Dose-Dependent Effects of Resveratrol on Cisplatin-Induced Hearing Loss


Abstract
:1. Introduction
2. Results
2.1. ABR Threshold Shift Following CXP and/or RV Administration
2.2. Cochlear Outer Hair Cell Injury
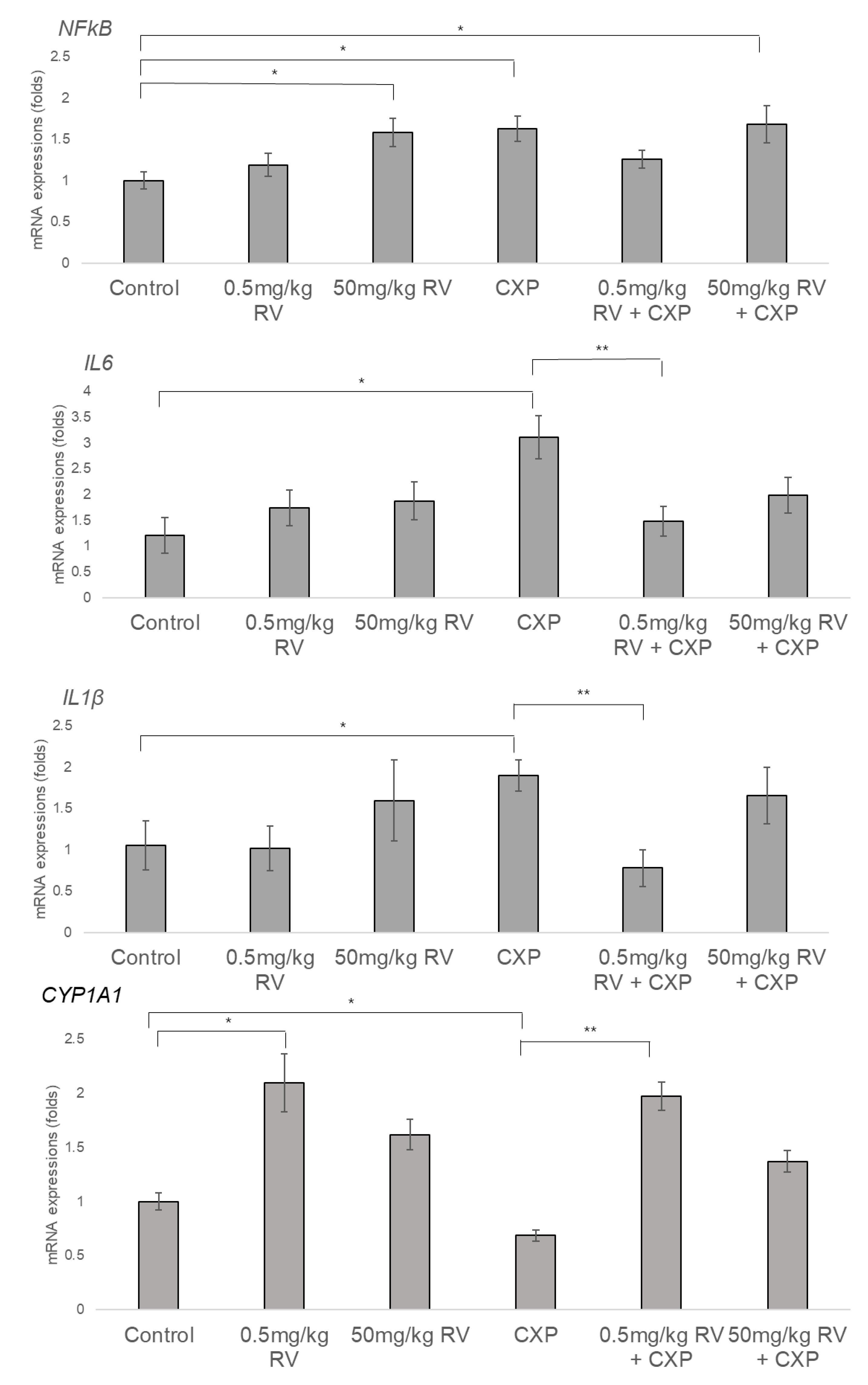
2.3. Increased NFκB, IL1β, and IL6 and Decreased CYP1A1 Levels in CXP-Induced Hearing Loss Rats and Partial Reversal of Expression Patterns Following 0.5 mg/kg RV Treatment
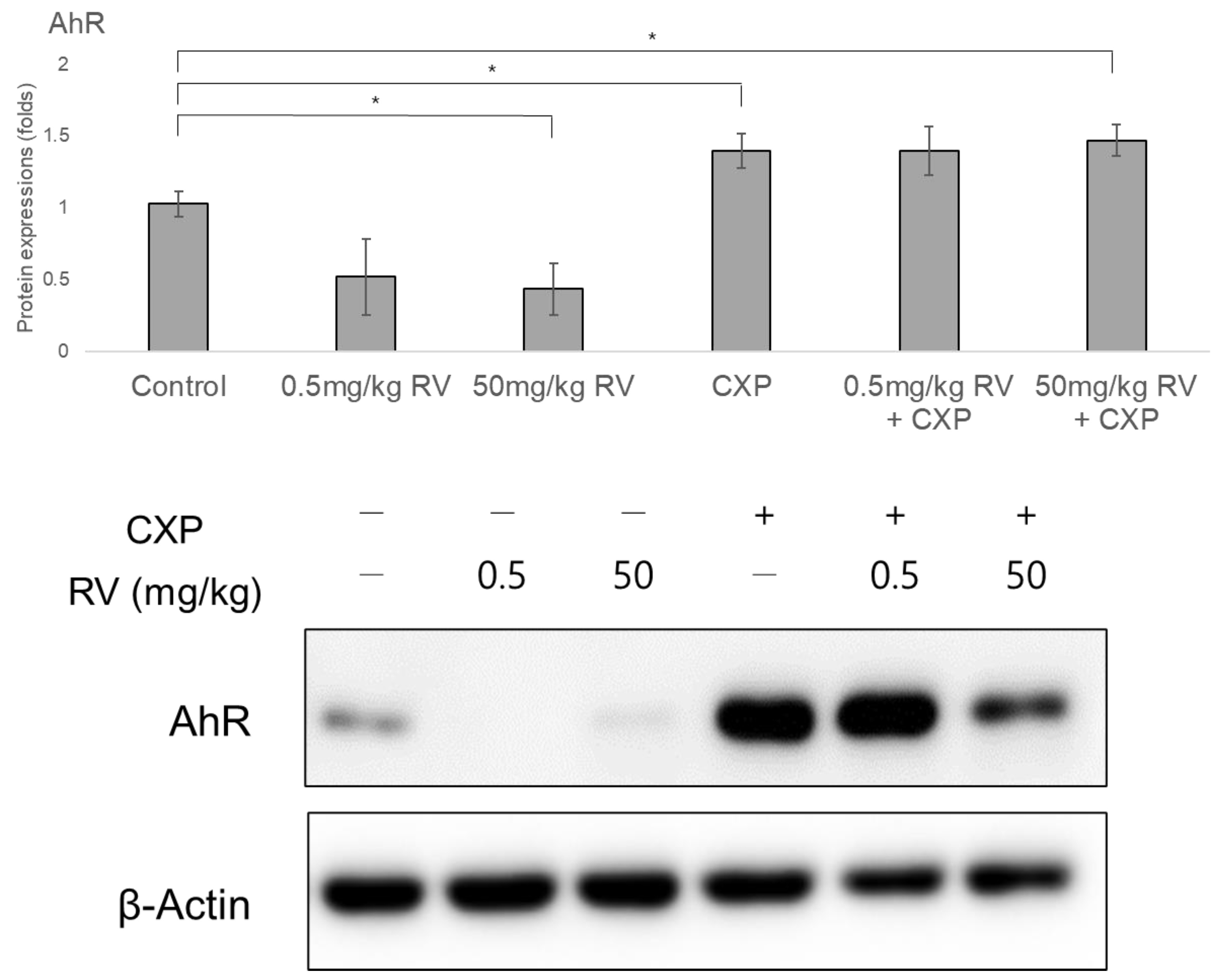
2.4. Decreased AhR Protein and Nuclear/Cytosolic RAGE Levels Following RV Administration
3. Discussion
4. Materials and Methods
4.1. Experimental Design
4.2. ABR Measurements
4.3. Cochlear Whole Mounts
4.4. mRNA Expression of Inflammatory Factors
4.5. Protein Levels of Aryl Hydrocarbon Receptor (AhR) and Receptor for Advanced Glycation Endproducts (RAGE)
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Soleas, G.J.; Diamandis, E.P.; Goldberg, D.M. Resveratrol: A molecule whose time has come? And gone? Clin Biochem. 1997, 30, 91–113. [Google Scholar] [CrossRef]
- Renaud, S.; de Lorgeril, M. Wine, alcohol, platelets, and the French paradox for coronary heart disease. Lancet 1992, 339, 1523–1526. [Google Scholar] [CrossRef]
- Bonnefont-Rousselot, D. Resveratrol and Cardiovascular Diseases. Nutrients 2016, 8, 250. [Google Scholar] [CrossRef] [PubMed]
- Catalgol, B.; Batirel, S.; Taga, Y.; Ozer, N.K. Resveratrol: French paradox revisited. Front Pharmacol. 2012, 3, 141. [Google Scholar] [CrossRef] [Green Version]
- Walle, T.; Hsieh, F.; DeLegge, M.H.; Oatis, J.E., Jr.; Walle, U.K. High absorption but very low bioavailability of oral resveratrol in humans. Drug Metab. Dispos. 2004, 32, 1377–1382. [Google Scholar] [CrossRef] [Green Version]
- Goldberg, D.M.; Yan, J.; Soleas, G.J. Absorption of three wine-related polyphenols in three different matrices by healthy subjects. Clin. Biochem. 2003, 36, 79–87. [Google Scholar] [CrossRef]
- Aziz, M.H.; Reagan-Shaw, S.; Wu, J.; Longley, B.J.; Ahmad, N. Chemoprevention of skin cancer by grape constituent resveratrol: Relevance to human disease? FASEB J. 2005, 19, 1193–1195. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, S.; Dudley, J.I.; Das, D.K. Dose-dependency of resveratrol in providing health benefits. Dose Response 2010, 8, 478–500. [Google Scholar] [CrossRef]
- Xiong, H.; Chen, S.; Lai, L.; Yang, H.; Xu, Y.; Pang, J.; Su, Z.; Lin, H.; Zheng, Y. Modulation of miR-34a/SIRT1 signaling protects cochlear hair cells against oxidative stress and delays age-related hearing loss through coordinated regulation of mitophagy and mitochondrial biogenesis. Neurobiol. Aging 2019, 79, 30–42. [Google Scholar] [CrossRef]
- Su, Z.; Xiong, H.; Pang, J.; Lin, H.; Lai, L.; Zhang, H.; Zhang, W.; Zheng, Y. LncRNA AW112010 Promotes Mitochondrial Biogenesis and Hair Cell Survival: Implications for Age-Related Hearing Loss. Oxid. Med. Cell. Longev. 2019, 2019, 6150148. [Google Scholar] [CrossRef] [Green Version]
- Xiong, H.; Pang, J.; Yang, H.; Dai, M.; Liu, Y.-M.; Ou, Y.; Huang, Q.; Chen, S.; Zhang, Z.; Xu, Y.; et al. Activation of miR-34a/SIRT1/p53 signaling contributes to cochlear hair cell apoptosis: Implications for age-related hearing loss. Neurobiol. Aging 2015, 36, 1692–1701. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.; Ou, Y.; Xu, Y.; Huang, Q.; Pang, J.; Lai, L.; Zheng, Y. Resveratrol Promotes Recovery of Hearing following Intense Noise Exposure by Enhancing Cochlear SIRT1 Activity. Audiol. Neurootol. 2017, 22, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Seidman, M.D.; Tang, W.; Bai, V.U.; Ahmad, N.; Jiang, H.; Media, J.; Patel, N.; Rubin, C.J.; Standring, R.T. Resveratrol decreases noise-induced cyclooxygenase-2 expression in the rat cochlea. Otolaryngol Head Neck Surg. 2013, 148, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Avci, D.; Erkan, M.; Sonmez, M.F.; Kokoglu, K.; Gunes, M.S.; Gundogdu, R.; Gulec, S.; Karabulut, D. A Prospective Experimental Study on the Protective Effect of Resveratrol against Amikacin-Induced Ototoxicity in Rats. J. Int. Adv. Otol. 2016, 12, 290–297. [Google Scholar] [CrossRef]
- Garcia-Alcantara, F.; Murillo-Cuesta, S.; Pulido, S.; Bermudez-Munoz, J.M.; Martinez-Vega, R.; Milo, M.; Varela-Nieto, I.; Rivera, T. The expression of oxidative stress response genes is modulated by a combination of resveratrol and N-acetylcysteine to ameliorate ototoxicity in the rat cochlea. Hear Res. 2018, 358, 10–21. [Google Scholar] [CrossRef]
- Simsek, G.; Tas, B.M.; Muluk, N.B.; Azman, M.; Kilic, R. Comparison of the protective efficacy between intratympanic dexamethasone and resveratrol treatments against cisplatin-induced ototoxicity: An experimental study. Eur. Arch. Otorhinolaryngol. 2019, 276, 3287–3293. [Google Scholar] [CrossRef]
- Yumusakhuylu, A.C.; Yazici, M.; Sari, M.; Binnetoglu, A.; Kosemihal, E.; Akdas, F.; Sirvanci, S.; Yüksel, M.; Üneri, C.; Tutkun, A. Protective role of resveratrol against cisplatin induced ototoxicity in guinea pigs. Int. J. Pediatr. Otorhinolaryngol. 2012, 76, 404–408. [Google Scholar] [CrossRef]
- Erdem, T.; Bayindir, T.; Filiz, A.; Iraz, M.; Selimoglu, E. The effect of resveratrol on the prevention of cisplatin ototoxicity. Eur. Arch. Otorhinolaryngol. 2012, 269, 2185–2188. [Google Scholar] [CrossRef]
- Pang, J.; Xiong, H.; Ou, Y.; Yang, H.; Xu, Y.; Chen, S.; Lai, L.; Ye, Y.; Su, Z.; Lin, H.; et al. SIRT1 protects cochlear hair cell and delays age-related hearing loss via autophagy. Neurobiol. Aging 2019, 80, 127–137. [Google Scholar] [CrossRef]
- Olgun, Y.; Kirkim, G.; Kolatan, E.; Kiray, M.; Bağriyanik, H.A.; Olgun, A.; Kızmazoglu, D.C.; Ellidokuz, H.; Serbetcioglu, B.; Altun, Z.S.; et al. Friend or foe? Effect of oral resveratrol on cisplatin ototoxicity. Laryngoscope 2014, 124, 760–766. [Google Scholar] [CrossRef]
- Li, I.-H.; Shih, J.-H.; Jhao, Y.-T.; Chen, H.-C.; Chiu, C.-H.; Chen, C.-F.F.; Huang, Y.-S.; Shiue, C.-Y.; Ma, K.-H. Regulation of Noise-Induced Loss of Serotonin Transporters with Resveratrol in a Rat Model Using 4-[(18)F]-ADAM/Small-Animal Positron Emission Tomography. Molecules 2019, 24, 1344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, J.; Wang, C.Q.; Fan, H.H.; Ding, H.Y.; Xie, X.L.; Xu, Y.M.; Wang, B.Y.; Huang, D.J. Effects of resveratrol on endothelial progenitor cells and their contributions to reendothelialization in intima-injured rats. J. Cardiovasc. Pharmacol. 2006, 47, 711–721. [Google Scholar] [CrossRef]
- Lee, C.H.; Lee, D.-H.; Lee, S.M.; Kim, S.Y. Otoprotective Effects of Zingerone on Cisplatin-Induced Ototoxicity. Int. J. Mol. Sci. 2020, 21, 3503. [Google Scholar] [CrossRef] [PubMed]
- Hajizadeh-Sharafabad, F.; Sahebkar, A.; Zabetian-Targhi, F.; Maleki, V. The impact of resveratrol on toxicity and related complications of advanced glycation end products: A systematic review. Biofactors 2019, 45, 651–665. [Google Scholar] [CrossRef] [PubMed]
- Hajizadeh-Sharafabad, F.; Sahebkar, A.; Zabetian-Targhi, F.; Alizadeh, M. Effects of Resveratrol on Receptor for Advanced Glycation End Products (RAGE) Expression and Oxidative Stress in the Liver of Rats with Type 2 Diabetes. Phytother. Res. 2016, 30, 66–71. [Google Scholar] [CrossRef]
- Ohtsu, A.; Shibutani, Y.; Seno, K.; Iwata, H.; Kuwayama, T.; Shirasuna, K. Advanced glycation end products and lipopolysaccharides stimulate interleukin-6 secretion via the RAGE/TLR4-NF-kappaB-ROS pathways and resveratrol attenuates these inflammatory responses in mouse macrophages. Exp. Ther. Med. 2017, 14, 4363–4370. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.H.; Park, S.-S.; Lee, D.-H.; Lee, S.M.; Kim, M.Y.; Choi, B.Y.; Kim, S.Y.; So, Y.K. Tauroursodeoxycholic acid attenuates cisplatin-induced hearing loss in rats. Neurosci. Lett. 2020, 722, 134838. [Google Scholar] [CrossRef]
- Assefa, E.G.; Yan, Q.; Gezahegn, S.B.; Salissou, M.T.M.; He, S.; Wu, N.; Zuo, X.; Ying, C. Role of Resveratrol on Indoxyl Sulfate-Induced Endothelial Hyperpermeability via Aryl Hydrocarbon Receptor (AHR)/Src-Dependent Pathway. Oxid. Med. Cell. Longev. 2019, 2019, 5847040. [Google Scholar] [CrossRef]
- Scott, E.; Steward, W.P.; Gescher, A.J.; Brown, K. Resveratrol in human cancer chemoprevention-choosing the ‘right’ dose. Mol. Nutr. Food Res. 2012, 56, 7–13. [Google Scholar] [CrossRef]
- Dudley, J.; Das, S.; Mukherjee, S.; Das, D.K. Resveratrol, a unique phytoalexin present in red wine, delivers either survival signal or death signal to the ischemic myocardium depending on dose. J. Nutr. Biochem. 2009, 20, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Wilson, T.; Knight, T.J.; Beitz, D.C.; Lewis, D.S.; Engen, R.L. Resveratrol promotes atherosclerosis in hypercholesterolemic rabbits. Life Sci. 1996, 59, PL15–PL21. [Google Scholar] [CrossRef]
- Ye, W.; Chen, R.; Chen, X.; Huang, B.; Lin, R.; Xie, X.; Chen, J.; Jiang, J.; Deng, Y.; Wen, J. AhR regulates the expression of human cytochrome P450 1A1 (CYP1A1) by recruiting Sp1. FEBS J. 2019, 286, 4215–4231. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Chiu, J.F.; Liu, J.; Deng, Y.; Xu, C.; Zhang, J.; Li, G. Resveratrol induces autophagy-dependent apoptosis in HL-60 cells. BMC Cancer 2018, 18, 581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapetanovic, I.M. Comment on ‘Resveratrol in human cancer chemoprevention-choosing the ‘right’ dose’. Mol. Nutr. Food Res. 2012, 56, 523. [Google Scholar] [CrossRef] [PubMed]
- Simsek, G.; Tokgoz, S.A.; Vuralkan, E.; Caliskan, M.; Besalti, O.; Akin, I. Protective effects of resveratrol on cisplatin-dependent inner-ear damage in rats. Eur. Arch. Otorhinolaryngol. 2013, 270, 1789–1793. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animals | Dose of Resveratrol (mg/kg) | Cumulative Dose (mg/kg) | Dose of Cisplatin (mg/kg) | Ototoxic/Otoprotective | References |
|---|---|---|---|---|---|
| SD 8-weeks rat | 0.5/day for 10 days (i.p.) | 5 | 3 for 5 days | Otoprotective | Current study |
| Wistar albino rat | 0.1, 1/day for 10 days (i.p.) | 1, 10 | 16 | Otoprotective | Olgun Y. et al., 2014 [20] |
| Adult albino guinea pig | 10/day for 2 days (i.p.) | 20 | 10 for 1 day | Otoprotective | Yumusakhuylu A.C. et al., 2013 [17] |
| Wistar albino | 10/day for 5 days (i.p.) | 50 | 12 | Otoprotective | Erdem T. et al., 2012 [18] |
| 3-months rat | |||||
| Wistar rat | 100/day for 1 day (i.p.) | 100 | 15 | Otoprotective | Simsek G. et al. [35] |
| Albino–Wistar 3-months rat | 20 mg/mL × 0.05 mL/day for 1 days (i.p.) | N/A | 15 for 1 day | Otoprotective | Simsek G. et al. [16] |
| Wistar albino rat | 10/day for 10 days (i.p.) | 100 | 16 | Ototoxic | Olgun Y. et al., 2014 [20] |
| SD 8-weeks rat | 50/day for 10 days (i.p.) | 500 | 3 for 5 days | Ototoxic | Current study |
| Gene | Primer Sequence (Forward) | Primer Sequence (Reverse) | Annealing Temperature (°C) | Product Size (bp) | RefSeq Number |
|---|---|---|---|---|---|
| IL6 | 5′-AGAGACTTCCAGCCAGTTGC-3′ | 5′-TGAAGTCTCCTCTCCGGACT-3′ | 60 | 88 | NM_012589.2 |
| IL1β | 5′-CACCTTCTTTTCCTTCATCTTTG-3′ | 5′-GTCGTTGCTTGTCTCTCCTTGTA-3′ | 60 | 241 | NM_031512.2 |
| NFκB | 5′-TGTCTGCACCTGTTCCAAAGA-3′ | 5′-TGCCAGGTCTGTGAACACTC-3′ | 60 | 143 | NM_199267.2 |
| CYP1A1 | 5′-CATCCCCCACAGCACCATAA-3′ | 5′-TTCGCTTGCCCAAACCAAAG-3′ | 60 | 212 | NM_012540.2 |
| GAPDH | 5′-ATTGTTGCCATCAACGACCC-3′ | 5′-TGACTGTGCCGTTGAACTTG-3′ | 60 | 94 | NM_017008.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, C.H.; Kim, K.W.; Lee, S.M.; Kim, S.Y. Dose-Dependent Effects of Resveratrol on Cisplatin-Induced Hearing Loss. Int. J. Mol. Sci. 2021, 22, 113. https://doi.org/10.3390/ijms22010113
Lee CH, Kim KW, Lee SM, Kim SY. Dose-Dependent Effects of Resveratrol on Cisplatin-Induced Hearing Loss. International Journal of Molecular Sciences. 2021; 22(1):113. https://doi.org/10.3390/ijms22010113
Chicago/Turabian StyleLee, Chang Ho, Kyung Woon Kim, So Min Lee, and So Young Kim. 2021. "Dose-Dependent Effects of Resveratrol on Cisplatin-Induced Hearing Loss" International Journal of Molecular Sciences 22, no. 1: 113. https://doi.org/10.3390/ijms22010113
APA StyleLee, C. H., Kim, K. W., Lee, S. M., & Kim, S. Y. (2021). Dose-Dependent Effects of Resveratrol on Cisplatin-Induced Hearing Loss. International Journal of Molecular Sciences, 22(1), 113. https://doi.org/10.3390/ijms22010113