Anticancer and Immunomodulatory Benefits of Taro (Colocasia esculenta) Corms, an Underexploited Tuber Crop

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
1.1. Underutilized and Neglected Crops Worldwide
1.2. Taro Consumption, Cultivation, and Nutritional Importance
1.3. Risk Factors Associated with Cancer and the Impact of Healthy Dietary Habits
2. Bioactive Compounds and Pharmacological Properties of Taro−Popular Medicinal Knowledge with Anticancer Potential
2.1. Taro Antioxidants
2.2. Taro Protection against Mutagenic and Carcinogenic Agents
2.3. Anticancer, Anti-Inflammatory, and Immunomodulatory Effectiveness of Taro Extracts or Their Components
2.4. Taro Compounds Effect on Type II Diabetes and Obesity
3. Conclusions
4. Methodology
4.1. Guiding Questions
- What are the biological activities associated with taro corms?
- What are the bioactive compounds responsible for taro corm health-promoting effects, and what is their mechanism of action?
- What is the impact of the cooking process on the presence of nutritional components and bioactive compounds of taro corms?
- How can taro corm bioactive compounds contribute to cancer fighting or prevention?
- What could be the benefits provided by taro corm consumption?
4.2. Databases, Descriptors, and/or Keywords
4.3. Eligibility Criteria for Each of the Articles Consulted
- Biological activities not directly related to cancer fighting or prevention and cancer risk factors (antimicrobial, anti-insect, antiviral, anti-helminthic, and others) for this study;
- Studies performed with parts of the plant other than the corm, such as leaves, petioles or roots;
- Unclear or wrong data.
- Experimental studies (in vitro, in vivo or clinical trial) that analyzed biological properties considered important for cancer fighting or prevention, such as antioxidant, antitumoral, antimetastatic, immunomodulatory, anti-hyperglycemic, antidiabetic, antimutagenic, and anti-hyperlipidemic activities;
- Studies performed with corms, edible part of taro;
- Articles published up to 2020 in the English language with no restriction regarding time period;
- Review articles describing Colocasia esculenta characteristics, production, nutritional importance, medicinal uses and other general information;
- Studies that purify or identify any taro component that has been proven to exhibit the claimed biological activities specified in item 1.
Author Contributions
Funding
Conflicts of Interest
References
- Abele, S.; Frohberg, K. Subsistence Agriculture in Central and Eastern Europe: How to Break the Vicious Circle? Oxford University Press: New York, NY, USA, 2004. [Google Scholar]
- Pereira, P.R.; Corrêa, A.C.N.T.F.; Vericimo, M.A.; Paschoalin, V.M.F. Tarin, a Potential Immunomodulator and COX-Inhibitor Lectin Found in Taro (Colocasia esculenta). Compr. Rev. Food Sci. Food Saf. 2018, 17, 878–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Temesgen, M.; Retta, N. Nutritional potential, health and food security benefits of taro Colocasia esculenta (L.): A Review. Food Sci. Qual. Manag. 2015, 36, 23–30. [Google Scholar]
- Siqueira, M.V. Yam: A neglected and underutilized crop in Brazil. Hortic. Bras. 2011, 29, 16–20. [Google Scholar] [CrossRef] [Green Version]
- Akwee, P.; Netondo, G.; Palapala, V.A. A critical review of the role of taro Colocasia esculenta L.(Schott) to food security: A comparative analysis of Kenya and Pacific Island taro germplasm. Sci. Agric. 2015, 9, 101–108. [Google Scholar] [CrossRef]
- Rao, V.R.; Hunter, D.; Eyzaguirre, P.B.; Matthews, P.J. Ethnobotany and global diversity of taro. In The Global Diversity of Taro: Ethnobotany and Conservation; Rao, V.R., Matthews, P.J., Eyzaguirre, P.B., Hunter, D., Eds.; Bioversity International: Rome, Italy, 2010; Volume 1. [Google Scholar]
- FAO. Food and Agriculture Organization of the United Nations. Statistics Division: Production of Taro. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 17 February 2020).
- Diop, N.; Jaffee, S.M. Fruits and vegetables: Global trade and competition in fresh and processed product markets. In Global Agricultural Trade and Developing Countries; Aksoy, M.A., Beghin, J.C., Eds.; The World Bank: Washington, DC, USA, 2005; pp. 237–254. [Google Scholar]
- Vieira, G.H.S.; Peterle, G.; Loss, J.B.; Peterle, G.; Poloni, C.M.M.; Colombo, J.N.; Monaco, P.A.V.L. Strategies for taro (Colocasia esculenta) irrigation. J. Exp. Agric. Int. 2018, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, E.E. A Cultura do Taro-Inhame (Colocasia esculenta (L.) Schott): Alternativa para o Estado de Roraima; Embrapa Roraima: Boa Vista, Roraima, Brazil, 2011. [Google Scholar]
- Nunes, R.S.C.; Del Aguila, E.; Paschoalin, V.; da Silva, J. DNA barcoding assessment of the genetic diversity of varieties of taro, Colocasia esculenta (L.) Schott in Brazil. In Breeding and Genetic Engineering: The Biology and Biotechnology Research; iConcept Press Ltd.: Hong Kong, China, 2014. [Google Scholar]
- USDA. FoodData Central. Available online: https://fdc.nal.usda.gov/ (accessed on 17 February 2020).
- Simsek, S.; El, S.N. In vitro starch digestibility, estimated glycemic index and antioxidant potential of taro (Colocasia esculenta L. Schott) corm. Food Chem. 2015, 168, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, R.; Qin, C.; Meng, Y.; Zhang, J.; Wang, Y.; Xu, G. Sources and intake of resistant starch in the Chinese diet. Asia Pac. J. Clin. Nutr. 2010, 19, 274–282. [Google Scholar]
- Ramdath, D.D.; Isaacs, R.L.; Teelucksingh, S.; Wolever, T.M. Glycaemic index of selected staples commonly eaten in the Caribbean and the effects of boiling v. crushing. Br. J. Nutr. 2004, 91, 971–977. [Google Scholar] [CrossRef] [Green Version]
- Bahado-Singh, P.; Wheatley, A.; Ahmad, M.; Morrison, E.S.A.; Asemota, H. Food processing methods influence the glycaemic indices of some commonly eaten West Indian carbohydrate-rich foods. Br. J. Nutr. 2006, 96, 476–481. [Google Scholar] [CrossRef]
- Foster-Powell, K.; Holt, S.H.; Brand-Miller, J.C. International table of glycemic index and glycemic load values: 2002. Am. J. Clin. Nutr. 2002, 76, 5–56. [Google Scholar] [CrossRef] [Green Version]
- Bsc, S.N.; Bsc, G.S. Oxalate content of foods and its effect on humans. Asia Pac. J. Clin. Nutr. 1999, 8, 64–74. [Google Scholar] [CrossRef]
- Brown, A.C.; Ibrahim, S.A.; Song, D. Poi history, uses, and role in health. In Fruits, Vegetables, and Herbs; Elsevier: Amsterdam, The Netherlands, 2016; pp. 331–342. [Google Scholar]
- Grimaldi, I.M.; Leke, W.N.; Nkeabeng, I.M.L.; van Andel, T. Traditional preparation of Achu, a cultural keystone dish in western Cameroon. Int. J. Gastron. Food Sci. 2018, 13, 25–28. [Google Scholar] [CrossRef]
- Maga, J.A. Taro: Composition and food uses. Food Rev. Int. 1992, 8, 443–473. [Google Scholar] [CrossRef]
- Kaushal, P.; Kumar, V.; Sharma, H. Utilization of taro (Colocasia esculenta): A review. J. Food Sci. Technol. 2015, 52, 27–40. [Google Scholar] [CrossRef]
- Hyacinthe, A.A.; Bedel, F.J.; Constant, Y.J.; Soumaila, D.; Patrice, K.L. Bread characteristics and descriptive analysis of taro (Colocasia esculenta, Cv Fouê): Wheat composite bread and some fritters. Int. J. Food Sci. Nutr. 2018, 3, 41–45. [Google Scholar]
- Ammar, M.; Hegazy, A.; Bedeir, S. Using of taro flour as partial substitute of wheat flour in bread making. World J. Dairy Food Sci. 2009, 4, 94–99. [Google Scholar]
- Tattiyakul, J.; Asavasaksakul, S.; Pradipasena, P. Chemical and physical properties of flour extracted from taro Colocasia esculenta (L.) Schott grown in different regions of Thailand. Sci. Asia 2006, 32, 279–284. [Google Scholar] [CrossRef]
- Simsek, S.; El, S.N. Production of resistant starch from taro (Colocasia esculenta L. Schott) corm and determination of its effects on health by in vitro methods. Carbohydr. Polym. 2012, 90, 1204–1209. [Google Scholar] [CrossRef]
- Atkinson, F.S.; Foster-Powell, K.; Brand-Miller, J.C. International tables of glycemic index and glycemic load values: 2008. Diabetes Care 2008, 31, 2281–2283. [Google Scholar] [CrossRef] [Green Version]
- Goñi, I.; Garcia-Alonso, A.; Saura-Calixto, F. A starch hydrolysis procedure to estimate glycemic index. Nutr. Res. 1997, 17, 427–437. [Google Scholar] [CrossRef]
- Sajilata, M.G.; Singhal, R.S.; Kulkarni, P.R. Resistant starch—A review. Compr. Rev. Food Sci. Food Saf. 2006, 5, 1–17. [Google Scholar] [CrossRef]
- Pearson-Stuttard, J.; Zhou, B.; Kontis, V.; Bentham, J.; Gunter, M.J.; Ezzati, M. Retraction and republication—Worldwide burden of cancer attributable to diabetes and high body-mass index: A comparative risk assessment. Lancet Diabetes Endocrinol. 2018, 6, S2213–S8587. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Zhu, W.; Thompson, P.; Hannun, Y.A. Evaluating intrinsic and non-intrinsic cancer risk factors. Nat. Commun. 2018, 9, 3490. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Powers, S.; Zhu, W.; Hannun, Y.A. Substantial contribution of extrinsic risk factors to cancer development. Nature 2016, 529, 43. [Google Scholar] [CrossRef]
- Bamia, C. Dietary patterns in association to cancer incidence and survival: Concept, current evidence, and suggestions for future research. Eur. J. Clin. Nutr. 2018, 72, 818–825. [Google Scholar] [CrossRef]
- Li, Y.; Schoufour, J.; Wang, D.D.; Dhana, K.; Pan, A.; Liu, X.; Song, M.; Liu, G.; Shin, H.J.; Sun, Q. Healthy lifestyle and life expectancy free of cancer, cardiovascular disease, and type 2 diabetes: Prospective cohort study. BMJ 2020, 368, 16669. [Google Scholar] [CrossRef] [Green Version]
- Steck, S.E.; Murphy, E.A. Dietary patterns and cancer risk. Nat. Rev. Cancer 2019, 20, 125–138. [Google Scholar] [CrossRef]
- Clinton, S.K.; Grainger, E.M.; Giovannucci, E.L. Diet and Nutrition in the Etiology and Prevention of Cancer. Holl. Frei. Cancer Med. 2016, 1–21. [Google Scholar] [CrossRef]
- Vanamala, J. Food systems approach to cancer prevention. Crit. Rev. Food Sci. Nutr. 2017, 57, 2573–2588. [Google Scholar] [CrossRef]
- Davis, C.D.; Ross, S. Diet and cancer prevention. In Lifestyle Medicine, 3rd ed.; Rippe, J.M., Ed.; CRC Press Taylor & Francis Group: New York, NY, USA, 2019; pp. 409–415. [Google Scholar]
- Prajapati, R.; Kalariya, M.; Umbarkar, R.; Parmar, S.; Sheth, N. Colocasia esculenta: A potent indigenous plant. Int. J. Nutr. Pharmacol. Neurol. Dis. 2011, 1, 90. [Google Scholar] [CrossRef]
- Lim, T. Colocasia esculenta. In Edible Medicinal and Non Medicinal Plants, 1st ed.; Lim, T., Ed.; Springer: Heidelberg, Germany, 2015; pp. 454–492. [Google Scholar]
- Brown, A.C.; Reitzenstein, J.E.; Liu, J.; Jadus, M.R. The anti-cancer effects of poi (Colocasia esculenta) on colonic adenocarcinoma cells In vitro. Phytother. Res. Ptr. 2005, 19, 767–771. [Google Scholar] [CrossRef]
- Kundu, N.; Campbell, P.; Hampton, B.; Lin, C.-Y.; Ma, X.; Ambulos, N.; Zhao, X.F.; Goloubeva, O.; Holt, D.; Fulton, A.M. Antimetastatic activity isolated from Colocasia esculenta (taro). Anti-Cancer Drugs 2012, 23, 200. [Google Scholar] [CrossRef] [Green Version]
- Yau Sang, C.; Jack Ho, W.; Tzi Bun, N. A Cytokine-Inducing Hemagglutinin from Small Taros. Protein Pept. Lett. 2010, 17, 823–830. [Google Scholar] [CrossRef]
- Corrêa, A.C.N.T.F.; Vericimo, M.A.; Dashevskiy, A.; Pereira, P.R.; Paschoalin, V.M.F. Liposomal Taro Lectin Nanocapsules Control Human Glioblastoma and Mammary Adenocarcinoma Cell Proliferation. Molecules 2019, 24, 471. [Google Scholar] [CrossRef] [Green Version]
- Park, H.R.; Lee, H.S.; Cho, S.Y.; Kim, Y.S.; Shin, K.S. Anti-metastatic effect of polysaccharide isolated from Colocasia esculenta is exerted through immunostimulation. Int. J. Mol. Med. 2013, 31, 361–368. [Google Scholar] [CrossRef] [Green Version]
- Kai, H.; Akamatsu, E.; Torii, E.; Kodama, H.; Yukizaki, C.; Sakakibara, Y.; Suiko, M.; Morishita, K.; Kataoka, H.; Matsuno, K. Inhibition of proliferation by agricultural plant extracts in seven human adult T-cell leukaemia (ATL)-related cell lines. J. Nat. Med. 2011, 65, 651–655. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Dong, Z.; Liu, X.; Chen, H.; Lai, F.; Zhang, M. Structure characterization of two novel polysaccharides from Colocasia esculenta (taro) and a comparative study of their immunomodulatory activities. J. Funct. Foods 2018, 42, 47–57. [Google Scholar] [CrossRef]
- Pereira, P.R.; Del Aguila, E.M.; Verícimo, M.A.; Zingali, R.B.; Paschoalin, V.M.F.; Silva, J.T. Purification and characterization of the lectin from taro (Colocasia esculenta) and its effect on mouse splenocyte proliferation in vitro and in vivo. Protein J. 2014, 33, 92–99. [Google Scholar] [CrossRef]
- Tulin, E.E.; Ecleo, Z.T. Cytokine-mimetic properties of some Philippine food and medicinal plants. J. Med. Food 2007, 10, 290–299. [Google Scholar] [CrossRef]
- Merida, L.A.; Mattos, E.B.; Correa, A.C.; Pereira, P.R.; Paschoalin, V.M.; Pinho, M.F.; Vericimo, M.A. Tarin stimulates granulocyte growth in bone marrow cell cultures and minimizes immunosuppression by cyclo-phosphamide in mice. PLoS ONE 2018, 13, e0206240. [Google Scholar] [CrossRef] [Green Version]
- Pereira, P.R.; Silva, J.T.; Verícimo, M.A.; Paschoalin, V.M.F.; Teixeira, G.A.P.B. Crude extract from taro (Colocasia esculenta) as a natural source of bioactive proteins able to stimulate haematopoietic cells in two murine models. J. Funct. Foods 2015, 18, 333–343. [Google Scholar] [CrossRef] [Green Version]
- Eleazu, C.O.; Eleazu, K.C.; Iroaganachi, M.A. Effect of cocoyam (Colocasia esculenta), unripe plantain (Musa paradisiaca) or their combination on glycated hemoglobin, lipogenic enzymes, and lipid metabolism of streptozotocin-induced diabetic rats. Pharm. Biol. 2016, 54, 91–97. [Google Scholar] [CrossRef] [Green Version]
- Eleazu, C.O.; Iroaganachi, M.A.; Eleazu, K.C. Ameliorative potentials of cocoyam (Colocasia esculenta L.) and unripe plantain (Musa paradisiaca L.) on the relative tissue weights of streptozotocin-induced diabetic rats. J. Diabetes Res. 2013, 2013, 160964. [Google Scholar] [CrossRef] [Green Version]
- Eleazu, C.O.; Okafor, P.N.; Ifeoma, I. Biochemical basis of the use of cocoyam (Colocassia esculenta L.) in the dietary management of diabetes and its complications in streptozotocin induced diabetes in rats. Asian Pac. J. Trop. Dis. 2014, 4, S705–S711. [Google Scholar] [CrossRef]
- Islam, M.H.; Mostafa, M.N.; Rahmatullah, M. Antihyperglycemic activity of methanolic extracts of corms of Colocasia esculenta var esculenta. Eur. J. Pharm. Med. Res. 2018, 5, 129–132. [Google Scholar]
- Chukwuma, C.I.; Islam, M.S.; Amonsou, E.O. A comparative study on the physicochemical, anti-oxidative, anti-hyperglycemic and anti-lipidemic properties of amadumbe (Colocasia esculenta) and okra (Abelmoschus esculentus) mucilage. J. Food Biochem. 2018, 42, e12601. [Google Scholar] [CrossRef]
- McEwan, R.; Madivha, R.; Djarova, T.; Oyedeji, O.; Opoku, A. Alpha-amylase inhibitor of amadumbe (Colocasia esculenta): Isolation, purification and selectivity toward-amylases from various sources. Afr. J. Biochem. Res. 2010, 4, 220–224. [Google Scholar]
- Fidyasari, A.; Raharjo, S.J.; Widiarto, E. Instant Tiwul Made of Colocasia esculenta (L.) Schott as A Current Functional Food Development for Hypercholesterolemic Patients. In Proceedings of the Innovation of Food Technology to Improve Food Security and Health, Surabaya, Indonesia, 20–21 October 2016; pp. 57–64. [Google Scholar]
- Sakano, Y.; Mutsuga, M.; Tanaka, R.; Suganuma, H.; Inakuma, T.; Toyoda, M.; Goda, Y.; Shibuya, M.; Ebizuka, Y. Inhibition of human lanosterol synthase by the constituents of Colocasia esculenta (taro). Biol. Pharm. Bull. 2005, 28, 299–304. [Google Scholar] [CrossRef] [Green Version]
- Boban, P.T.; Nambisan, B.; Sudhakaran, P.R. Hypolipidaemic effect of chemically different mucilages in rats: A comparative study. Br. J. Nutr. 2006, 96, 1021–1029. [Google Scholar] [CrossRef]
- Ferguson, L.R.; Roberton, A.M.; McKenzie, R.J.; Watson, M.E.; Harris, P.J. Adsorption of a hydrophobic mutagen to dietary fiber from taro (Colocasia esculenta), an important food plant of the South Pacific. Nutr. Cancer 1992, 17, 85–95. [Google Scholar] [CrossRef]
- Nakamura, Y.; Suganuma, E.; Kuyama, N.; Sato, K.; Ohtsuki, K. Comparative bio-antimutagenicity of common vegetables and traditional vegetables in Kyoto. Biosci. Biotechnol. Biochem. 1998, 62, 1161–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botting, K.; Young, M.; Pearson, A.; Harris, P.; Ferguson, L. Antimutagens in food plants eaten by Polynesians: Micronutrients, phytochemicals and protection against bacterial mutagenicity of the heterocyclic amine 2-amino-3-methylimidazo [4, 5-f] quinoline. Food Chem. Toxicol. 1999, 37, 95–103. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M. Free Radicals in Biology and Medicine; Oxford University Press: New York, NY, USA, 2015. [Google Scholar]
- Osawa, T.; Kato, Y. Protective role of antioxidative food factors in oxidative stress caused by hyperglycemia. Ann. N. Y. Acad. Sci. 2005, 1043, 440–451. [Google Scholar] [CrossRef] [PubMed]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef]
- Valtuena, S.; Pellegrini, N.; Franzini, L.; Bianchi, M.A.; Ardigo, D.; Del Rio, D.; Piatti, P.; Scazzina, F.; Zavaroni, I.; Brighenti, F. Food selection based on total antioxidant capacity can modify antioxidant intake, systemic inflammation, and liver function without altering markers of oxidative stress. Am. J. Clin. Nutr. 2008, 87, 1290–1297. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Anil Kumar, N.V.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Tsouh Fokou, P.V.; Azzini, E.; Peluso, I.; et al. Lifestyle, Oxidative Stress, and Antioxidants: Back and Forth in the Pathophysiology of Chronic Diseases. Front. Physiol. 2020, 11, 694. [Google Scholar] [CrossRef]
- Agbor-Egbe, T.; Rickard, J.E. Identification of phenolic compounds in edible aroids. J. Sci. Food Agric. 1990, 51, 215–221. [Google Scholar] [CrossRef]
- Isabelle, M.; Lee, B.L.; Lim, M.T.; Koh, W.-P.; Huang, D.; Ong, C.N. Antioxidant activity and profiles of common fruits in Singapore. Food Chem. 2010, 123, 77–84. [Google Scholar] [CrossRef]
- El-Dardiry, A.I.; Ewis, A.M.; Abo-Srea, M. Impact of Taro Corms on Functional Low Fat Ice Cream Properties. J. Food Dairy Sci. 2018, 9, 399–402. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.S.; Adeyemi, D.; Korovulavula, P.; Jang, D.W.; Park, M.-K. Effect of steaming on the functional compounds and antioxidant activity of Fijian taro (Colocasia esculenta L. Schott) corms. Korean J. Food Preserv. 2019, 26, 449–454. [Google Scholar] [CrossRef]
- Tuti, M.; Pal, R.; Arun Kumar, R.; Bist, J.; Bhatt, J. Colocasia based cropping systems affects the antioxidant properties and productivity of colocasia [Colocasia esculenta (l.) schott] tuber. Bioscan Int. Q. J. Life Sci. 2015, 10, 117–123. [Google Scholar]
- Chakraborty, P.; Deb, P.; Chakraborty, S.; Chatterjee, B.; Abraham, J. Cytotoxicity and antimicrobial activity of Colocasia esculenta. J. Chem. Pharm. Res. 2015, 7, 627–635. [Google Scholar]
- Akshatha, M.; Kavadikeri, S.; Rao, N.N. In vitro micropropagation and antioxidant assay in Colocasia esculenta. Plant Tissue Cult. Biotechnol. 2018, 28, 183–190. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Sharma, H. Process optimization for extraction of bioactive compounds from taro (Colocasia esculenta), using RSM and ANFIS modeling. J. Food Meas. Charact. 2017, 11, 704–718. [Google Scholar] [CrossRef]
- Takebayashi, J.; Oki, T.; Watanabe, J.; Yamasaki, K.; Chen, J.; Sato-Furukawa, M.; Tsubota-Utsugi, M.; Taku, K.; Goto, K.; Matsumoto, T. Hydrophilic antioxidant capacities of vegetables and fruits commonly consumed in Japan and estimated average daily intake of hydrophilic antioxidants from these foods. J. Food Compos. Anal. 2013, 29, 25–31. [Google Scholar] [CrossRef]
- Lee, S.; Wee, W.; Yong, J.; Syamsumir, D. Antimicrobial, antioxidant, anticancer property and chemical composition of different parts (corm, stem and leave) of Colocasia esculenta extract. Ann. Univ. Mariae Curie-Skłodowska 2011, 24, 9–16. [Google Scholar]
- Awa, E.; Eleazu, C. Bioactive constituents and antioxidant activities of raw and processed cocoyam (Colocasia esculenta). Nutrafoods 2015, 14, 133–140. [Google Scholar] [CrossRef]
- Eleazu, C.O. Characterization of the natural products in cocoyam (Colocasia esculenta) using GC–MS. Pharm. Biol. 2016, 54, 2880–2885. [Google Scholar] [CrossRef] [Green Version]
- Alcantara, M.; Hurtada, A.; Dizon, I. The nutritional value and phytochemical components of taro [Colocasia esculenta (L.) Schott] powder and its selected processed foods. Nutr. Food Sci. 2013, 3, 3. [Google Scholar] [CrossRef] [Green Version]
- Lindsey, K.; Motsei, M.; Jäger, A. Screening of South African food plants for antioxidant activity. J. Food Sci. 2002, 67, 2129–2131. [Google Scholar] [CrossRef]
- Akyuz, M. Determination of Antioxidant Activity of Ethanol Extract of Gölevez [(Colocasia esculenta (L.)] Tubers. Ksu J. Agric. Nat. 2019, 22, 388–394. [Google Scholar] [CrossRef]
- Goncalves, R.F.; Silva, A.M.; Silva, A.M.; Valentao, P.; Ferreres, F.; Gil-Izquierdo, A.; Silva, J.B.; Santos, D.; Andrade, P.B. Influence of taro (Colocasia esculenta L. Shott) growth conditions on the phenolic composition and biological properties. Food Chem. 2013, 141, 3480–3485. [Google Scholar] [CrossRef] [PubMed]
- Ferreres, F.; Goncalves, R.F.; Gil-Izquierdo, A.; Valentao, P.; Silva, A.M.; Silva, J.B.; Santos, D.; Andrade, P.B. Further knowledge on the phenolic profile of Colocasia esculenta (L.) Shott. J. Agric. Food Chem. 2012, 60, 7005–7015. [Google Scholar] [CrossRef] [PubMed]
- Chipurura, B.; Muchuweti, M.; Manditseraa, F. Effects of thermal treatment on the phenolic content and antioxidant activity of some vegetables. Asian J. Clin. Nutr. 2010, 2, 93–100. [Google Scholar] [CrossRef]
- Amin, I.; Lee, W.Y. Effect of different blanching times on antioxidant properties in selected cruciferous vegetables. J. Sci. Food Agric. 2005, 85, 2314–2320. [Google Scholar] [CrossRef]
- Rebey, I.B.; Bourgou, S.; Debez, I.B.S.; Karoui, I.J.; Sellami, I.H.; Msaada, K.; Limam, F.; Marzouk, B. Effects of extraction solvents and provenances on phenolic contents and antioxidant activities of cumin (Cuminum cyminum L.) seeds. Food Bioprocess Technol. 2012, 5, 2827–2836. [Google Scholar] [CrossRef]
- Bae, H.; Jayaprakasha, G.K.; Crosby, K.; Jifon, J.L.; Patil, B.S. Influence of extraction solvents on antioxidant activity and the content of bioactive compounds in non-pungent peppers. Plant Foods Hum. Nutr. 2012, 67, 120–128. [Google Scholar] [CrossRef]
- Collins, A.R. Antioxidant intervention as a route to cancer prevention. Eur. J. Cancer 2005, 41, 1923–1930. [Google Scholar] [CrossRef]
- Bellik, Y.; Boukraâ, L.; Alzahrani, H.A.; Bakhotmah, B.A.; Abdellah, F.; Hammoudi, S.M.; Iguer-Ouada, M. Molecular mechanism underlying anti-inflammatory and anti-allergic activities of phytochemicals: An update. Molecules 2013, 18, 322. [Google Scholar] [CrossRef] [Green Version]
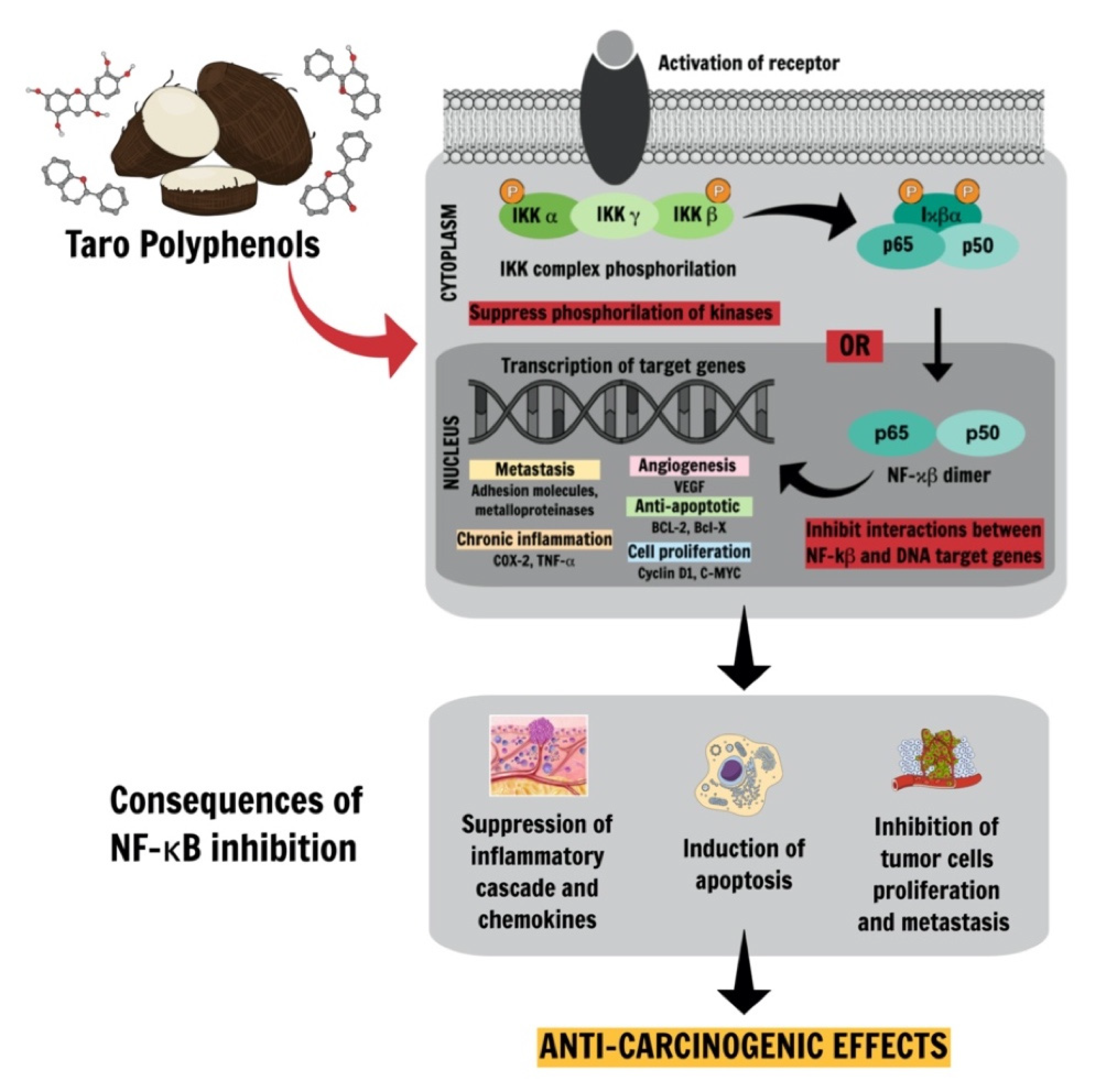
- Khan, H.; Ullah, H.; Castilho, P.C.M.F.; Gomila, A.S.; D’Onofrio, G.; Filosa, R.; Wang, F.; Nabavi, S.M.; Daglia, M.; Silva, A.S. Targeting NF-κB signaling pathway in cancer by dietary polyphenols. Crit. Rev. Food Sci. Nutr. 2019, 60, 2790–2800. [Google Scholar] [CrossRef]
- Dewanto, V.; Wu, X.; Liu, R.H. Processed sweet corn has higher antioxidant activity. J. Agric. Food Chem. 2002, 50, 4959–4964. [Google Scholar] [CrossRef] [PubMed]
- Ajila, C.; Brar, S.; Verma, M.; Tyagi, R.; Valéro, J. Solid-state fermentation of apple pomace using Phanerocheate chrysosporium–Liberation and extraction of phenolic antioxidants. Food Chem. 2011, 126, 1071–1080. [Google Scholar] [CrossRef]
- Kamiloglu, S.; Toydemir, G.; Boyacioglu, D.; Beekwilder, J.; Hall, R.D.; Capanoglu, E. A review on the effect of drying on antioxidant potential of fruits and vegetables. Crit. Rev. Food Sci. Nutr. 2016, 56, S110–S129. [Google Scholar] [CrossRef] [PubMed]
- Asadi-Samani, M.; Farkhad, N.K.; Mahmoudian-Sani, M.R.; Shirzad, H. Antioxidants as a Double-Edged Sword in the Treatment of Cancer; Shalaby, E., Ed.; IntechOpen: London, UK, 2019. [Google Scholar]
- Wang, J.-K.; Higa, S. Taro, a Review of Colocasia Esculenta and Its Potentials; University of Hawaii Press: Honolulu, HI, USA, 1983. [Google Scholar]
- National Research Council. Underexploited Tropical Plants with Promising Economic Value: Report of an Ad Hoc Panel of the Advisory Committee on Technology Innovation, Board on Science and Technology for International Development, Commission on International Relations; National Academy of Sciences: Washington, DC, USA, 1975. [Google Scholar]
- Chandra, S. Tropical root crops and their potential for food in the less developed countries. Food Rev. Int. 1986, 2, 143–169. [Google Scholar] [CrossRef]
- Słoczyńska, K.; Powroźnik, B.; Pękala, E.; Waszkielewicz, A.M. Antimutagenic compounds and their possible mechanisms of action. J. Appl. Genet. 2014, 55, 273–285. [Google Scholar] [CrossRef] [Green Version]
- Makhafola, T.J.; Elgorashi, E.E.; McGaw, L.J.; Verschaeve, L.; Eloff, J.N. The correlation between antimutagenic activity and total phenolic content of extracts of 31 plant species with high antioxidant activity. BMC Complement. Altern. Med. 2016, 16, 490. [Google Scholar] [CrossRef] [Green Version]
- López-Romero, D.; Izquierdo-Vega, J.A.; Morales-González, J.A.; Madrigal-Bujaidar, E.; Chamorro-Cevallos, G.; Sánchez-Gutiérrez, M.; Betanzos-Cabrera, G.; Alvarez-Gonzalez, I.; Morales-González, Á.; Madrigal-Santillán, E. Evidence of Some Natural Products with Antigenotoxic Effects. Part 2: Plants, Vegetables, and Natural Resin. Nutrients 2018, 10, 1954. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Huang, P.; Hu, M.; Ling, S.; Li, Y.; Chen, B.; Xu, Y. Dietary fiber intake and risks of proximal and distal colon cancers: A systematic review and meta-analysis. J. Clin. Oncol. 2017, 35, e15080. [Google Scholar] [CrossRef]
- McRae, M.P. The benefits of dietary fiber intake on reducing the risk of cancer: An umbrella review of meta-analyses. J. Chiropr. Med. 2018, 17, 90–96. [Google Scholar] [CrossRef]
- Zhang, Z.; Xu, G.; Ma, M.; Yang, J.; Liu, X. Dietary fiber intake reduces risk for gastric cancer: A meta-analysis. Gastroenterology 2013, 145, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Ben, Q.; Sun, Y.; Chai, R.; Qian, A.; Xu, B.; Yuan, Y. Dietary fiber intake reduces risk for colorectal adenoma: A meta-analysis. Gastroenterology 2014, 146, 689–699. [Google Scholar] [CrossRef] [PubMed]
- NIH. Naitional Cancer Institute. Available online: https://www.cancer.gov (accessed on 16 April 2020).
- American Institute for Cancer Research. Cancer Prevention. Available online: https://www.aicr.org/cancer-prevention/healthy-eating/ (accessed on 16 April 2020).
- Society, A.C. Stay Healthy. Available online: https://www.cancer.org (accessed on 16 April 2020).
- Zeng, H.; Lazarova, D.L.; Bordonaro, M. Mechanisms linking dietary fiber, gut microbiota and colon cancer prevention. World J. Gastrointest. Oncol. 2014, 6, 41. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capuano, E. The behavior of dietary fiber in the gastrointestinal tract determines its physiological effect. Crit. Rev. Food Sci. Nutr. 2017, 57, 3543–3564. [Google Scholar] [CrossRef] [Green Version]
- Brister, M.M.; Piñero-Santiago, L.E.; Morel, M.; Arce, R.; Crespo-Hernández, C.E. Photochemical Relaxation Pathways in Dinitropyrene Isomer Pollutants. J. Phys. Chem. A 2017, 121, 8197–8206. [Google Scholar] [CrossRef]
- Veronese, N.; Solmi, M.; Caruso, M.G.; Giannelli, G.; Osella, A.R.; Evangelou, E.; Maggi, S.; Fontana, L.; Stubbs, B.; Tzoulaki, I. Dietary fiber and health outcomes: An umbrella review of systematic reviews and meta-analyses. Am. J. Clin. Nutr. 2018, 107, 436–444. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, H.; Hagerling, C.; Werb, Z. Roles of the immune system in cancer: From tumor initiation to metastatic progression. Genes Dev. 2018, 32, 1267–1284. [Google Scholar] [CrossRef] [Green Version]
- Pereira, P.R.; Winter, H.C.; Verícimo, M.A.; Meagher, J.L.; Stuckey, J.A.; Goldstein, I.J.; Paschoalin, V.M.F.; Silva, J.T. Structural analysis and binding properties of isoforms of tarin, the GNA-related lectin from Colocasia esculenta. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2015, 1854, 20–30. [Google Scholar] [CrossRef] [Green Version]
- Pereira, P.R.; Meagher, J.L.; Winter, H.C.; Goldstein, I.J.; Paschoalin, V.M.F.; Silva, J.T.; Stuckey, J.A. High-resolution crystal structures of Colocasia esculenta tarin lectin. Glycobiology 2017, 27, 50–56. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Qu, L.; Yan, S. Cyclooxygenase-2 promotes tumor growth and suppresses tumor immunity. Cancer Cell Int. 2015, 15, 106. [Google Scholar] [CrossRef] [Green Version]
- Pannunzio, A.; Coluccia, M. Cyclooxygenase-1 (COX-1) and COX-1 inhibitors in cancer: A review of oncology and medicinal chemistry literature. Pharmaceuticals 2018, 11, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashemi Goradel, N.; Najafi, M.; Salehi, E.; Farhood, B.; Mortezaee, K. Cyclooxygenase-2 in cancer: A review. J. Cell. Physiol. 2019, 234, 5683–5699. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.E.; Beebe, J.; Alshafie, G.A. Reduction in cancer risk by selective and nonselective cyclooxygenase-2 (COX-2) inhibitors. J. Exp. Pharmacol. 2012, 4, 91–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Bao, J.-k. Anti-tumor and anti-viral activities of Galanthus nivalis agglutinin (GNA)-related lectins. Glycoconj. J. 2013, 30, 269–279. [Google Scholar] [CrossRef]
- Mazalovska, M.; Kouokam, J.C. Plant-Derived Lectins as Potential Cancer Therapeutics and Diagnostic Tools. Biomed. Res. Int. 2020, 2020, 1631394. [Google Scholar] [CrossRef]
- Wang, D.; DuBois, R.N. Eicosanoids and cancer. Nat. Rev. Cancer 2010, 10, 181–193. [Google Scholar] [CrossRef]
- Muñoz-Cuervo, I.; Malapa, R.; Michalet, S.; Lebot, V.; Legendre, L. Secondary metabolite diversity in taro, Colocasia esculenta (L.) Schott, corms. J. Food Compos. Anal. 2016, 52, 24–32. [Google Scholar] [CrossRef]
- Pawar, H.A.; Choudhary, P.D.; Gavasane, A.J. Isolation and Characterisation of Water Soluble Polysaccharide from Colocasia esculenta Tubers. In Handbook of Composites from Renewable Materials; Kessler, M.R., Ed.; Scrivener Publishing and Wiley & Sons, Inc.: Beverly, MA, USA, 2017; Volume 1, pp. 221–241. [Google Scholar]
- Yamasaki, M.; Mukai, A.; Ohba, M.; Mine, Y.; Sakakibara, Y.; Suiko, M.; Morishita, K.; Nishiyama, K. Genistein induced apoptotic cell death in adult T-cell leukemia cells through estrogen receptors. Biosci. Biotechnol. Biochem. 2010, 74, 2113–2115. [Google Scholar] [CrossRef] [Green Version]
- García-Jiménez, C.; Gutiérrez-Salmerón, M.; Chocarro-Calvo, A.; García-Martinez, J.M.; Castaño, A.; De la Vieja, A. From obesity to diabetes and cancer: Epidemiological links and role of therapies. Br. J. Cancer 2016, 114, 716–722. [Google Scholar] [CrossRef] [Green Version]
- Tie, G.; Yan, J.; Khair, L.; Messina, J.A.; Deng, A.; Kang, J.; Fazzio, T.; Messina, L.M. Hypercholesterolemia Increases Colorectal Cancer Incidence by Reducing Production of NKT and γδ T Cells from Hematopoietic Stem Cells. Cancer Res. 2017, 77, 2351. [Google Scholar] [CrossRef] [Green Version]
- WHO. Obesity. Available online: https://www.who.int/health-topics/obesity#tab=tab_1 (accessed on 19 April 2020).
- Jakobek, L.; Matić, P. Non-covalent dietary fiber—Polyphenol interactions and their influence on polyphenol bioaccessibility. Trends Food Sci. Technol. 2019, 83, 235–247. [Google Scholar] [CrossRef]
- Ismaiel, M.; Yang, H.; Min, C. Dietary fiber role in type 2 diabetes prevention. Br. Food J. 2016, 118, 961–975. [Google Scholar] [CrossRef]
- Sefi, M.; Chaâbane, M.; Rafrafi, M.; Zeghal, N. Hypoglycemic and Hypolipidemic Activities of Aloe vera Leaf Mucilage in Alloxan-Induced Diabetic Rats. Pharm. Biomed. Res. 2019, 5. [Google Scholar] [CrossRef]
- Anbalahan, D. Pharmacological activity of mucilage isolated from medicinal plants. Diabetes 2017, 3, 98–113. [Google Scholar]
- Hajian, S.; Asgary, S.; Rafieian-Kopaei, M.; Sahebkar, A.; Goli-Malekabady, N.; Rashidi, B. Hibiscus esculentus seed and mucilage beneficial effects in reducing complications of diabetes in streptozotocin-induced diabetic rats. Ann. Res. Antioxid. 2016, 1, e23. [Google Scholar]
- Lin, D.; Xiao, M.; Zhao, J.; Li, Z.; Xing, B.; Li, X.; Kong, M.; Li, L.; Zhang, Q.; Liu, Y.; et al. An Overview of Plant Phenolic Compounds and Their Importance in Human Nutrition and Management of Type 2 Diabetes. Molecules 2016, 21, 1374. [Google Scholar] [CrossRef]
- Hoda, M.; Hemaiswarya, S.; Doble, M. Mechanisms of Action of Phenolic Phytochemicals in Diabetes Management. In Role of Phenolic Phytochemicals in Diabetes Management, 1st ed.; Hoda, M., Hemaiswarya, S., Doble, M., Eds.; Springer: Singapore, 2019; pp. 83–121. [Google Scholar]
- Aryaeian, N.; Sedehi, S.K.; Arablou, T. Polyphenols and their effects on diabetes management: A review. Med J. Islamic Repub. Iran. 2017, 31, 134. [Google Scholar] [CrossRef] [Green Version]
- Loy, T.H.; Spriggs, M.; Wickler, S. Direct evidence for human use of plants 28,000 years ago: Starch residues on stone artefacts from the northern Solomon Islands. Antiquity 1992, 66, 898. [Google Scholar] [CrossRef]
- Amadi, J.A. Glycaemic index of three cocoyam varieties consumed in imo state, Nigeria. J. Dietit. Assoc. Niger. 2017, 8, 96–103. [Google Scholar]
- Eleazu, C.O. The concept of low glycemic index and glycemic load foods as panacea for type 2 diabetes mellitus; prospects, challenges and solutions. Afr. Health Sci. 2016, 16, 468–479. [Google Scholar] [CrossRef] [Green Version]
- Washio, M.; Mori, M.; Sakauchi, F.; Watanabe, Y.; Ozasa, K.; Hayashi, K.; Miki, T.; Nakao, M.; Mikami, K.; Ito, Y. Risk factors for kidney cancer in a Japanese population: Findings from the JACC Study. J. Epidemiol. 2005, 15, S203–S211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Washio, M.; Mori, M. Risk factors for renal cell cancer in a Japanese population. Clin. Med. Oncol. 2009, 3, 71–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Washio, M.; Mori, M.; Mikami, K.; Miki, T.; Watanabe, Y.; Nakao, M.; Kubo, T.; Suzuki, K.; Ozasa, K.; Wakai, K. Risk factors for renal cell carcinoma in a Japanese population. Asian Pac. J. Cancer Prev. 2014, 15, 9065–9070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teramoto, T.; Yokode, M.; Iso, H.; Kitamura, A.; Shiomi, H.; Kimura, T.; Matsumoto, M.; Iida, M.; Sasaki, J.; Inoue, S.; et al. Comprehensive risk management for the prevention of cerebro-cardiovascular diseases in Japan. Hypertens. Res. 2017, 40, 847–855. [Google Scholar] [CrossRef] [Green Version]
- Kraft, P.; Schuhmann, M.K.; Garz, C.; Jandke, S.; Urlaub, D.; Mencl, S.; Zernecke, A.; Heinze, H.-J.; Carare, R.O.; Kleinschnitz, C. Hypercholesterolemia induced cerebral small vessel disease. PLoS ONE 2017, 12, e0182822. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutritional Composition | |||
|---|---|---|---|
| Principle * | Nutrient per 100 g of Dry Weight | ||
| Crude Taro | Cooked Taro | Baked Taro with Salt | |
| Water | 70.64 g | 63.8 g | 60.98 g |
| Energy | 112 kcal | 142 kcal | 144 kcal |
| Carbohydrates | 26.46 g | 34.6 g | 34.09 g |
| Protein | 1.5 g | 0.52 g | 1.93 g |
| Total fat | 0.20 g | 0.11 g | 0.26 g |
| Cholesterol | 0 mg | 0 mg | 0 mg |
| Dietary fibers | 4.1 g | 5.1 g | 5.3 g |
| Ash | 1.2 g | 0.97 g | na |
| Vitamins * | |||
| Folates | 0.022 mg | 0.019 mg | 0.023 mg |
| Niacin | 0.600 mg | 0.510 mg | 0.734 mg |
| Pantothenic acid | 0.303 mg | 0.336 mg | na |
| Pyridoxine | 0.283 mg | Na | na |
| Riboflavin | 0.025 mg | 0.028 mg | 0.031 mg |
| Thiamin | 0.095 mg | 0.107 mg | 0.110 mg |
| Vitamin A | 0.004 mg | 0.004 mg | 0.005 mg |
| Vitamin C | 4.5 mg | 5 mg | 4.3 mg |
| Vitamin E | 2.38 mg | 2.93 mg | 3.07 mg |
| Vitamin K | 0.001 mg | 0.0012 mg | 0.0013 mg |
| Electrolytes * | |||
| Sodium | 11 mg | 15 mg | 475 mg |
| Potassium | 591 mg | 484 mg | 762 mg |
| Minerals * | |||
| Calcium | 43 mg | 18 mg | 56 mg |
| Copper | 0.172 mg | 0.201 mg | 0.222 mg |
| Iron | 0.550 mg | 0.720 mg | 0.710 mg |
| Magnesium | 33 mg | 30 mg | 43 mg |
| Manganese | 0.383 mg | 0.449 mg | na |
| Selenium | 0.0007 mg | 0.0009 mg | 0.0009 mg |
| Zinc | 0.230 mg | 0.270 mg | 0.300 mg |
| Starch ** (g starch/100 g) | |||
| Total starch | 18.8 | 14.2 | na |
| Resistant Starch—RS | 5.2 | 2.1 | na |
| Slowly digestible starch—SDS | 13.6 (SDS+RDS) | 2.5 | na |
| Rapidly digestible starch—RDS | 9.6 | na | |
| Glycemic Index ** | na | Medium | Medium |
| Property | Taro Corm Preparation | Bioactive Compound Class | Active Principle | Screening | Biological Effect of Its Derivatives | Reference | Origin of Taro Corms |
|---|---|---|---|---|---|---|---|
| Antitumoral/Antimetastatic | poi extract | - | - | In vitro | Inhibition of rat YYT colon cancer cells (nearly 80–90%) | [41] | USA |
| Crude taro extract | Protein | Tarin | In vitro | Inhibition of murine breast cancer cell lines: 66.1 (54%), 410.4 (24%) and EpH4 (21%) | [42] | ||
| Inhibition of human breast cancer cell lines: MCF-7 (65%), MDA-MB-231 (26%), MCF10A (31%) | |||||||
| Reduction of PGE2 release (63%) and mRNA COX-1/2 expression | |||||||
| In vivo | Inhibition lung and heart colonization by murine lineages 66.1 and 410.4 (85–99% reduction) | ||||||
| In vitro | Inhibition of human hepatoma HepG2 cells (25%) | [43] | China | ||||
| Tarin nano-liposomal capsules | In vitro | Inhibition of human breast cancer MDA-MB-231 (41%) | [44] | Brazil | |||
| Inhibition of human Glioblastoma U87 MG lineage (65%) | |||||||
| Polysaccharide | Taro-4-I | In vivo | Inhibition of lung colonization by murine B16BL6 melanoma cells (96.2%) | [45] | Korea | ||
| Ethanolic crude taro extract | - | - | In vitro | Inhibition of various adult T-cells leukemia (ATL) lineages IC50 from 25 to 106.9 μg/mL | [46] | Japan | |
| Immunomodulatory | poi extract | - | - | In vitro | Proliferation of spleen cells (T, B and NK cells) | [41] | USA |
| Crude taro extract | Polysaccharide | Taro-4-I | In vitro | Activation of complement system by alternative and classical pathways | [45] | Korea | |
| Release of IL-6, IL-12 and TNF-α by macrophages | |||||||
| Activation of NK cells cytotoxicity against Yac-1 cells | |||||||
| TPS-1 and TPS-2 | In vitro | Activation of macrophage (RAW 264.7) with releasing of NO, IL-6 and TNF-α | [47] | China | |||
| Protein | Tarin | In vitro | Cytokine expression IL-2, IL1β, INF-γ and TNF-α | [43] | |||
| Total spleen cells proliferation | |||||||
| Proliferation of mice spleen cells | [48] | Brazil; Philippines; China | |||||
| Proliferation of mice bone marrow cells | [49] | ||||||
| Protection of BM mice granulocytic progenitor cells | [50] | Brazil | |||||
| Promotion of repopulation of BM granulocytic Gr1+ cells | |||||||
| In vivo | Attenuation of leukopenia in immunosuppressed mice | ||||||
| Enhancement of BM granulocytic progenitors | |||||||
| Proliferation/differentiation into mature BM granulocytes | |||||||
| Protection of BM erythroid cells from CY-cytotoxicity | |||||||
| 3.3-fold enhancement on 5th day (mice spleen cells) 4.1-fold enhancement on 5th day (mice Spleen B lymphocytes) | [48] | ||||||
| - | - | In vivo | Proliferation of total mice spleen cells in 5 days | [51] | |||
| Proliferation of B220+ lymphocytes from mice spleen in 5 or 10 days | |||||||
| Proliferation of total mice bone marrow cells in 10 days | |||||||
| In vitro | Proliferation of total mice spleen cells | ||||||
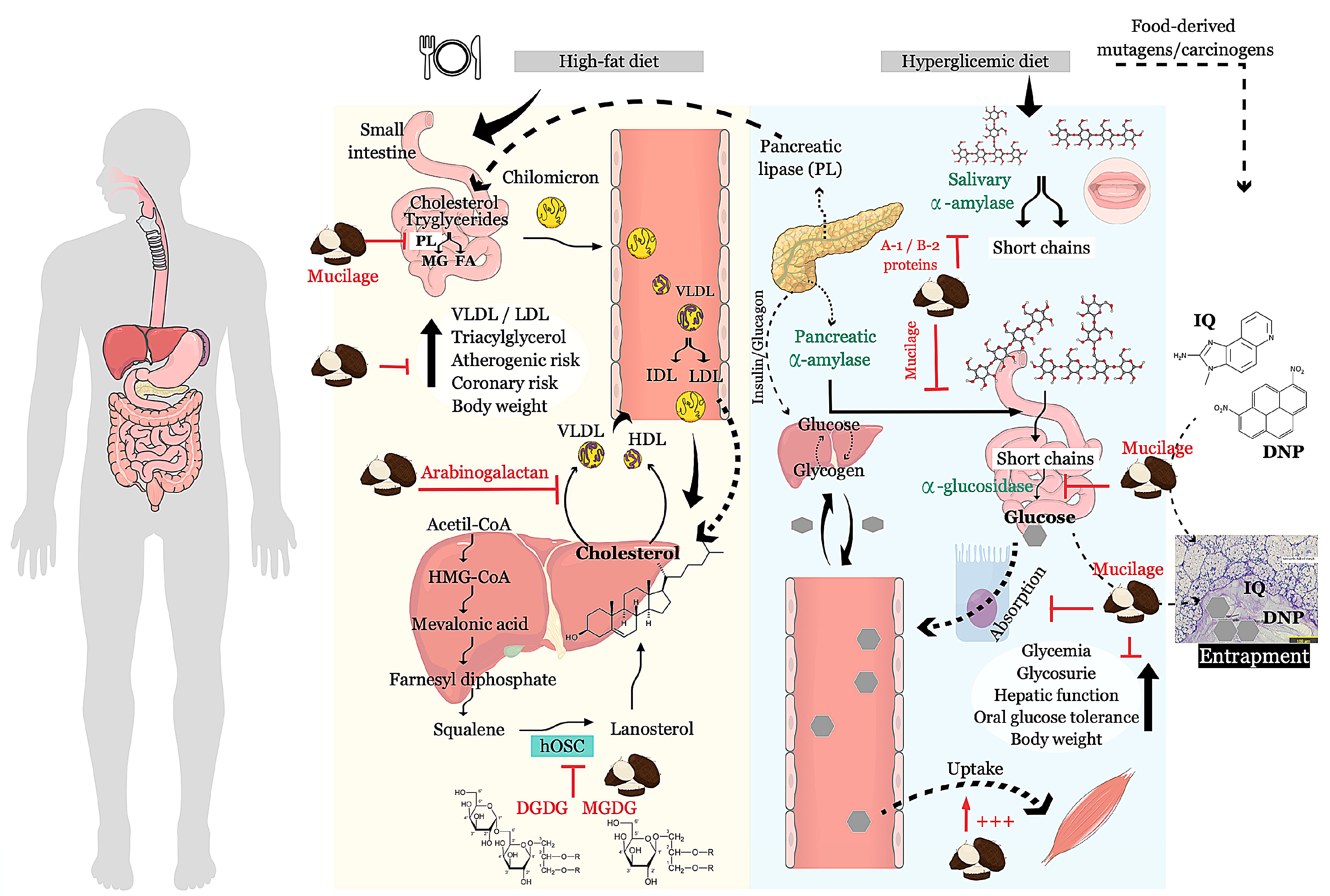
| Anti-hyperglycemic | Taro flour | Flavonoid; Alkaloid; saponin; tannin | - | In vivo | Decrease of blood glucose on hyperglycemic rats Amelioration of biochemical parameters in the urine Amelioration of liver, hepatic function and, body weight | [52,53,54] | Nigeria |
| Methanolic extract of taro flour | Alkaloid; flavonoid; steroid | Improvement of oral glucose tolerance | [55] | Bangladesh | |||
| Decrease of blood glucose levels (35.8%) | |||||||
| mucilage-rich extract from crude taro flour | Neutral sugar; protein, polyphenols | - | In vitro | Inhibition of alpha-amylase IC50 = 2.23 ± 0.24 mg/mL | [56] | South Africa | |
| Inhibition of alpha-glucosidase IC50 = 1.60 ± 0.17 mg/mL | |||||||
| Ex vivo | Inhibits glucose absorption in isolated rat jejunum IC50 = 1.82 ± 0.24 mg/mL | ||||||
| Enhances glucose uptake by rat psoas muscle GU50 = 0.78 ± 0.13 mg/mL | |||||||
| Extract from defatted crude taro flour | Protein | A-1 and B-2 | In vitro | Inhibition of human pancreatic (28.5 and 48.5%) and salivary alpha-amylase (62 and 56%) | [57] | ||
| Anti-hypercholesterolemic or Anti-hyperlipidemic | Taro flour | Flavonoid; Alkaloid; saponin; tannin | - | In vivo | Decrease of total cholesterol, VLDL-, LDL-cholesterol, triacylglycerol, atherogenic and coronary risk. Increase of HDL-cholesterol | [52,54] | Nigeria |
| Reduction of serum pancreatic lipase levels | |||||||
| Extract from cooked taro flour | - | Reduction of total blood cholesterol (36.41%) | [58] | Indonesia | |||
| mucilage-rich extract from crude taro flour | Neutral sugar; protein, polyphenols | - | In vitro | Inhibition of pancreatic lipase IC50 = 1.63 ± 0.15 mg/mL | [56] | South Africa | |
| Ethanolic extract from crude taro | Lipid | Extract | Inhibition (55%) of human lanosterol synthase (hOSC) | [59] | Japan | ||
| MGDG 1-3 | Inhibition (28–67%) of human lanosterol synthase (hOSC) | ||||||
| DGDG 1-4 | |||||||
| Mucilage-rich extract from taro flour | Polysaccharide | Arabinogalactan | In vivo | Decrease lipid levels in serum and tissues; decrease synthesis/secretion of apoB-containing lipoproteins, mainly VLDL, by hepatocytes. | [60] | India | |
| Anti-mutagenic | Dietary fiber-rich extract from crude taro | Polysaccharide | - | In vitro | Avoid mutation induced by DNP (87%) | [61] | Samoa |
| Crude taro extract | - | Avoid mutation by UV radiation with ID50 = 0.6 mg/plate | [62] | Japan | |||
| Heptane extract from cooked taro | Avoid mutation by IQ with IC50 = 200–500 μg/plate | [63] | New Zealand | ||||
| Antioxidant | See Table 3 | ||||||
| Taro Corm Origin | Taro Derivatives and Method of Extraction | Antioxidant Molecules | Methods Used for Antioxidant Evaluation | Quantification | Antioxidant Capacity | Ref. |
|---|---|---|---|---|---|---|
| Cameroon | Raw Taro Corms Acetone: water extraction | Polyphenols Chlorogenic acid; Catechin; Epicatechin; Epigallocatechin (flavan-3-01s); Gallic acid; Proanthocyanidins. | TLC and HPLC. | Non applicable | Non applicable | [69] |
| China | Raw Taro Corms Acetone: water: acetic acid extraction | Ascorbic acid; Violaxanthin; Lutein; β-Carotene; δ-γ-α-Tocopherol; δ-γ-Tocotrienol. | H-ORAC, Folin–Ciocalteu method (a) ascorbic acid determination (b) and HPLC evaluation. | (a). 1.8 mg GAE/g FW (b). 29.5 µg AA/g FW | 12.96 µmol TE/g FW | [70] |
| Egypt | 1. Fresh Taro Corm Ice Cream 2. Boiled Taro Corm Ice Cream | (a) Flavonoids (b) Tanins (c) Ascorbic acid (d) Carotenoids | Free-radical scavenging activity (RSA), total flavonoid contents and chemical composition of taro corms. | 1. (a). 88 μg/100 g (b). 0.92 (c). 31.54 mg/100 g (d). 328 μg/100 g 2. (a). 73 μg/100 g (b). 1.84 (c). 29.18 mg/100 g (d). 273 μg/100 g | CControl: 25.14% 11. 45.84% 22. 44.73% | [71] |
| Fiji | 1.Steamed corms 2. Unsteamedcorms Methanol extraction | Phenols Flavonoids | Folin-Ciocalteu method, aluminium chloride colorimetric method, DPPH, FRAP e ABTS. | TPC (mg GAE1)/g dry weight) 1. (a) 42.77 ± 3.39 2. (a) 32.32 ± 4.56 TFC (mg CE/g dry weight) 1. (a).12.68 ± 4.85 2. (a).10.24 ± 3.51 | DPPH (%) 1. (a) 4.82 ± 2.91 2. (a) 24.37 ± 4.23 FRAP (mM TE) 1. (a) 339.08 ± 20.50 2. (a) 224.72 ± 13.24 ABTS (%) 1. (a) 56.34 ± 3.54 2. (a) 42.33 ± 0.31 | [72] |
| India | Raw Taro Corms Methanol: acetone: water: acetic acid extraction | Phenols Tanins Condensed anthocyanins | Folin-Ciocalteu method, condensed tannins, total anthocyanins, DPPH, ABTS and FRAP. | TPC—0.887 ± 0.016 mg GAE/100 mg FW | DPPH—EC50 from 1.390 to 2.890 mg/mL | [73] |
| CTC—0.015 ± 0.001 mg CE/100 mg FW | ABTS—EC50 from 1.720 to 2.360 mg/mL | |||||
| TA—4.09 ± mg/100 mg FW | RPA -EC50 from 4410 to 5190 mg/mL | |||||
| Raw Taro Corms Methanol extraction | Alkaloids Tanins Terpenoids Flavonoids Fatty acids 9,12,15- octadecatrienoic Acid (?) Decanoic acid (?) 10 Fluoro trimethyl Ester (?) Pentadecanoic acid | Hager’s test and Wagner’s test, Ferric chloride test, Lead acetate test, Salkowski test and tannins, TLC, GC-MS and DPPH. | Non applicable | 78.73% of inhibition | [74] | |
| 1. Raw Taro Corms from Greenhouse Plants; 2. Micropropagated Plants; Methanol extraction | Phenols | DPPH and Folin-Ciocalteu method. | 1. 88.1 ± 2.0 mg/g 2. 93.5 ± 1.3 mg/g | 1. from 39 ± 1.2 to 92 ± 0.7 2. from 39 ± 1.2 to 92 ± 0.7 | [75] | |
| Raw Taro Corms Methanol extraction | Catechin Epi-catechin 1-O-feruloyl-D-glucoside 3, 5-DiCQ acid Vitexin Isovitexin Cyanidin-3-glucoside Luteolin-7-O-ruti-noside Vicenin-2-Cafeic acid Cyanidin-3-rhamnoside Chlorogenic acid Quercetin Hyperoside | Folin-Ciocalteu method, aluminium chloride colorimetric method, DPPH, FRAP, phos-phomolybdenum method and LC-MS. | TFC—10.78 mg RE/100 g | DPPH—21.80% | [76] | |
| TAA—63.47 mg AAE/100 g | ||||||
| TPC—14.17 mg GAE/100 g | ||||||
| FRAP—63.71 mg BHT Eq/100 g | ||||||
| Japan | Raw Taro Corms Methanol/water/acetic acid extraction | Polyphenolic compounds | H-ORAC (a), Folin-Ciocalteu method and DPPH (b). | 0.46 mg GAE/g | a. 10.10 mmol TE/g b. 2.47 mmol TE/g | [77] |
| Raw Taro Corms Methanol extraction | a. 8,11-octadecadienoic acid, methyl ester; b. hexadecanoic acid, methyl ester; c. 9,12,15-octadecatrienoic acid, methyl ester (Z,Z,Z); d. 9-octadecenoic acid, methyl ester (E); e 3,5-di-tert-butyl-4-trimethylsiloxytoluene; f. cyclohexanol, 2-nethyl-5-(1-methylethenyl)-(1.alpha.,2.beta.,5alpha). | GC-MS, DPPH. | a. 54.62% b. 20.55% c. 12.06% d. 06.42% e. 01.96% f. 01.88% | Strip > Leaf > Root | [78] | |
| Malaysia | 1. Raw taro flour 2. Boiled in Water 3. Fried in groundnut oil | (a) Flavonoids (b) Tannins (c) Alkaloids (d) Carotenoid Vitamins ThiamineRiboflavin | Flavonoid and tannin contents according to AOAC, Gravimetric method, DPPH and reducing power assay. | 1. (a). 0.88 ± 0.11% (b). 0.90 ± 0.00% (c). 3.68 ± 0.95% (d). 3.28 ± 0.04 μg/g 2. (a). 0.43 ± 0.32% (b). 1.83 ± 0.04% (c). 1.35 ± 0.92% (d). 1.73 ± 0.02 μg/g 3. (a). 0.38 ± 0.11% (b). 1.42 ± 0.02% (c). 1.03 ± 0.04% (d). 1.54 ± 0.02μg/g | DPPH (%) 1. 64.05 ± 2.50 2. 56.88 ± 3.77 3. 44.20 ± 3.10 Reducing power (nm) 1. 1.60 ± 0.33 2. 1.58 ± 0.37 3. 1.34 ± 0.30 | [79] |
| Nigeria | Rat Feed Taro Corm Based | a. Flavonoids b. Alkaloids c. Saponins d. Tannins | Gravimetric method of Harbone, DPPH and TLC. | a. 2.65% b. 1.01% c. 0.70% d. 1.06% | Moderate (+++) | [53] |
| Taro Corm Flour | Gravimetric method. | a. 0.64 ± 0.10% b. 0.37 ± 0.05% c. 0.51 ± 0.04% d. 0.28 ± 0.03% | Non applicable | [52] | ||
| Taro Corm Flour Methanol extraction | a. hexadecanoic acid methyl ester; b. octadecanoic acid; c. 12-octadecadienoyl chloride; d. 11-octadecenoic acid methyl ester; e. 9-octadecenoic acid; f. 3-hexadecyloxycarbonyl-5- (2-hydroxylethyl)-4-methylimidazolium ion; g. hexanedioic acid, bis(2-ethylhexyl) ester; h. 3, 5-di-t-butyl phenol. i. flavonoids—mg quercetin equivalent/g dry weight j. Phenols—mggGallic acid equivalent/g dry weight k. Condensed tanins—mg Catechin Equivalent/g dry weight | GC–MS, polyphenol assays, total flavonoids, total condensed tannins, DPPH and reducing power assay. | a. 0.43% b. 20.91% c. 0.77% d. 2.12% e. 64.37% f. 1.36% g. 1.36% h. 3.27% i. 8.50 ± 0.42 j. 15.15 ± 0.35 k. 4.40 ± 0.14 | 0.58 ± 0.36 nm | [80] | |
| 1. Raw taro corms 2. Taro powder 3. Taro-based noodles 4. Taro-based cookies | (a) Phenols—mg/100g expressed as g/catechin eq/g (b) Tanins—mg/100g expressed as vanillin eq/g (c) Flavonoids—mg/100g expressed as mg gallic acid eq/g (d) Saponins—mg/100g saponins | Lipid peroxidation (%), Folin-Ciocalteu method, Vanillin method, flavonoid content, saponin content. | 1. (a). 34.83 ± 0.28 (b). 32.24 ± 0.35 (c). 28.56 ± 0.23 (d). 14.22 ± 0.36 2. (a). 78.33 ± 0.66 (b).32.24 ± 0.35 (c). 64.23 ± 0.54 (d). 26.96 ± 0.61 3. (a). 16.27 ± 0.90 (b). Negative (c). 2.96 ± 0.323 (d). 5.01 ± 0.61 4. (a). 3.68 ± 0.10 (b). Negative (c). 0.90 ± 0.16 (d). 2.73 ± 0.18 | 1. 74.68 ± 0.44% LP 2. 81.77 ± 0.47% LP 3. 65.91 ± 0.27% LP 4. 28.00 ± 0.12% LP | [81] | |
| Philippines | 1. Aqueous extraction 2. Boiled taro corms 3. Ethanol extraction 4. Hexane extraction | Non-identified | % inhibition of lipid peroxidation. | Non applicable | 1. 100% 2. 100% 3. 70 ± 16% 4. 83 ± 7% | [82] |
| South Africa | Raw Taro Corms Methanol: water extraction | a. Flavonoids b. Phenols | Folin–Ciocalteu method, total flavonoid content, ABTS(c), DPPH (d) | a. 61 ± 9 CAE/100 g b. 205 ± 53 CAE/100 g | (c). 452 ± 72 mM TEAC/100 g (d). 244 ± 73 mM TEAC/100 g | [13] |
| Turkey | Raw Taro Corms Ethanolic extraction | a. Phenols b. Flavonoids | DPPH (c), ABTS (d), reducing power assay (e), Folin–Ciocalteu method and total flavonoid amount. | a. 2400 mg GAE/kg b. 2050 mg QE/kg | (c). Vitamin C > Trolox > C. esculenta > BHA—95.4, 93.6, 83.8 and 78.8% (d). Trolox = BHA > C. esculenta—100, 100 and 94.6% (e). BHA > Trolox > C. esculenta, | [83] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ribeiro Pereira, P.; Bertozzi de Aquino Mattos, É.; Nitzsche Teixeira Fernandes Corrêa, A.C.; Afonso Vericimo, M.; Margaret Flosi Paschoalin, V. Anticancer and Immunomodulatory Benefits of Taro (Colocasia esculenta) Corms, an Underexploited Tuber Crop. Int. J. Mol. Sci. 2021, 22, 265. https://doi.org/10.3390/ijms22010265
Ribeiro Pereira P, Bertozzi de Aquino Mattos É, Nitzsche Teixeira Fernandes Corrêa AC, Afonso Vericimo M, Margaret Flosi Paschoalin V. Anticancer and Immunomodulatory Benefits of Taro (Colocasia esculenta) Corms, an Underexploited Tuber Crop. International Journal of Molecular Sciences. 2021; 22(1):265. https://doi.org/10.3390/ijms22010265
Chicago/Turabian StyleRibeiro Pereira, Patrícia, Érika Bertozzi de Aquino Mattos, Anna Carolina Nitzsche Teixeira Fernandes Corrêa, Mauricio Afonso Vericimo, and Vania Margaret Flosi Paschoalin. 2021. "Anticancer and Immunomodulatory Benefits of Taro (Colocasia esculenta) Corms, an Underexploited Tuber Crop" International Journal of Molecular Sciences 22, no. 1: 265. https://doi.org/10.3390/ijms22010265
APA StyleRibeiro Pereira, P., Bertozzi de Aquino Mattos, É., Nitzsche Teixeira Fernandes Corrêa, A. C., Afonso Vericimo, M., & Margaret Flosi Paschoalin, V. (2021). Anticancer and Immunomodulatory Benefits of Taro (Colocasia esculenta) Corms, an Underexploited Tuber Crop. International Journal of Molecular Sciences, 22(1), 265. https://doi.org/10.3390/ijms22010265