
Incorporation of Sulfonamide Moiety into Biguanide Scaffold Results in Apoptosis Induction and Cell Cycle Arrest in MCF-7 Breast Cancer Cells

 ,
,  and
and 
Abstract
:1. Introduction
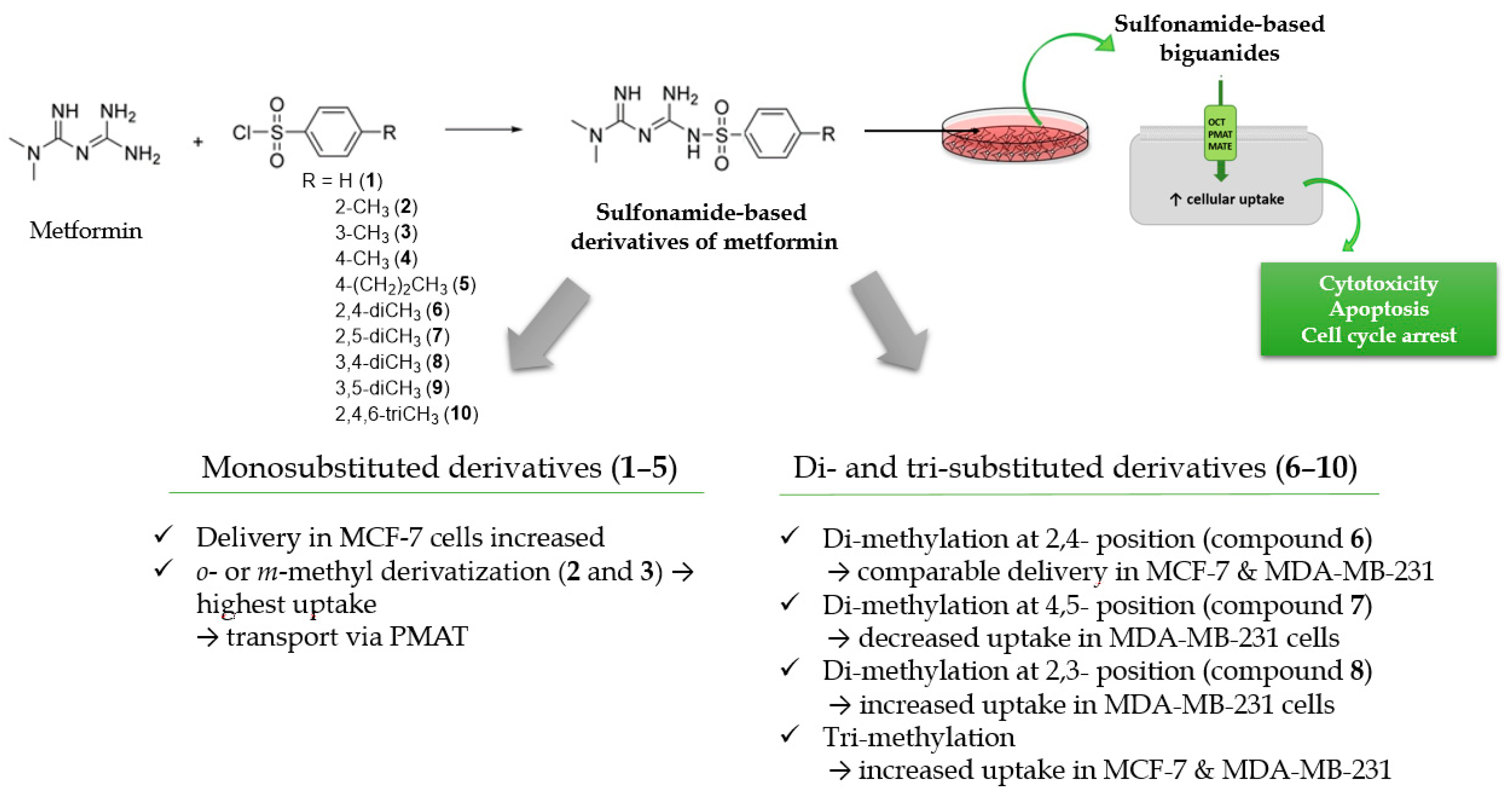
2. Results and Discussion
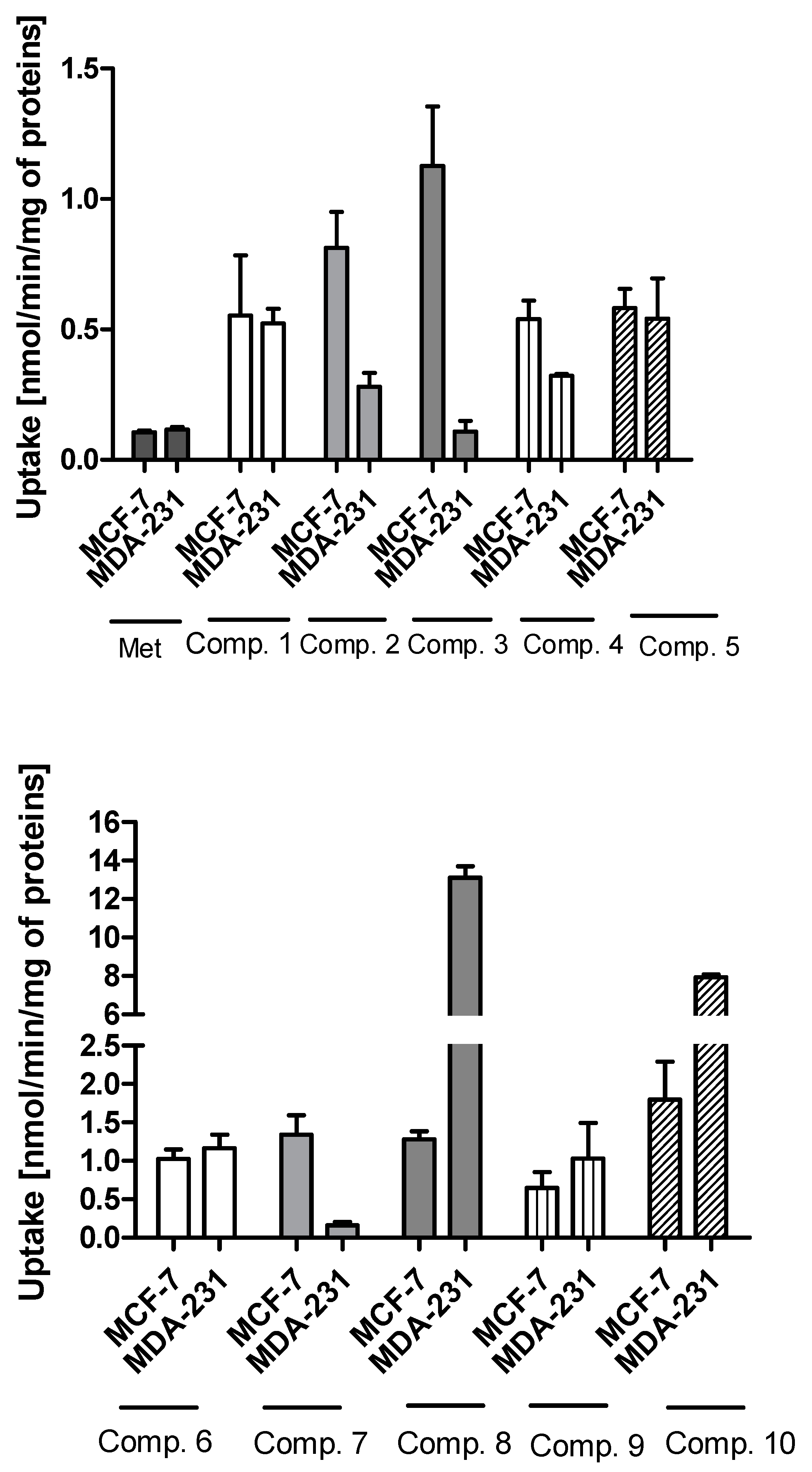
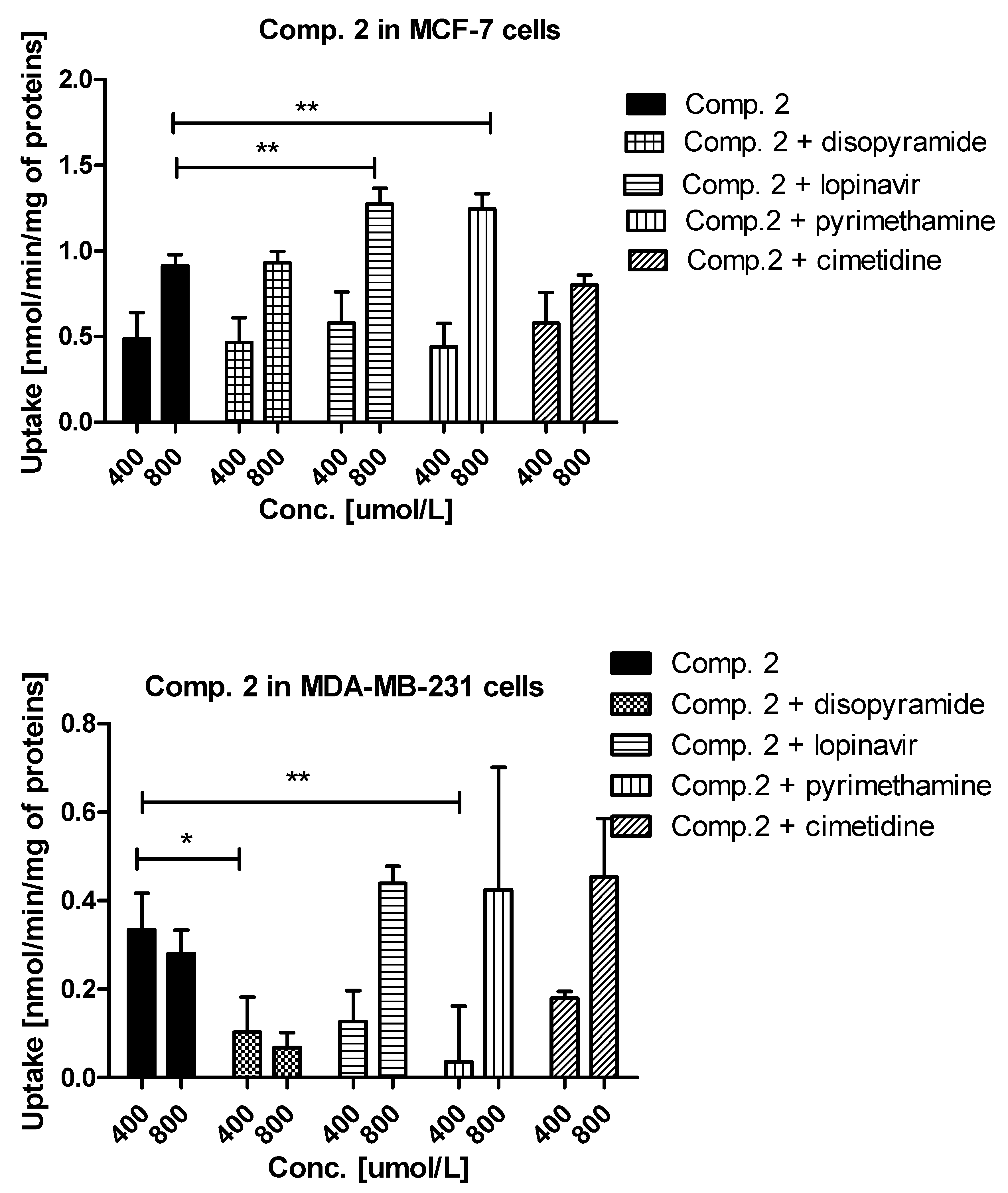
2.1. Cellular Uptake of Metformin Derivatives
2.1.1. General Characterization
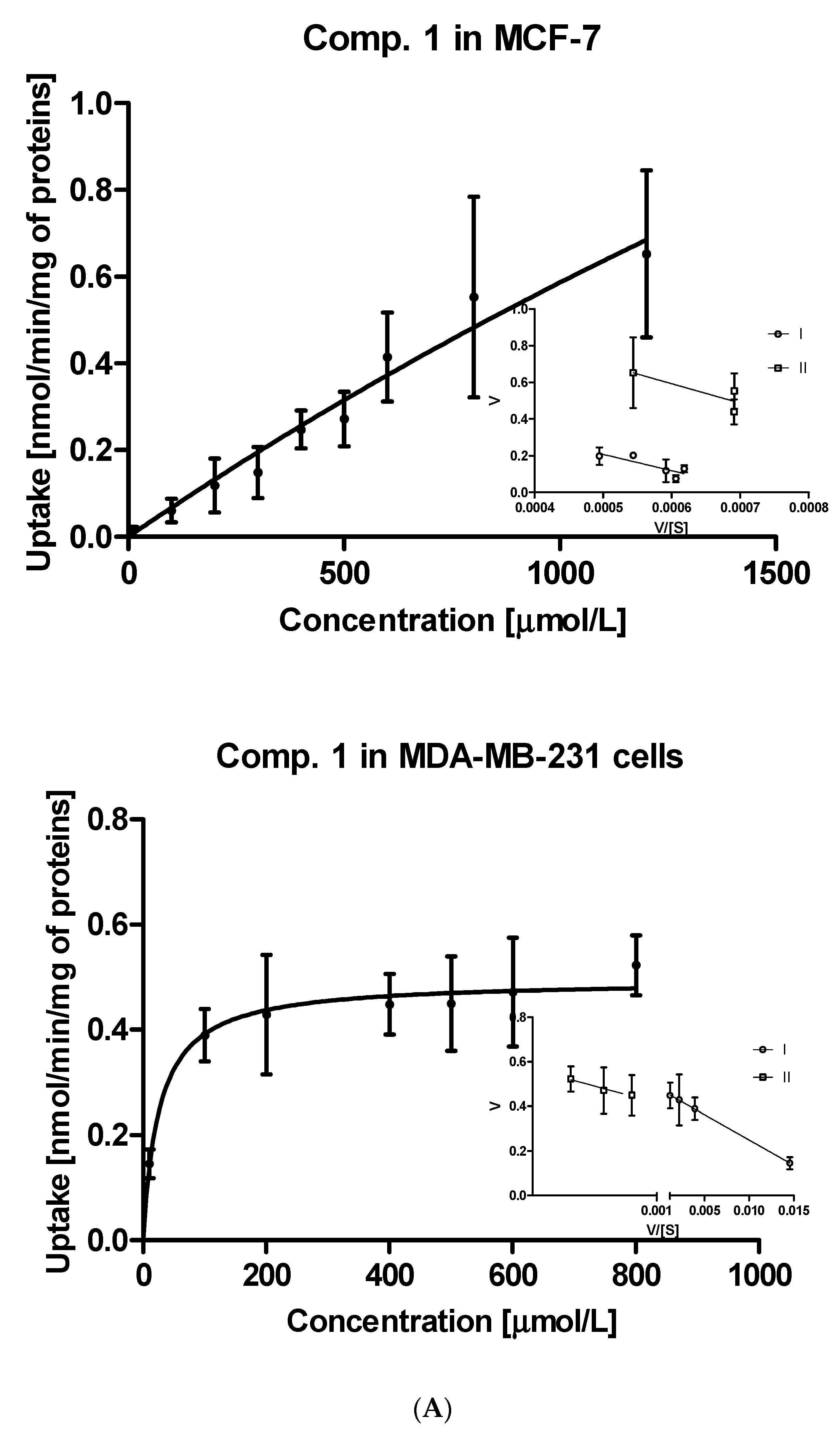
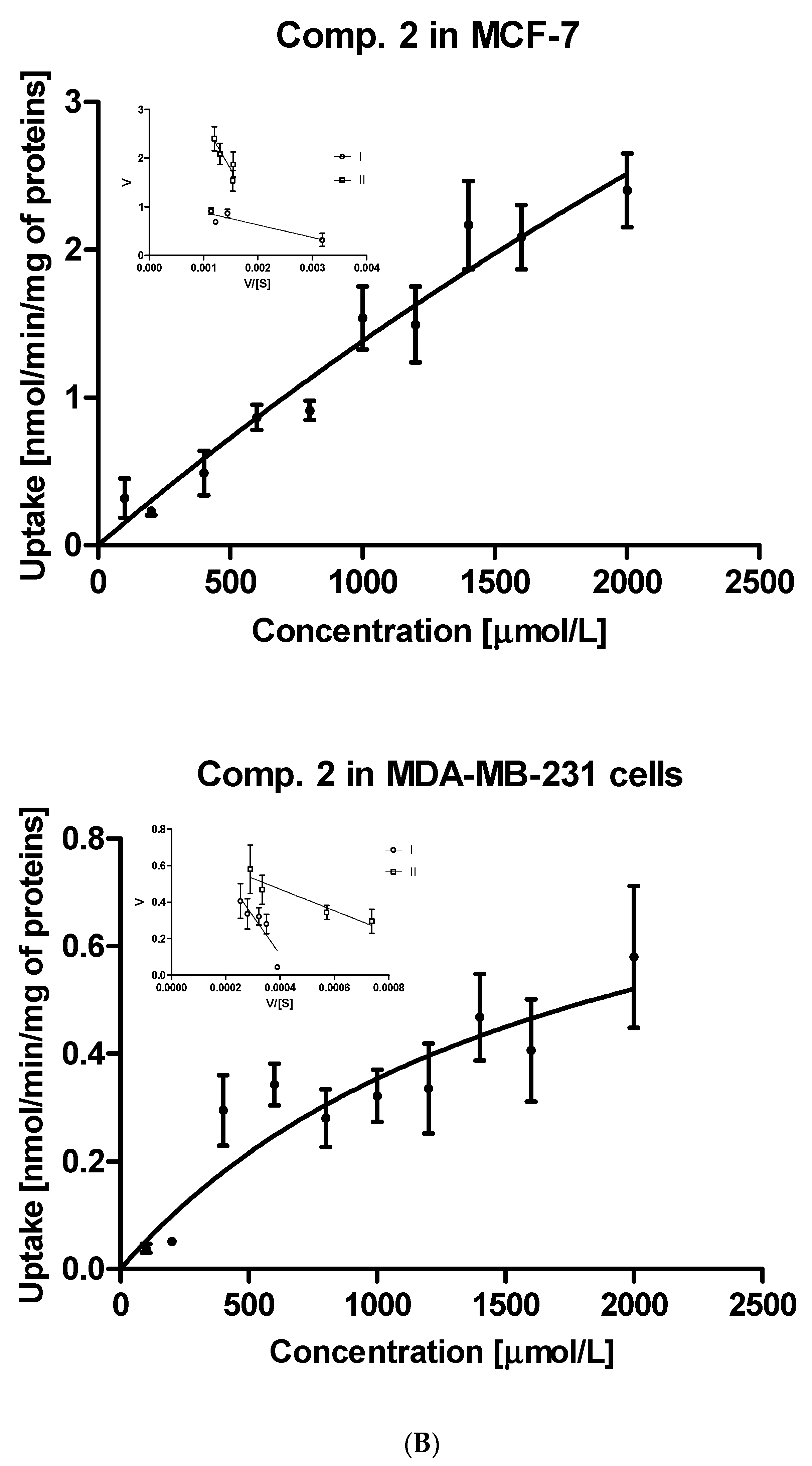
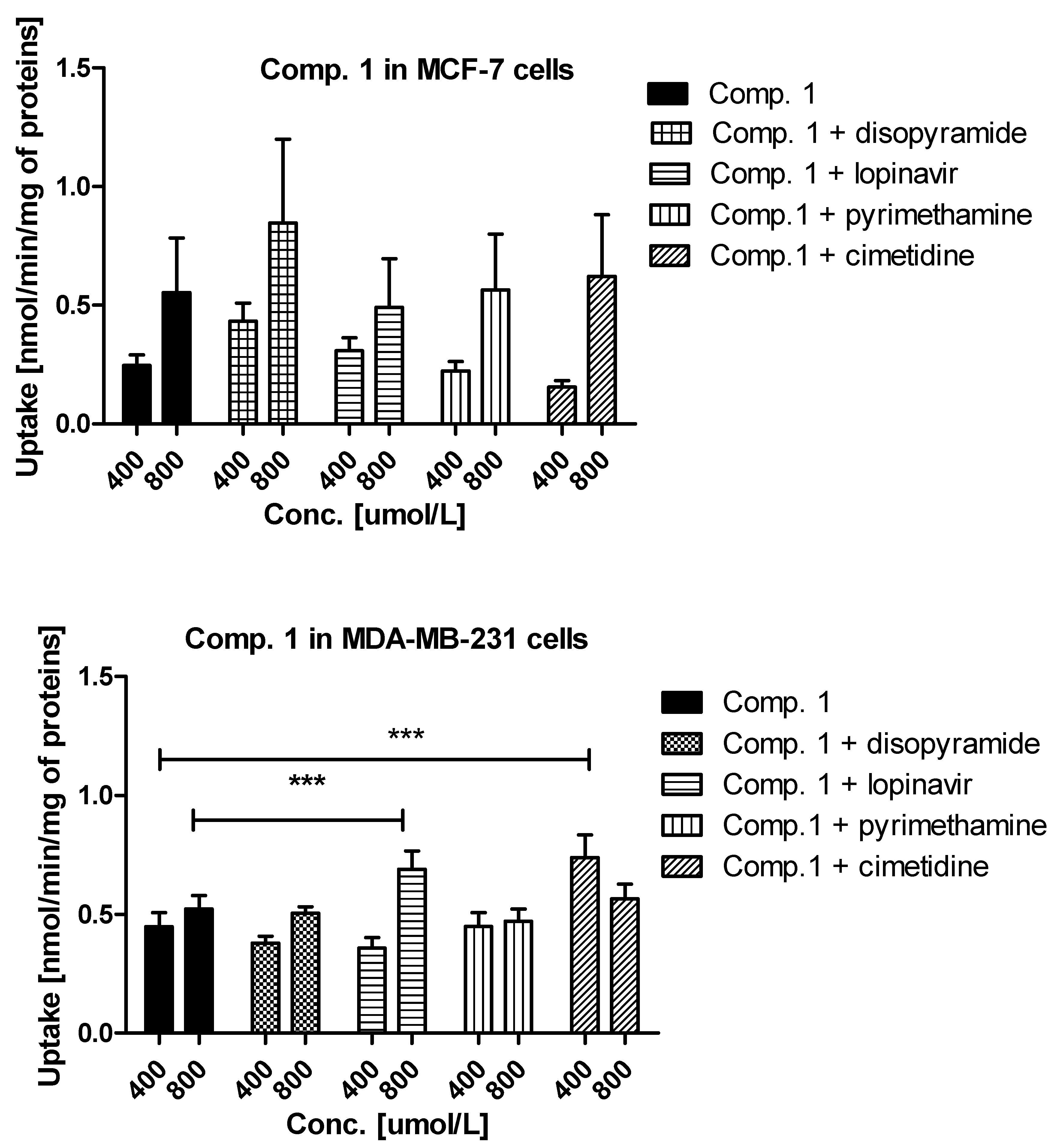
2.1.2. Kinetic Analysis of Sulfonamide Uptake
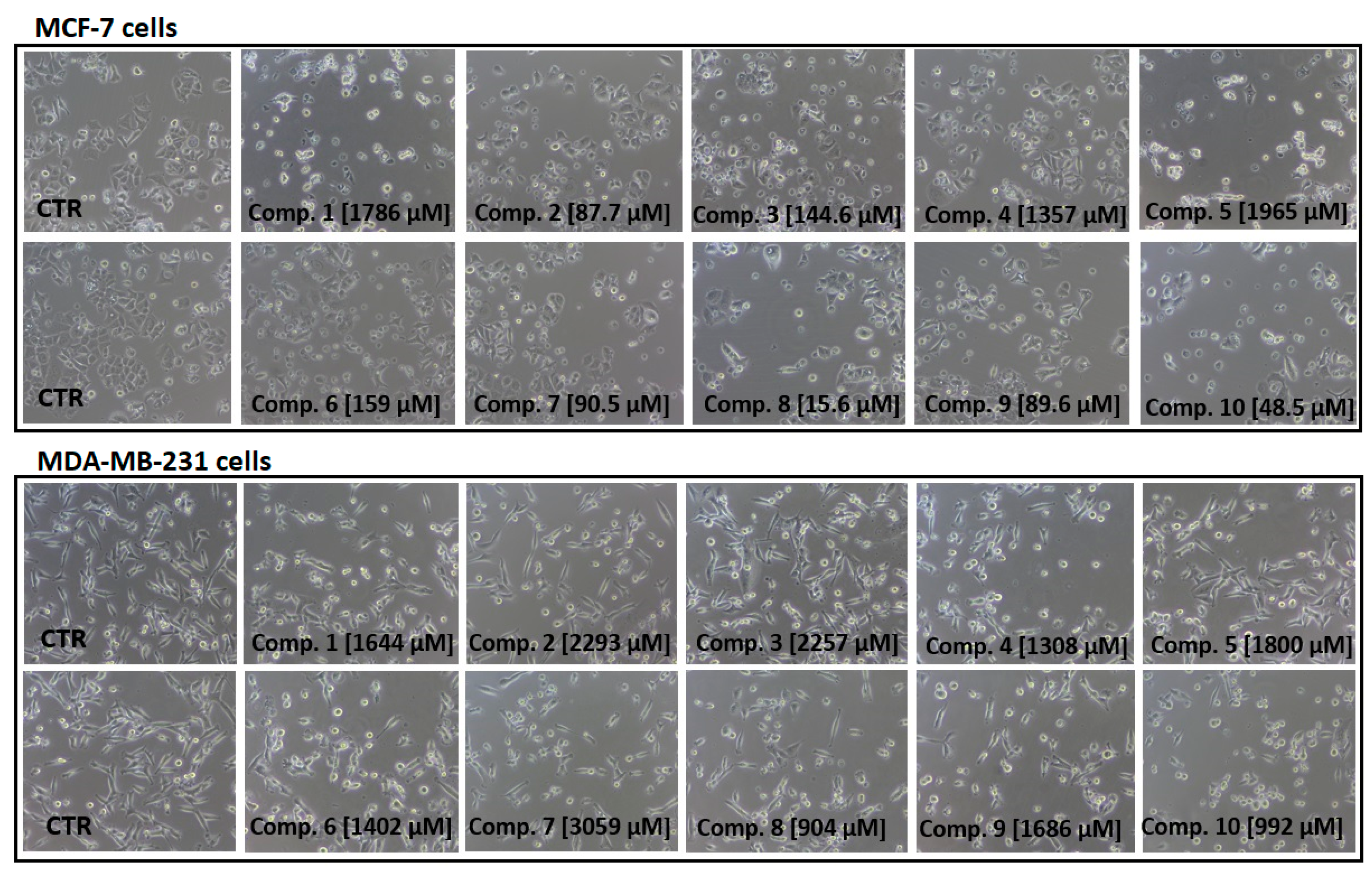
2.2. Cytotoxicity of Compounds 1–10
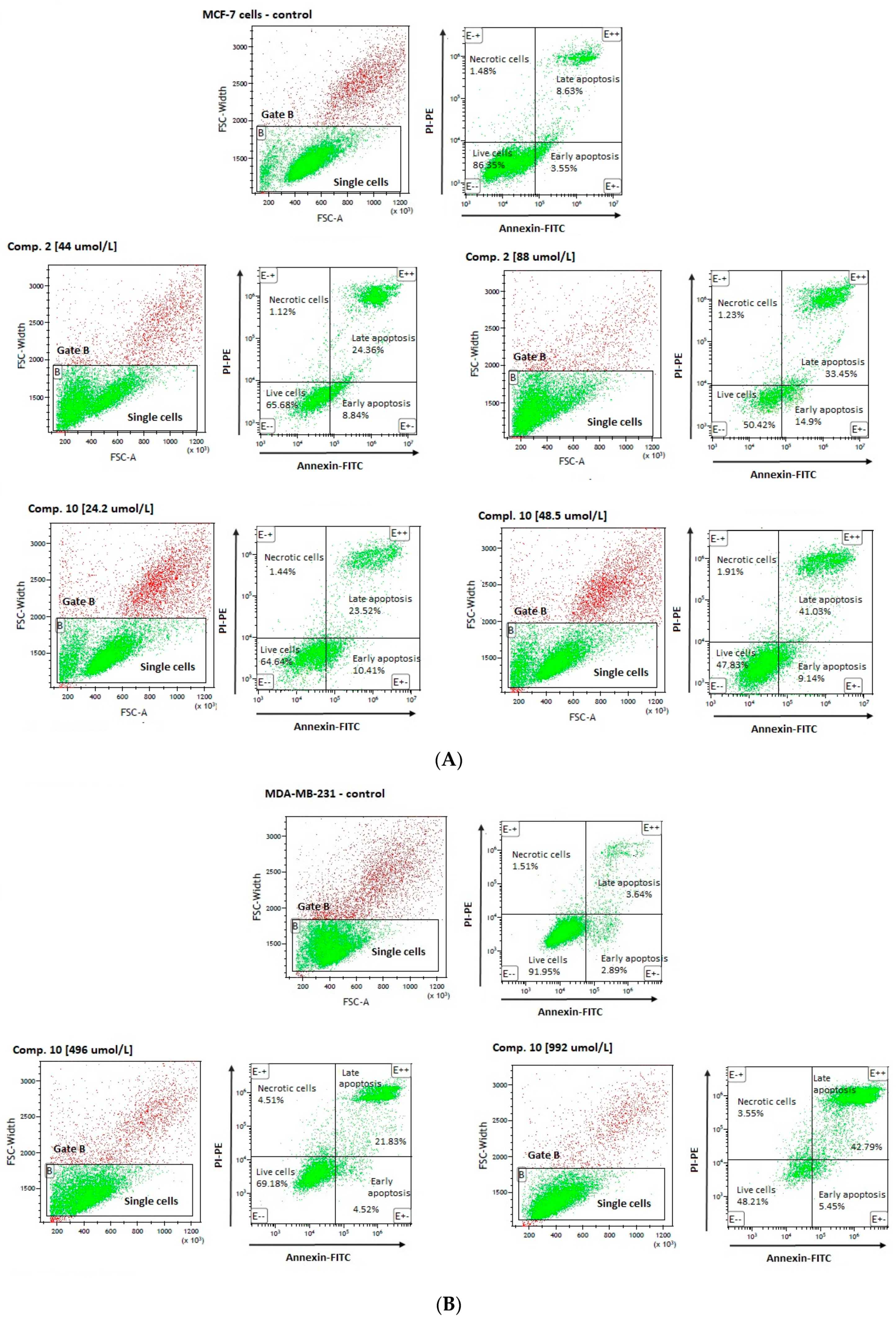
2.3. Apoptosis Assay
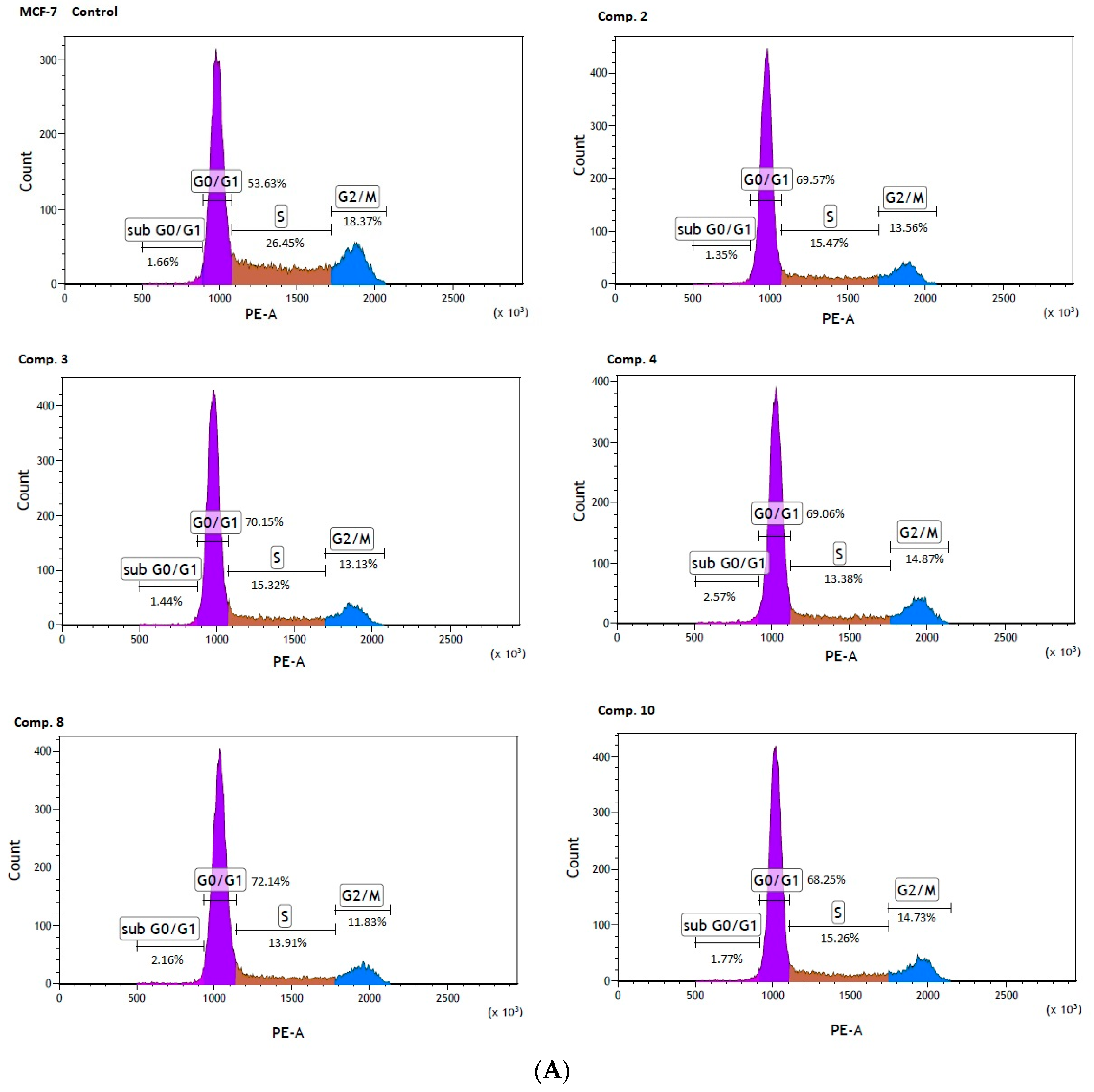
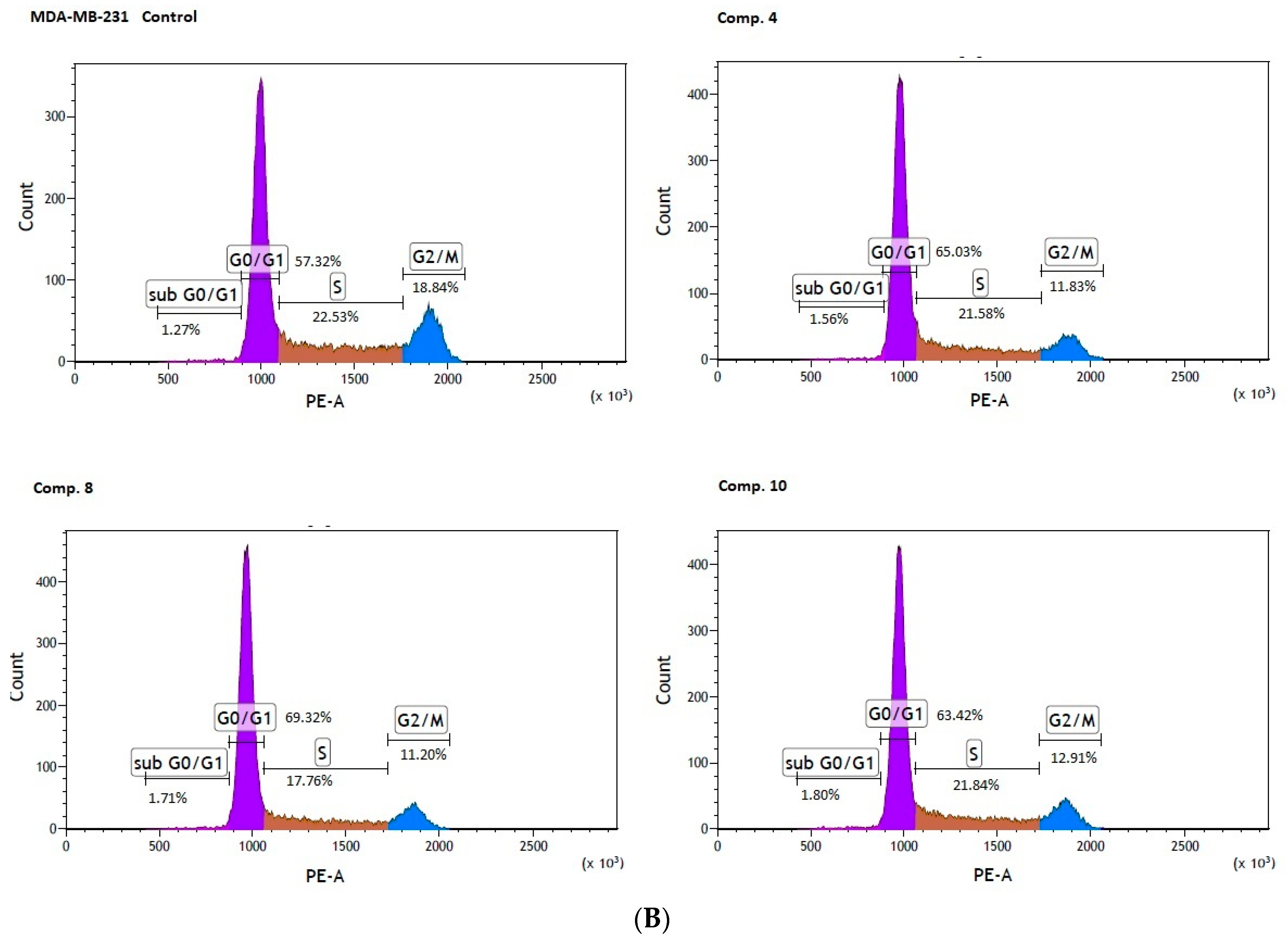
2.4. Cell Cycle Arrest
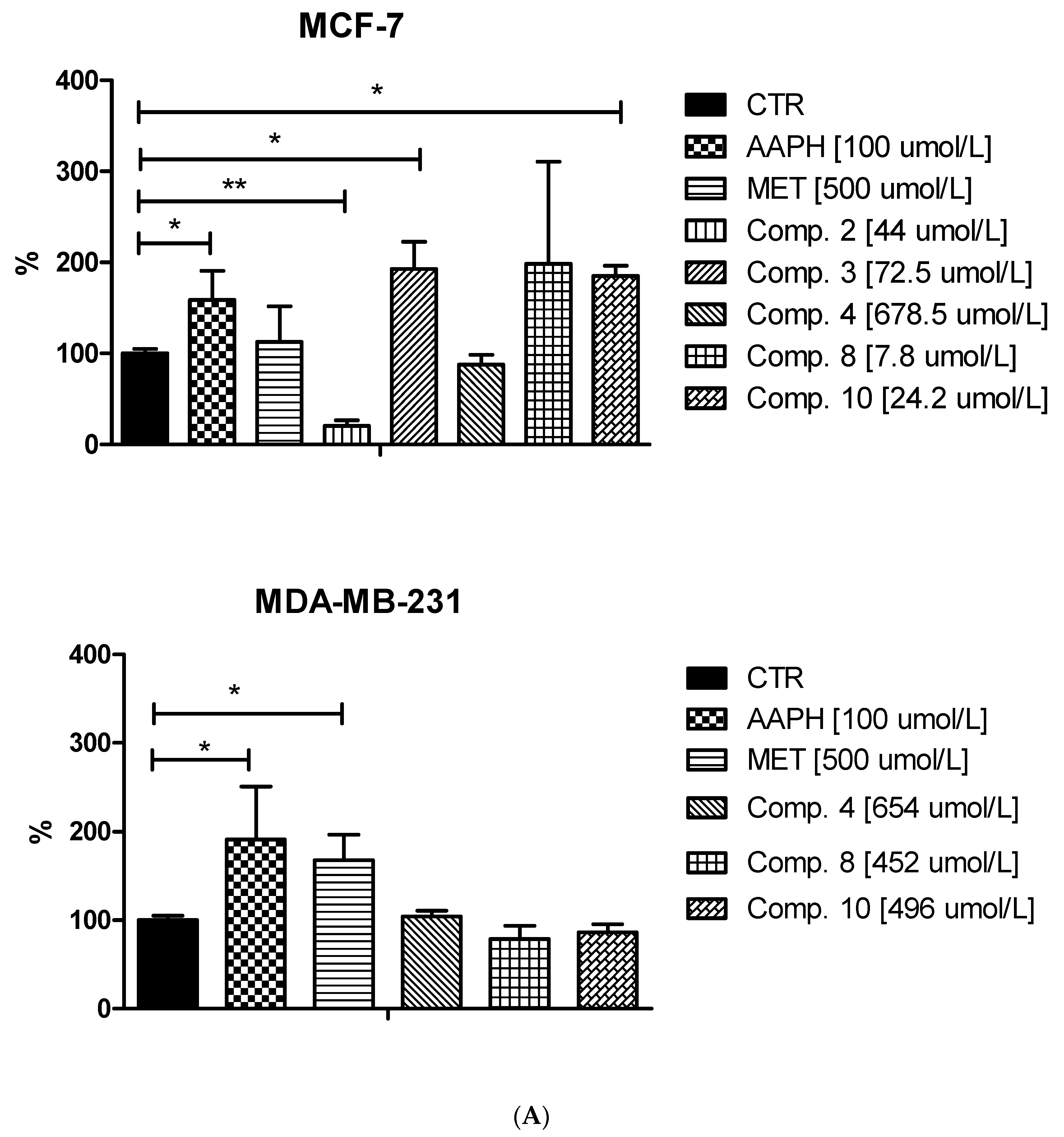
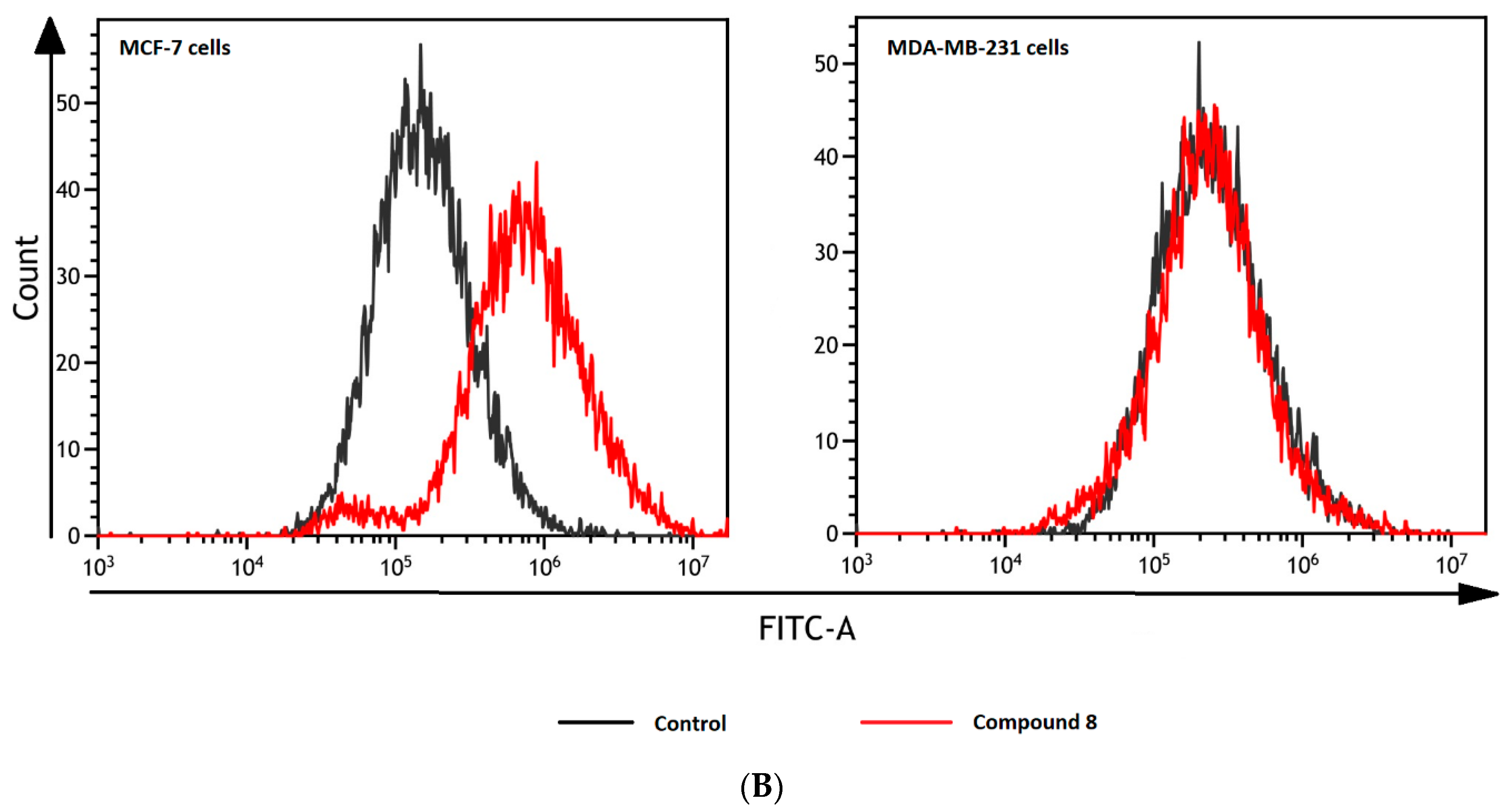
2.5. Intracellular ROS Generation
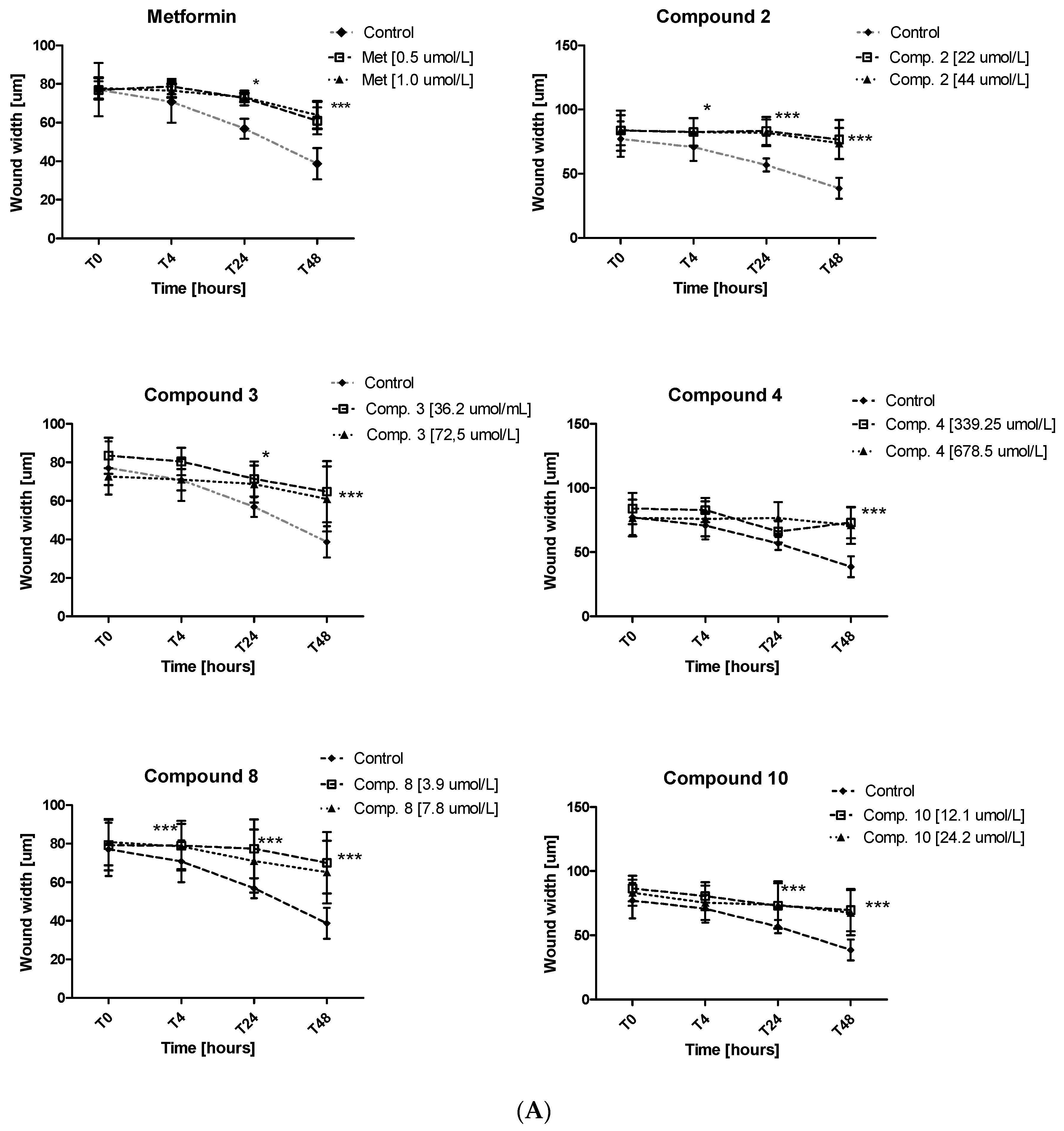
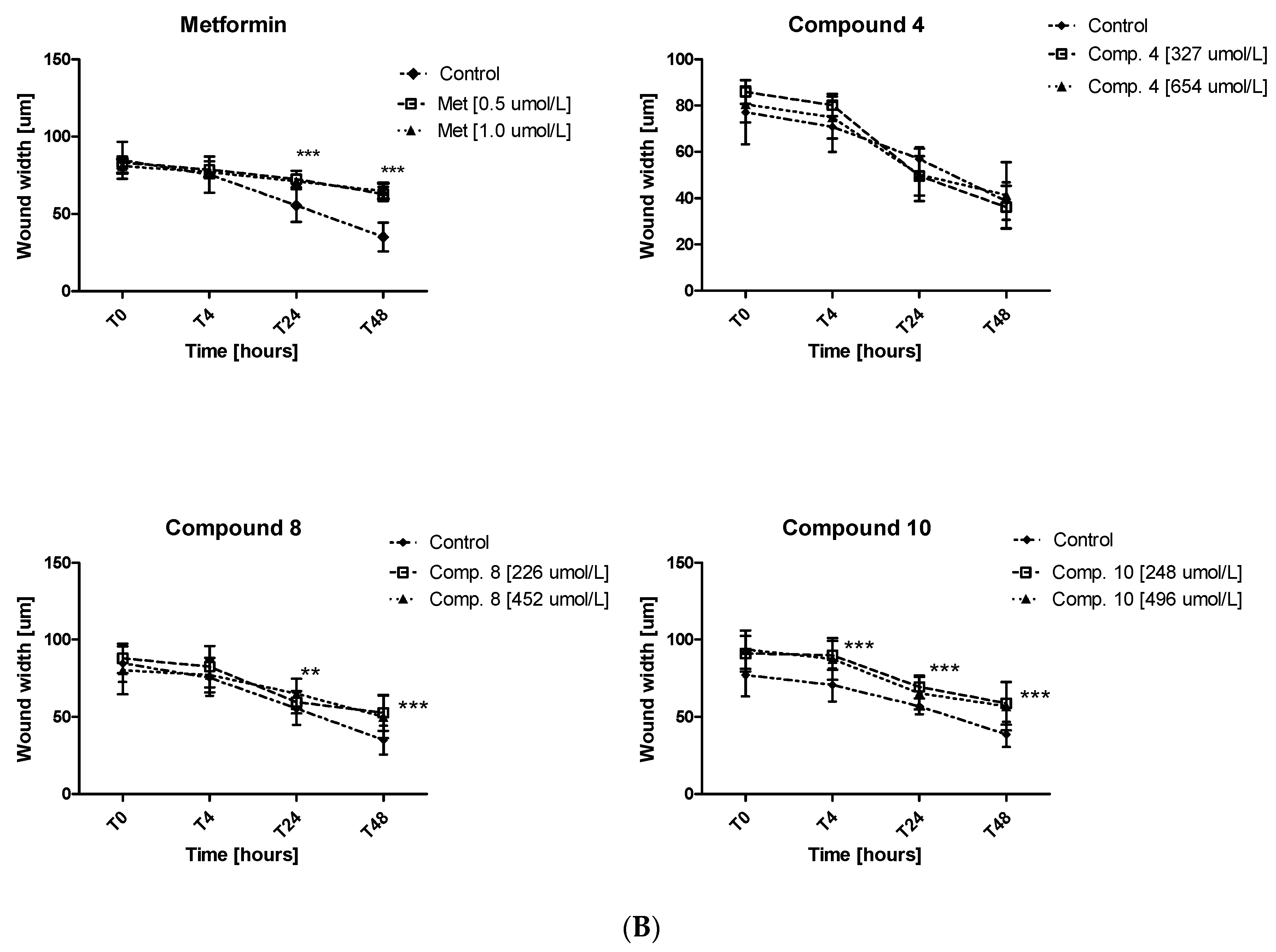
2.6. Migration Test
3. Materials and Methods
3.1. Breast Cancer Cell Culturing
3.2. Studied Compounds
3.3. Uptake Studies of Metformin Derivatives
3.4. Uptake of Metformin Derivatives in the Presence of OCT and MATE Inhibitors
3.5. Cancer Cell Viability and Morphology
3.6. Cell Apoptosis Assay
3.7. Cell Cycle Analysis
3.8. Intracellular ROS Generation
3.9. Wound Healing Assay
3.10. Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dey, G. An Overview of Drug Repurposing: Review Article. J. Med. Sci. Clin. Res. 2019, 7, 3–5. [Google Scholar] [CrossRef]
- Lee, H.M.; Kim, Y. Drug Repurposing Is a New Opportunity for Developing Drugs against Neuropsychiatric Disorders. Schizophr. Res. Treat. 2016, 2016. [Google Scholar] [CrossRef] [Green Version]
- Prattichizzo, F.; Giuliani, A.; Mensà, E.; Sabbatinelli, J.; De Nigris, V.; Rippo, M.R.; La Sala, L.; Procopio, A.D.; Olivieri, F.; Ceriello, A. Pleiotropic effects of metformin: Shaping the microbiome to manage type 2 diabetes and postpone ageing. Ageing Res. Rev. 2018, 48, 87–98. [Google Scholar] [CrossRef]
- Mahmood, K.; Naeem, M.; Ali, N. Metformin: The hidden chronicles of a magic drug. Eur. J. Intern. Med. 2013, 24, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Markowicz-Piasecka, M.; Huttunen, K.M.; Mikiciuk-Olasik, E.; Mateusiak, Ł.; Sikora, J. Metformin-from anti-diabetic drug to anti-cancer drug. Acta Pol. Pharm. Drug Res. 2018, 75, 3–18. [Google Scholar]
- Luo, Q.; Hu, D.; Hu, S.; Yan, M.; Sun, Z.; Chen, F. In vitro and in vivo anti-tumor effect of metformin as a novel therapeutic agent in human oral squamous cell carcinoma. BMC Cancer 2012, 12, 1. [Google Scholar] [CrossRef] [Green Version]
- Yousef, M.; Tsiani, E. Metformin in lung cancer: Review of in vitro and in vivo animal studies. Cancers 2017, 9, 45. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Luo, J.; Yu, T. Anticancer mechanisms of metformin: A review of the current evidence. Life Sci. 2020, 254, 117717. [Google Scholar] [CrossRef] [PubMed]
- Franciosi, M.; Lucisano, G.; Lapice, E.; Strippoli, G.F.M.; Pellegrini, F.; Nicolucci, A. Metformin Therapy and Risk of Cancer in Patients with Type 2 Diabetes: Systematic Review. PLoS ONE 2013, 8, e71583. [Google Scholar] [CrossRef] [PubMed]
- Samsuri, N.A.B.; Leech, M.; Marignol, L. Metformin and improved treatment outcomes in radiation therapy—A review. Cancer Treat. Rev. 2017, 55, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Daugan, M.; Wojcicki, A.D.; d’Hayer, B.; Boudy, V. Metformin: An anti-diabetic drug to fight cancer. Pharmacol. Res. 2016, 113, 675–685. [Google Scholar] [CrossRef]
- Saraei, P.; Asadi, I.; Kakar, M.A.; Moradi-Kor, N. The beneficial effects of metformin on cancer prevention and therapy: A comprehensive review of recent advances. Cancer Manag. Res. 2019, 11, 3295–3313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias Lopes, N.M.; Marinello, P.C.; Sanches, L.J.; da Silva Brito, W.A.; Lovo-Martins, M.I.; Pinge-Filho, P.; Luiz, R.C.; Cecchini, R.; Cecchini, A.L. Patterns of cell death induced by metformin in human MCF-7 breast cancer cells. Pathol. Res. Pract. 2020, 216, 153199. [Google Scholar] [CrossRef]
- De, A.; Kuppusamy, G. Metformin in breast cancer: Preclinical and clinical evidence. Curr. Probl. Cancer 2020, 44, 100488. [Google Scholar] [CrossRef] [PubMed]
- Marinello, P.C.; da Silva, T.N.X.; Panis, C.; Neves, A.F.; Machado, K.L.; Borges, F.H.; Guarnier, F.A.; Bernardes, S.S.; de-Freitas-Junior, J.C.M.; Morgado-Díaz, J.A.; et al. Mechanism of metformin action in MCF-7 and MDA-MB-231 human breast cancer cells involves oxidative stress generation, DNA damage, and transforming growth factor β1 induction. Tumor Biol. 2016, 37, 5337–5346. [Google Scholar] [CrossRef] [PubMed]
- Jiralerspong, S.; Palla, S.L.; Giordano, S.H.; Meric-Bernstam, F.; Liedtke, C.; Barnett, C.M.; Hsu, L.; Hung, M.C.; Hortobagyi, G.N.; Gonzalez-Angulo, A.M. Metformin and pathologic complete responses to neoadjuvant chemotherapy in diabetic patients with breast cancer. J. Clin. Oncol. 2009, 27, 3297–3302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirsch, H.A.; Iliopoulos, D.; Tsichlis, P.N.; Struhl, K. Metformin Selectively Targets Cancer Stem Cells, and Acts Together with Chemotherapy to Block Tumor Growth and Prolong Remission. Cancer Res. 2009, 69, 7509–7511. [Google Scholar] [CrossRef] [Green Version]
- Marinello, P.C.; Panis, C.; Silva, T.N.X.; Binato, R.; Abdelhay, E.; Rodrigues, J.A.L.; Mencalha, A.; Lopes, N.M.D.; Borges, F.H.; Luiz, R.C.; et al. Oxidative stress and TGF-β1 induction by metformin in MCF-7 and MDA-MB-231 human breast cancer cells are accompanied with the downregulation of genes related to cell proliferation, invasion and metastasis. Pathol. Res. Pract. 2020, 216, 153135. [Google Scholar] [CrossRef]
- Cai, H.; Zhang, Y.; Han, T.; Everett, R.S.; Thakker, D.R. Cation-selective transporters are critical to the AMPK-mediated antiproliferative effects of metformin in human breast cancer cells. Int. J. Cancer 2016, 138, 2281–2292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, D.J.; Hu, T.; Wang, J. Polyspecific organic cation transporters and their impact on drug intracellular levels and pharmacodynamics. Pharmacol. Res. 2016, 111, 237–246. [Google Scholar] [CrossRef] [Green Version]
- Markowicz-Piasecka, M.; Huttunen, K.M.; Mateusiak, L.; Mikiciuk-Olasik, E.; Sikora, J. Is Metformin a Perfect Drug? Updates in Pharmacokinetics and Pharmacodynamics. Curr. Pharm. Des. 2017, 23, 2532–2550. [Google Scholar] [CrossRef]
- Maideen, N.M.P.; Jumale, A.; Balasubramaniam, R. Drug interactions of metformin involving drug transporter proteins. Adv. Pharm. Bull. 2017, 7, 501–505. [Google Scholar] [CrossRef] [Green Version]
- Graham, G.G.; Punt, J.; Arora, M.; Day, R.O.; Doogue, M.P.; Duong, J.K.; Furlong, T.J.; Greenfield, J.R.; Greenup, L.C.; Kirkpatrick, C.M.; et al. Clinical pharmacokinetics of metformin. Clin. Pharmacokinet. 2011, 50, 81–98. [Google Scholar] [CrossRef]
- Cheng, G.; Zielonka, J.; Ouari, O.; Lopez, M.; McAllister, D.; Boyle, K.; Barrios, C.S.; Weber, J.J.; Johnson, B.D.; Hardy, M.; et al. Mitochondria-targeted analogues of metformin exhibit enhanced antiproliferative and radiosensitizing effects in pancreatic cancer cells. Cancer Res. 2016, 76, 3904–3915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markowicz-Piasecka, M.; Huttunen, J.; Sikora, J.; Huttunen, K.M. Sulfenamide derivatives can improve transporter-mediated cellular uptake of metformin and induce cytotoxicity in human breast adenocarcinoma cell lines. Bioorg. Chem. 2019, 87, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Markowicz-Piasecka, M.; Komeil, I.; Huttunen, J.; Sikora, J.; Huttunen, K.M. Effective Cellular Transport of Ortho -Halogenated Sulfonamide Derivatives of Metformin Is Related to Improved Antiproliferative Activity and Apoptosis Induction in MCF-7 Cells. Int. J. Mol. Sci. 2020, 21, 2389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houston, J.B.; Kenworthy, K.E. In vitro-in vivo scaling of CYP kinetic data not consistent with the classical Michaelis-Menten model. Drug Metab. Dispos. 2000, 28, 246–254. [Google Scholar] [PubMed]
- Checkley, L.A.; Rudolph, M.C.; Wellberg, E.A.; Giles, E.D.; Wahdan-Alaswad, R.S.; Houck, J.A.; Edgerton, S.M.; Thor, A.D.; Schedin, P.; Anderson, S.M.; et al. Metformin accumulation correlates with organic cation transporter 2 protein expression and predicts mammary tumor regression in vivo. Cancer Prev. Res. 2017, 10, 198–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markowicz-Piasecka, M.; Sadkowska, A.; Sikora, J.; Broncel, M.; Huttunen, K.M. Novel Sulfonamide-Based Analogs of Metformin Exert Promising Anti-Coagulant Effects without Compromising Glucose-Lowering Activity. Pharmaceuticals 2020, 13, 323. [Google Scholar] [CrossRef] [PubMed]
- Yasmeen, A.; Beauchamp, M.C.; Piura, E.; Segal, E.; Pollak, M.; Gotlieb, W.H. Induction of apoptosis by metformin in epithelial ovarian cancer: Involvement of the Bcl-2 family proteins. Gynecol. Oncol. 2011, 121, 492–498. [Google Scholar] [CrossRef]
- Moro, M.; Caiola, E.; Ganzinelli, M.; Zulato, E.; Rulli, E.; Marabese, M.; Centonze, G.; Busico, A.; Pastorino, U.; de Braud, F.G.; et al. Metformin Enhances Cisplatin-Induced Apoptosis and Prevents Resistance to Cisplatin in Co-mutated KRAS/LKB1 NSCLC. J. Thorac. Oncol. 2018, 13, 1692–1704. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Kong, P.; Wang, J.; Xu, Y.; Gao, C.; Guo, G. Effects of metformin on proliferation and apoptosis of human megakaryoblastic Dami and MEG-01 cells. J. Pharmacol. Sci. 2017, 135, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Markowicz-Piasecka, M.; Sikora, J.; Zajda, A.; Huttunen, K.M. Novel halogenated sulfonamide biguanides with anti-coagulation properties. Bioorg. Chem. 2020, 94, 103444. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, W.; Yan, Z.; Zhao, W.; Mi, J.; Li, J.; Yan, H. Metformin induces autophagy and G0/G1 phase cell cycle arrest in myeloma by targeting the AMPK/mTORC1 and mTORC2 pathways. J. Exp. Clin. Cancer Res. 2018, 37, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.H.; Kim, Y.; Lee, B.; Han, J.; Kim, H.K.; Shim, Y.M.; Kim, D.H. Metformin induces cell cycle arrest at the G1 phase through E2F8 suppression in lung cancer cells. Oncotarget 2017, 8, 101509–101519. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Wen, Z.H.; Jia, C.H.; Li, M.; Luo, S.Q.; Bai, X.C. Metformin Induces G1 Cell Cycle Arrest and Inhibits Cell Proliferation in Nasopharyngeal Carcinoma Cells. Anat. Rec. 2011, 294, 1337–1343. [Google Scholar] [CrossRef] [PubMed]
- Jingwen, B.; Yaochen, L.; Guojun, Z. Cell cycle regulation and anticancer drug discovery. Cancer Biol. Med. 2017, 14, 348. [Google Scholar] [CrossRef] [PubMed]
- Geou-Yarh Liou, P.S. Reactive oxygen species in cancer. Free Radic. Res. 2010, 44, 479–496. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Roper, M.G. Measurement of DCF fluorescence as a measure of reactive oxygen species in murine islets of Langerhans. Anal. Methods 2014, 6, 3019–3024. [Google Scholar] [CrossRef] [Green Version]
- Ariaans, G.; Jalving, M.; de Vries, E.G.E.; de Jong, S. Anti-tumor effects of everolimus and metformin are complementary and glucose-dependent in breast cancer cells. BMC Cancer 2017, 17, 232. [Google Scholar] [CrossRef] [Green Version]
- Li, S.M.H.; Liu, S.T.; Chang, Y.L.; Ho, C.L.; Huang, S.M. Metformin causes cancer cell death through downregulation of p53-dependent differentiated embryo chondrocyte 1. J. Biomed. Sci. 2018, 25, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.C.; Li, H.; Wang, J. Mechanisms of metformin inhibiting cancer invasion and migration. Am. J. Transl. Res. 2020, 12, 4885–4901. [Google Scholar] [PubMed]
- He, L.; Wondisford, F.E. Metformin action: Concentrations matter. Cell Metab. 2015, 21, 159–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 | |||
|---|---|---|---|
| Compound | R Group | Mw (g/mol) | cLogD a |
| Metformin | - | 129.17 | −1.32 |
| 1 | H | 269.32 | 0.80 |
| 2 | 2-CH3 | 283.35 | 1.30 |
| 3 | 3-CH3 | 283.35 | 1.30 |
| 4 | 4-CH3 | 283.35 | 1.30 |
| 5 | 4-(CH2)2CH3 | 297.38 | 1.47 |
| 6 | 2,4-diCH3 | 297.38 | 1.80 |
| 7 | 2,5-diCH3 | 297.38 | 1.80 |
| 8 | 3,4-diCH3 | 297.38 | 1.75 |
| 9 | 3,5-diCH3 | 297.38 | 1.80 |
| 10 | 2,4,6-triCH3 | 311.40 | 2.30 |
| Kinetic Parameters of Sulfonamide Uptake | ||||||
|---|---|---|---|---|---|---|
| MCF-7 Cells | MDA-231 Cells | |||||
| Compound | Km [μmol/L] | Vmax [nmol/min/mg] | Vmax/Km | Km [μmol/L] | Vmax [nmol/min/mg] | Vmax/Km |
| Metformin | 5583 ± 1560 # | 0.801 ± 0.296 # | 0.00014 # | 3375.0 ± 952 # | 0.718 ± 0.181 # | 0.000212 # |
| 1 | 6170 ± 778 | 4.206 ± 2.666 | 0.00068 | 26.21 ± 8.798 | 0.4946 ± 0.021 | 0.01887 |
| 2 | 9070 ± 3913 | 13.92 ± 6.151 | 0.00153 | 1784 ± 714.1 | 0.985 ± 0.293 | 0.00055 |
| 3 | 312.9 ± 116.7 | 1.684 ± 0.168 | 0.00538 | NE | NE | NE |
| 4 | NE | NE | NE | 716.6 ± 187.6 | 0.563 ± 0.132 | 0.00078 |
| 5 | 4508 ± 1992 | 3.664 ± 2.056 | 0.00081 | 1178 ± 525.3 | 1.292 ± 0.360 | 0.00109 |
| 6 | 819.5 ± 251.4 | 2.180 ± 0.399 | 0.00266 | NE | NE | NE |
| 7 | 613.2 ± 202.8 | 2.291 ± 0.321 | 0.00374 | 167.6 ± 91.38 | 0.225 ± 0.024 | 0.00134 |
| 8 | 3454 ± 1103 | 5.787 ± 2.520 | 0.00167 | 3140 ± 1876 | 65.66 ± 27.85 | 0.02091 |
| 9 | 2601 ± 849 | 3.174 ± 1.619 | 0.00122 | NE | NE | NE |
| 10 | 843.6 ± 256.1 | 3.243 ± 0.593 | 0.00384 | 110.6 ± 8.991 | 8.789 ± 0.119 | 0.07946 |
| Compound | MCF-7 Cells [µmol/L] | MDA-MB-231 Cells [µmol/L] |
|---|---|---|
| 1 | 1786 ± 123.8 | 1644 ± 122.87 |
| 2 | 87.7 ± 1.18 | 2293 ± 1261 |
| 3 | 144.6 ± 12.2 | 2257 ± 1095 |
| 4 | 1357 ± 119.7 | 1308 ± 118.7 |
| 5 | 1965 ± 136.1 | 1800 ± 142.42 |
| 6 | 159.0 ± 12.34 | 1402 ± 119.2 |
| 7 | 90.49 ± 11.79 | 3059 ± 1291 |
| 8 | 15.65 ± 1.22 | 903.9 ± 115.6 |
| 9 | 89.56 ± 11.7 | 1686 ± 124.3 |
| 10 | 48.46 ± 11.79 | 992.1 ± 115.9 |
| Compound [μmol/L] | Living Cells [E − −] [%] | Necrotic Cells [E − +] [%] | Early Apoptotic [E + −] [%] | Late Apoptotic [E + +] [%] |
|---|---|---|---|---|
| Control MCF-7 | 83.57 ± 3.60 | 1.92 ± 0.67 | 5.29 ± 1.13 | 9.23 ± 2.91 |
| Comp. 2 [44 μmol/L] | 67.49 ± 4.12 *** | 1.78 ± 1.84 | 9.95 ± 3.93 * | 20.74 ± 6.80 * |
| Comp. 2 [88 μmol/L] | 51.39 ± 4.15 *** | 1.19 ± 0.05 * | 10.04 ± 4.90 * | 37.38 ± 8.36 *** |
| Comp. 3 [72.5 μmol/L] | 69.50 ± 5.08 ** | 0.88 ± 0.15 ** | 9.27 ± 4.24 # | 23.68 ± 12.42 * |
| Comp. 3 [145 μmol/L] | 21.15 ± 3.92 ** | 0.56 ± 0.14 * | 7.16 ± 1.92 | 71.13 ± 2.14 ** |
| Comp. 4 [678.5 μmol/L] | 61.14 ± 0.74 ** | 1.41 ± 0.30 | 12.41 ± 2.03 * | 25.03 ± 1.60 *** |
| Comp. 4 [1357 μmol/mL] | 31.32 ± 4.67 ** | 2.44 ± 0.65 * | 7.91 ± 1.32 * | 58.32 ± 4.58 ** |
| Comp. 8 [7.8 μmol/L] | 70.75 ± 2.68 ** | 1.74 ± 0.29 | 7.34 ± 2.01 * | 23.90 ± 3.64 ** |
| Comp. 8 [15.6 μmol/L] | 30.72 ± 1.10 ** | 1.07 ± 0.09 * | 9.07 ± 0.26 * | 59.14 ± 1.21 *** |
| Comp. 10 [24.2 μmol/L] | 65.58 ± 3.41 ** | 1.64 ± 0.24 | 9.57 ± 1.67 * | 23.21 ± 1.99 ** |
| Comp. 10 [48.5 μmol/L] | 47.48 ± 1.70 ** | 1.93 ± 0.22 | 8.93 ± 0.22 | 41.65 ± 1.05 ** |
| Compound [μmol/L] | Living Cells [E − −] [%] | Necrotic Cells [E − +] [%] | Early Apoptotic [E + −] [%] | Late Apoptotic [E + +] [%] |
|---|---|---|---|---|
| Control MDA-MB-231 | 86.20 ± 5.45 | 1.92 ± 0.60 | 6.12 ± 2.94 | 5.75 ± 2.37 |
| Comp. 4 [654 μmol/L] | 80.76 ± 2.22 * | 1.61 ± 0.51 | 4.78 ± 0.62 * | 12.84 ± 1.40 ** |
| Comp. 4 [1308 μmol/L] | 49.68 ± 1.41 *** | 1.10 ± 0.11 | 3.90 ± 0.39 * | 45.36 ± 1.64 *** |
| Comp. 8 [452 μmol/L] | 80.75 ± 0.92 | 0.60 ± 0.36 ** | 1.61 ± 1.17 * | 17.02 ± 2.39 * |
| Comp. 8 [904 μmol/L] | 34.94 ± 8.78 ** | 4.04 ± 0.80 | 3.07 ± 1.06 ** | 57.94 ± 8.65 ** |
| Comp. 10 [496 μmol/L] | 69.21 ± 0.89 * | 4.50 ± 0.89 * | 3.94 ± 0.59 * | 22.34 ± 1.48 * |
| Comp. 10 [992 μmol/mL] | 50.44 ± 4.35 ** | 3.65 ± 0.39 * | 5.01 ± 0.42 ** | 40.90 ± 4.04 ** |
| Compound [µmol/L] | Sub G0/G1 [%] | G0/G1 [%] | S [%] | G2/M [%] |
|---|---|---|---|---|
| Control MCF-7 | 1.24 ± 0.70 | 52.30 ± 1.84 | 26.01 ± 0.70 | 20.45 ± 1.30 |
| Comp. 2 [88 µmol/L] | 1.55 ± 0.18 | 69.69 ± 0.94 *** | 15.42 ± 0.71 *** | 13.28 ± 0.38 *** |
| Comp. 3 [145 µmol/L] | 1.47 ± 0.07 | 70.34 ± 0.71 *** | 14.89 ± 0.38 *** | 13.27 ± 0.58 *** |
| Comp. 4 [1357 µmol/L] | 1.96 ± 0.60 | 70.63 ± 1.47 *** | 12.82 ± 0.50 *** | 14.54 ± 0.31 *** |
| Comp. 8 [15.6 µmol/L] | 2.75 ± 0.51 *** | 72.79 ± 0.56 *** | 12.83 ± 1.19 *** | 11.61 ± 0.74 *** |
| Comp. 10 [48.5 µmol/L] | 1.71 ± 0.19 | 68.23 ± 0.60 *** | 15.52 ± 0.22 *** | 14.54 ± 0.43 *** |
| Control MDA-MB-231 | 1.40 ± 0.14 | 57.09 ± 0.91 | 23.27 ± 0.65 | 18.20 ± 0.99 |
| Comp. 4 [1308 µmol/L] | 1.77 ± 0.19 | 65.77 ± 0.93 | 20.95 ± 0.71 | 11.54 ± 1.11 *** |
| Comp. 8 [904 µmol/L] | 1.83 ± 0.21 * | 70.32 ± 1.95 ** | 16.61 ± 1.36 ** | 11.20 ± 0.96 *** |
| Comp. 10 [992 µmol/L] | 1.73 ± 0.19 | 60.65 ± 7.04 | 20.65 ± 1.05 * | 13.07 ± 0.30 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Markowicz-Piasecka, M.; Sadowski, K.; Huttunen, J.; Sikora, J.; Huttunen, K.M. Incorporation of Sulfonamide Moiety into Biguanide Scaffold Results in Apoptosis Induction and Cell Cycle Arrest in MCF-7 Breast Cancer Cells. Int. J. Mol. Sci. 2021, 22, 5642. https://doi.org/10.3390/ijms22115642
Markowicz-Piasecka M, Sadowski K, Huttunen J, Sikora J, Huttunen KM. Incorporation of Sulfonamide Moiety into Biguanide Scaffold Results in Apoptosis Induction and Cell Cycle Arrest in MCF-7 Breast Cancer Cells. International Journal of Molecular Sciences. 2021; 22(11):5642. https://doi.org/10.3390/ijms22115642
Chicago/Turabian StyleMarkowicz-Piasecka, Magdalena, Karol Sadowski, Johanna Huttunen, Joanna Sikora, and Kristiina M. Huttunen. 2021. "Incorporation of Sulfonamide Moiety into Biguanide Scaffold Results in Apoptosis Induction and Cell Cycle Arrest in MCF-7 Breast Cancer Cells" International Journal of Molecular Sciences 22, no. 11: 5642. https://doi.org/10.3390/ijms22115642
APA StyleMarkowicz-Piasecka, M., Sadowski, K., Huttunen, J., Sikora, J., & Huttunen, K. M. (2021). Incorporation of Sulfonamide Moiety into Biguanide Scaffold Results in Apoptosis Induction and Cell Cycle Arrest in MCF-7 Breast Cancer Cells. International Journal of Molecular Sciences, 22(11), 5642. https://doi.org/10.3390/ijms22115642