
MMP-9 Deletion Attenuates Arteriovenous Fistula Neointima through Reduced Perioperative Vascular Inflammation


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Neointima Increases Progressively in the AV Fistula Venous Segment of Wild-Type (WT) Mice
2.2. Vascular Inflammation Was Detected in the Early Perioperative Phase of AV Fistula
2.3. MMP-9 Expression Elevates Significantly during the Perioperative Period after AV Fistula Creation
2.4. MMP-9 Knockout Does Not Have Any Detrimental Influence on AVF Venous Segment OR, but Attenuates Neointimal Hyperplasia and Increases Lumen Area
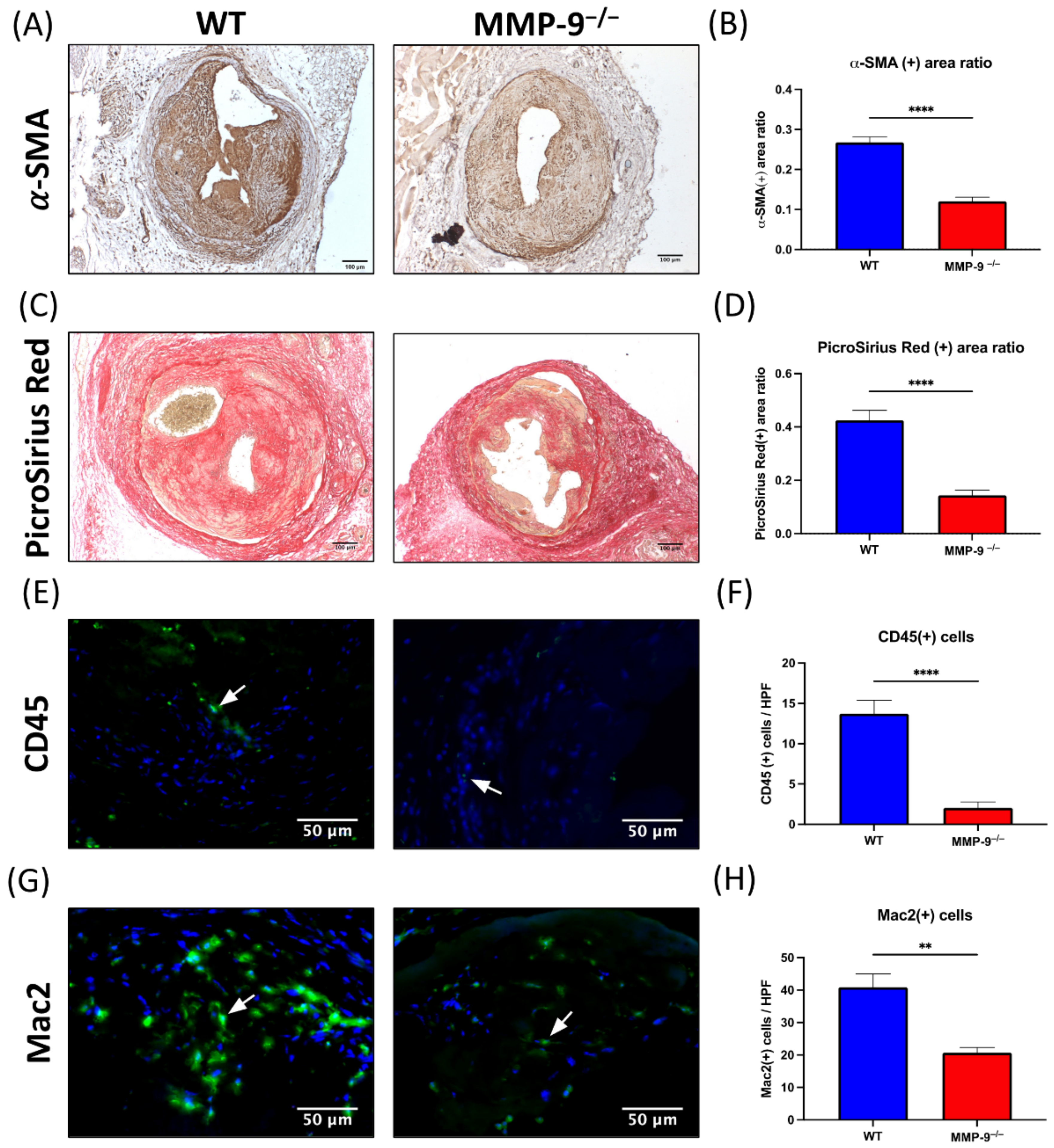
2.5. MMP-9 Knockout Significantly Diminished α-Smooth Muscle Actin (α-SMA) and Collagen Content in Neointima
2.6. MMP-9 Knockout Reduced CD45(+) and Mac2(+) Cells within Neointima
2.7. MMP-9 Knockout Downregulates Perioperative Inflammation in AV Fistula
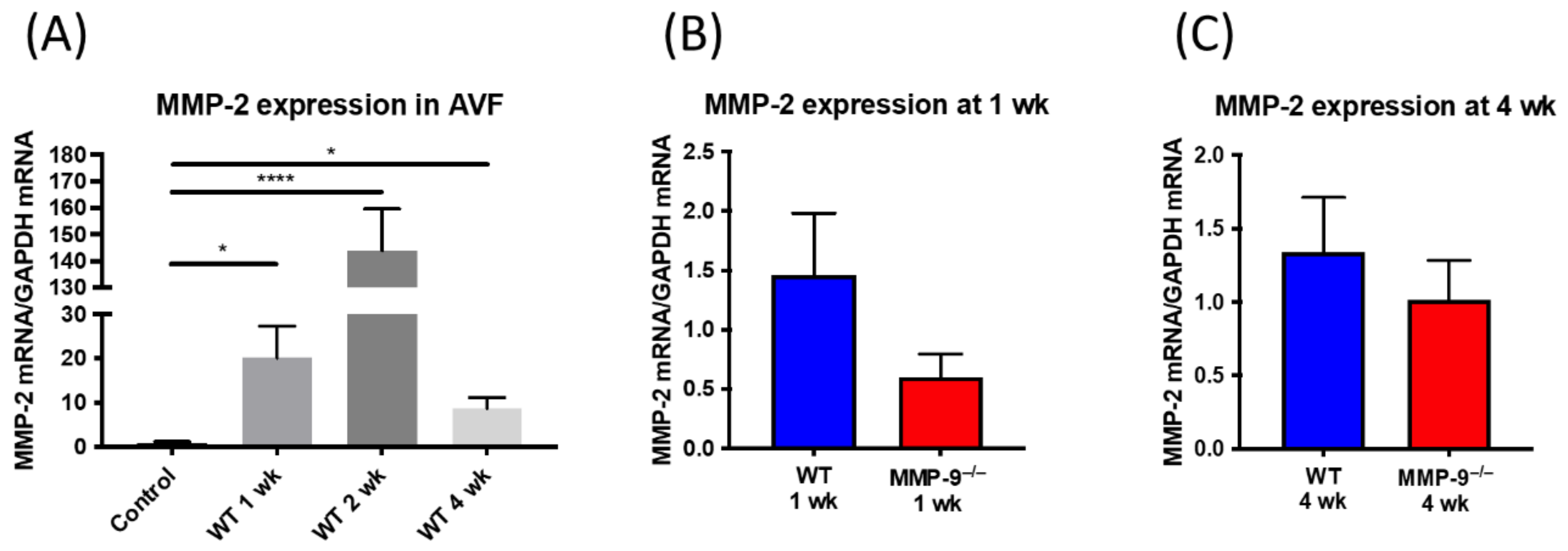
2.8. MMP-2 Expression in AV Fistula Venous Segment Was Not Significantly Altered in MMP-9 Knockout Mice
2.9. MMP-9 Knockout Downregulated CD44 and RAC-Alpha Serine/Threonine-Protein Kinase (Akt) and Extracellular Signal-Regulated Kinase (ERK) Phosphorylation in AV Fistula Venous Segment
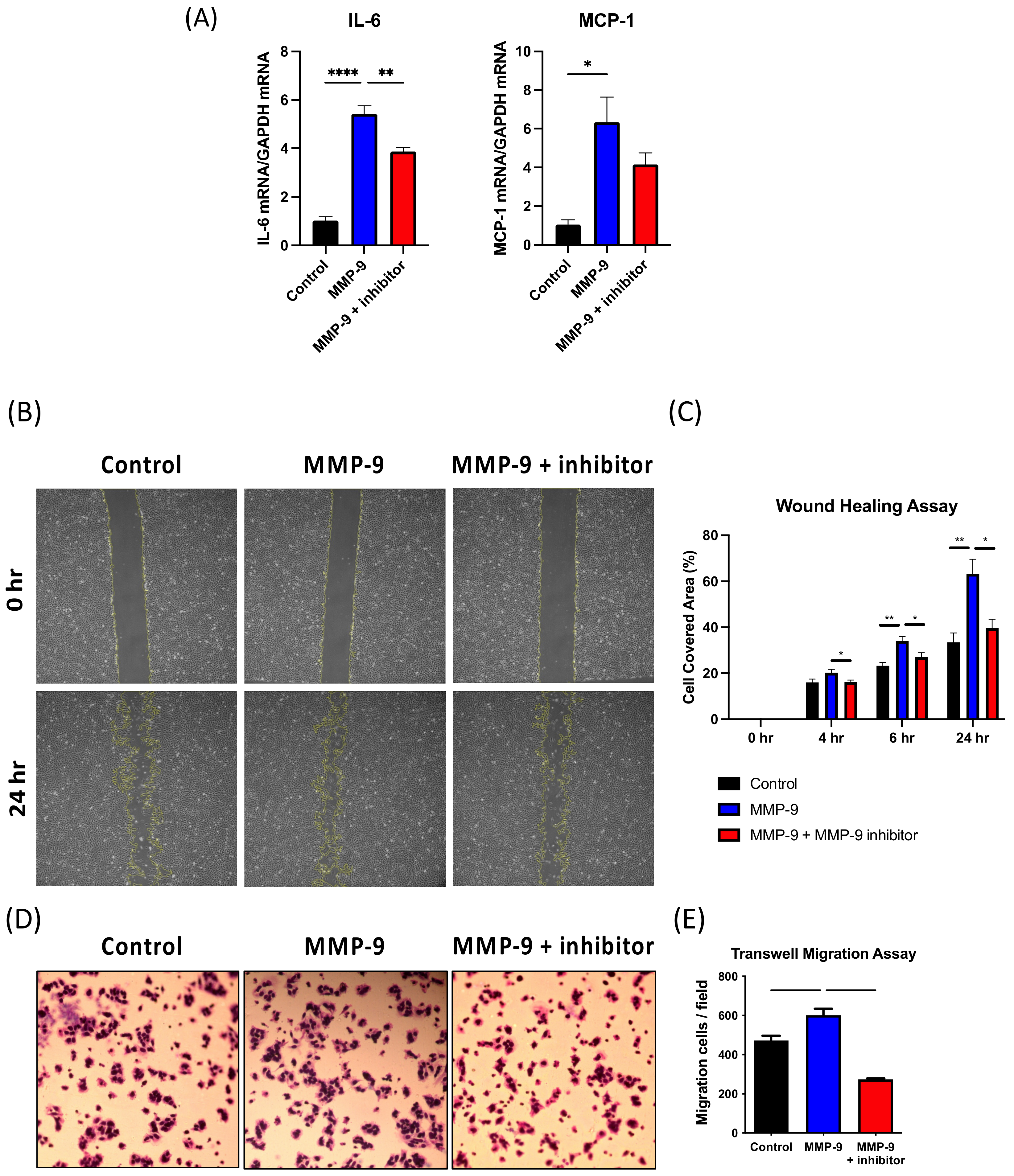
2.10. MMP-9 Increases MCP-1 and IL-6 Expression and Migration in VSMCs
3. Discussion
4. Materials and Methods
4.1. Animal Study
4.2. Tissue Harvesting and Processing
4.3. Measurement of Serum BUN and Creatinine
4.4. Morphometric Analysis
4.5. Collagen Quantification
4.6. Immunofluorescence and Immunohistochemical Staining
4.7. RNA Extraction and Quantitative PCR Analysis
4.8. RNA Sequencing, Differential Gene Expression and GSEA
4.9. Western Blotting
4.10. Cell Culture
4.11. Cell Migration Assay
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sands, J.J. Increasing AV fistulas: Revisiting a time-tested solution. Semin. Dial. 2000, 13, 351–353. [Google Scholar] [CrossRef]
- Roy-Chaudhury, P.; Lee, T.C. Vascular stenosis: Biology and interventions. Curr. Opin. Nephrol. Hypertens. 2007, 16, 516–522. [Google Scholar] [CrossRef] [PubMed]
- Rooijens, P.P.; Tordoir, J.H.; Stijnen, T.; Burgmans, J.P.; Smet de, A.A.; Yo, T.I. Radiocephalic wrist arteriovenous fistula for hemodialysis: Meta-analysis indicates a high primary failure rate. Eur. J. Vasc. Endovasc. Surg. 2004, 28, 583–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huijbregts, H.J.; Bots, M.L.; Wittens, C.H.; Schrama, Y.C.; Moll, F.L.; Blankestijn, P.J.; CIMINO Study Group. Hemodialysis arteriovenous fistula patency revisited: Results of a prospective, multicenter initiative. Clin. J. Am. Soc. Nephrol. 2008, 3, 714–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Jaishi, A.A.; Oliver, M.J.; Thomas, S.M.; Lok, C.E.; Zhang, J.C.; Garg, A.X.; Kosa, S.D.; Quinn, R.R.; Moist, L.M. Patency rates of the arteriovenous fistula for hemodialysis: A systematic review and meta-analysis. Am. J. Kidney Dis. 2014, 63, 464–478. [Google Scholar] [CrossRef] [PubMed]
- Rothuizen, T.C.; Wong, C.; Quax, P.H.; van Zonneveld, A.J.; Rabelink, T.J.; Rotmans, J.I. Arteriovenous access failure: More than just intimal hyperplasia? Nephrol. Dial. Transplant. 2013, 28, 1085–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brahmbhatt, A.; Remuzzi, A.; Franzoni, M.; Misra, S. The molecular mechanisms of hemodialysis vascular access failure. Kidney Int. 2016, 89, 303–316. [Google Scholar] [CrossRef] [Green Version]
- Wasse, H.; Huang, R.; Naqvi, N.; Smith, E.; Wang, D.; Husain, A. Inflammation, oxidation and venous neointimal hyperplasia precede vascular injury from AVF creation in CKD patients. J. Vasc. Access. 2012, 13, 168–174. [Google Scholar] [CrossRef]
- Roy-Chaudhury, P.; Wang, Y.; Krishnamoorthy, M.; Zhang, J.; Banerjee, R.; Munda, R.; Heffelfinger, S.; Arend, L. Cellular phenotypes in human stenotic lesions from haemodialysis vascular access. Nephrol. Dial. Transplant. 2009, 24, 2786–2791. [Google Scholar] [CrossRef] [Green Version]
- Roy-Chaudhury, P.; Arend, L.; Zhang, J.; Krishnamoorthy, M.; Wang, Y.; Banerjee, R.; Samaha, A.; Munda, R. Neointimal hyperplasia in early arteriovenous fistula failure. Am. J. Kidney Dis. 2007, 50, 782–790. [Google Scholar] [CrossRef] [Green Version]
- Stracke, S.; Konner, K.; Kostlin, I.; Friedl, R.; Jehle, P.M.; Hombach, V.; Keller, F.; Waltenberger, J. Increased expression of TGF-beta1 and IGF-I in inflammatory stenotic lesions of hemodialysis fistulas. Kidney Int. 2002, 61, 1011–1019. [Google Scholar] [CrossRef] [Green Version]
- Simone, S.; Loverre, A.; Cariello, M.; Divella, C.; Castellano, G.; Gesualdo, L.; Pertosa, G.; Grandaliano, G. Arteriovenous fistula stenosis in hemodialysis patients is characterized by an increased adventitial fibrosis. J. Nephrol. 2014, 27, 555–562. [Google Scholar] [CrossRef]
- Zempo, N.; Kenagy, R.D.; Au, Y.P.; Bendeck, M.; Clowes, M.M.; Reidy, M.A.; Clowes, A.W. Matrix metalloproteinases of vascular wall cells are increased in balloon-injured rat carotid artery. J. Vasc. Surg. 1994, 20, 209–217. [Google Scholar] [CrossRef] [Green Version]
- Abbruzzese, T.A.; Guzman, R.J.; Martin, R.L.; Yee, C.; Zarins, C.K.; Dalman, R.L. Matrix metalloproteinase inhibition limits arterial enlargements in a rodent arteriovenous fistula model. Surgery 1998, 124, 328–334. [Google Scholar] [CrossRef]
- Tronc, F.; Mallat, Z.; Lehoux, S.; Wassef, M.; Esposito, B.; Tedgui, A. Role of matrix metalloproteinases in blood flow-induced arterial enlargement: Interaction with NO. Arterioscler. Thromb. Vasc. Biol. 2000, 20, E120–E126. [Google Scholar] [CrossRef]
- Visse, R.; Nagase, H. Matrix metalloproteinases and tissue inhibitors of metalloproteinases: Structure, function, and biochemistry. Circ. Res. 2003, 92, 827–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sternlicht, M.D.; Werb, Z. How matrix metalloproteinases regulate cell behavior. Annu. Rev. Cell Dev. Biol. 2001, 17, 463–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yabluchanskiy, A.; Ma, Y.; Iyer, R.P.; Hall, M.E.; Lindsey, M.L. Matrix metalloproteinase-9: Many shades of function in cardiovascular disease. Physiology 2013, 28, 391–403. [Google Scholar] [CrossRef] [Green Version]
- Mott, J.D.; Werb, Z. Regulation of matrix biology by matrix metalloproteinases. Curr. Opin. Cell Biol. 2004, 16, 558–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newby, A.C. Dual role of matrix metalloproteinases (matrixins) in intimal thickening and atherosclerotic plaque rupture. Physiol. Rev. 2005, 85, 1–31. [Google Scholar] [CrossRef] [Green Version]
- Chan, C.Y.; Chen, Y.S.; Ma, M.C.; Chen, C.F. Remodeling of experimental arteriovenous fistula with increased matrix metalloproteinase expression in rats. J. Vasc. Surg. 2007, 45, 804–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.J.; Ko, Y.S.; Ko, P.J.; Hsu, L.A.; Chen, C.F.; Yang, C.W.; Hsu, T.S.; Pang, J.H. Thrombosed arteriovenous fistula for hemodialysis access is characterized by a marked inflammatory activity. Kidney Int. 2005, 68, 1312–1319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croatt, A.J.; Grande, J.P.; Hernandez, M.C.; Ackerman, A.W.; Katusic, Z.S.; Nath, K.A. Characterization of a model of an arteriovenous fistula in the rat: The effect of L-NAME. Am. J. Pathol. 2010, 176, 2530–2541. [Google Scholar] [CrossRef] [PubMed]
- Juncos, J.P.; Tracz, M.J.; Croatt, A.J.; Grande, J.P.; Ackerman, A.W.; Katusic, Z.S.; Nath, K.A. Genetic deficiency of heme oxygenase-1 impairs functionality and form of an arteriovenous fistula in the mouse. Kidney Int. 2008, 74, 47–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misra, S.; Fu, A.A.; Anderson, J.L.; Sethi, S.; Glockner, J.F.; McKusick, M.A.; Bjarnason, H.; Woodrum, D.A.; Mukhopadhyay, D. The rat femoral arteriovenous fistula model: Increased expression of matrix metalloproteinase-2 and -9 at the venous stenosis. J. Vasc. Interv. Radiol. 2008, 19, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Misra, S.; Fu, A.A.; Rajan, D.K.; Juncos, L.A.; McKusick, M.A.; Bjarnason, H.; Mukhopadhyay, D. Expression of hypoxia inducible factor-1 alpha, macrophage migration inhibition factor, matrix metalloproteinase-2 and -9, and their inhibitors in hemodialysis grafts and arteriovenous fistulas. J. Vasc. Interv. Radiol. 2008, 19, 252–259. [Google Scholar] [CrossRef]
- Lin, C.C.; Yang, W.C.; Chung, M.Y.; Lee, P.C. Functional polymorphisms in matrix metalloproteinases-1, -3, -9 are associated with arteriovenous fistula patency in hemodialysis patients. Clin. J. Am. Soc. Nephrol. 2010, 5, 1805–1814. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Ning, W.; Tan, Z.; Gong, Z.; Li, X. Mechanism of matrix metalloproteinase axis-induced neointimal growth. J. Mol. Cell Cardiol. 2014, 66, 116–125. [Google Scholar] [CrossRef]
- Janardhanan, R.; Yang, B.; Vohra, P.; Roy, B.; Withers, S.; Bhattacharya, S.; Mandrekar, J.; Kong, H.; Leof, E.B.; Mukhopadhyay, D.; et al. Simvastatin reduces venous stenosis formation in a murine hemodialysis vascular access model. Kidney Int. 2013, 84, 338–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nath, K.A.; Grande, J.P.; Kang, L.; Juncos, J.P.; Ackerman, A.W.; Croatt, A.J.; Katusic, Z.S. ss-Catenin is markedly induced in a murine model of an arteriovenous fistula: The effect of metalloproteinase inhibition. Am. J. Physiol. Renal. Physiol. 2010, 299, F1270–F1277. [Google Scholar] [CrossRef]
- Shih, Y.C.; Wu, C.C.; Wang, S.C.; Liou, J.Y.; Huang, P.H.; Tarng, D.C. Oral Charcoal Adsorbents Attenuate Neointima Formation of Arteriovenous Fistulas. Toxins 2020, 12, 237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vu, T.H.; Shipley, J.M.; Bergers, G.; Berger, J.E.; Helms, J.A.; Hanahan, D.; Shapiro, S.D.; Senior, R.M.; Werb, Z. MMP-9/gelatinase B is a key regulator of growth plate angiogenesis and apoptosis of hypertrophic chondrocytes. Cell 1998, 93, 411–422. [Google Scholar] [CrossRef] [Green Version]
- Wong, C.Y.; de Vries, M.R.; Wang, Y.; van der Vorst, J.R.; Vahrmeijer, A.L.; van Zonneveld, A.J.; Roy-Chaudhury, P.; Rabelink, T.J.; Quax, P.H.; Rotmans, J.I. Vascular remodeling and intimal hyperplasia in a novel murine model of arteriovenous fistula failure. J. Vasc. Surg. 2014, 59, 192–201.e191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Liang, A.; Luo, J.; Liang, M.; Han, G.; Mitch, W.E.; Cheng, J. Blocking Notch in endothelial cells prevents arteriovenous fistula failure despite CKD. J. Am. Soc. Nephrol. 2014, 25, 773–783. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Mootha, V.K.; Lindgren, C.M.; Eriksson, K.F.; Subramanian, A.; Sihag, S.; Lehar, J.; Puigserver, P.; Carlsson, E.; Ridderstrale, M.; Laurila, E.; et al. PGC-1alpha-responsive genes involved in oxidative phosphorylation are coordinately downregulated in human diabetes. Nat. Genet. 2003, 34, 267–273. [Google Scholar] [CrossRef]
- Liberzon, A.; Birger, C.; Thorvaldsdottir, H.; Ghandi, M.; Mesirov, J.P.; Tamayo, P. The Molecular Signatures Database (MSigDB) hallmark gene set collection. Cell Syst. 2015, 1, 417–425. [Google Scholar] [CrossRef] [Green Version]
- Kuwahara, G.; Hashimoto, T.; Tsuneki, M.; Yamamoto, K.; Assi, R.; Foster, T.R.; Hanisch, J.J.; Bai, H.; Hu, H.; Protack, C.D.; et al. CD44 Promotes Inflammation and Extracellular Matrix Production During Arteriovenous Fistula Maturation. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 1147–1156. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Fereydooni, A.; Isaji, T.; Gorecka, J.; Liu, S.; Hu, H.; Ono, S.; Alozie, M.; Lee, S.R.; Taniguchi, R.; et al. Inhibition of the Akt1-mTORC1 Axis Alters Venous Remodeling to Improve Arteriovenous Fistula Patency. Sci. Rep. 2019, 9, 11046. [Google Scholar] [CrossRef]
- Ene-Iordache, B.; Remuzzi, A. Disturbed flow in radial-cephalic arteriovenous fistulae for haemodialysis: Low and oscillating shear stress locates the sites of stenosis. Nephrol. Dial. Transplant. 2012, 27, 358–368. [Google Scholar] [CrossRef]
- Chen, N.X.; O’Neill, K.D.; Chen, X.; Kiattisunthorn, K.; Gattone, V.H.; Moe, S.M. Activation of arterial matrix metalloproteinases leads to vascular calcification in chronic kidney disease. Am. J. Nephrol. 2011, 34, 211–219. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Vohra, P.K.; Janardhanan, R.; Misra, K.D.; Misra, S. Expression of profibrotic genes in a murine remnant kidney model. J. Vasc. Interv. Radiol. 2011, 22, 1765–1772.e1761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Ding, W.; Wei, Q.; Shen, Z.; Wang, C. Dysregulated gene expression of extracellular matrix and adhesion molecules in saphenous vein conduits of hemodialysis patients. J. Thorac. Cardiovasc. Surg. 2012, 144, 684–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henaut, L.; Mary, A.; Chillon, J.M.; Kamel, S.; Massy, Z.A. The Impact of Uremic Toxins on Vascular Smooth Muscle Cell Function. Toxins 2018, 10, 218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, A.; Wang, Y.; Han, G.; Truong, L.; Cheng, J. Chronic kidney disease accelerates endothelial barrier dysfunction in a mouse model of an arteriovenous fistula. Am. J. Physiol. Renal. Physiol. 2013, 304, F1413–F1420. [Google Scholar] [CrossRef] [Green Version]
- Khandoga, A.; Kessler, J.S.; Hanschen, M.; Khandoga, A.G.; Burggraf, D.; Reichel, C.; Hamann, G.F.; Enders, G.; Krombach, F. Matrix metalloproteinase-9 promotes neutrophil and T cell recruitment and migration in the postischemic liver. J. Leukoc. Biol. 2006, 79, 1295–1305. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Hart, E.; Shchurin, A.; Hoover-Plow, J. Inflammatory macrophage migration requires MMP-9 activation by plasminogen in mice. J. Clin. Investig. 2008, 118, 3012–3024. [Google Scholar] [CrossRef] [Green Version]
- Juncos, J.P.; Grande, J.P.; Kang, L.; Ackerman, A.W.; Croatt, A.J.; Katusic, Z.S.; Nath, K.A. MCP-1 contributes to arteriovenous fistula failure. J. Am. Soc. Nephrol. 2011, 22, 43–48. [Google Scholar] [CrossRef] [Green Version]
- Moon, S.K.; Cha, B.Y.; Kim, C.H. ERK1/2 mediates TNF-alpha-induced matrix metalloproteinase-9 expression in human vascular smooth muscle cells via the regulation of NF-kappaB and AP-1: Involvement of the ras dependent pathway. J. Cell. Physiol. 2004, 198, 417–427. [Google Scholar] [CrossRef]
- Sakata, K.; Hamaoka, K.; Ozawa, S.; Niboshi, A.; Yahata, T.; Fujii, M.; Hamaoka, A.; Toiyama, K.; Nishida, M.; Itoi, T. Matrix metalloproteinase-9 in vascular lesions and endothelial regulation in Kawasaki disease. Circ. J. 2010, 74, 1670–1675. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.Q.; Li, W.; Li, S.Q.; Li, J.; Li, Y.W.; Kong, S.X.; Liu, R.M.; Wang, S.M.; Lv, W.M. MCP-1 stimulates MMP-9 expression via ERK 1/2 and p38 MAPK signaling pathways in human aortic smooth muscle cells. Cell Physiol. Biochem. 2014, 34, 266–276. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.; Kuppler, C.; He, Y.; Tao, M.; Ding, K.; Longchamp, A.; Dember, L.M.; Ozaki, C.K.; Berceli, S.A. Local Adipose-Associated Mediators and Adaptations Following Arteriovenous Fistula Creation. Kidney Int. Rep. 2018, 3, 970–978. [Google Scholar] [CrossRef] [Green Version]
- Deleon-Pennell, K.Y.; Altara, R.; Yabluchanskiy, A.; Modesti, A.; Lindsey, M.L. The circular relationship between matrix metalloproteinase-9 and inflammation following myocardial infarction. IUBMB Life 2015, 67, 611–618. [Google Scholar] [CrossRef] [Green Version]
- Malik, M.; Bakshi, C.S.; McCabe, K.; Catlett, S.V.; Shah, A.; Singh, R.; Jackson, P.L.; Gaggar, A.; Metzger, D.W.; Melendez, J.A.; et al. Matrix metalloproteinase 9 activity enhances host susceptibility to pulmonary infection with type A and B strains of Francisella tularensis. J. Immunol. 2007, 178, 1013–1020. [Google Scholar] [CrossRef] [Green Version]
- Gambillara, V.; Montorzi, G.; Haziza-Pigeon, C.; Stergiopulos, N.; Silacci, P. Arterial wall response to ex vivo exposure to oscillatory shear stress. J. Vasc. Res. 2005, 42, 535–544. [Google Scholar] [CrossRef]
- Cho, A.; Reidy, M.A. Matrix metalloproteinase-9 is necessary for the regulation of smooth muscle cell replication and migration after arterial injury. Circ. Res. 2002, 91, 845–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berceli, S.A.; Jiang, Z.; Klingman, N.V.; Pfahnl, C.L.; Abouhamze, Z.S.; Frase, C.D.; Schultz, G.S.; Ozaki, C.K. Differential expression and activity of matrix metalloproteinases during flow-modulated vein graft remodeling. J. Vasc. Surg. 2004, 39, 1084–1090. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.J.; Chen, C.C.; Hsu, L.A.; Chang, G.J.; Ko, Y.H.; Chen, C.F.; Chen, M.Y.; Yang, S.H.; Pang, J.H. Degradation of the internal elastic laminae in vein grafts of rats with aortocaval fistulae: Potential impact on graft vasculopathy. Am. J. Pathol. 2009, 174, 1837–1846. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Zuckerman, S.T.; Cai, C.; Kilari, S.; Singh, A.; Simeon, M.; von Recum, H.A.; Korley, J.N.; Misra, S. Periadventitial Delivery of Simvastatin-Loaded Microparticles Attenuate Venous Neointimal Hyperplasia Associated with Arteriovenous Fistula. J. Am. Heart Assoc. 2020, 9, e018418. [Google Scholar] [CrossRef] [PubMed]
- Bonetti, P.O.; Lerman, L.O.; Napoli, C.; Lerman, A. Statin effects beyond lipid lowering--are they clinically relevant? Eur. Heart J. 2003, 24, 225–248. [Google Scholar] [CrossRef] [Green Version]
- Johnson, P.; Ruffell, B. CD44 and its role in inflammation and inflammatory diseases. Inflamm. Allergy Drug Targets 2009, 8, 208–220. [Google Scholar] [CrossRef]
- Pan, L.; Ni, H.; Jin, W.; Su, X. Inhibition of ERK or Akt ameliorates intimal hyperplasia via up-regulation of Cx37 and down-regulation of Cx43 in balloon injury rat model. Cardiovasc. Diagn. Ther. 2020, 10, 658–666. [Google Scholar] [CrossRef] [PubMed]
- Sedding, D.G.; Trobs, M.; Reich, F.; Walker, G.; Fink, L.; Haberbosch, W.; Rau, W.; Tillmanns, H.; Preissner, K.T.; Bohle, R.M.; et al. 3-Deazaadenosine prevents smooth muscle cell proliferation and neointima formation by interfering with Ras signaling. Circ. Res. 2009, 104, 1192–1200. [Google Scholar] [CrossRef] [Green Version]
- Wan, J.; Lata, C.; Santilli, A.; Green, D.; Roy, S.; Santilli, S. Supplemental oxygen reverses hypoxia-induced smooth muscle cell proliferation by modulating HIF-alpha and VEGF levels in a rabbit arteriovenous fistula model. Ann. Vasc. Surg. 2014, 28, 725–736. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Ren, X.; Zhao, M.; Zhou, B.; Han, Y. Angiotensin-(1-7) abrogates angiotensin II-induced proliferation, migration and inflammation in VSMCs through inactivation of ROS-mediated PI3K/Akt and MAPK/ERK signaling pathways. Sci. Rep. 2016, 6, 34621. [Google Scholar] [CrossRef] [PubMed]
- Campbell, M.; Trimble, E.R. Modification of PI3K- and MAPK-dependent chemotaxis in aortic vascular smooth muscle cells by protein kinase CbetaII. Circ. Res. 2005, 96, 197–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vigetti, D.; Viola, M.; Karousou, E.; Rizzi, M.; Moretto, P.; Genasetti, A.; Clerici, M.; Hascall, V.C.; De Luca, G.; Passi, A. Hyaluronan-CD44-ERK1/2 regulate human aortic smooth muscle cell motility during aging. J. Biol. Chem. 2008, 283, 4448–4458. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.; Cai, X.; Wu, C.; Wu, L.; Wang, Y.; Liu, Y.; Yu, Z.; Qin, S.; Ma, F.; Thiery, J.P.; et al. Adhesion glycoprotein CD44 functions as an upstream regulator of a network connecting ERK, AKT and Hippo-YAP pathways in cancer progression. Oncotarget 2015, 6, 2951–2965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, J.; Liu, J.; Zhou, C.; Alexander, J.S.; Nanda, A.; Granger, D.N.; Zhang, J.H. Mmp-9 deficiency enhances collagenase-induced intracerebral hemorrhage and brain injury in mutant mice. J. Cereb. Blood Flow Metab. 2004, 24, 1133–1145. [Google Scholar] [CrossRef] [Green Version]
- Kang, L.; Grande, J.P.; Hillestad, M.L.; Croatt, A.J.; Barry, M.A.; Katusic, Z.S.; Nath, K.A. A new model of an arteriovenous fistula in chronic kidney disease in the mouse: Beneficial effects of upregulated heme oxygenase-1. Am. J. Physiol. Renal. Physiol. 2016, 310, F466–F476. [Google Scholar] [CrossRef] [Green Version]
- Kuo, K.L.; Hung, S.C.; Lee, T.S.; Tarng, D.C. Iron sucrose accelerates early atherogenesis by increasing superoxide production and upregulating adhesion molecules in CKD. J. Am. Soc. Nephrol. 2014, 25, 2596–2606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahraeian, S.M.E.; Mohiyuddin, M.; Sebra, R.; Tilgner, H.; Afshar, P.T.; Au, K.F.; Bani Asadi, N.; Gerstein, M.B.; Wong, W.H.; Snyder, M.P.; et al. Gaining comprehensive biological insight into the transcriptome by performing a broad-spectrum RNA-seq analysis. Nat. Commun. 2017, 8, 59. [Google Scholar] [CrossRef]
- Tseng, W.C.; Chuang, C.W.; Yang, M.H.; Pan, C.C.; Tarng, D.C. Kruppel-like factor 4 is a novel prognostic predictor for urothelial carcinoma of bladder and it regulates TWIST1-mediated epithelial-mesenchymal transition. Urol. Oncol. 2016, 34, 485.e415–485.e424. [Google Scholar] [CrossRef]
- Afroze, T.; Yang, L.L.; Wang, C.; Gros, R.; Kalair, W.; Hoque, A.N.; Mungrue, I.N.; Zhu, Z.; Husain, M. Calcineurin-independent regulation of plasma membrane Ca2+ ATPase-4 in the vascular smooth muscle cell cycle. Am. J. Physiol. Cell Physiol. 2003, 285, C88–C95. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.C.; Park, A.Y.; Guan, J.L. In vitro scratch assay: A convenient and inexpensive method for analysis of cell migration in vitro. Nat. Protoc. 2007, 2, 329–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shih, Y.-C.; Chen, P.-Y.; Ko, T.-M.; Huang, P.-H.; Ma, H.; Tarng, D.-C. MMP-9 Deletion Attenuates Arteriovenous Fistula Neointima through Reduced Perioperative Vascular Inflammation. Int. J. Mol. Sci. 2021, 22, 5448. https://doi.org/10.3390/ijms22115448
Shih Y-C, Chen P-Y, Ko T-M, Huang P-H, Ma H, Tarng D-C. MMP-9 Deletion Attenuates Arteriovenous Fistula Neointima through Reduced Perioperative Vascular Inflammation. International Journal of Molecular Sciences. 2021; 22(11):5448. https://doi.org/10.3390/ijms22115448
Chicago/Turabian StyleShih, Yu-Chung, Po-Yuan Chen, Tai-Ming Ko, Po-Hsun Huang, Hsu Ma, and Der-Cherng Tarng. 2021. "MMP-9 Deletion Attenuates Arteriovenous Fistula Neointima through Reduced Perioperative Vascular Inflammation" International Journal of Molecular Sciences 22, no. 11: 5448. https://doi.org/10.3390/ijms22115448
APA StyleShih, Y. -C., Chen, P. -Y., Ko, T. -M., Huang, P. -H., Ma, H., & Tarng, D. -C. (2021). MMP-9 Deletion Attenuates Arteriovenous Fistula Neointima through Reduced Perioperative Vascular Inflammation. International Journal of Molecular Sciences, 22(11), 5448. https://doi.org/10.3390/ijms22115448