
The Role of ZEB2 in Human CD8 T Lymphocytes: Clinical and Cellular Immune Profiling in Mowat–Wilson Syndrome

 , ,
, , 
Abstract
:1. Introduction
2. Results
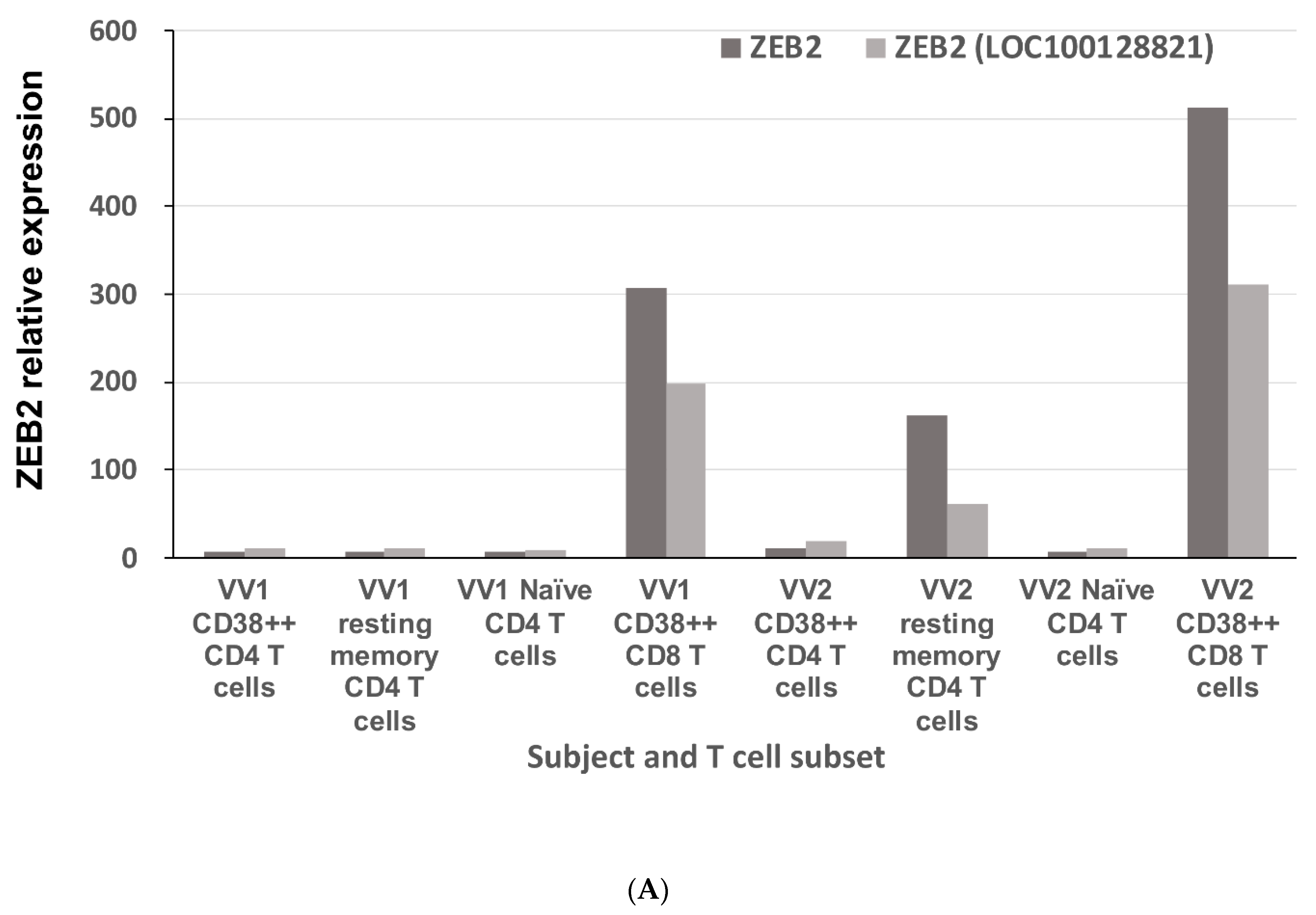
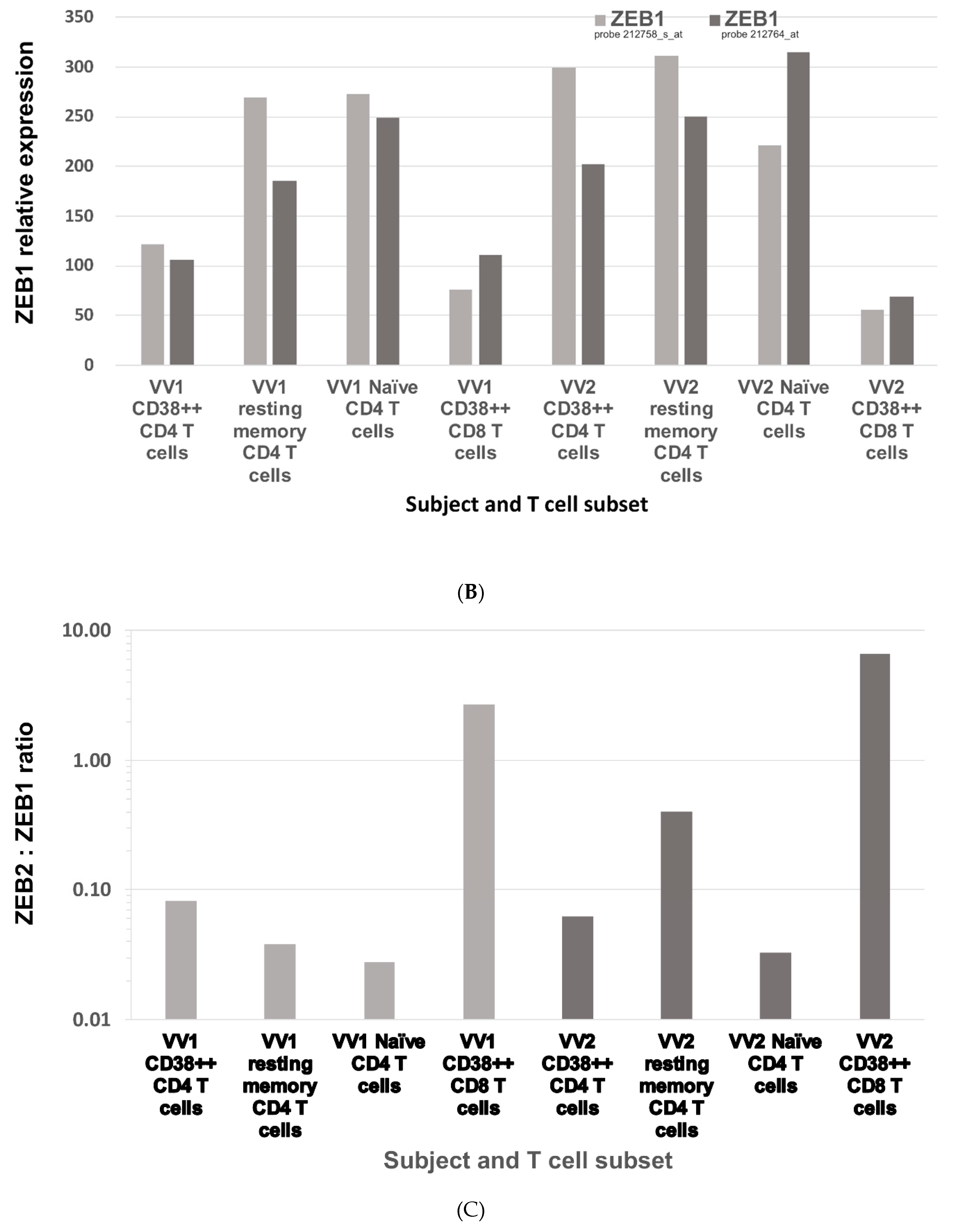
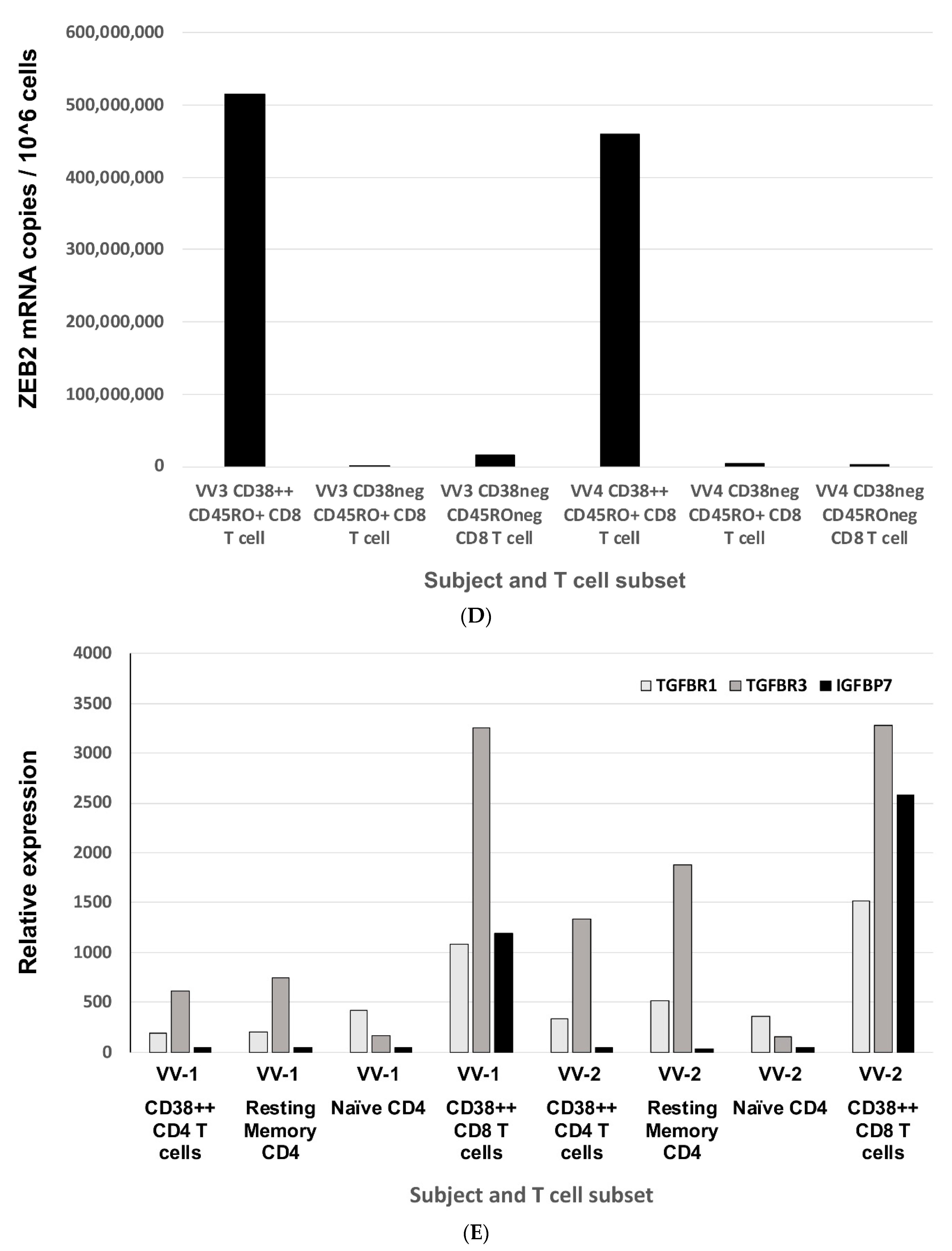
2.1. ZEB2 Expression in Early Antiviral Human Effector CD8 T Cells
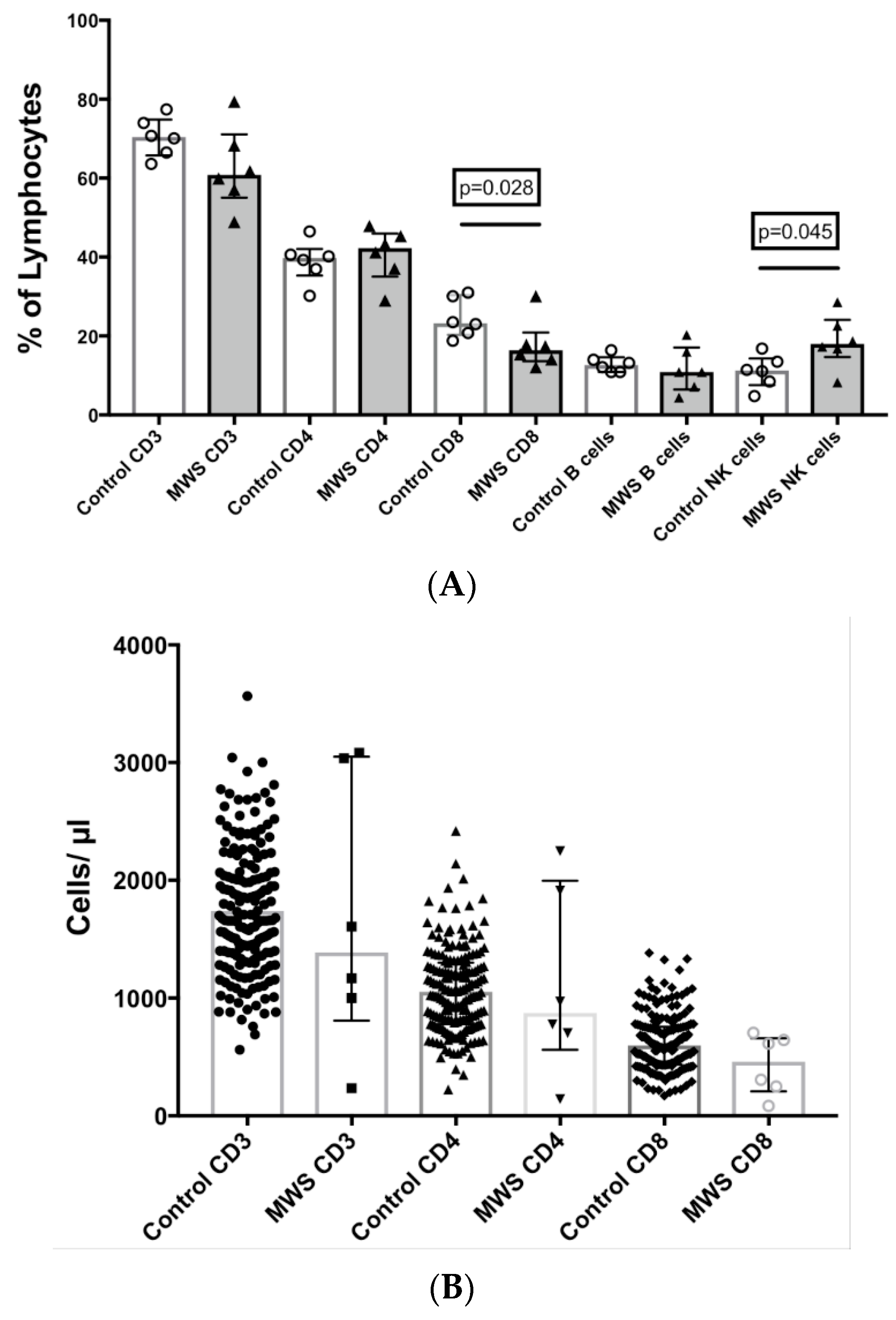
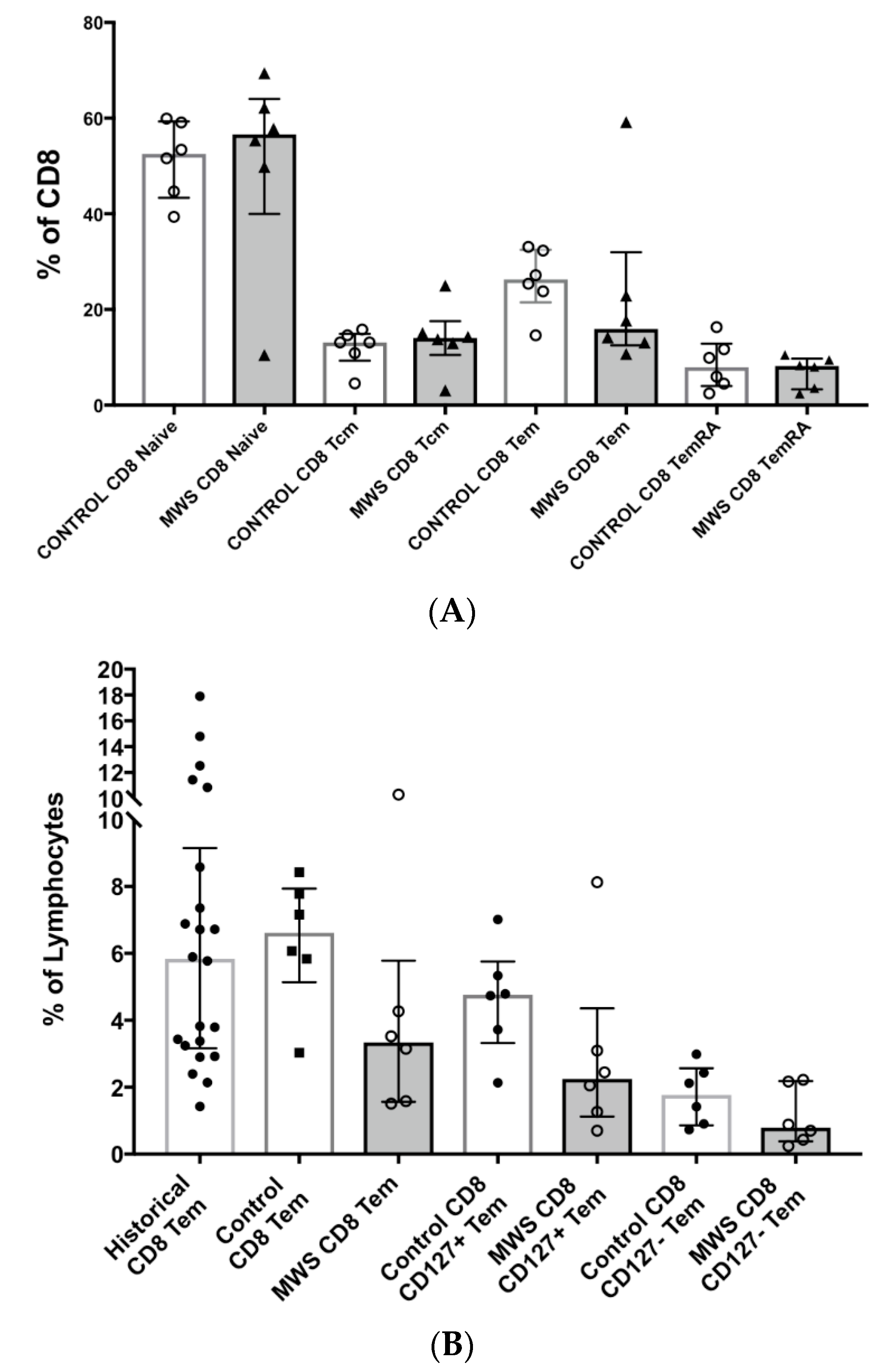
2.2. Lymphocyte Subsets
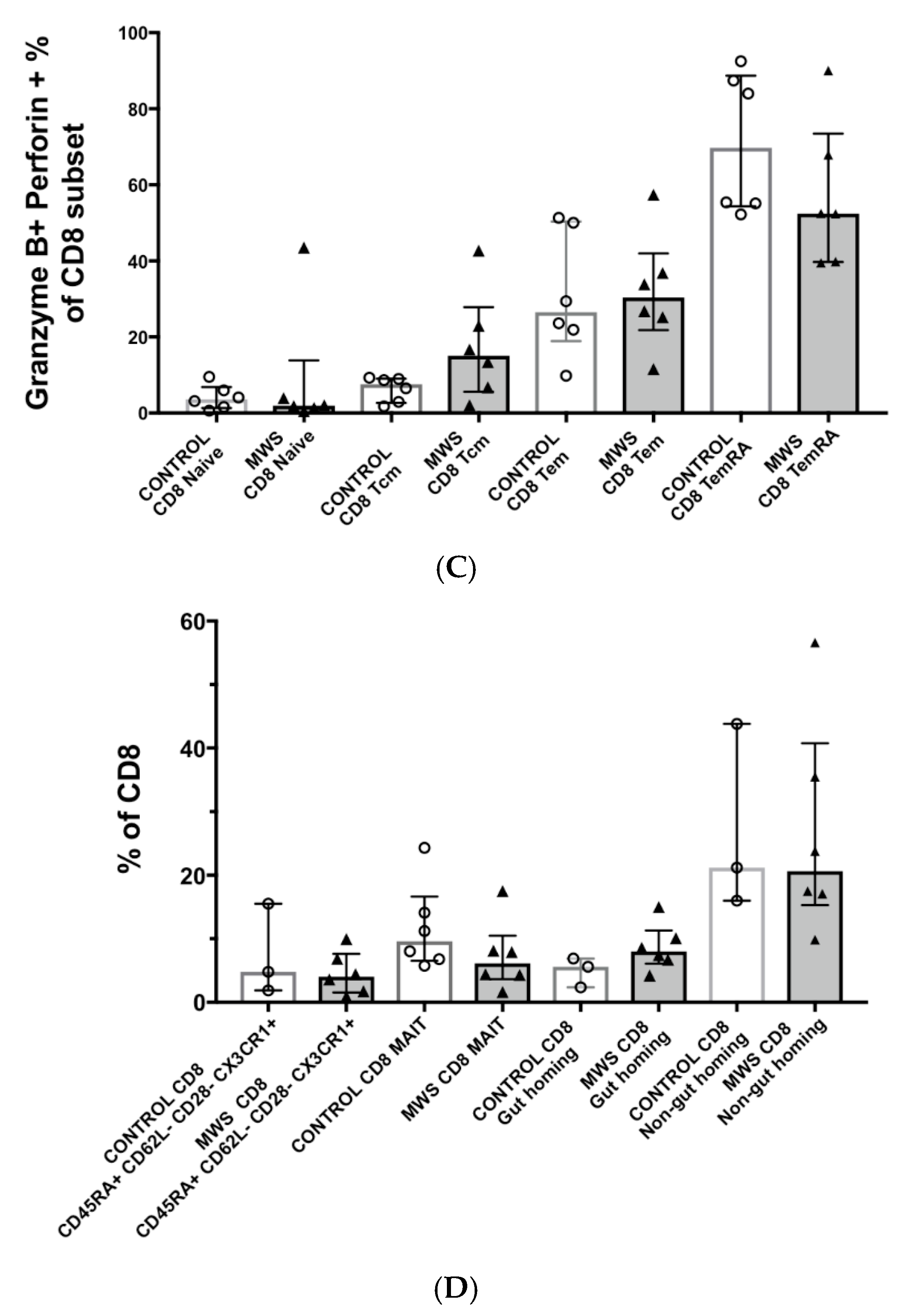
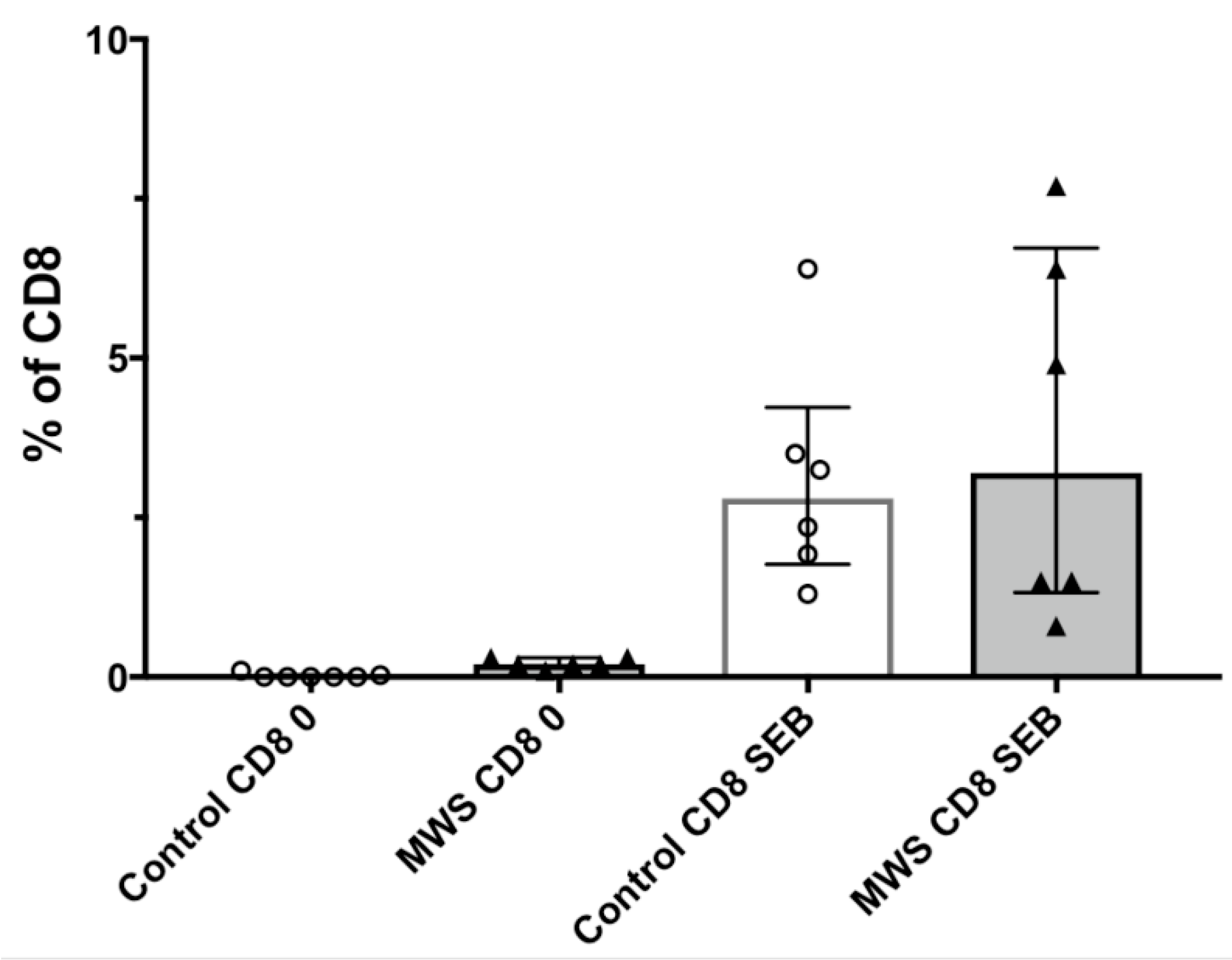
2.3. Lymphocyte Function
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Flow Cytometry
4.3. Lymphocyte Function
4.4. Cell Sorting, RNA Extraction, Preparation of cRNA for Microarray Analysis
4.5. Real-Time PCR
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACD | Acid citrate dextrose |
| ANA | Antinuclear antibody test |
| AOM | Acute otitis media |
| Bcl-2 | B-cell lymphoma 2 |
| CRIM1 | Cysteine rich transmembrane BMP regulator 1 |
| EMT | Epithelial-to-mesenchymal transition |
| HAEC | Hirshsprung associated enterocolitis |
| IGFBP7 | Insulin-like growth factor-binding protein 7 |
| IL- | Interleukin- |
| KLF10 | Kruppel-like factor 10 |
| MAIT | Mucosal associated invariant T cell |
| MWS | Mowat–Wilson syndrome |
| NaHep | Sodium heparin |
| PBMC | Peripheral blood mononuclear cells |
| PBS | Phosphate buffered saline |
| PDGFD | Platelet-derived growth factor |
| RT | Room temperature |
| SMAD5 | SMAD family member 5 |
| TFG-β1 | Transforming growth factor beta 1 |
| TFGBR | Transforming growth factor beta receptor |
| TIEG1 | TFG-β inducible early gene 1 |
| tTG | Transglutaminase |
| VV | Vaccinia virus |
| ZEB2 | Zinc finger E-box binding homeobox 2 gene |
Appendix A
References
- Mowat, D.R.; Croaker, G.D.; Cass, D.T.; Kerr, B.A.; Chaitow, J.; Ades, L.C.; Chia, N.L.; Wilson, M.J. Hirschsprung disease, microcephaly, mental retardation, and characteristic facial features: Delineation of a new syndrome and identification of a locus at chromosome 2q22-q23. J. Med. Genet. 1998, 35, 617–623. [Google Scholar] [CrossRef] [PubMed]
- Ivanovski, I.; Djuric, O.; Caraffi, S.G.; Santodirocco, D.; Pollazzon, M.; Rosato, S.; Cordelli, D.M.; Abdalla, E.; Accorsi, P.; Adam, M.P.; et al. Phenotype and genotype of 87 patients with Mowat–Wilson syndrome and recommendations for care. Genet. Med. 2018, 20, 965–975. [Google Scholar] [CrossRef] [Green Version]
- Ho, S.; Luk, H.-M.; Chung, B.H.-Y.; Fung, J.L.-F.; Mak, H.H.-Y.; Lo, I.F.M. Mowat–Wilson syndrome in a Chinese population: A case series. Am. J. Med. Genet. Part A 2020, 182, 1336–1341. [Google Scholar] [CrossRef] [PubMed]
- Zweier, C.; Thiel, C.T.; Dufke, A.; Crow, Y.J.; Meinecke, P.; Suri, M.; Ala-Mello, S.; Beemer, F.; Bernasconi, S.; Bianchi, P.; et al. Clinical and Mutational Spectrum of Mowat–Wilson Syndrome. Eur. J. Med. Genet. 2005, 48, 97–111. [Google Scholar] [CrossRef] [PubMed]
- Omilusik, K.D.; Best, J.A.; Yu, B.; Goossens, S.; Weidemann, A.; Nguyen, J.V.; Seuntjens, E.; Stryjewska, A.; Zweier, C.; Roychoudhuri, R.; et al. Transcriptional repressor ZEB2 promotes terminal differentiation of CD8+ effector and memory T cell populations during infection. J. Exp. Med. 2015, 212, 2027–2039. [Google Scholar] [CrossRef] [Green Version]
- Arsenio, J.; Kakaradov, B.; Metz, P.J.; Kim, S.H.; Yeo, G.W.; Chang, J.T. Early specification of CD8+ T lymphocyte fates during adaptive immunity revealed by single-cell gene-expression analyses. Nat. Immunol. 2014, 15, 365–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Best, J.A.; Blair, D.A.; Knell, J.; Yang, E.; Mayya, V.; Doedens, A.; Dustin, M.L.; Goldrath, A.W.; The Immunological Genome Project Consortium. Transcriptional insights into the CD8+ T cell response to infection and memory T cell formation. Nat. Immunol. 2013, 14, 404–412. [Google Scholar] [CrossRef]
- Pons, L.; Dupuis-Girod, S.; Cordier, M.-P.; Edery, P.; Rossi, M. ZEB2, a new candidate gene for asplenia. Orphanet J. Rare Dis. 2014, 9, 2. [Google Scholar] [CrossRef] [Green Version]
- Nevarez Flores, C.G.N.; Sun, A.P.; Hast, H. Mowat-Wilson Syndrome Presenting With Purpura Fulminans. Pediatrics 2019, 143, e20180922. [Google Scholar] [CrossRef] [Green Version]
- Sgruletti, M.; Di Cesare, K.; Chini, L.; Graziani, D.S.; Di Matteo, G.; D’Agostini, C.; Falvo, F.; Concolino, D.; Miniero, R.; Moschese, V. Hypogammaglobulinaemia in a patient with Mowat Wilson Syndrome. In Proceedings of the European Society for Immunodeficiencies 17th Biennial Meeting, Barcelona, Spain, 21–24 September 2016. [Google Scholar]
- Zaunders, J.J.; Dyer, W.B.; Munier, M.L.; Ip, S.; Liu, J.; Amyes, E.; Rawlinson, W.; De Rose, R.; Kent, S.J.; Sullivan, J.S.; et al. CD127+CCR5+CD38+++ CD4+ Th1 Effector Cells Are an Early Component of the Primary Immune Response to Vaccinia Virus and Precede Development of Interleukin-2+ Memory CD4+ T Cells. J. Virol. 2006, 80, 10151–10161. [Google Scholar] [CrossRef] [Green Version]
- Munier, C.M.L.; van Bockel, D.; Bailey, M.; Ip, S.; Xu, Y.; Alcantara, S.; Liu, S.M.; Denyer, G.; Kaplan, W.; Suzuki, K.; et al. The primary immune response to Vaccinia virus vaccination includes cells with a distinct cytotoxic effector CD4 T-cell phenotype. Vaccine 2016, 34, 5251–5261. [Google Scholar] [CrossRef] [PubMed]
- Vander Ark, A.; Cao, J.; Li, X. TGF-β receptors: In and beyond TGF-β signaling. Cell Signal 2018, 52, 112–120. [Google Scholar] [CrossRef]
- Dongre, A.; Weinberg, R.A. New insights into the mechanisms of epithelial–mesenchymal transition and implications for cancer. Nat. Rev. Mol. Cell Biol. 2019, 20, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Guan, T.; Dominguez, C.X.; Amezquita, R.A.; Laidlaw, B.J.; Cheng, J.; Henao-Mejia, J.; Williams, A.; Flavell, R.A.; Lu, J.; Kaech, S.M. ZEB1, ZEB2, and the miR-200 family form a counterregulatory network to regulate CD8+ T cell fates. J. Exp. Med. 2018, 215, 1153–1168. [Google Scholar] [CrossRef] [PubMed]
- Memon, A.; Lee, W.K. KLF10 as a Tumor Suppressor Gene and Its TGF-β Signaling. Cancers 2018, 10, 161. [Google Scholar] [CrossRef] [Green Version]
- Trojanowska, M. Role of PDGF in fibrotic diseases and systemic sclerosis. Rheumatology 2008, 47 (Suppl. 5), v2–v4. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, L.; Kolle, G.; Wen, D.; Piper, M.; Scott, J.; Little, M. CRIM1 Regulates the Rate of Processing and Delivery of Bone Morphogenetic Proteins to the Cell Surface. J. Biol. Chem. 2003, 278, 34181–34188. [Google Scholar] [CrossRef] [Green Version]
- Daly, A.C.; Randall, R.A.; Hill, C.S. Transforming growth factor beta-induced Smad1/5 phosphorylation in epithelial cells is mediated by novel receptor complexes and is essential for anchorage-independent growth. Mol. Cell Biol. 2008, 28, 6889–6902. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, J.; Takiyama, Y.; Honjyo, J.; Makino, Y.; Fujita, Y.; Tateno, M.; Haneda, M. Role of IGFBP7 in Diabetic Nephropathy: TGF-beta1 Induces IGFBP7 via Smad2/4 in Human Renal Proximal Tubular Epithelial Cells. PLoS ONE 2016, 11, e0150897. [Google Scholar] [CrossRef] [PubMed]
- Hegarty, S.V.; Sullivan, A.M.; O’Keeffe, G.W. Zeb2: A multifunctional regulator of nervous system development. Prog. Neurobiol. 2015, 132, 81–95. [Google Scholar] [CrossRef]
- Li, H.; Mar, B.G.; Zhang, H.; Puram, R.V.; Vazquez, F.; Weir, B.A.; Hahn, W.C.; Ebert, B.; Pellman, D. The EMT regulator ZEB2 is a novel dependency of human and murine acute myeloid leukemia. Blood 2017, 129, 497–508. [Google Scholar] [CrossRef] [Green Version]
- van Helden, M.J.; Goossens, S.; Daussy, C.; Mathieu, A.L.; Faure, F.; Marçais, A.; Vandamme, N.; Farla, N.; Mayol, K.; Viel, S.; et al. Terminal NK cell maturation is controlled by concerted actions of T-bet and Zeb2 and is essential for melanoma rejection. J. Exp. Med. 2015, 212, 2015–2025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goossens, S.; Janzen, V.; Bartunkova, S.; Yokomizo, T.; Drogat, B.; Crisan, M.; Haigh, K.; Seuntjens, E.; Umans, L.; Riedt, T.; et al. The EMT regulator Zeb2/Sip1 is essential for murine embryonic hematopoietic stem/progenitor cell differentiation and mobilization. Blood 2011, 117, 5620–5630. [Google Scholar] [CrossRef]
- Dominguez, C.X.; Amezquita, R.A.; Guan, T.; Marshall, H.D.; Joshi, N.S.; Kleinstein, S.H.; Kaech, S.M. The transcription factors ZEB2 and T-bet cooperate to program cytotoxic T cell terminal differentiation in response to LCMV viral infection. J. Exp. Med. 2015, 212, 2041–2056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garavelli, L.; Mainardi, P.C. Mowat-Wilson syndrome. Orphanet J. Rare Dis. 2007, 2, 42. [Google Scholar] [CrossRef] [Green Version]
- Gosain, A.; Frykman, P.K.; Cowles, R.A.; Horton, J.; Levitt, M.; Rothstein, D.H.; Langer, J.C.; Goldstein, A.M.; On behalf of the American Pediatric Surgical Association Hirschsprung Disease Interest Group. Guidelines for the diagnosis and management of Hirschsprung-associated enterocolitis. Pediatr. Surg. Int. 2017, 33, 517–521. [Google Scholar] [CrossRef] [Green Version]
- Fischer, A.; Provot, J.; Jais, J.-P.; Alcais, A.; Mahlaoui, N.; Adoue, D.; Aladjidi, N.; Amoura, Z.; Arlet, P.; Armari-Alla, C.; et al. Autoimmune and inflammatory manifestations occur frequently in patients with primary immunodeficiencies. J. Allergy Clin. Immunol. 2017, 140, 1388–1393.e8. [Google Scholar] [CrossRef] [Green Version]
- Conrad, K.; Röber, N.; Andrade, L.E.C.; Mahler, M. The Clinical Relevance of Anti-DFS70 Autoantibodies. Clin. Rev. Allergy Immunol. 2016, 52, 202–216. [Google Scholar] [CrossRef]
- Cook, T.; Gebelein, B.; Mesa, K.; Mladek, A.; Urrutia, R. Molecular cloning and characterization of TIEG2 reveals a new subfamily of transforming growth factor-beta-inducible Sp1-like zinc finger-encoding genes involved in the regulation of cell growth. J. Biol. Chem. 1998, 273, 25929–25936. [Google Scholar] [CrossRef] [Green Version]
- Lewis, G.M.; Wehrens, E.J.; Labarta-Bajo, L.; Streeck, H.; Zuniga, E.I. TGF-beta receptor maintains CD4 T helper cell identity during chronic viral infections. J. Clin. Investig. 2016, 126, 3799–3813. [Google Scholar] [CrossRef]
- Sanjabi, S.; Mosaheb, M.M.; Flavell, R.A. Opposing effects of TGF-beta and IL-15 cytokines control the number of short-lived effector CD8+ T cells. Immunity 2009, 31, 131–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tinoco, R.; Alcalde, V.; Yang, Y.; Sauer, K.; Zuniga, E.I. Cell-intrinsic transforming growth factor-beta signaling mediates virus-specific CD8+ T cell deletion and viral persistence in vivo. Immunity 2009, 31, 145–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouyang, W.; Oh, S.A.; Ma, Q.; Bivona, M.R.; Zhu, J.; Li, M.O. TGF-beta cytokine signaling promotes CD8+ T cell development and low-affinity CD4+ T cell homeostasis by regulation of interleukin-7 receptor alpha expression. Immunity 2013, 39, 335–346. [Google Scholar] [CrossRef] [Green Version]
- Zaunders, J.J.; Lévy, Y.; Seddiki, N. Exploiting differential expression of the IL-7 receptor on memory T cells to modulate immune responses. Cytokine Growth Factor Rev. 2014, 25, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Zaunders, J.J.; Parseval, L.M.-D.; Kitada, S.; Reed, J.C.; Rought, S.; Genini, D.; Leoni, L.; Kelleher, A.; Cooper, D.A.; Smith, D.E.; et al. Polyclonal Proliferation and Apoptosis of CCR5+T Lymphocytes during Primary Human Immunodeficiency Virus Type 1 Infection: Regulation by Interleukin (IL)–2, IL-15, and Bcl-2. J. Infect. Dis. 2003, 187, 1735–1747. [Google Scholar] [CrossRef] [Green Version]
- Zaunders, J.; Jing, J.; Leipold, M.; Maecker, H.; Kelleher, A.D.; Koch, I. Computationally efficient multidimensional analysis of complex flow cytometry data using second order polynomial histograms. Cytom. Part A 2016, 89, 44–58. [Google Scholar] [CrossRef]
- Xu, Y.; Phetsouphanh, C.; Suzuki, K.; Aggrawal, A.; Graff-Dubois, S.; Roche, M.; Bailey, M.; Alcantara, S.; Cashin, K.; Sivasubramaniam, R.; et al. HIV-1 and SIV Predominantly Use CCR5 Expressed on a Precursor Population to Establish Infection in T Follicular Helper Cells. Front. Immunol. 2017, 8, 376. [Google Scholar] [CrossRef] [PubMed]
- Zaunders, J.J.; Munier, M.L.; Seddiki, N.; Pett, S.; Ip, S.; Bailey, M.; Xu, Y.; Brown, K.; Dyer, W.B.; Kim, M.; et al. High Levels of Human Antigen-Specific CD4+ T Cells in Peripheral Blood Revealed by Stimulated Coexpression of CD25 and CD134 (OX40). J. Immunol. 2009, 183, 2827–2836. [Google Scholar] [CrossRef] [Green Version]
- Baugh, L.R. Quantitative analysis of mRNA amplification by in vitro transcription. Nucleic Acids Res. 2001, 29, E29. [Google Scholar] [CrossRef]
- Chtanova, T.; Tangye, S.G.; Newton, R.; Frank, N.; Hodge, M.R.; Rolph, M.S.; Mackay, C.R. T Follicular Helper Cells Express a Distinctive Transcriptional Profile, Reflecting Their Role as Non-Th1/Th2 Effector Cells That Provide Help for B Cells. J. Immunol. 2004, 173, 68–78. [Google Scholar] [CrossRef]
- Liu, S.M.; Xavier, R.; Good, K.L.; Chtanova, T.; Newton, R.; Sisavanh, M.; Zimmer, S.; Deng, C.; Silva, D.G.; Frost, M.J. Immune cell transcriptome datasets reveal novel leukocyte subset–specific genes and genes associated with allergic processes. J. Allergy Clin. Immunol. 2006, 118, 496–503. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age (y)/Gender (M/F) | ZEB2 Mutation | Infections | Spleen | Immune Function Tests | ||
|---|---|---|---|---|---|---|
| Hypo/asplenia | Humoral | Cellular | ||||
| Nevarez Flores 2019 [9] | 0.8 F | c.1426dup | S. pneumoniae meningitis and purpura fulminans | Asplenia | NR | NR |
| Sgruletti 2016 [10] | 37 F | 4.6 Mb microdeletion 22q (22q22.3–22q23.2) | Nil significant | No | -Severe pan-hypogammaglobulinaemia; -Absent functional IgG response to tetanus | -Increased B cell numbers; -Dramatically reduced switched and non-switched memory B cells; -Reduced CD4 memory cells; -Increased terminally differentiated CD8 cells |
| Pons 2014 [8] | 0.7 F | c.2083C > T | S. pneumoniae purpura fulminans | Asplenia | NR | NR |
| 1 F | c.600_640dup | S. pneumoniae meningitis | Asplenia | NR | NR | |
| 2 F | c.1762G > T | S. pneumoniae sepsis ×2 | Hyposplenia | NR | NR | |
| F | c.1426dup | Nil reported | Hypoplasia | NR | NR | |
| Zweier 2005 [4]/ Ivanovski 2018 [2] | 1 F | c.696C > G | Ni reported | Asplenia | NR | NR |
| Age (y) Gender (M/F) | ZEB2 Mutation | Infections | Splenic Hypo/ Asplenia | Immunoglobulins (g/L) | Lymphocyte Subsets (×109/L) | Vaccine Responses * | Other Clinical History | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sinopulmonary | Severe viral | Opportunistic | Yes/No | IgG | IgA | IgM | CD4 | CD8 | CD19 | CD16+CD56 | Protein | Polysaccharide (pre & post Pneumovax 23) | |||||
| 1 | 22 M | NM_014795.4:c.1426dupA | AOMp+, pneumonia+, sepsis+ | No | Candida (Inv, Sup+) | N | 11.5 | 3.5 | 1.7 | 0.2 ↓ | 0.1 ↓ | 0.05 ↓ | 0.2 | BR | Poor | AR | ID, HD with colitis; thrombocytopenia; epilepsy; tetralogy of Fallot; submucosal cleft palate; recurrent encephalopathy with autonomic dysfunction; osteopenia and # |
| 2 | 21 M | partial deletion of chromosome 2q22 | AOMp+, sinusitis+, pneumonia+, sepsis | No | Candida (Sup+) | N | 11.5 | 3.6 | 0.2 ↓ | 1.2 | 0.8 ↑ | 0.1 ↓ | 0.2 | BR | Poor | ND | ID, HD with colitis; thrombocytopenia; epilepsy; scoliosis; #; iron deficiency anaemia; neurogenic bladder; atopy |
| 3 | 20 M | NM_014795.4: c.2798delC: p.Pro933HisfsTer44 | AOM | No | No | N | 11.5 | 2.2 | 0.6 | 1.1 | 0.4 | 0.3 | 0.5 ↑ | AR | BR | ND | ID; epilepsy |
| 4 | 12 F | c.690–721del | AOM | No | No | N | 7.9 | 0.8 | 1.1 | 0.9 | 0.4 | 0.3 | 0.4 | poor | poor | ND | ID; epilepsy; constipation |
| 5 | 11 M | NM_014795.4: c.3211T > C: p.Ser1071Pro | pneumonia, tonsillitis+ | No | No | N | 9.7 | 1.6 | 1.6 | 2.7 | 1.9 | 0.95 ↑ | 0.8 | AR | poor | ND | DD; epilepsy |
| 6 | 3 M | NM_014795.4: c.1106T > G: p.Leu369Ter | AOM | No | No | N | 7.4 | 0.3 | 0.6 | 2.6 | 1.6 | 0.8 | 0.8 | AR | poor | AR | ID;HD; L esotropia |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frith, K.; Munier, C.M.L.; Hastings, L.; Mowat, D.; Wilson, M.; Seddiki, N.; Macintosh, R.; Kelleher, A.D.; Gray, P.; Zaunders, J.J. The Role of ZEB2 in Human CD8 T Lymphocytes: Clinical and Cellular Immune Profiling in Mowat–Wilson Syndrome. Int. J. Mol. Sci. 2021, 22, 5324. https://doi.org/10.3390/ijms22105324
Frith K, Munier CML, Hastings L, Mowat D, Wilson M, Seddiki N, Macintosh R, Kelleher AD, Gray P, Zaunders JJ. The Role of ZEB2 in Human CD8 T Lymphocytes: Clinical and Cellular Immune Profiling in Mowat–Wilson Syndrome. International Journal of Molecular Sciences. 2021; 22(10):5324. https://doi.org/10.3390/ijms22105324
Chicago/Turabian StyleFrith, Katie, C. Mee Ling Munier, Lucy Hastings, David Mowat, Meredith Wilson, Nabila Seddiki, Rebecca Macintosh, Anthony D. Kelleher, Paul Gray, and John James Zaunders. 2021. "The Role of ZEB2 in Human CD8 T Lymphocytes: Clinical and Cellular Immune Profiling in Mowat–Wilson Syndrome" International Journal of Molecular Sciences 22, no. 10: 5324. https://doi.org/10.3390/ijms22105324
APA StyleFrith, K., Munier, C. M. L., Hastings, L., Mowat, D., Wilson, M., Seddiki, N., Macintosh, R., Kelleher, A. D., Gray, P., & Zaunders, J. J. (2021). The Role of ZEB2 in Human CD8 T Lymphocytes: Clinical and Cellular Immune Profiling in Mowat–Wilson Syndrome. International Journal of Molecular Sciences, 22(10), 5324. https://doi.org/10.3390/ijms22105324