
Gonadotropin Releasing Hormone (GnRH) Triggers Neurogenesis in the Hypothalamus of Adult Zebrafish


Abstract
:1. Introduction
2. Results
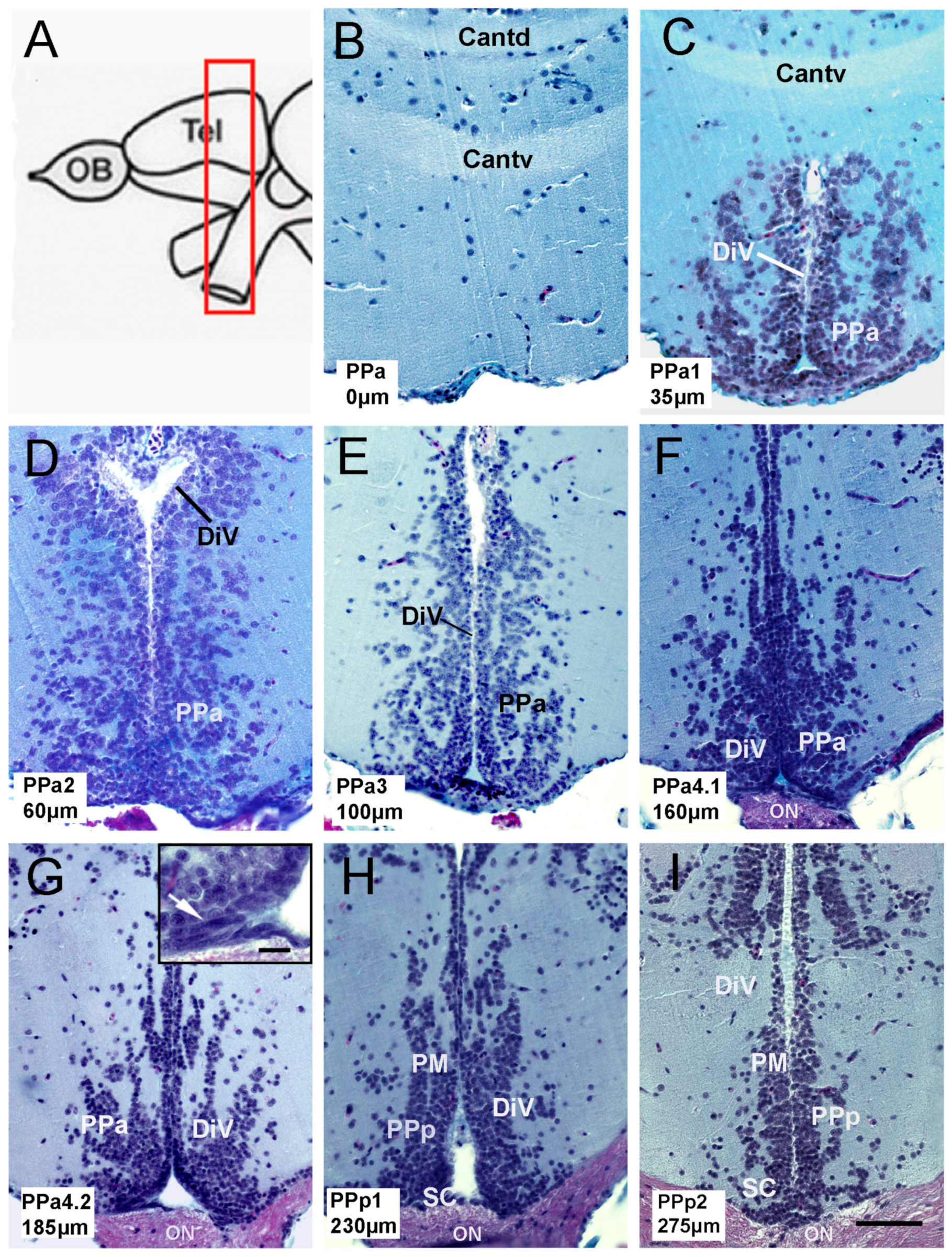
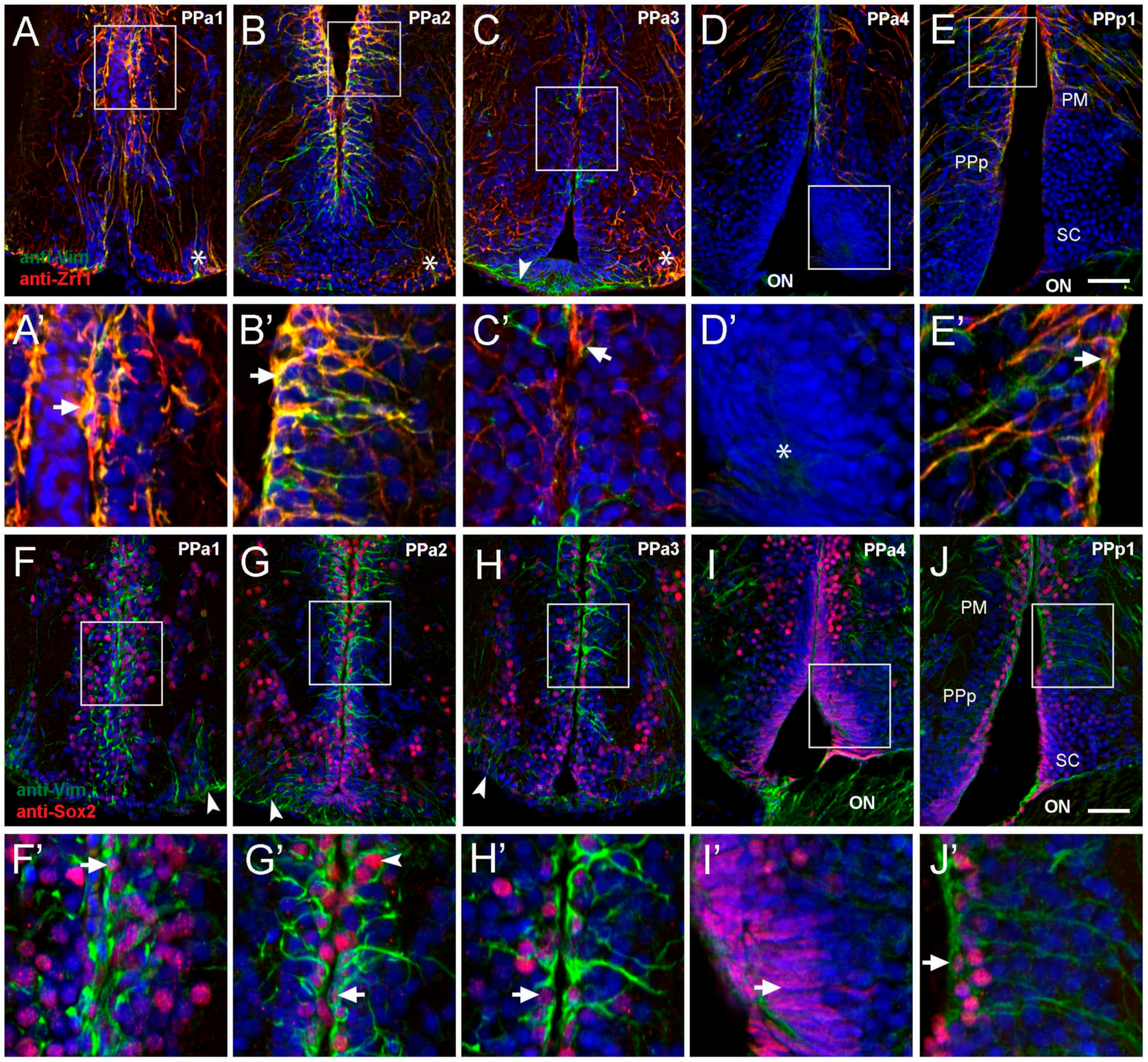
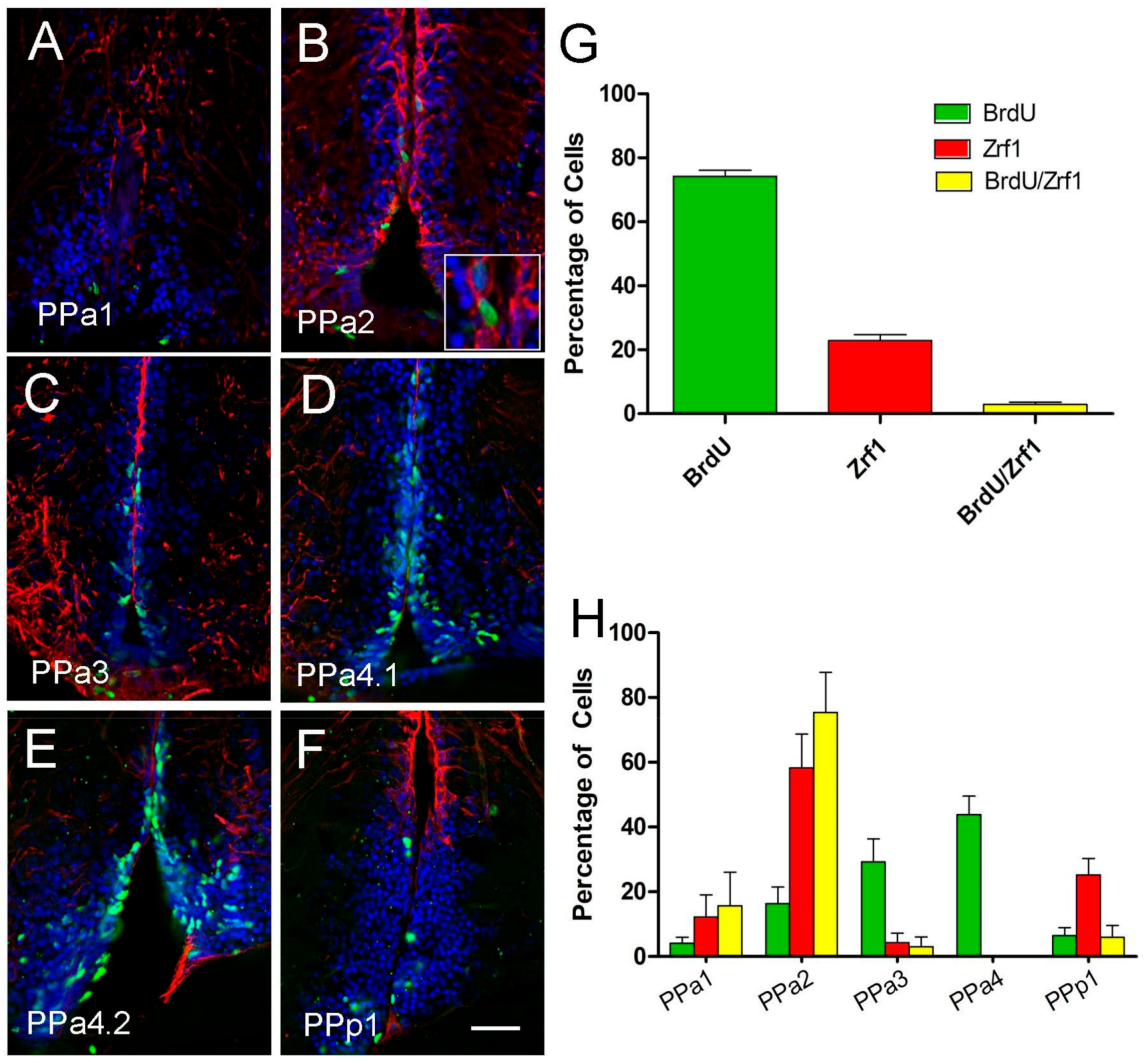
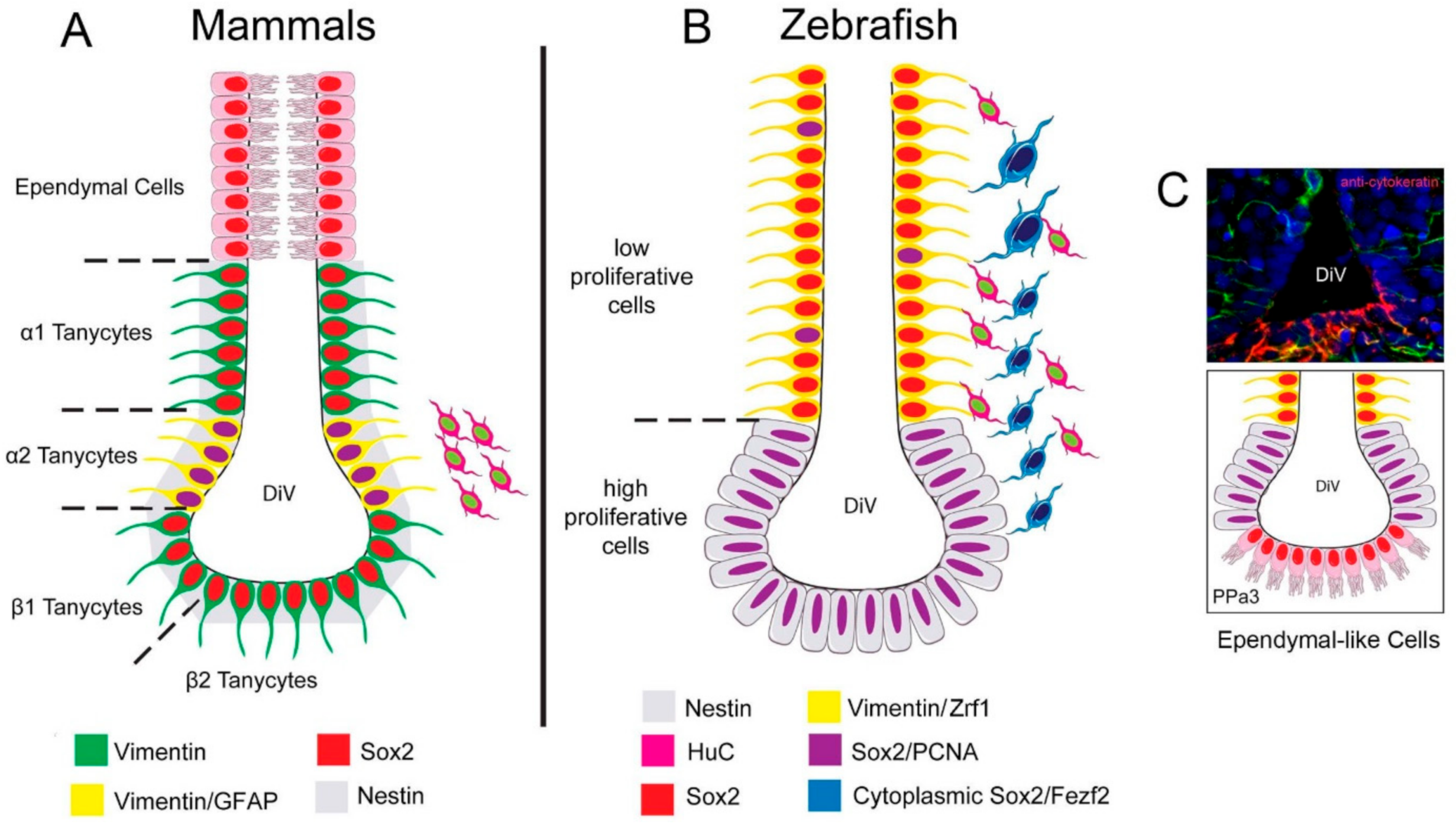
2.1. Hypothalamic Neural Progenitors Are Located in the POA
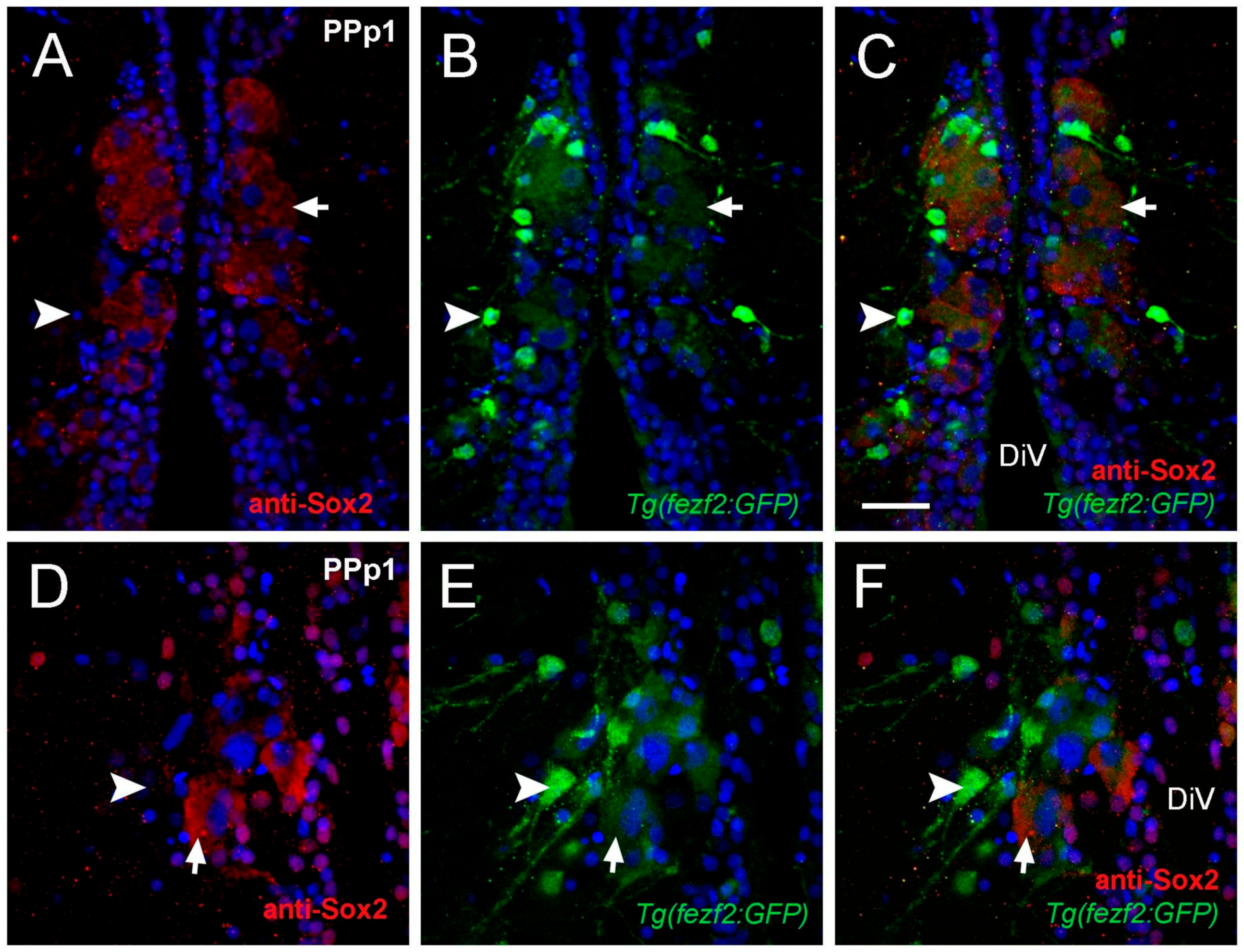
2.2. Cytoplasmic Sox2 Cells Express Fezf2:GFP in Ventral Region of the POA
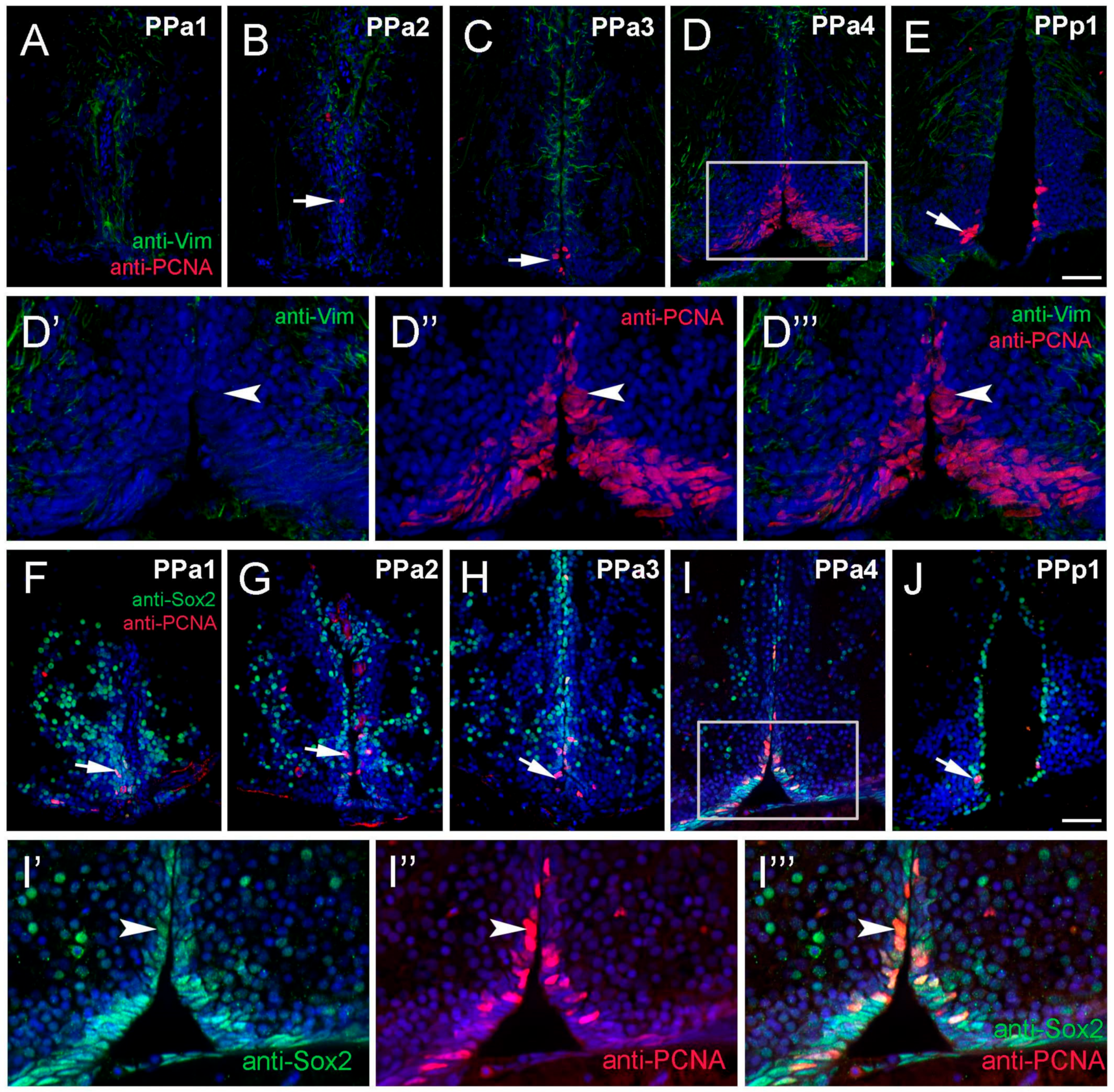
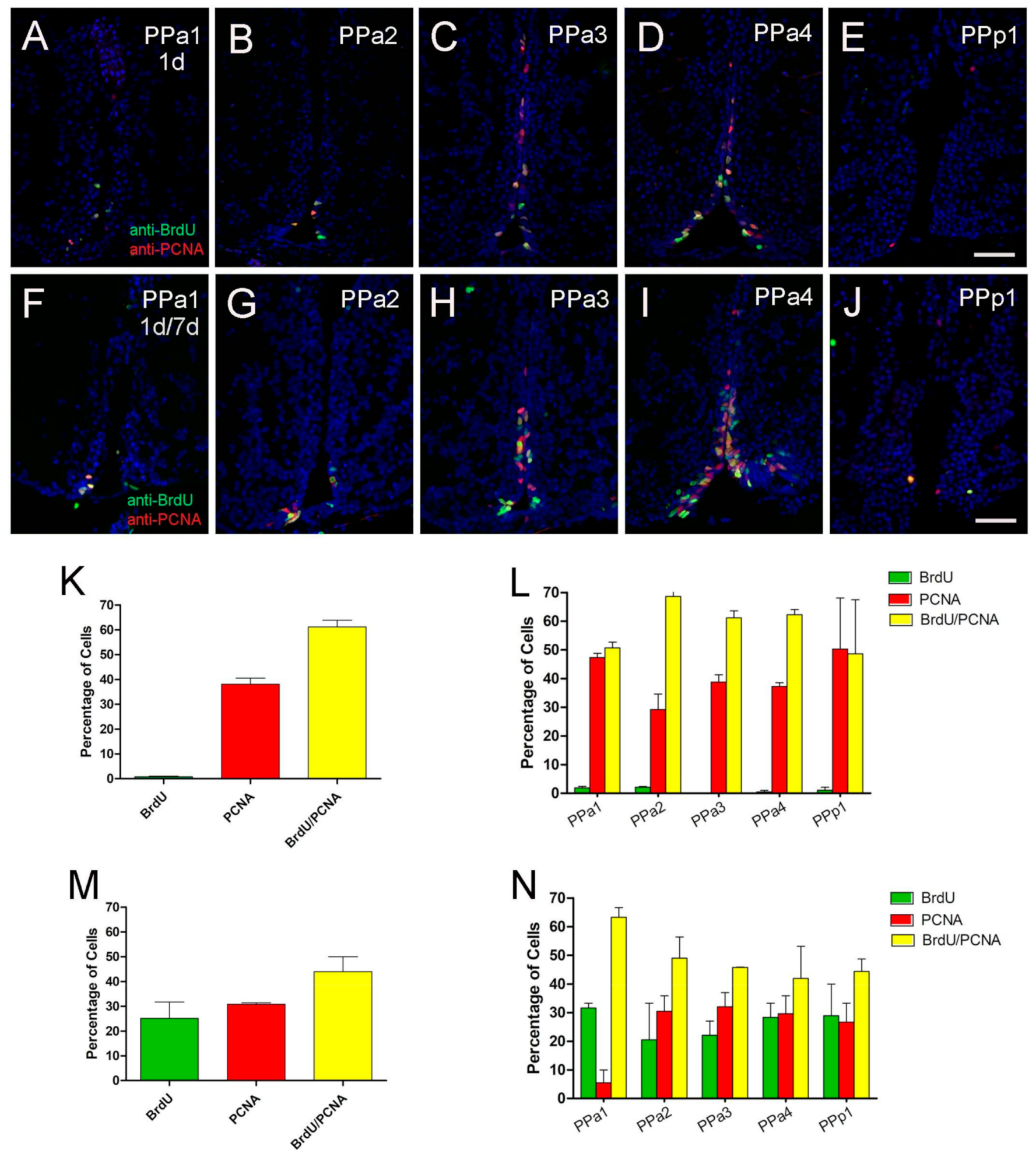
2.3. Proliferative Cells in the POA
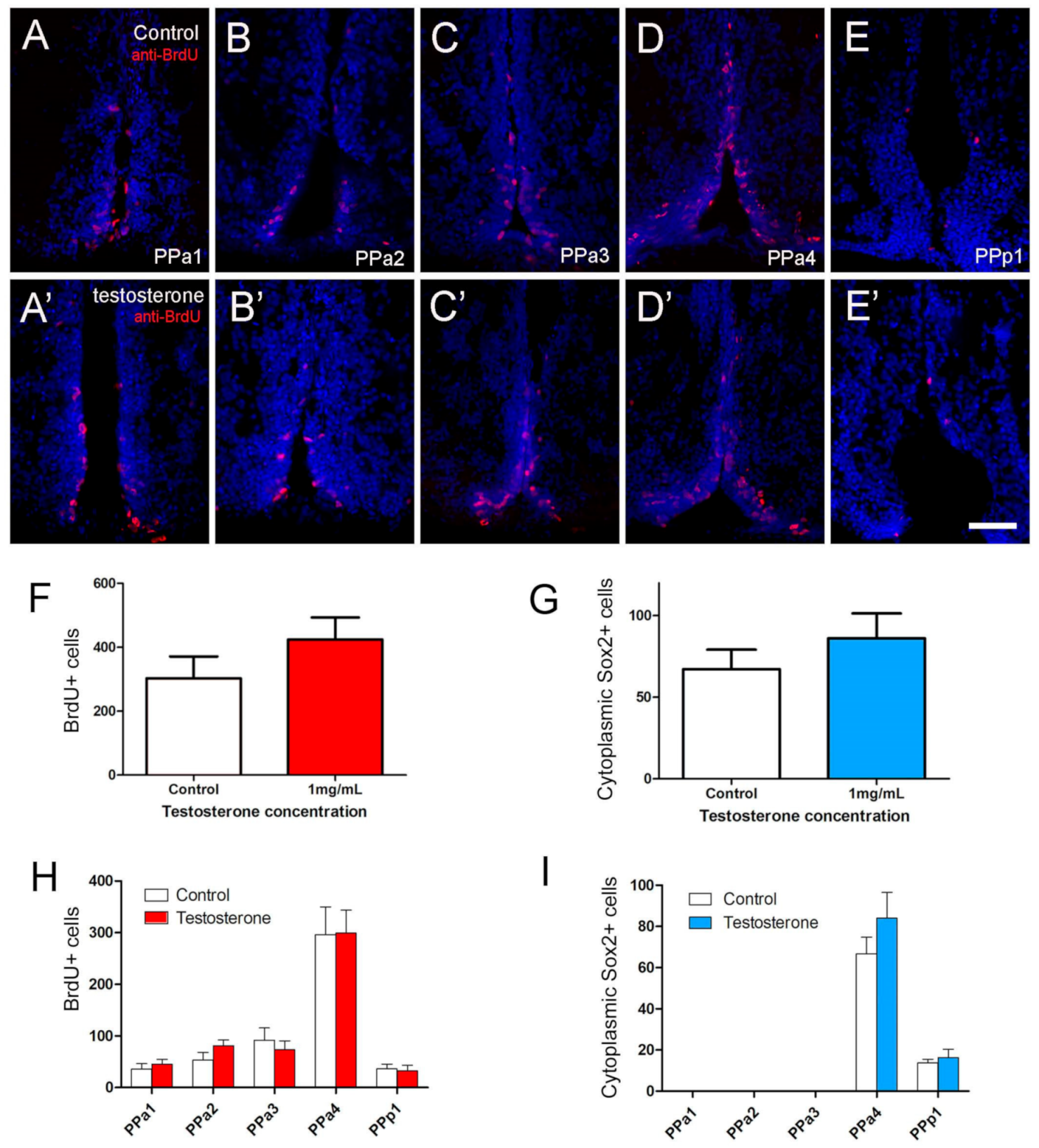
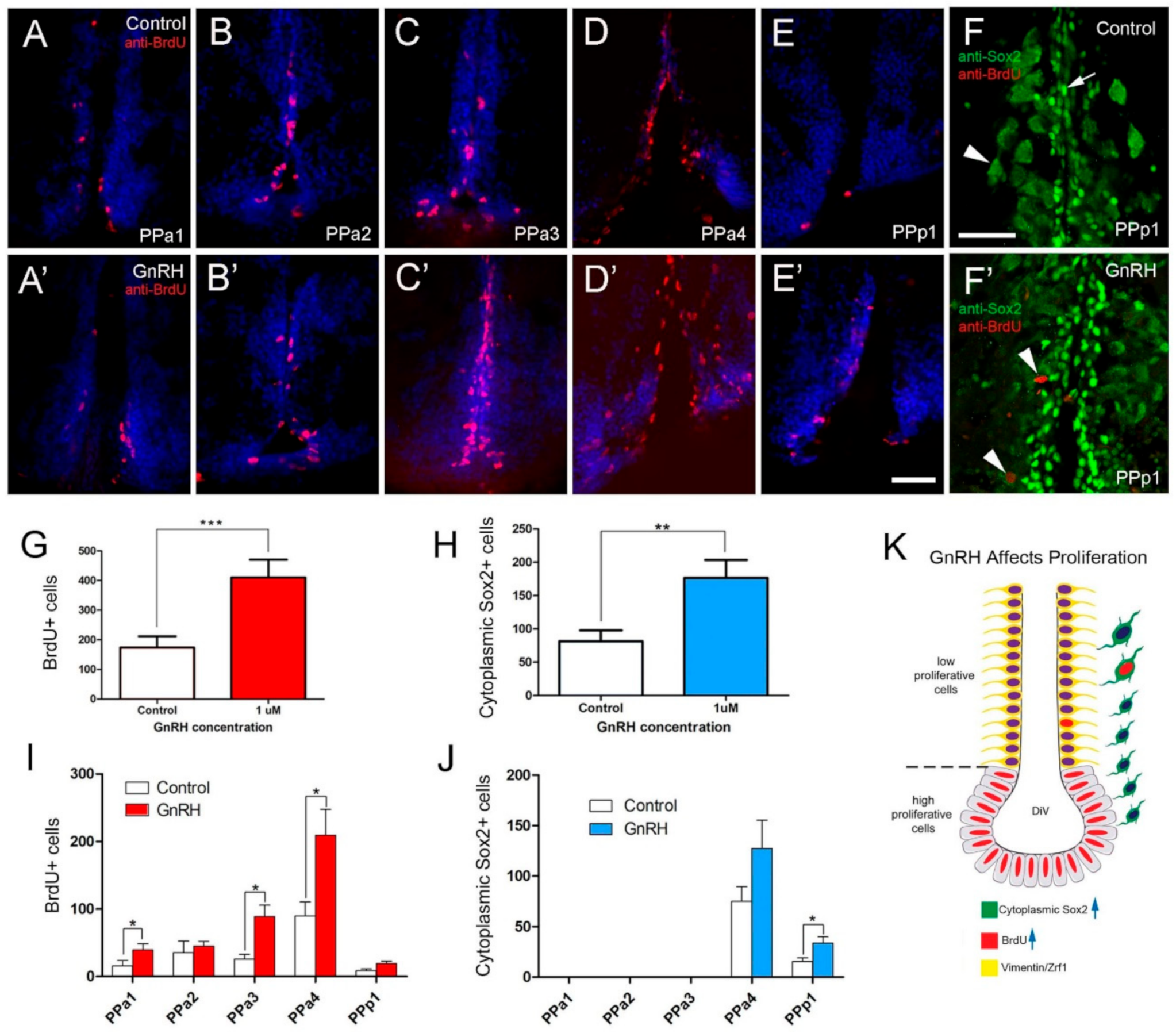
2.4. GnRH and Testosterone Treatments Cause Neurogenesis in the POA
3. Discussion
3.1. Tanycytes as Adult Neural Progenitors
3.2. Cytoplasmic Sox2 Cells
3.3. Hormone Treatment and Neurogenesis
3.4. GnRH, Neurogenesis, and Longevity
4. Materials and Methods
4.1. Animals
4.2. Histochemistry
4.2.1. Trichromic Stain in Paraffin Sections
4.2.2. Immunocytochemistry in Paraffin Sections
4.2.3. Immunocytochemistry in Cryosections
4.2.4. Antibodies
4.2.5. Hormone Injection and BrdU Incubation
4.3. Microscopy
4.4. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cameron, H.A.; McKay, R.D. Adult neurogenesis produces a large pool of new granule cells in the dentate gyrus. J. Comp. Neurol. 2001, 435, 406–417. [Google Scholar] [CrossRef] [PubMed]
- Kempermann, G.; Wiskott, L.; Gage, F.H. Functional significance of adult neurogenesis. Curr. Opin. Neurobiol. 2004, 14, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Bond, A.M.; Ming, G.-L.; Song, H. Ontogeny of adult neural stem cells in the mammalian brain. Curr. Top. Dev. Biol. 2021, 142, 67–98. [Google Scholar] [CrossRef] [PubMed]
- 4Doetsch, F.; Alvarez-Buylla, A. Network of tangential pathways for neuronal migration in adult mammalian brain. Proc. Natl. Acad. Sci. USA 1996, 93, 14895–14900. [Google Scholar] [CrossRef] [Green Version]
- Ghaddar, B.; Lübke, L.; Couret, D.; Rastegar, S.; Diotel, N. Cellular Mechanisms Participating in Brain Repair of AdultZebrafish and Mammals after Injury. Cells 2021, 10, 391. [Google Scholar] [CrossRef]
- Losurdo, M.; Grilli, M. Extracellular Vesicles, Influential Players of Intercellular Communication within Adult Neurogenic Niches. Int. J. Mol. Sci. 2020, 21, 8819. [Google Scholar] [CrossRef]
- Grandel, H.; Kaslin, J.; Ganz, J.; Wenzel, I.; Brand, M. Neural stem cells and neurogenesis in the adult zebrafish brain: Origin, proliferation dynamics, migration and cell fate. Dev. Biol. 2006, 295, 263–277. [Google Scholar] [CrossRef] [Green Version]
- Kizil, C.; Kaslin, J.; Kroehne, V.; Brand, M. Adult neurogenesis and brain regeneration in zebrafish. Dev. Neurobiol. 2012, 72, 429–461. [Google Scholar] [CrossRef]
- Than-Trong, E.; Bally-Cuif, L. Radial glia and neural progenitors in the adult zebrafish central nervous system. Glia 2015, 63, 1406–1428. [Google Scholar] [CrossRef]
- Burbridge, S.; Stewart, I.; Placzek, M. Development of the Neuroendocrine Hypothalamus. Compr. Physiol. 2016, 6, 623–643. [Google Scholar] [CrossRef]
- Nagpal, J.; Herget, U.; Choi, M.K.; Ryu, S. Anatomy, development, and plasticity of the neurosecretory hypothalamus in zebrafish. Cell Tissue Res. 2018, 375, 5–22. [Google Scholar] [CrossRef]
- Prevot, V.; Dehouck, B.; Sharif, A.; Ciofi, P.; Giacobini, P.; Clasadonte, J. The Versatile Tanycyte: A Hypothalamic Integrator of Reproduction and Energy Metabolism. Endocr. Rev. 2018, 39, 333–368. [Google Scholar] [CrossRef]
- Rodríguez, E.M.; Blázquez, J.L.; Pastor, F.E.; Peláez, B.; Peña, P.; Peruzzo, B.; Amat, P. Hypothalamic Tanycytes: A Key Component of Brain–Endocrine Interaction. Int. Rev. Cytol. 2005, 247, 89–164. [Google Scholar] [CrossRef]
- Yoo, S.; Blackshaw, S. Regulation and function of neurogenesis in the adult mammalian hypothalamus. Prog. Neurobiol. 2018, 170, 53–66. [Google Scholar] [CrossRef]
- Bolborea, M.; Dale, N. Hypothalamic tanycytes: Potential roles in the control of feeding and energy balance. Trends Neurosci. 2013, 36, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.A.; Bedont, J.L.; Pak, T.; Wang, H.; Song, J.; Miranda-Angulo, A.; Takiar, V.; Charubhumi, V.; Balordi, F.; Takebayashi, H.; et al. Tanycytes of the hypothalamic median eminence form a diet-responsive neurogenic niche. Nat. Neurosci. 2012, 15, 700–702. [Google Scholar] [CrossRef] [Green Version]
- Robins, S.; Stewart, I.; McNay, D.; Taylor, V.; Giachino, C.; Goetz, M.; Ninkovic, J.; Briancon, N.; Maratos-Flier, E.; Flier, J.; et al. α-Tanycytes of the adult hypothalamic third ventricle include distinct populations of FGF-responsive neural progenitors. Nat. Commun. 2013, 4, 2049. [Google Scholar] [CrossRef] [Green Version]
- Kostin, A.; Alam, M.A.; McGinty, D.; Alam, M.N. Adult hypothalamic neurogenesis and sleep-wake dysfunction in aging. Sleep 2021, 44, zsaa173. [Google Scholar] [CrossRef]
- Lindvall, O.; Kokaia, Z. Stem cells for the treatment of neurological disorders. Nature 2006, 441, 1094–1096. [Google Scholar] [CrossRef]
- Gore, A.C. GnRH: Tha Master Molecule of Reproduction; Kluwer Academic Publishers: Norwell, MA, USA, 2002; p. 227. [Google Scholar]
- Terasawa, E.; Fernandez, D.L. Neurobiological mechanisms of the onset of puberty in primates. Endocr. Rev. 2001, 22, 111–151. [Google Scholar] [CrossRef]
- Dodé, C.; Hardelin, J.P. Kallmann syndrome. Eur. J. Hum. Genet. 2009, 17, 139–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raivio, T.; Falardeau, J.; Dwyer, A.; Quinton, R.; Hayes, F.J.; Hughes, V.A.; Cole, L.W.; Pearce, S.; Lee, H.; Boepple, P.; et al. Reversal of Idiopathic Hypogonadotropic Hypogonadism. N. Engl. J. Med. 2007, 357, 863–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sidhoum, V.F.; Chan, Y.-M.; Lippincott, M.F.; Balasubramanian, R.; Quinton, R.; Plummer, L.; Dwyer, A.; Pitteloud, N.; Hayes, F.J.; Hall, J.; et al. Reversal and Relapse of Hypogonadotropic Hypogonadism: Resilience and Fragility of the Reproductive Neuroendocrine System. J. Clin. Endocrinol. Metab. 2014, 99, 861–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamson, D.K.; Wainwright, S.R.; Taylor, J.R.; Jones, B.A.; Watson, N.V.; Galea, L.A.M. Androgens Increase Survival of Adult-Born Neurons in the Dentate Gyrus by an Androgen Receptor-Dependent Mechanism in Male Rats. Endocrinology 2013, 154, 3294–3304. [Google Scholar] [CrossRef] [Green Version]
- Farinetti, A.; Tomasi, S.; Foglio, B.; Ferraris, A.; Ponti, G.; Gotti, S.; Peretto, P.; Panzica, G.; Farinetti, A.; Tomasi, S.; et al. Testosterone and estradiol differentially affect cell proliferation in the subventricular zone of young adult gonadectomized male and female rats. Neuroscience 2015, 286, 162–170. [Google Scholar] [CrossRef] [Green Version]
- Purkayastha, S.; Cai, D. Disruption of neurogenesis by hypothalamic inflammation in obesity or aging. Rev. Endocr. Metab. Disord. 2013, 14, 351–356. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Li, J.; Purkayastha, S.; Tang, Y.; Zhang, H.; Yin, Y.; Li, B.; Liu, G.; Cai, D. Hypothalamic programming of systemic ageing involving IKK-β, NF-κB and GnRH. Nat. Cell Biol. 2013, 497, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Campos, C.C.; Letelier, J.; Ceriani, R.; Whitlock, K.E. Zebrafish adult-derived hypothalamic neurospheres generate gonadotropin-releasing hormone (GnRH) neurons. Biol. Open 2015, 4, 1077–1086. [Google Scholar] [CrossRef] [Green Version]
- Whitlock, K.E. The loss of scents: Do defects in olfactory sensory neuron development underlie human disease? Birth Defects Res. Part C Embryo Today Rev. 2015, 105, 114–125. [Google Scholar] [CrossRef]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Petree, C.; Requena, T.; Varshney, P.; Varshney, G.K. Expanding the CRISPR Toolbox in Zebrafish for Studying Development and Disease. Front. Cell Dev. Biol. 2019, 7, 13. [Google Scholar] [CrossRef] [Green Version]
- Rupp, B.; Reichert, H.; Wullimann, M.F. The zebrafish brain: A neuroanatomical comparison with the goldfish. Brain Struct. Funct. 1996, 194, 187–203. [Google Scholar] [CrossRef] [PubMed]
- Furube, E.; Ishii, H.; Nambu, Y.; Kurganov, E.; Nagaoka, S.; Morita, M.; Miyata, S. Neural stem cell phenotype of tanycyte-like ependymal cells in the circumventricular organs and central canal of adult mouse brain. Sci. Rep. 2020, 10, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercurio, S.; Serra, L.; Nicolis, S.K. More than just Stem Cells: Functional Roles of the Transcription Factor Sox2 in Differentiated Glia and Neurons. Int. J. Mol. Sci. 2019, 20, 4540. [Google Scholar] [CrossRef] [Green Version]
- Blechman, J.; Borodovsky, N.; Eisenberg, M.; Nabel-Rosen, H.; Grimm, J.; Levkowitz, G. Specification of hypothalamic neurons by dual regulation of the homeodomain protein Orthopedia. Development 2007, 134, 4417–4426. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, A.M.; Beddows, E.; Filippi, A.; Driever, W. Orthopedia Transcription Factor otpa and otpb Paralogous Genes Function during Dopaminergic and Neuroendocrine Cell Specification in Larval Zebrafish. PLoS ONE 2013, 8, e75002. [Google Scholar] [CrossRef]
- Berberoglu, M.A.; Dong, Z.; Mueller, T.; Guo, S. fezf2 expression delineates cells with proliferative potential and expressing markers of neural stem cells in the adult zebrafish brain. Gene Expr. Patterns 2009, 9, 411–422. [Google Scholar] [CrossRef] [Green Version]
- Garcia, A.D.R.; Doan, N.B.; Imura, T.; Bush, T.G.; Sofroniew, M.V. GFAP-expressing progenitors are the principal source of constitutive neurogenesis in adult mouse forebrain. Nat. Neurosci. 2004, 7, 1233–1241. [Google Scholar] [CrossRef]
- Connolly, K.M.; Bogdanffy, M.S. Evaluation of proliferating cell nuclear antigen (PCNA) as an endogenous marker of cell proliferation in rat liver: A dual-stain comparison with 5-bromo-2’-deoxyuridine. J. Histochem. Cytochem. 1993, 41, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Rojczyk-Gołębiewska, E.; Pałasz, A.; Wiaderkiewicz, R. Hypothalamic subependymal niche: A novel site of the adult neurogenesis. Cell. Mol. Neurobiol. 2014, 34, 631–642. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Ma, L.; Cristofanilli, M.; Hart, R.; Hao, A.; Schachner, M. Transcription factor Sox11b is involved in spinal cord regeneration in adult zebrafish. Neuroscience 2011, 172, 329–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paquette, C.E.; Kent, M.L.; Peterson, T.S.; Wang, R.; Dashwood, R.H.; Löhr, C.V. Immunohistochemical characterization of intestinal neoplasia in zebrafish Danio rerio indicates epithelial origin. Dis. Aquat. Org. 2015, 116, 191–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa, J.S.; Sanchez-Gonzalez, R.; Di Giaimo, R.; Baumgart, E.V.; Theis, F.J.; Götz, M.; Ninkovic, M.G.A.J. Live imaging of adult neural stem cell behavior in the intact and injured zebrafish brain. Science 2015, 348, 789–793. [Google Scholar] [CrossRef] [PubMed]
- Ganz, J.; Kaslin, J.; Hochmann, S.; Freudenreich, D.; Brand, M. Heterogeneity and Fgf dependence of adult neural progenitors in the zebrafish telencephalon. Glia 2010, 58, 1345–1363. [Google Scholar] [CrossRef]
- Matsuzaki, K.; Katakura, M.; Sugimoto, N.; Hara, T.; Hashimoto, M.; Shido, O. Neural progenitor cell proliferation in the hypothalamus is involved in acquired heat tolerance in long-term heat-acclimated rats. PLoS ONE 2017, 12, e0178787. [Google Scholar] [CrossRef] [Green Version]
- März, M.; Chapouton, P.; Diotel, N.; Vaillant, C.; Hesl, B.; Takamiya, M.; Lam, C.S.; Kah, O.; Bally-Cuif, L.; Strähle, U. Heterogeneity in progenitor cell subtypes in the ventricular zone of the zebrafish adult telencephalon. Glia 2010, 58, 870–888. [Google Scholar] [CrossRef]
- Batailler, M.; Droguerre, M.; Baroncini, M.; Fontaine, C.; Prevot, V.; Migaud, M. DCX-expressing cells in the vicinity of the hypothalamic neurogenic niche: A comparative study between mouse, sheep, and human tissues. J. Comp. Neurol. 2014, 522, 1966–1985. [Google Scholar] [CrossRef]
- Lindsey, B.W.; Darabie, A.; Tropepe, V. The cellular composition of neurogenic periventricular zones in the adult zebrafish forebrain. J. Comp. Neurol. 2012, 520, 2275–2316. [Google Scholar] [CrossRef]
- Kaslin, J.; Kroehne, V.; Ganz, J.; Hans, S.; Brand, M. Distinct roles of neuroepithelial-like and radial glia-like progenitor cells in cerebellar regeneration. Development 2017, 144, 1462–1471. [Google Scholar] [CrossRef] [Green Version]
- Van Schaijik, B.; Davis, P.F.; Wickremesekera, A.C.; Tan, S.T.; Itinteang, T. Subcellular localisation of the stem cell markers OCT4, SOX2, NANOG, KLF4 and c-MYC in cancer: A review. J. Clin. Pathol. 2018, 71, 88–91. [Google Scholar] [CrossRef]
- Schaefer, T.; Lengerke, C. SOX2 protein biochemistry in stemness, reprogramming, and cancer: The PI3K/AKT/SOX2 axis and beyond. Oncogene 2020, 39, 278–292. [Google Scholar] [CrossRef] [Green Version]
- Almeida, O.; Oliveira, R.F. Social Status and Arginine Vasotocin Neuronal Phenotypes in a Cichlid Fish. Brainbehav. Evol. 2015, 85, 203–213. [Google Scholar] [CrossRef]
- Eaton, J.L.; Holmqvist, B.; Glasgow, E. Ontogeny of vasotocin-expressing cells in zebrafish: Selective requirement for the transcriptional regulatorsorthopediaandsingle-minded 1in the preoptic area. Dev. Dyn. 2008, 237, 995–1005. [Google Scholar] [CrossRef]
- Alvarez-Bolado, G. Development of neuroendocrine neurons in the mammalian hypothalamus. Cell Tissue Res. 2019, 375, 23–39. [Google Scholar] [CrossRef]
- Levkowitz, G.; Zeller, J.; Sirotkin, H.I.; French, R.; Schilbach, S.; Hashimoto, H.; Hibi, M.; Talbot, W.S.; Rosenthal, A. Zinc finger protein too few controls the development of monoaminergic neurons. Nat. Neurosci. 2002, 6, 28–33. [Google Scholar] [CrossRef]
- Caldwell, L.J.; Davies, N.O.; Cavone, L.; Mysiak, K.S.; Semenova, S.A.; Panula, P.; Armstrong, J.D.; Becker, C.; Becker, T. Regeneration of Dopaminergic Neurons in Adult Zebrafish Depends on Immune System Activation and Differs for Distinct Populations. J. Neurosci. 2019, 39, 4694–4713. [Google Scholar] [CrossRef] [Green Version]
- Millimaki, B.B.; Sweet, E.M.; Riley, B.B. Sox2 is required for maintenance and regeneration, but not initial development, of hair cells in the zebrafish inner ear. Dev. Biol. 2010, 338, 262–269. [Google Scholar] [CrossRef] [Green Version]
- Narita, Y.; Tsutiya, A.; Nakano, Y.; Ashitomi, M.; Sato, K.; Hosono, K.; Kaneko, T.; Chen, R.-D.; Lee, J.-R.; Tseng, Y.-C.; et al. Androgen induced cellular proliferation, neurogenesis, and generation of GnRH3 neurons in the brain of mature female Mozambique tilapia. Sci. Rep. 2018, 8, 16855. [Google Scholar] [CrossRef]
- Fowler, C.D.; Freeman, M.E.; Wang, Z. Newly proliferated cells in the adult male amygdala are affected by gonadal steroid hormones. J. Neurobiol. 2003, 57, 257–269. [Google Scholar] [CrossRef]
- Antzoulatos, E.; Magorien, J.E.; Wood, R.I. Cell proliferation and survival in the mating circuit of adult male hamsters: Effects of testosterone and sexual behavior. Horm. Behav. 2008, 54, 735–740. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Chadwick, W.; Park, S.-S.; Zhou, Y.; Silver, N.; Martin, B.; Maudsley, S. Gonadotropin-Releasing Hormone Receptor System: Modulatory Role in Aging and Neurodegeneration. CNS Neurol. Disord. Drug Targets 2010, 9, 651–660. [Google Scholar] [CrossRef] [Green Version]
- Barardo, D.G.; Newby, D.; Thornton, D.; Ghafourian, T.; De Magalhães, J.P.; Freitas, A.A. Machine learning for predicting lifespan-extending chemical compounds. Aging 2017, 9, 1721–1737. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Choe, H.K. Role of hypothalamus in aging and its underlying cellular mechanisms. Mech. Ageing Dev. 2019, 177, 74–79. [Google Scholar] [CrossRef]
- Mariën, V.; Zandecki, C.; Seuntjens, E.; Ayana, R.; Arckens, L. Modeling Neuroregeneration and Neurorepair in an Aging Context: The Power of a Teleost Model. Front. Cell Dev. Biol. 2021, 9, 619197. [Google Scholar]
- Krylov, V.V.; Izvekov, E.I.; Pavlova, V.V.; Pankova, N.A.; Osipova, E.A. Circadian rhythms in zebrafish (Danio rerio) behaviour and the sources of their variability. Biol. Rev. Camb. Philos. Soc. 2020, 96, 785–797. [Google Scholar] [CrossRef]
- Edelmann, K.; Glashauser, L.; Sprungala, S.; Hesl, B.; Fritschle, M.; Ninkovic, J.; Godinho, L.; Chapouton, P. Increased radial glia quiescence, decreased reactivation upon injury and unaltered neuroblast behavior underlie decreased neurogenesis in the aging zebrafish telencephalon. J. Comp. Neurol. 2013, 521, 3099–3115. [Google Scholar] [CrossRef]
- Xi, Y.; Yu, M.; Godoy, R.; Hatch, G.; Poitras, L.; Ekker, M. Transgenic zebrafish expressing green fluorescent protein in dopaminergic neurons of the ventral diencephalon. Dev. Dyn. 2011, 240, 2539–2547. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.J.; Fossum, S.L.; Fimbel, S.M.; Montgomery, J.E.; Hyde, D.R. The inhibitor of phagocytosis, O-phospho-l-serine, suppresses Müller glia proliferation and cone cell regeneration in the light-damaged zebrafish retina. Exp. Eye Res. 2010, 91, 601–612. [Google Scholar] [CrossRef] [Green Version]
- Dirian, L.; Galant, S.; Coolen, M.; Chen, W.; Bedu, S.; Houart, C.; Bally-Cuif, L.; Foucher, I. Spatial Regionalization and Heterochrony in the Formation of Adult Pallial Neural Stem Cells. Dev. Cell 2014, 30, 123–136. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Fogarty, B.; LaForge, B.; Aziz, S.; Pham, T.; Lai, L.; Bai, S. Delivery of Small Interfering RNA to Inhibit Vascular Endothelial Growth Factor in Zebrafish Using Natural Brain Endothelia Cell-Secreted Exosome Nanovesicles for the Treatment of Brain Cancer. AAPS J. 2016, 19, 475–486. [Google Scholar] [CrossRef]
- Kinkel, M.D.; Eames, S.C.; Philipson, L.H.; Prince, V.E. Intraperitoneal Injection into Adult Zebrafish. J. Vis. Exp. 2010, e2126. [Google Scholar] [CrossRef] [PubMed]
- Wullimann, M.F.; Mueller, T. Expression of Zash-1a in the postembryonic zebrafish brain allows comparison to mouse Mash1 domains. Brain Res. Gene Expr. Patterns 2002, 1, 187–192. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigen | Host | Dilution | Manufacturer | Cat. No | Immunogen Organism | Reference Use in Zebrafish |
|---|---|---|---|---|---|---|
| BrdU | Rabbit | 1:500 | Invitrogen | PA5-32256 | BrdU conjugated to KLH | Not identified |
| BrdU | Rat | 1:500 | Abcam | Ab6326 | Not available | [67] |
| GFP | Mouse | 1:500 | Invitrogen | A-11120 | Aequorea victoria | [68] |
| PCNA | Mouse | 1:100 | Sigma-Aldrich | P8825 | Rat | [69] |
| Sox2 | Mouse | 1:200 | Abcam | ab137385 | Human | Not identified |
| Sox2 | Rabbit | 1:500 | Abcam | Ab97959 | Human | [70] |
| Vimentin | Chicken | 1:500 | Millipore | AB5733 | Human | [29] |
| Zrf1 | Mouse | 1:500 | Abcam | ab154474 | Zebrafish | [71] |
| Cytokeratin(clones AE1/AE3) | Mouse | 1:500 | Dako | M3515 | Human | [43] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ceriani, R.; Whitlock, K.E. Gonadotropin Releasing Hormone (GnRH) Triggers Neurogenesis in the Hypothalamus of Adult Zebrafish. Int. J. Mol. Sci. 2021, 22, 5926. https://doi.org/10.3390/ijms22115926
Ceriani R, Whitlock KE. Gonadotropin Releasing Hormone (GnRH) Triggers Neurogenesis in the Hypothalamus of Adult Zebrafish. International Journal of Molecular Sciences. 2021; 22(11):5926. https://doi.org/10.3390/ijms22115926
Chicago/Turabian StyleCeriani, Ricardo, and Kathleen E. Whitlock. 2021. "Gonadotropin Releasing Hormone (GnRH) Triggers Neurogenesis in the Hypothalamus of Adult Zebrafish" International Journal of Molecular Sciences 22, no. 11: 5926. https://doi.org/10.3390/ijms22115926
APA StyleCeriani, R., & Whitlock, K. E. (2021). Gonadotropin Releasing Hormone (GnRH) Triggers Neurogenesis in the Hypothalamus of Adult Zebrafish. International Journal of Molecular Sciences, 22(11), 5926. https://doi.org/10.3390/ijms22115926