
Essential Role of CRIM1 on Endometrial Receptivity in Goat

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
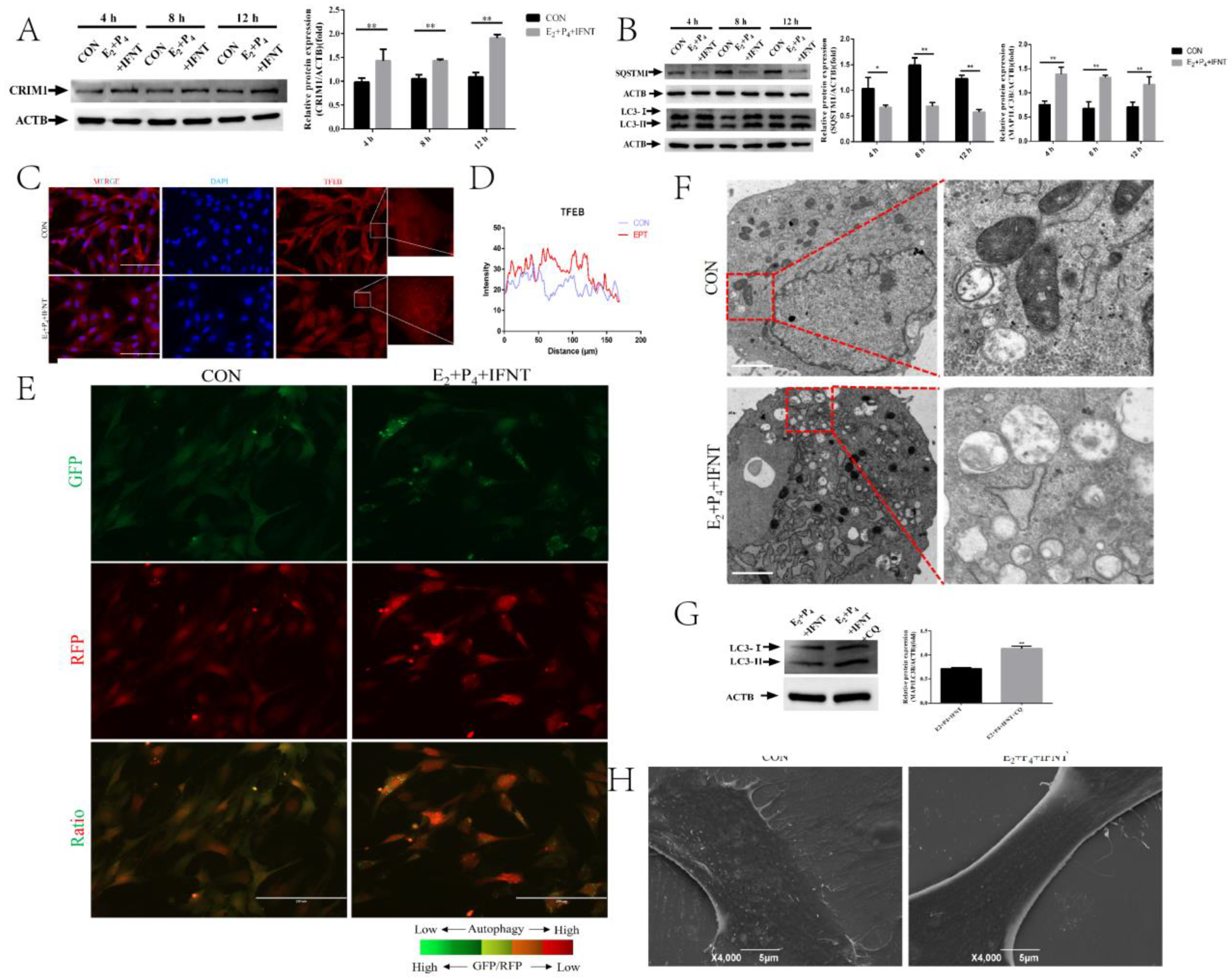
2.1. Hormones and IFN-τ Treatment Increased CRIM1 and Activated Autophagy
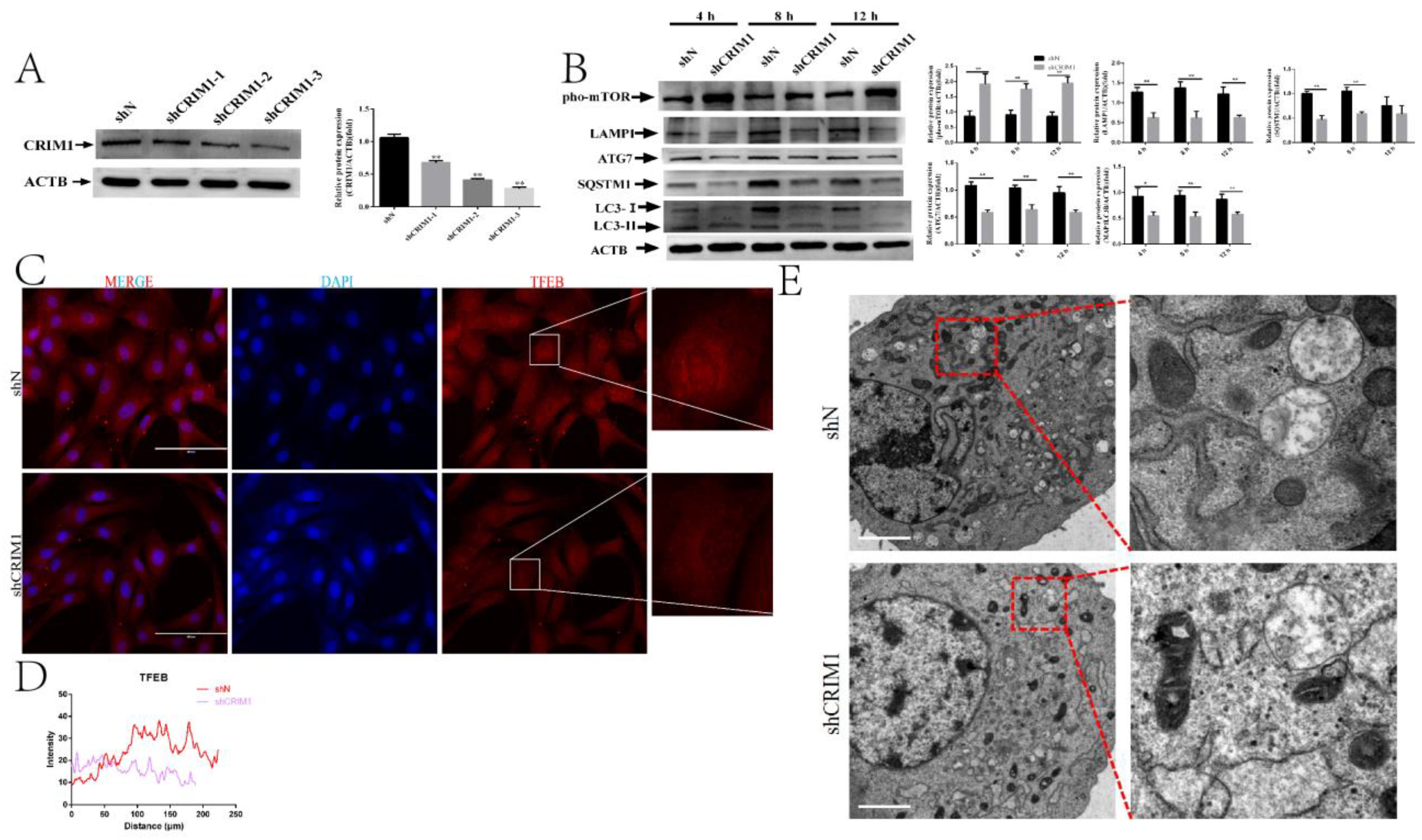
2.2. Knockdown of CRIM1 Inhibited Autophagy
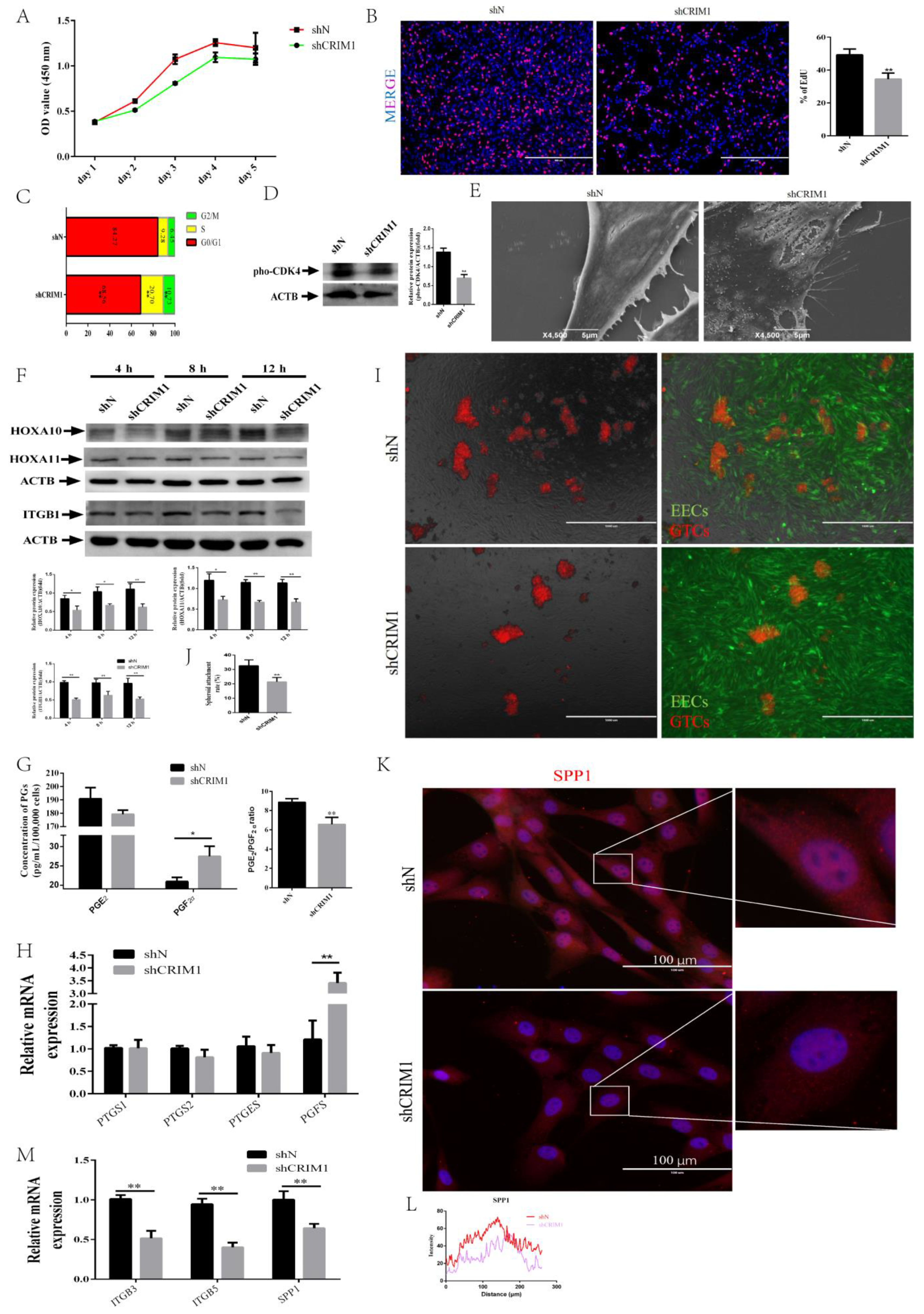
2.3. Knockdown of CRIM1 Jeopardized Endometrial Receptivity
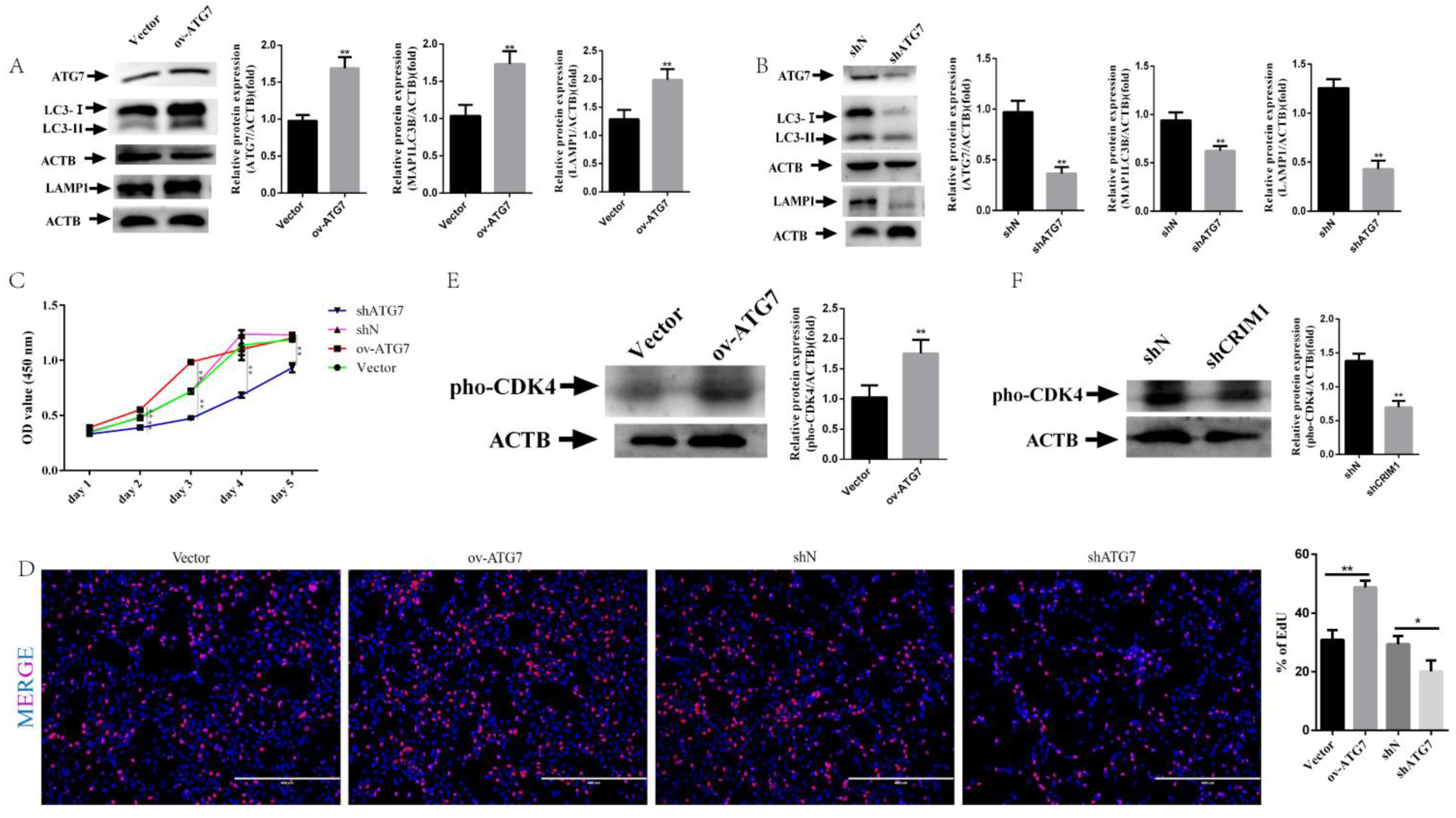
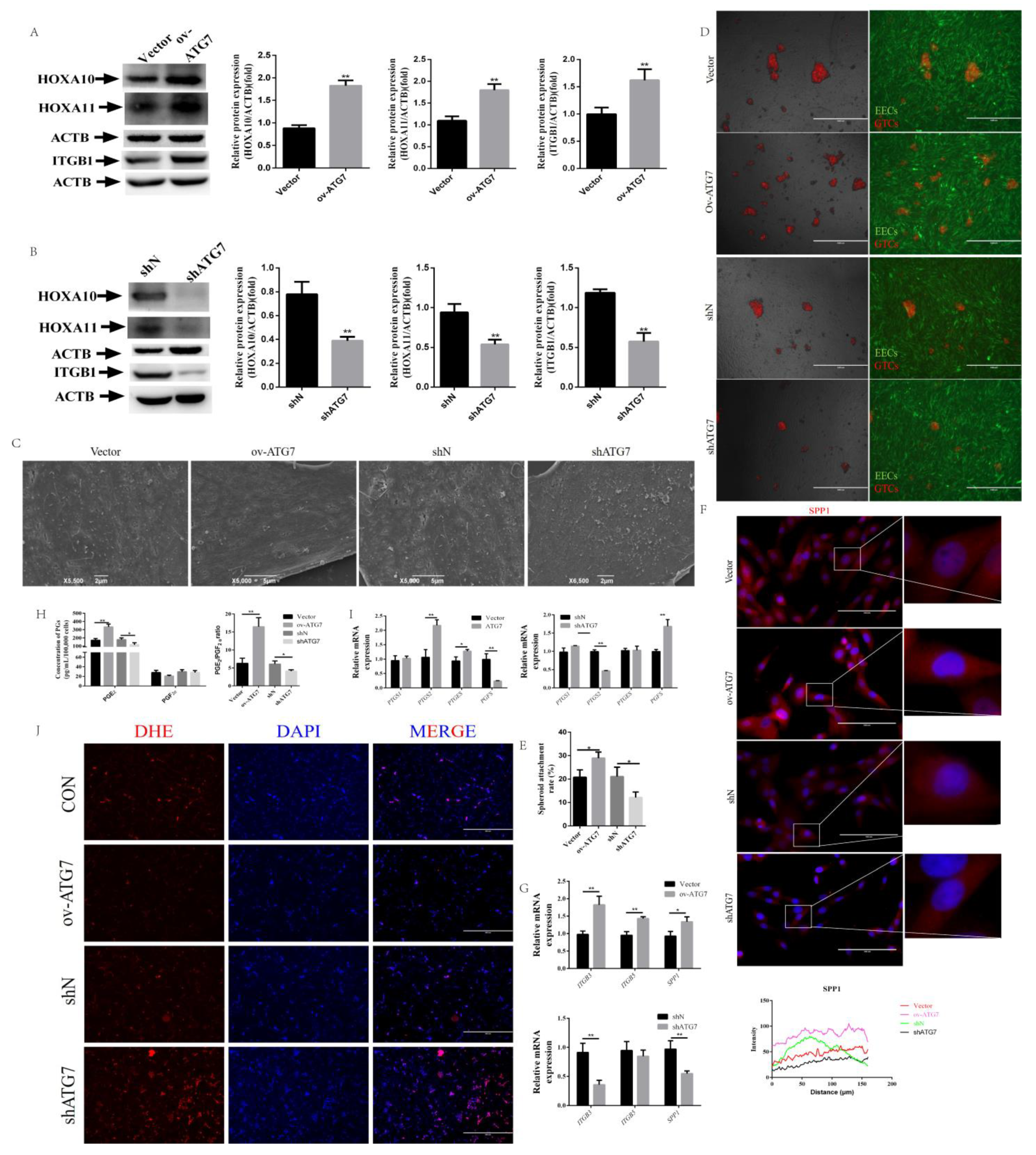
2.4. Overexpression of ATG7 Reversed Receptivity Defects Phenotype
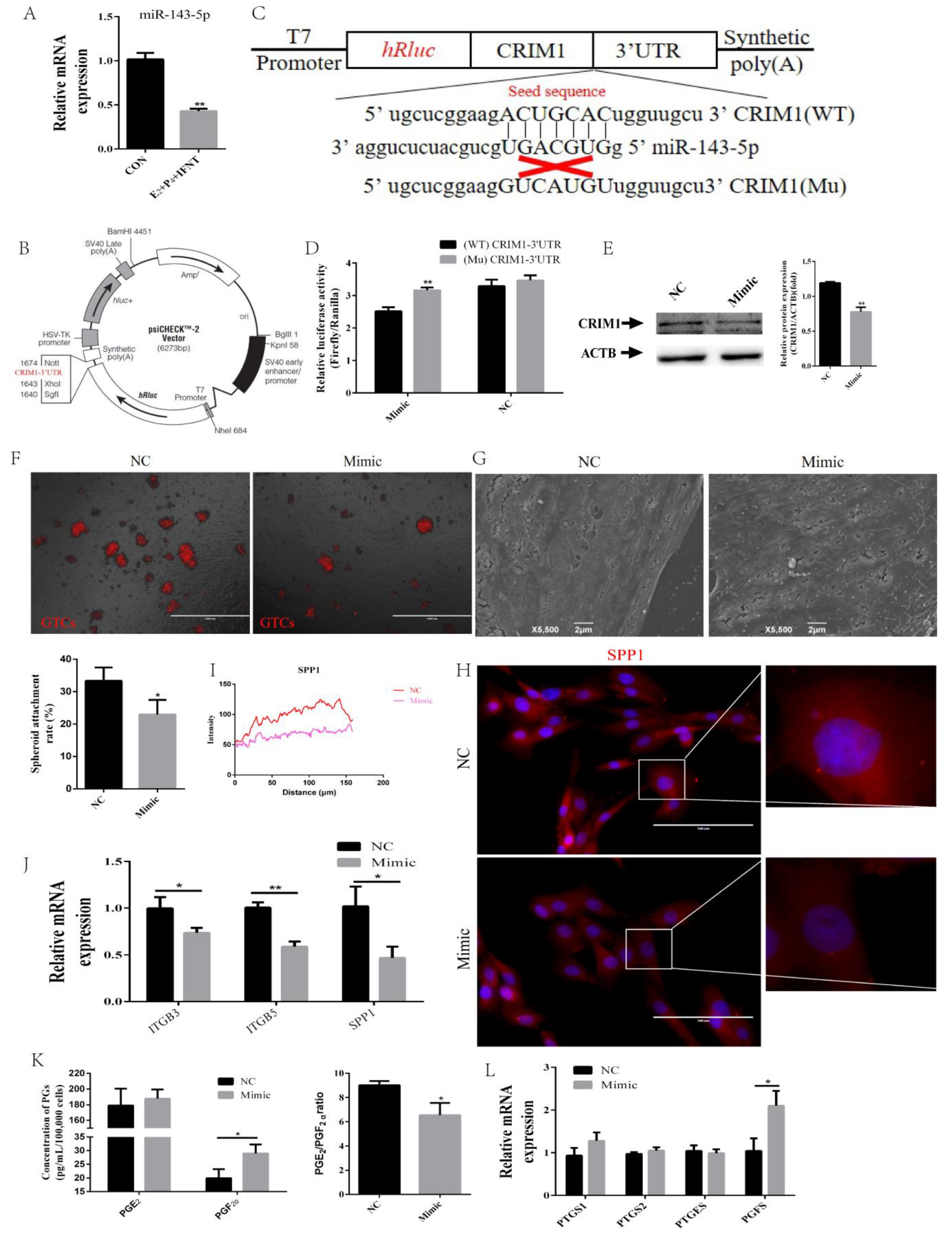
2.5. miR-143-5p Targeted the CRIM1 3′UTR to Regulate Endometrial Receptivity
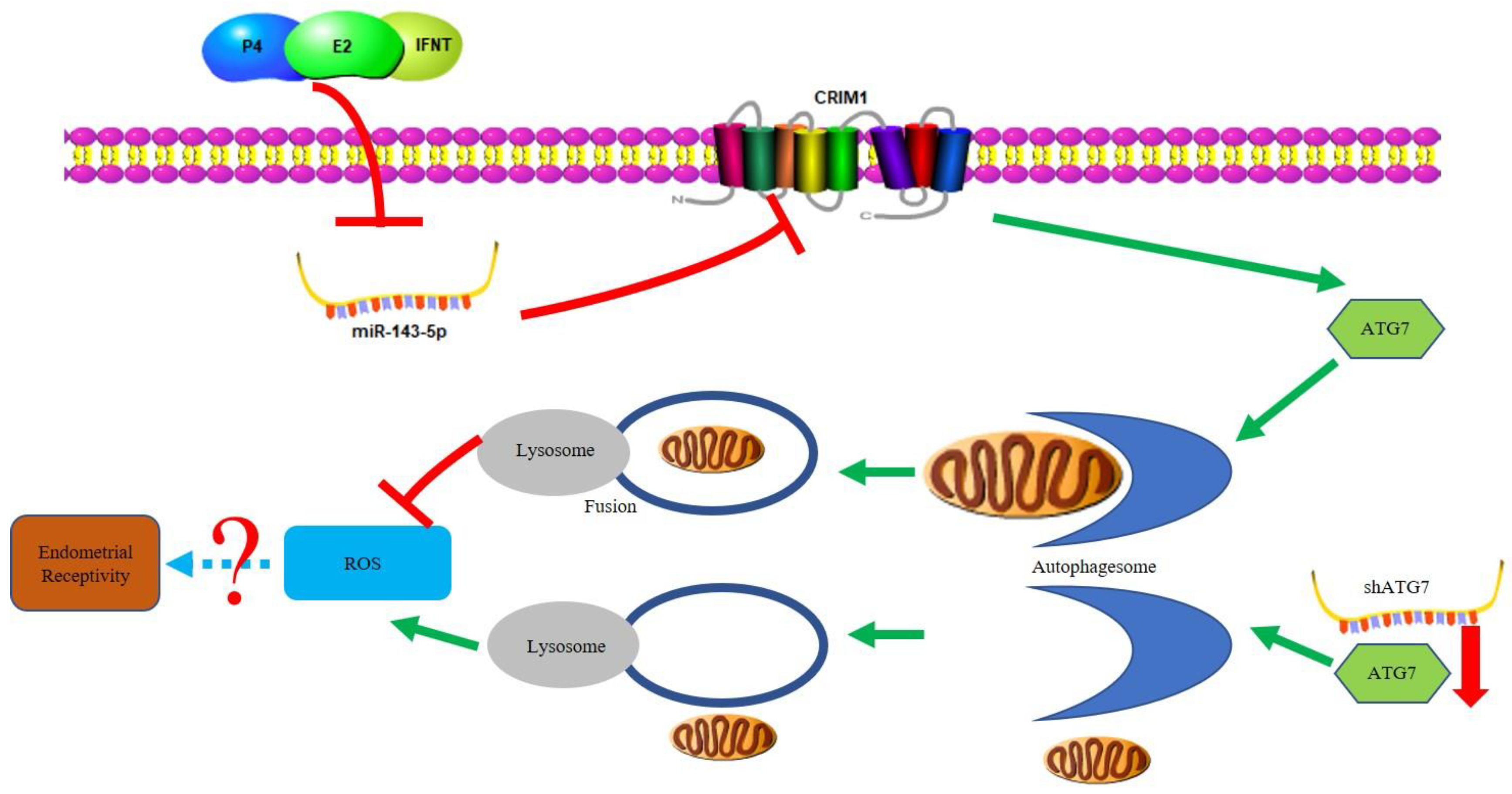
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Drug Treatment
4.2. Cell Transfection and Fluorescence Measurement
4.3. Spheroid Co-Culture Assay
4.4. RNA Extraction and Real-Time Quantitative PCR
4.5. Western Blot Analysis
4.6. Immunofluorescent Staining
4.7. Prostaglandins Measurement
4.8. Measurement of Cell Viability
4.9. Cell Cycle Analysis
4.10. EdU Proliferation Assay
4.11. Transmission Electron Microscopy (TEM)
4.12. Scanning Electron Microscopy (SEM)
4.13. Assessment of Reactive Oxygen Species (ROS) Generation
4.14. 3′-UTR Constructs/Luciferase Assay
4.15. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, Q.; Zhang, A.; Yu, F.; Gao, J.; Liu, Y.; Yu, C.; Zhou, H.; Xu, C. Label-free proteomics uncovers energy metabolism and focal adhesion regulations responsive for endometrium receptivity. J. Proteome Res. 2015, 14, 1831–1842. [Google Scholar] [CrossRef]
- Craciunas, L.; Gallos, I.; Chu, J.; Bourne, T.; Quenby, S.; Brosens, J.J.; Coomarasamy, A. Conventional and modern markers of endometrial receptivity: A systematic review and meta-analysis. Hum. Reprod. Update 2019, 25, 202–223. [Google Scholar] [CrossRef]
- Simmons, R.M.; Satterfield, M.C.; Welsh, T.H., Jr.; Bazer, F.W.; Spencer, T.E. HSD11B1, HSD11B2, PTGS2, and NR3C1 expression in the peri-implantation ovine uterus: Effects of pregnancy, progesterone, and interferon tau. Biol. Reprod. 2010, 82, 35–43. [Google Scholar] [CrossRef] [Green Version]
- Vasquez, Y.M.; Wang, X.; Wetendorf, M.; Franco, H.L.; Mo, Q.; Wang, T.; Lanz, R.B.; Young, S.L.; Lessey, B.A.; Spencer, T.E.; et al. FOXO1 regulates uterine epithelial integrity and progesterone receptor expression critical for embryo implantation. PLoS Genet. 2018, 14, e1007787. [Google Scholar] [CrossRef] [PubMed]
- Benson, G.V.; Lim, H.; Paria, B.C.; Satokata, I.; Dey, S.K.; Maas, R.L. Mechanisms of reduced fertility in Hoxa-10 mutant mice: Uterine homeosis and loss of maternal Hoxa-10 expression. Development 1996, 122, 2687–2696. [Google Scholar] [CrossRef]
- Gendron, R.L.; Paradis, H.; Hsieh-Li, H.M.; Lee, D.W.; Potter, S.S.; Markoff, E. Abnormal uterine stromal and glandular function associated with maternal reproductive defects in Hoxa-11 null mice. Biol. Reprod. 1997, 56, 1097–1105. [Google Scholar] [CrossRef]
- MacIntyre, D.M.; Lim, H.C.; Ryan, K.; Kimmins, S.; Small, J.A.; MacLaren, L.A. Implantation-associated changes in bovine uterine expression of integrins and extracellular matrix. Biol. Reprod. 2002, 66, 1430–1436. [Google Scholar] [CrossRef] [Green Version]
- Johnson, G.A.; Burghardt, R.C.; Bazer, F.W. Osteopontin: A leading candidate adhesion molecule for implantation in pigs and sheep. J. Anim. Sci. Biotechnol. 2014, 5, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, S.F.; Brown, J.K.; Campbell, L.L.; Koscielniak, M.; Oliver, C.; Wheelhouse, N.; Entrican, G.; McFee, S.; Wills, G.S.; McClure, M.O.; et al. Pelvic Chlamydial Infection Predisposes to Ectopic Pregnancy by Upregulating Integrin beta1 to Promote Embryo-tubal Attachment. EBioMedicine 2018, 29, 159–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, B.; Geerts, D.; Hu, S.; Yue, J.; Li, Z.; Zhu, G.; Jin, L. The depot GnRH agonist protocol improves the live birth rate per fresh embryo transfer cycle, but not the cumulative live birth rate in normal responders: A randomized controlled trial and molecular mechanism study. Hum. Reprod. 2020, 35, 1306–1318. [Google Scholar] [CrossRef] [PubMed]
- Paravati, R.; De Mello, N.; Onyido, E.K.; Francis, L.W.; Brusehafer, K.; Younas, K.; Spencer-Harty, S.; Conlan, R.S.; Gonzalez, D.; Margarit, L. Differential regulation of osteopontin and CD44 correlates with infertility status in PCOS patients. J. Mol. Med. 2020, 98, 1713–1725. [Google Scholar] [CrossRef]
- Altmae, S.; Esteban, F.J.; Stavreus-Evers, A.; Simon, C.; Giudice, L.; Lessey, B.A.; Horcajadas, J.A.; Macklon, N.S.; D’Hooghe, T.; Campoy, C.; et al. Guidelines for the design, analysis and interpretation of ‘omics’ data: Focus on human endometrium. Hum. Reprod. Update 2014, 20, 12–28. [Google Scholar] [CrossRef] [PubMed]
- Perez-Deben, S.; Bellver, J.; Alama, P.; Salsano, S.; Quinonero, A.; Sebastian-Leon, P.; Diaz-Gimeno, P.; Dominguez, F. iTRAQ comparison of proteomic profiles of endometrial receptivity. J. Proteom. 2019, 203, 103381. [Google Scholar] [CrossRef]
- Zhang, L.; An, X.P.; Liu, X.R.; Fu, M.Z.; Han, P.; Peng, J.Y.; Hou, J.X.; Zhou, Z.Q.; Cao, B.Y.; Song, Y.X. Characterization of the Transcriptional Complexity of the Receptive and Pre-receptive Endometria of Dairy Goats. Sci. Rep. 2015, 5, 14244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Liu, X.; Liu, J.; Ma, L.; Zhou, Z.; Song, Y.; Cao, B. The developmental transcriptome landscape of receptive endometrium during embryo implantation in dairy goats. Gene 2017, 633, 82–95. [Google Scholar] [CrossRef]
- Zhang, Y.; Fan, J.; Ho, J.W.; Hu, T.; Kneeland, S.C.; Fan, X.; Xi, Q.; Sellarole, M.A.; de Vries, W.N.; Lu, W.; et al. Crim1 regulates integrin signaling in murine lens development. Development 2016, 143, 356–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, J.; Ponferrada, V.G.; Sato, T.; Vemaraju, S.; Fruttiger, M.; Gerhardt, H.; Ferrara, N.; Lang, R.A. Crim1 maintains retinal vascular stability during development by regulating endothelial cell Vegfa autocrine signaling. Development 2014, 141, 448–459. [Google Scholar] [CrossRef] [Green Version]
- Iyer, S.; Chhabra, Y.; Harvey, T.J.; Wang, R.; Chiu, H.S.; Smith, A.G.; Thomas, W.G.; Pennisi, D.J.; Piper, M. CRIM1 is necessary for coronary vascular endothelial cell development and homeostasis. J. Mol. Histol. 2017, 48, 53–61. [Google Scholar] [CrossRef]
- Oestreich, A.K.; Chadchan, S.B.; Popli, P.; Medvedeva, A.; Rowen, M.N.; Stephens, C.S.; Xu, R.; Lydon, J.P.; Demayo, F.J.; Jungheim, E.S.; et al. The Autophagy Gene Atg16L1 is Necessary for Endometrial Decidualization. Endocrinology 2020, 161. [Google Scholar] [CrossRef] [PubMed]
- Popova, N.V.; Jucker, M. The Role of mTOR Signaling as a Therapeutic Target in Cancer. Int. J. Mol. Sci. 2021, 22, 1743. [Google Scholar] [CrossRef] [PubMed]
- Arakawa, S.; Honda, S.; Yamaguchi, H.; Shimizu, S. Molecular mechanisms and physiological roles of Atg5/Atg7-independent alternative autophagy. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2017, 93, 378–385. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Vargas, J.M.; Ruiz-Magana, M.J.; Ruiz-Ruiz, C.; Majuelos-Melguizo, J.; Peralta-Leal, A.; Rodriguez, M.I.; Munoz-Gamez, J.A.; de Almodovar, M.R.; Siles, E.; Rivas, A.L.; et al. ROS-induced DNA damage and PARP-1 are required for optimal induction of starvation-induced autophagy. Cell Res. 2012, 22, 1181–1198. [Google Scholar] [CrossRef] [Green Version]
- Hansen, M.; Rubinsztein, D.C.; Walker, D.W. Autophagy as a promoter of longevity: Insights from model organisms. Nat. Rev. Mol. Cell Biol. 2018, 19, 579–593. [Google Scholar] [CrossRef] [PubMed]
- Lotfi, P.; Tse, D.Y.; Di Ronza, A.; Seymour, M.L.; Martano, G.; Cooper, J.D.; Pereira, F.A.; Passafaro, M.; Wu, S.M.; Sardiello, M. Trehalose reduces retinal degeneration, neuroinflammation and storage burden caused by a lysosomal hydrolase deficiency. Autophagy 2018, 14, 1419–1434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eskelinen, E.L. Roles of LAMP-1 and LAMP-2 in lysosome biogenesis and autophagy. Mol. Asp. Med. 2006, 27, 495–502. [Google Scholar] [CrossRef]
- Choi, J.; Jo, M.; Lee, E.; Oh, Y.K.; Choi, D. The role of autophagy in human endometrium. Biol. Reprod. 2012, 86, 70. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.L.; Xu, J.; Zhang, S.S.; Zhu, Q.Y.; Jin, B.H.; ZhuGe, D.L.; Shen, B.X.; Wu, X.Q.; Xiao, J.; Zhao, Y.Z. Temperature-sensitive heparin-modified poloxamer hydrogel with affinity to KGF facilitate the morphologic and functional recovery of the injured rat uterus. Drug Deliv. 2017, 24, 867–881. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Li, J.; Xu, G.; Zhang, R.; Zhou, C.; Qian, Y.; Liu, Y.; Chen, L.; Zhu, B.; Ye, X.; et al. Follicle-stimulating hormone promotes age-related endometrial atrophy through cross-talk with transforming growth factor beta signal transduction pathway. Aging Cell 2015, 14, 284–287. [Google Scholar] [CrossRef]
- Yang, S.; Wang, H.; Li, D.; Li, M. Role of Endometrial Autophagy in Physiological and Pathophysiological Processes. J. Cancer 2019, 10, 3459–3471. [Google Scholar] [CrossRef]
- Yang, D.; Jiang, T.; Liu, J.; Hong, J.; Lin, P.; Chen, H.; Zhou, D.; Tang, K.; Wang, A.; Jin, Y. Hormone regulates endometrial function via cooperation of endoplasmic reticulum stress and mTOR-autophagy. J. Cell. Physiol. 2018, 233, 6644–6659. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Jiang, T.; Liu, J.; Zhang, B.; Lin, P.; Chen, H.; Zhou, D.; Tang, K.; Wang, A.; Jin, Y. CREB3 regulatory factor -mTOR-autophagy regulates goat endometrial function during early pregnancy. Biol. Reprod. 2018, 98, 713–721. [Google Scholar] [CrossRef]
- Yang, D.; Zhang, B.; Wang, Z.; Zhang, L.; Chen, H.; Zhou, D.; Tang, K.; Wang, A.; Lin, P.; Jin, Y. COPS5 negatively regulates goat endometrial function via the ERN1 and mTOR-autophagy pathways during early pregnancy. J. Cell. Physiol. 2019, 234, 18666–18678. [Google Scholar] [CrossRef]
- Gao, K.; Wang, P.; Peng, J.; Xue, J.; Chen, K.; Song, Y.; Wang, J.; Li, G.; An, X.; Cao, B. Regulation and function of runt-related transcription factors (RUNX1 and RUNX2) in goat granulosa cells. J. Steroid Biochem. Mol. Biol. 2018, 181, 98–108. [Google Scholar] [CrossRef] [PubMed]
- An, X.; Liu, X.; Zhang, L.; Liu, J.; Zhao, X.; Chen, K.; Ma, H.; Li, G.; Cao, B.; Song, Y. MiR-449a regulates caprine endometrial stromal cell apoptosis and endometrial receptivity. Sci. Rep. 2017, 7, 12248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Liu, X.; Liu, J.; Ma, X.; Zhou, Z.; Song, Y.; Cao, B. miR-26a promoted endometrial epithelium cells (EECs) proliferation and induced stromal cells (ESCs) apoptosis via the PTEN-PI3K/AKT pathway in dairy goats. J. Cell. Physiol. 2018, 233, 4688–4706. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; An, X.; Zhang, L.; Fu, M.; Peng, J.; Han, P.; Hou, J.; Zhou, Z.; Cao, B. Identification and profiling of microRNAs in goat endometrium during embryo implantation. PLoS ONE 2015, 10, e0122202. [Google Scholar] [CrossRef]
- Tian, S.; Su, X.; Qi, L.; Jin, X.H.; Hu, Y.; Wang, C.L.; Ma, X.; Xia, H.F. MiR-143 and rat embryo implantation. Biochim. Biophys. Acta 2015, 1850, 708–721. [Google Scholar] [CrossRef]
- Nakamura, K.; Kusama, K.; Ideta, A.; Imakawa, K.; Hori, M. IFNT-independent effects of intrauterine extracellular vesicles (EVs) in cattle. Reproduction 2020, 159, 503–511. [Google Scholar] [CrossRef]
- Kaizuka, T.; Morishita, H.; Hama, Y.; Tsukamoto, S.; Matsui, T.; Toyota, Y.; Kodama, A.; Ishihara, T.; Mizushima, T.; Mizushima, N. An Autophagic Flux Probe that Releases an Internal Control. Mol. Cell 2016, 64, 835–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, H.; Yancey, P.G.; Blakemore, J.L.; Zhang, Y.; Ding, L.; Jerome, W.G.; Brown, J.D.; Vickers, K.C.; Linton, M.F. Macrophage SR-BI modulates autophagy via VPS34 complex and PPARalpha transcription of Tfeb in atherosclerosis. J. Clin. Investig. 2021, 131. [Google Scholar] [CrossRef]
- Juncadella, I.J.; Kadl, A.; Sharma, A.K.; Shim, Y.M.; Hochreiter-Hufford, A.; Borish, L.; Ravichandran, K.S. Apoptotic cell clearance by bronchial epithelial cells critically influences airway inflammation. Nature 2013, 493, 547–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubin, S.M.; Sage, J.; Skotheim, J.M. Integrating Old and New Paradigms of G1/S Control. Mol. Cell 2020, 80, 183–192. [Google Scholar] [CrossRef]
- Raspe, E.; Coulonval, K.; Pita, J.M.; Paternot, S.; Rothe, F.; Twyffels, L.; Brohee, S.; Craciun, L.; Larsimont, D.; Kruys, V.; et al. CDK4 phosphorylation status and a linked gene expression profile predict sensitivity to palbociclib. EMBO Mol. Med. 2017, 9, 1052–1066. [Google Scholar] [CrossRef] [PubMed]
- Rarani, F.Z.; Borhani, F.; Rashidi, B. Endometrial pinopode biomarkers: Molecules and microRNAs. J. Cell. Physiol. 2018, 233, 9145–9158. [Google Scholar] [CrossRef]
- Ott, T.L. Symposium review: Immunological detection of the bovine conceptus during early pregnancy. J. Dairy Sci. 2019, 102, 3766–3777. [Google Scholar] [CrossRef] [Green Version]
- Qiao, Z.; Xu, Z.; Xiao, Q.; Yang, Y.; Ying, J.; Xiang, L.; Zhang, C. Dysfunction of ATG7-dependent autophagy dysregulates the antioxidant response and contributes to oxidative stress-induced biological impairments in human epidermal melanocytes. Cell Death Discov. 2020, 6, 31. [Google Scholar] [CrossRef]
- Georgas, K.; Bowles, J.; Yamada, T.; Koopman, P.; Little, M.H. Characterisation of Crim1 expression in the developing mouse urogenital tract reveals a sexually dimorphic gonadal expression pattern. Dev. Dyn. 2000, 219, 582–587. [Google Scholar] [CrossRef]
- Tam, O.H.; Pennisi, D.; Wilkinson, L.; Little, M.H.; Wazin, F.; Wan, V.L.; Lovicu, F.J. Crim1 is required for maintenance of the ocular lens epithelium. Exp. Eye Res. 2018, 170, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Ka, H.; Seo, H.; Choi, Y.; Yoo, I.; Han, J. Endometrial response to conceptus-derived estrogen and interleukin-1β at the time of implantation in pigs. J. Anim. Sci. Biotechnol. 2018, 9, 44. [Google Scholar] [CrossRef]
- D’Occhio, M.J.; Campanile, G.; Zicarelli, L.; Visintin, J.A.; Baruselli, P.S. Adhesion molecules in gamete transport, fertilization, early embryonic development, and implantation-role in establishing a pregnancy in cattle: A review. Mol. Reprod. Dev. 2020, 87, 206–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazer, F.W.; Wu, G.; Johnson, G.A.; Kim, J.; Song, G. Uterine histotroph and conceptus development: Select nutrients and secreted phosphoprotein 1 affect mechanistic target of rapamycin cell signaling in ewes. Biol. Reprod. 2011, 85, 1094–1107. [Google Scholar] [CrossRef] [Green Version]
- Arosh, J.A.; Banu, S.K.; McCracken, J.A. Novel concepts on the role of prostaglandins on luteal maintenance and maternal recognition and establishment of pregnancy in ruminants. J. Dairy Sci. 2016, 99, 5926–5940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wathes, D.C.; Abayasekara, D.R.; Aitken, R.J. Polyunsaturated fatty acids in male and female reproduction. Biol. Reprod. 2007, 77, 190–201. [Google Scholar] [CrossRef]
- Zhang, B.; He, L.; Liu, Y.; Zhang, J.; Zeng, Q.; Wang, S.; Fan, Z.; Fang, F.; Chen, L.; Lv, Y.; et al. Prostaglandin E2 Is Required for BMP4-Induced Mesoderm Differentiation of Human Embryonic Stem Cells. Stem Cell Rep. 2018, 10, 905–919. [Google Scholar] [CrossRef] [Green Version]
- Devis-Jauregui, L.; Eritja, N.; Davis, M.L.; Matias-Guiu, X.; Llobet-Navàs, D. Autophagy in the physiological endometrium and cancer. Autophagy 2020. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Yang, H.L.; Zhou, W.J.; Lai, Z.Z.; Qiu, X.M.; Fu, Q.; Zhao, J.Y.; Wang, J.; Li, D.J.; Li, M.Q. Rapamycin prevents spontaneous abortion by triggering decidual stromal cell autophagy-mediated NK cell residence. Autophagy 2020, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Toschi, P.; Czernik, M.; Zacchini, F.; Fidanza, A.; Loi, P.; Ptak, G.E. Evidence of Placental Autophagy during Early Pregnancy after Transfer of In Vitro Produced (IVP) Sheep Embryos. PLoS ONE 2016, 11, e0157594. [Google Scholar] [CrossRef]
- James, R.E.; Broihier, H.T. Crimpy inhibits the BMP homolog Gbb in motoneurons to enable proper growth control at the Drosophila neuromuscular junction. Development 2011, 138, 3273–3286. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Kitami, M.; Pan, H.; Nakamura, M.T.; Zhang, H.; Liu, F.; Zhu, L.; Komatsu, Y.; Mishina, Y. Augmented BMP signaling commits cranial neural crest cells to a chondrogenic fate by suppressing autophagic beta-catenin degradation. Sci. Signal. 2021, 14. [Google Scholar] [CrossRef] [PubMed]
- Towers, C.G.; Thorburn, A. Circumventing autophagy inhibition. Cell Cycle 2019, 18, 3421–3431. [Google Scholar] [CrossRef] [PubMed]
- Aoki, A.; Nakashima, A.; Kusabiraki, T.; Ono, Y.; Yoshino, O.; Muto, M.; Kumasawa, K.; Yoshimori, T.; Ikawa, M.; Saito, S. Trophoblast-Specific Conditional Atg7 Knockout Mice Develop Gestational Hypertension. Am. J. Pathol. 2018, 188, 2474–2486. [Google Scholar] [CrossRef] [Green Version]
- Hamanaka, R.B.; Chandel, N.S. Mitochondrial reactive oxygen species regulate cellular signaling and dictate biological outcomes. Trends Biochem. Sci. 2010, 35, 505–513. [Google Scholar] [CrossRef] [Green Version]
- Noh, S.; Go, A.; Kim, D.B.; Park, M.; Jeon, H.W.; Kim, B. Role of Antioxidant Natural Products in Management of Infertility: A Review of Their Medicinal Potential. Antioxidants 2020, 9, 957. [Google Scholar] [CrossRef] [PubMed]
- Muralimanoharan, S.; Maloyan, A.; Myatt, L. Mitochondrial function and glucose metabolism in the placenta with gestational diabetes mellitus: Role of miR-143. Clin. Sci. 2016, 130, 931–941. [Google Scholar] [CrossRef] [Green Version]
- Yuan, D.-Z.; Lei, Y.; Zhao, D.; Pan, J.-L.; Zhao, Y.-B.; Nie, L.; Liu, M.; Long, Y.; Zhang, J.-H.; Yue, L.-M. Progesterone-Induced miR-145/miR-143 Inhibits the Proliferation of Endometrial Epithelial Cells. Reprod. Sci. 2019, 26, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Gomez, Y.; Organista-Nava, J.; Ocadiz-Delgado, R.; Garcia-Villa, E.; Leyva-Vazquez, M.A.; Illades-Aguiar, B.; Lambert, P.F.; Garcia-Carranca, A.; Gariglio, P. The expression of miR-21 and miR-143 is deregulated by the HPV16 E7 oncoprotein and 17beta-estradiol. Int. J. Oncol. 2016, 49, 549–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.Y.; Wang, A.H.; Wu, Q.X.; Sheng, H.X.; Jin, Y.P. Establishment and Characteristics of Immortal Goat Endometrial Epithelial Cells and Stromal Cells with hTERT. J. Anim. Vet. Adv. 2010, 9, 2738–2747. [Google Scholar] [CrossRef]
- Dong, F.; Huang, Y.; Li, W.; Zhao, X.; Zhang, W.; Du, Q.; Zhang, H.; Song, X.; Tong, D. The isolation and characterization of a telomerase immortalized goat trophoblast cell line. Placenta 2013, 34, 1243–1250. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lin, P.; Yin, Y.; Zhou, J.; Lei, L.; Zhou, X.; Jin, Y.; Wang, A. Brucella suis vaccine strain S2-infected immortalized caprine endometrial epithelial cell lines induce non-apoptotic ER-stress. Cell Stress Chaperones 2015, 20, 399–409. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Liu, A.; Wu, Y.; Li, B.; Nan, S.; Yin, R.; Zhu, H.; Chen, J.; Ding, Y.; Ding, M. BCL2L15 Depletion Inhibits Endometrial Receptivity via the STAT1 Signaling Pathway. Genes 2020, 11, 816. [Google Scholar] [CrossRef]
- Yang, D.; Jiang, T.; Lin, P.; Chen, H.; Wang, L.; Wang, N.; Zhao, F.; Tang, K.; Zhou, D.; Wang, A.; et al. Apoptosis inducing factor gene depletion inhibits zearalenone-induced cell death in a goat Leydig cell line. Reprod. Toxicol. 2017, 67, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Wang, L.; Lin, P.; Jiang, T.; Wang, N.; Zhao, F.; Chen, H.; Tang, K.; Zhou, D.; Wang, A.; et al. An immortalized steroidogenic goat granulosa cell line as a model system to study the effect of the endoplasmic reticulum (ER)-stress response on steroidogenesis. J. Reprod. Dev. 2017, 63, 27–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Luo, Z.; Yan, C.Y.; Wang, X.H.; He, Z.J.; Ouyang, S.H.; Yan, C.; Liu, L.F.; Zhou, Q.Q.; Mu, H.L.; et al. Autophagic degradation of PML promotes susceptibility to HSV-1 by stress-induced corticosterone. Theranostics 2020, 10, 9032–9049. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, D.; Liu, A.; Zhang, Y.; Nan, S.; Yin, R.; Lei, Q.; Zhu, H.; Chen, J.; Han, L.; Ding, M.; et al. Essential Role of CRIM1 on Endometrial Receptivity in Goat. Int. J. Mol. Sci. 2021, 22, 5323. https://doi.org/10.3390/ijms22105323
Yang D, Liu A, Zhang Y, Nan S, Yin R, Lei Q, Zhu H, Chen J, Han L, Ding M, et al. Essential Role of CRIM1 on Endometrial Receptivity in Goat. International Journal of Molecular Sciences. 2021; 22(10):5323. https://doi.org/10.3390/ijms22105323
Chicago/Turabian StyleYang, Diqi, Ai Liu, Yanyan Zhang, Sha Nan, Ruiling Yin, Qianghui Lei, Hongmei Zhu, Jianguo Chen, Li Han, Mingxing Ding, and et al. 2021. "Essential Role of CRIM1 on Endometrial Receptivity in Goat" International Journal of Molecular Sciences 22, no. 10: 5323. https://doi.org/10.3390/ijms22105323
APA StyleYang, D., Liu, A., Zhang, Y., Nan, S., Yin, R., Lei, Q., Zhu, H., Chen, J., Han, L., Ding, M., & Ding, Y. (2021). Essential Role of CRIM1 on Endometrial Receptivity in Goat. International Journal of Molecular Sciences, 22(10), 5323. https://doi.org/10.3390/ijms22105323