci. Rep. 2015, 5, 11262. [Google Scholar] [CrossRef] [Green Version]
- Chua, J.S.; Cowley, C.J.; Manavis, J.; Rofe, A.M.; Coyle, P. Prenatal exposure to lipopolysaccharide results in neurodevelopmental damage that is ameliorated by zinc in mice. Brain Behav. Immun. 2012, 26, 326–336. [Google Scholar] [CrossRef]
- Labrousse, V.F.; Leyrolle, Q.; Amadieu, C.; Aubert, A.; Sere, A.; Coutureau, E.; Grégoire, S.; Bretillon, L.; Pallet, V.; Gressens, P.; et al. Dietary omega-3 deficiency exacerbates inflammation and reveals spatial memory deficits in mice exposed to lipopolysaccharide during gestation. Brain Behav. Immun. 2018, 7, 427–440. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Chen, Y.H.; Fu, L.; Yu, Z.; Xia, M.Z.; Hu, X.G.; Wang, H.; Xu, D.X. Vitamin D3 pretreatment protects against lipopolysaccharide-induced early embryo loss through its anti-inflammatory effects. Am. J. Reprod. Immunol. 2017, 77. [Google Scholar] [CrossRef]
- Maugeri, A.; Barchitta, M.; Blanco, I.; Agodi, A. Effects of Vitamin D Supplementation During Pregnancy on Birth Size: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutrients 2019, 11, 442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langley-Evans, S.C. Developmental programming of health and disease. Proc. Nutr. Soc. 2006, 65, 97–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, S.S.; Zhou, Y.M.; Li, D.; Ma, Q. Early Infant Exposure to Excess Multivitamin: A Risk Factor for Autism? Autism Res Treat. 2013, 2913, 963697. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, M.; Laskou, F.; Stapleton, P.P.; Hadavi, S.; Dasgupta, B. Tocilizumab (Actemra). Hum. Vaccin. Immunother. 2017, 13, 1972–1988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Medicines Agency. 1st Edition. London, UK, 2009. Available online: https://www.ema.europa.eu/en/documents/assessment-report/invokana-epar-public-assessment-report_en.pdf (accessed on 3 January 2017).
- Smith, S.E.; Li, J.; Garbett, K.; Mirnics, K.; Patterson, P.H. Maternal immune activation alters fetal brain development through interleukin-6. J. Neurosci. 2007, 27, 10695–10702. [Google Scholar] [CrossRef] [Green Version]
- Domínguez-Soto, Á.; Simón-Fuentes, M.; de Las Casas-Engel, M.; Cuevas, V.D.; López-Bravo, M.; Domínguez-Andrés, J.; Saz-Leal, P.; Sancho, D.; Ardavín, C.; Ochoa-Grullón, J.; et al. IVIg promote cross-tolerance against inflammatory stimuli in vitro and in vivo. J. Immunol. 2018, 201, 41–52. [Google Scholar] [CrossRef]
- Sawa, T.; Kinoshita, M.; Inoue, K.; Ohara, J.; Moriyama, K. Immunoglobulin for treating bacterial infections: One more mechanism of action. Antibodies 2019, 8, 52. [Google Scholar] [CrossRef] [Green Version]
- Inci, A.; Űnal, D.S.; Őzeş, N.O.; Erin, N.; Akçakuş, M.; Oygűr, N. The efficacy of intravenous immunoglobulin on lipopolysaccharide-induced fetal brain inflammation in preterm rats. Am. J. Obstet. Gynecol. 2013, 209, 1–8. [Google Scholar] [CrossRef]
- Kyvelidou, C.; Sotiriou, D.; Zerva, I.; Athanassakis, I. Protection against lipopolysaccharide-induced immunosuppression by IgG and IgM. Shock 2018, 49, 474–482. [Google Scholar] [CrossRef]
- Chaigne, B.; Mouthon, L. Mechanisms of action of intravenous immunoglobulin. Transfus. Apher. Sci. 2017, 56, 45–49. [Google Scholar] [CrossRef]
- Murakami, K.; Suzuki, C.; Kobayashi, F.; Nakano, A.; Fujii, A.; Sakai, K.; Imada, T. Intravenous immunoglobulin preparation attenuates LPS-induced production of pro-inflammatory cytokines in human monocytic cells by modulating TLR4-mediated signaling pathways. Naunyn Schmiedebergs Arch. Pharmacol. 2012, 385, 891–898. [Google Scholar] [CrossRef] [PubMed]
- Anthoney, N.; Foldi, I.; Hidalgo, A. Toll and Toll-like receptor signalling in development. Development 2018, 145, dev156018. [Google Scholar] [CrossRef] [PubMed] [Green Version]




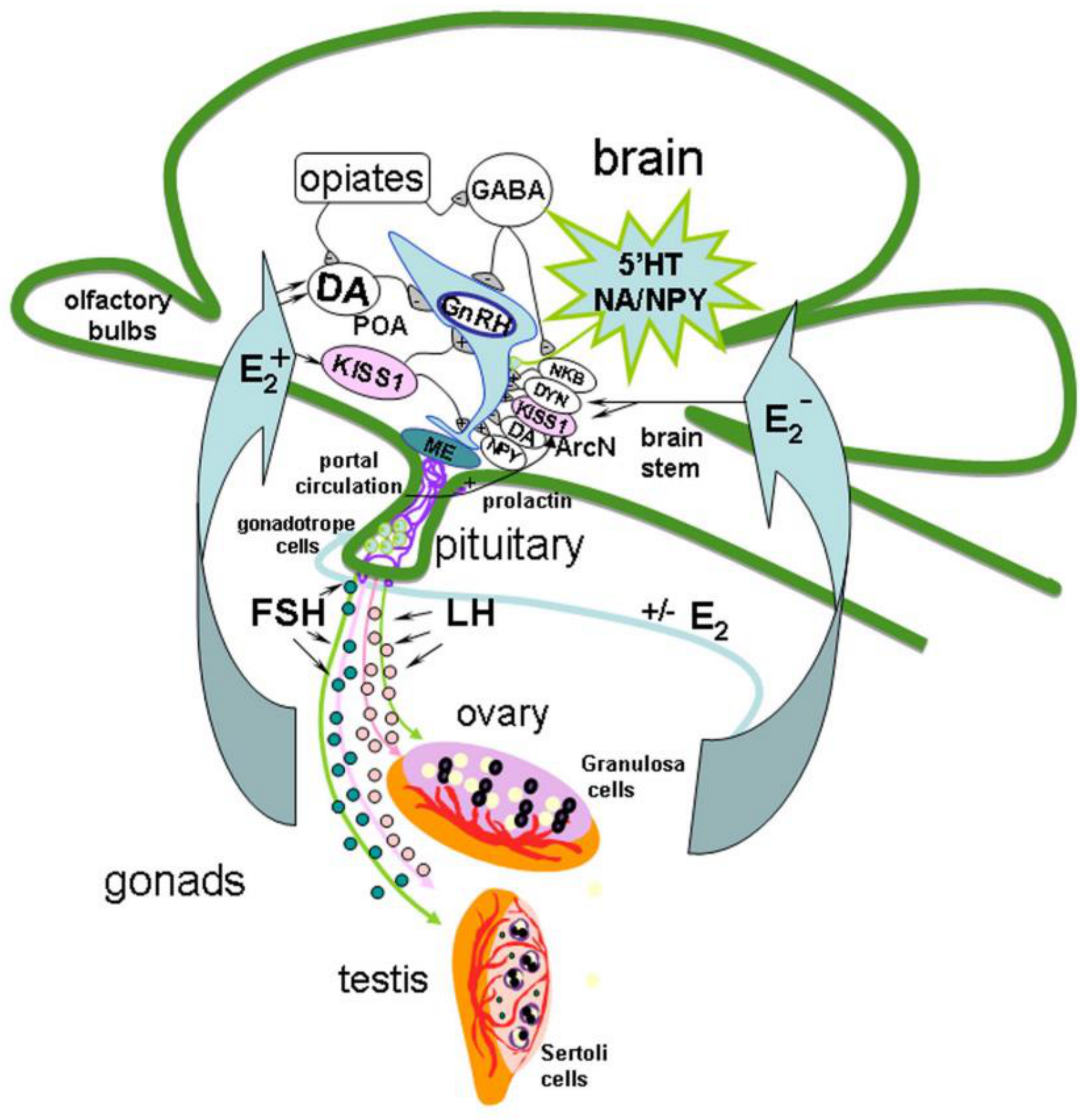{kind=link}
{kind=link}
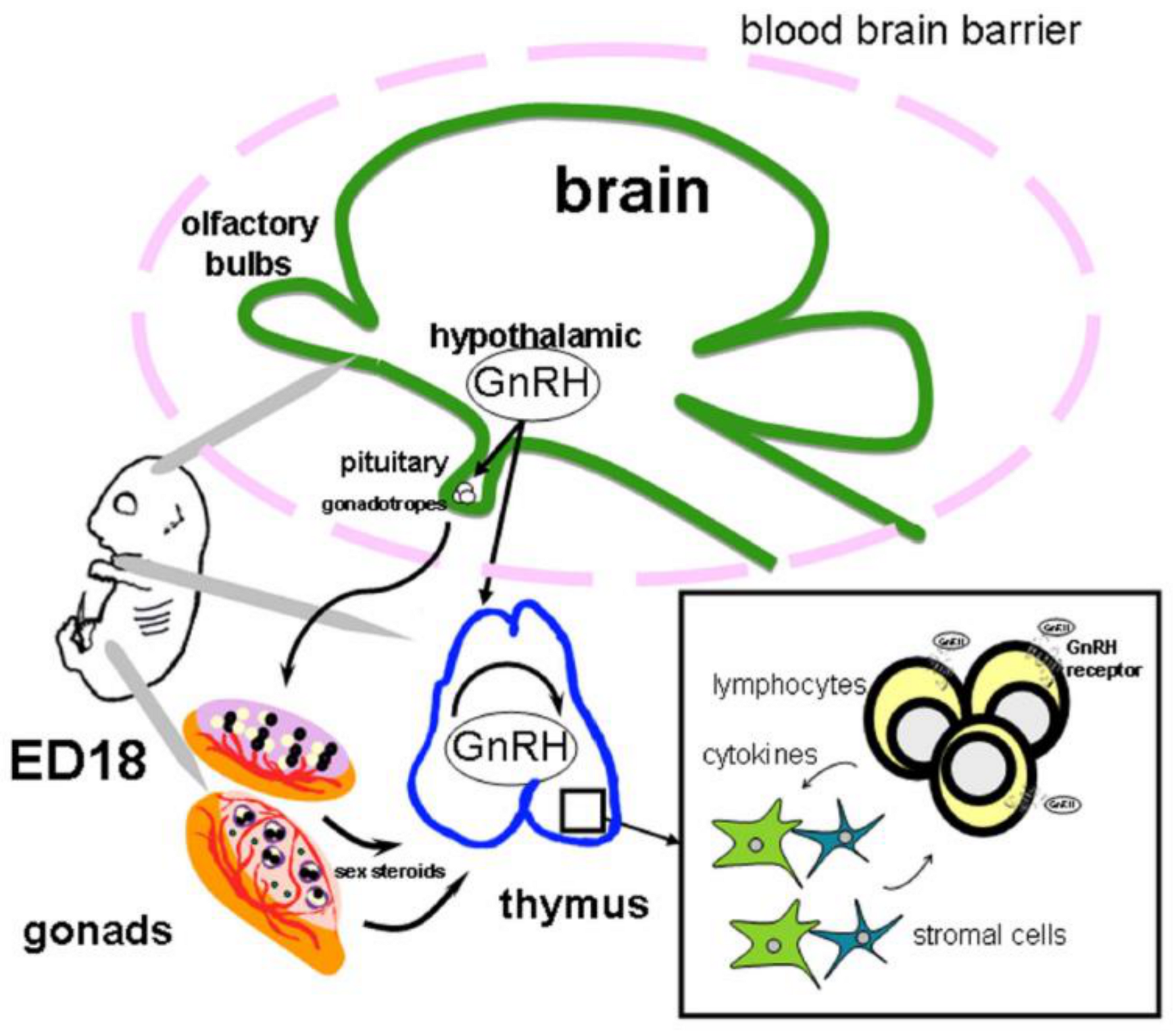{kind=link}
| Transcriptional Factor | General Action | Action on GnRH | References |
|---|---|---|---|
| Pax-6,7 | The role of Pax-6 in eye and nasal development | Its expression appears in the olfactory ensheathing cells and vomeronasal organ. PAX6 and AP-2 define distinct progenitor cells mixed within the developing nasal placode | [96,97] |
| Eya-1,2 | The role in developmental process conserved across vertebrates | It is initially expressed in a row of eight cells located immediately anterior to the neural plate | [98,99] |
| Six-1,3,6 | Six is restricted to the developing eye and brain | Mice lacking Six1 (Six1−/−) exhibited defective early neurogenesis in the olfactory epithelium. Loss of Six6 expression disrupts male fertility because of decreased follicle-stimulating hormone release | [99,100,101] |
| OTX-1,2 | Involvement in the early development of CNS | The role in the initiation and guidance of directed migration. In mice with deletion of the transcription factor Otx2 found fewer GnRH neurons | [102,103] |
| Gli3 | It is necessary for the development of olfactory system | Regulates vomeronasal neurogenesis, olfactory ensheathing cell formation and GnRH neuronal migration | [104,105] |
| Ebf2 | Involvement in numerous developmental processes, ranging from B-cell development to neuronal differentiation | It is expressed in migrating GnRH neurons on embryonic day 11 (ED11). Mice with disrupted expression of Ebf2 retained GnRH neurons clustered in the nasal mesenchyme | [106,107] |
| Nhlh2 | Its expression in regions of the hypothalamus as well as the pituitary | Mice with disrupted expression of Nhlh2 had a loss of GnRH neurons in adulthood that occurred sometime between birth and adulthood (loss of 60% in females and 30% in males) | [108,109] |
| VAX1 | It is essential for the formation of the eye, ventral forebrain and palate | GnRH staining in Vax1null mice show a total absence of GnRH expression in the adult. Using the immortalized model GnRH neuron cell lines, GN11 and GT1-7, it was show that VAX1 is a direct regulator of GnRH transcription by binding key ATTA sites within the GnRH promoter. | [110,111] |
| Mash-1, Math4A, Math4/neurogenin1, NeuroD | Involvement in gliogenesis and neurogenesis | Differentiation into the olfactory epithelium | [101,112] |
| Olf-1, GATA-4 | Involvement in brain development | Its expression in the olfactory epithelium Its expression is necessary for GnRH pulse activity | [113,114] |
| AP-2α | Involvement in craniofacial morphogenesis | Its expression only in the respiratory epithelium. It prevents recapitulation of developmental programs within the respiratory epithelium that lead to expression of GnRH and peripherin phenotypes | [113] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zakharova, L.; Sharova, V.; Izvolskaia, M. Mechanisms of Reciprocal Regulation of Gonadotropin-Releasing Hormone (GnRH)-Producing and Immune Systems: The Role of GnRH, Cytokines and Their Receptors in Early Ontogenesis in Normal and Pathological Conditions. Int. J. Mol. Sci. 2021, 22, 114. https://doi.org/10.3390/ijms22010114
Zakharova L, Sharova V, Izvolskaia M. Mechanisms of Reciprocal Regulation of Gonadotropin-Releasing Hormone (GnRH)-Producing and Immune Systems: The Role of GnRH, Cytokines and Their Receptors in Early Ontogenesis in Normal and Pathological Conditions. International Journal of Molecular Sciences. 2021; 22(1):114. https://doi.org/10.3390/ijms22010114
Chicago/Turabian StyleZakharova, Liudmila, Viktoria Sharova, and Marina Izvolskaia. 2021. "Mechanisms of Reciprocal Regulation of Gonadotropin-Releasing Hormone (GnRH)-Producing and Immune Systems: The Role of GnRH, Cytokines and Their Receptors in Early Ontogenesis in Normal and Pathological Conditions" International Journal of Molecular Sciences 22, no. 1: 114. https://doi.org/10.3390/ijms22010114
APA StyleZakharova, L., Sharova, V., & Izvolskaia, M. (2021). Mechanisms of Reciprocal Regulation of Gonadotropin-Releasing Hormone (GnRH)-Producing and Immune Systems: The Role of GnRH, Cytokines and Their Receptors in Early Ontogenesis in Normal and Pathological Conditions. International Journal of Molecular Sciences, 22(1), 114. https://doi.org/10.3390/ijms22010114