
Extracellular Vesicles: Versatile Nanomediators, Potential Biomarkers and Therapeutic Agents in Atherosclerosis and COVID-19-Related Thrombosis


Abstract
:1. Introduction
2. Extracellular Vesicles: Our Current Understanding
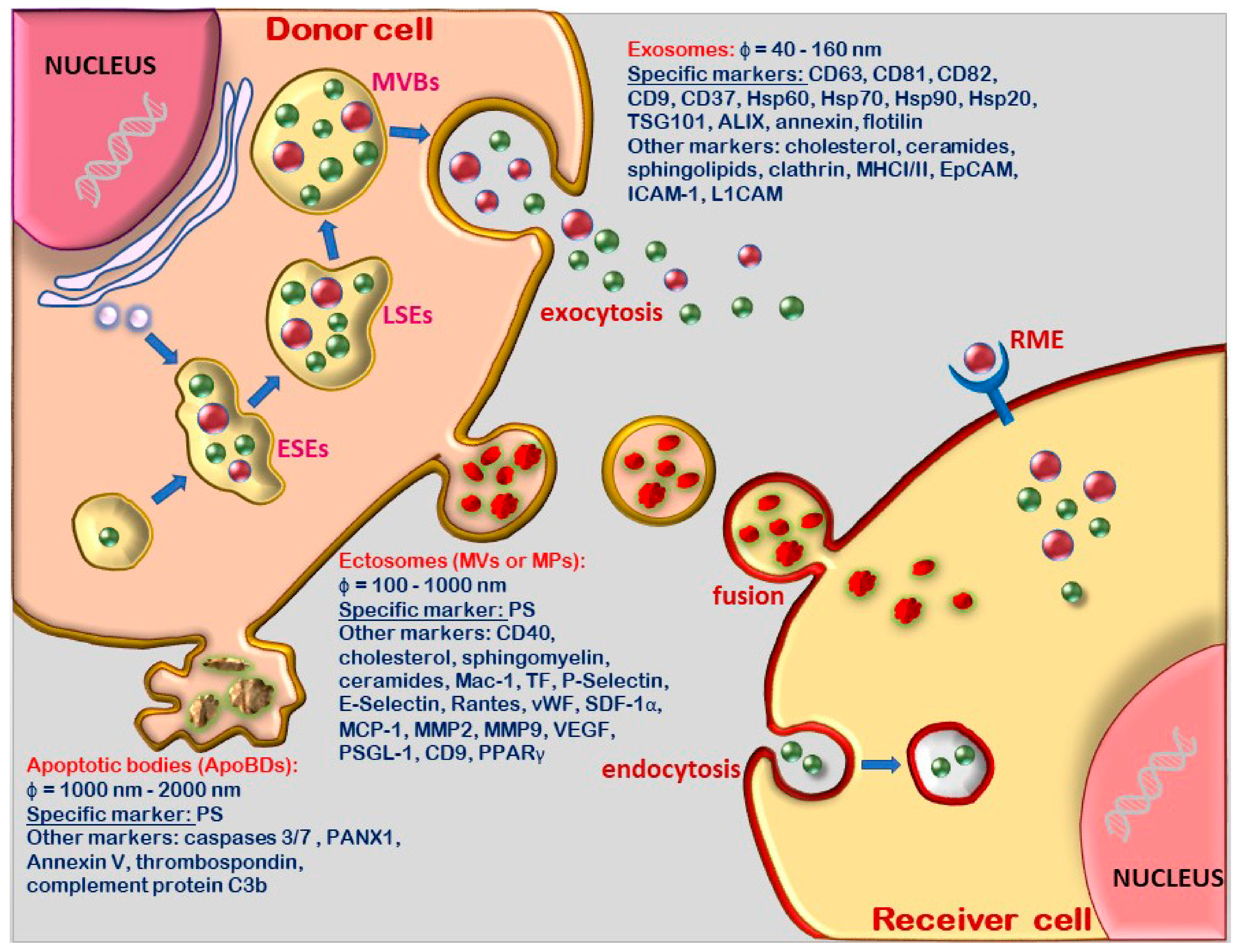
2.1. Terminology and Biogenesis Pathways of the Distinct Extracellular Vesicle Population
2.2. Molecular Content and Biological Functions of Extracellular Vesicles
3. Pathophysiology of Atherosclerosis
3.1. Consecutive Stages, the Cells and Molecules Involved in the Formation of Atheroma
3.2. Immune Cell Activation and Inflammatory Process in Atherogenesis
4. Extracellular Vesicles Have the Potential to Predict, Monitor and Act as Therapeutic Agents in Atherosclerosis
4.1. Extracellular Vesicles of Various Cell Origin Are Implicated in All Stages of Atheroma Formation
4.2. Extracellular Vesicles as Nanomediators in Atherosclerosis
4.3. Extracellular Vesicles as Biomarkers in Atherosclerosis
4.4. Extracellular Vesicles Deliver Molecules to Target Cells Acting as Therapeutic Agents
5. COVID-19-Associated Thrombosis and Extracellular Vesicles
5.1. Thrombosis Is a Common Occurrence in Corona-Virus Disease 19 (COVID-19)
5.2. Extracellular Vesicles as Biomarkers in COVID-19-Associated Thrombosis
5.3. Extracellular Vesicles as Therapeutic Agents in COVID-19-Related Thrombosis
6. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ABCA1 | ATP binding cassette A1 |
| ABCG-1 | ATP-binding cassette sub-family G member 1 |
| ACE2 | angiotensin-converting enzyme 2 receptor |
| Akt | protein kinase B |
| ALIX | apoptosis-linked gene 2-interacting protein X |
| ApoBDs | apoptotic bodies |
| ARDS | acute respiratory distress syndrome |
| ARF6 | ADP-ribosylation factor 6 |
| CAPN2 | calpain 2 |
| CCL5 | chemokine (C-C motif) ligand 5 |
| CHIP | clonal hematopoiesis of indeterminate potential |
| COPB2 | coat complex subunit beta 2 |
| COVID-19 | coronavirus disease-2019 |
| CRP | C-reactive protein |
| DLL4 | Notch ligand Delta-like 4 |
| E | envelope proteins |
| ECM1 | extracellular matrix protein 1 |
| ECs | endothelial cells |
| EGFRs | epidermal growth factor receptors |
| EMVs | endothelial-derived microvesicles |
| eNOS | endothelial nitric oxide synthase |
| EpCAM | epithelial cell adhesion molecule |
| EPCs | endothelial progenitor cells |
| ERK | extracellular regulated protein kinase |
| ESCRT | endosomal sorting complexes required for transport |
| ESEs | early sorting endosomes |
| EVs | extracellular vesicles |
| Exo-MSC | bone marrow mesenchymal stem cells |
| FDP | fibrin degradation product |
| FGG | fibrinogen gamma chain |
| Hsp | heat-shock proteins |
| ICAM-1 | intercellular adhesion molecule-1 |
| IFNγ | interferon gamma |
| IL-1 | interleukin-1 |
| ILVs | intraluminal vesicles |
| INR | international normalized ratio |
| L1CAM | L1 cell adhesion molecule |
| LC-MS | liquid chromatograph mass spectrometers |
| LFA-1 | lymphocyte function-associated antigen 1 |
| LMVs | leukocyte-derived microvesicles |
| LSEs | late sorting endosomes |
| M | membrane proteins |
| M2 Exo | M2 macrophage-derived exosomes |
| Mac-1 | macrophage integrin-1 |
| MCP-1 | monocyte chemoattractant protein-1 |
| M-CSF | monocyte colony stimulating factor |
| miRNA | microRNA |
| MMP2 | matrix metallopeptidase2 |
| MPs | microparticles |
| MSCs | mesenchymal stem cells |
| MVBs | multivesicular bodies |
| MVs | microvesicles |
| N | nucleocapsid proteins |
| ncRNA | non-coding RNAs |
| NLRP3 | nod-like receptor protein 3 |
| oxLDL | oxidized low-density lipoproteins |
| PDGF | platelet-derived growth factor |
| PDGF | platelet-derived growth factor |
| PMVs | platelet-derived microvesicles |
| PPAR-γ | proliferator-activated receptor-γ |
| PRKCB | protein kinase C beta |
| PS | phosphatidylserine |
| PSGL-1 | P-Selectin glycoprotein ligand 1 |
| PT | prothrombin time |
| RhoC | ras homolog family member C, |
| S | spike proteins |
| SARS-CoV-2 | severe acute respiratory syndrome coronavirus-2 |
| SDF-1α | stromal cell-derived factor 1 α |
| SMCs | smooth muscle cells |
| TF | tissue factor |
| TGF-β | transforming growth factor β |
| TMPRSS2 | transmembrane protease, serine 2 |
| TNFα | tumour necrosis factor α |
| Th-1 | T helper 1 cells |
| T-reg | regulatory T cells |
| TSG101 | tumor susceptibility gene 101 |
| VCAM-1 | vascular-cell adhesion molecule-1 |
| VEGF | vascular endothelial growth factor |
| vWF | von Willebrand factor |
References
- Boulanger, C.M.; Scoazec, A.; Ebrahimian, T.; Henry, P.; Mathieu, E.; Tedgui, A.; Mallat, Z. Circulating Microparticles From Patients with Myocardial Infarction Cause Endothelial Dysfunction. Circulation 2001, 104, 2649–2652. [Google Scholar] [CrossRef]
- Boulanger, C.M.; Amabile, N.; Tedgui, A. Circulating microparticles: A potential prognostic marker for atherosclerotic vascular disease. Hypertension 2006, 48, 180–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tokarz, A.; Szuścik, I.; Kuśnierz-Cabala, B.; Kapusta, M.; Konkolewska, M.; Żurakowski, A.; Georgescu, A.; Stępień, E. Extracellular vesicles participate in the transport of cytokines and angiogenic factors in diabetic patients with ocular complications. Folia Med. Cracov. 2015, 55, 35–48. [Google Scholar]
- Georgescu, A.; Alexandru, N.; Popov, D.; Amuzescu, M.; Andrei, E.; Zamfir, C.; Maniu, H.; Badila, A. Chronic venous insufficiency is associated with elevated level of circulating microparticles. J. Thromb. Haemost. 2009, 7, 1566–1575. [Google Scholar] [CrossRef] [PubMed]
- Georgescu, A.; Alexandru, N.; Andrei, E.; Titorencu, I.; Dragan, E.; Tarziu, C.; Ghiorghe, S.; Badila, E.; Bartos, D.; Popov, D. Circulating microparticles and endothelial progenitor cells in atherosclerosis: Pharmacological effects of irbesartan. J. Thromb. Haemost. 2012, 10, 680–691. [Google Scholar] [CrossRef] [PubMed]
- Koenen, R.R.; Aikawa, E. Editorial: Extracellular Vesicle-Mediated Processes in Cardiovascular Diseases. Front. Cardiovasc. Med. 2018, 5, 133. [Google Scholar] [CrossRef]
- Rajendran, L.; Bali, J.; Barr, M.M.; Court, F.A.; Krämer-Albers, E.-M.; Picou, F.; Raposo, G.; Van Der Vos, K.E.; Van Niel, G.; Wang, J.; et al. Emerging Roles of Extracellular Vesicles in the Nervous System. J. Neurosci. 2014, 34, 15482–15489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endzeliņš, E.; Berger, A.; Melne, V.; Bajo-Santos, C.; Soboļevska, K.; Ābols, A.; Rodriguez, M.; Šantare, D.; Rudņickiha, A.; Lietuvietis, V.; et al. Detection of circulating miRNAs: Comparative analysis of extracellular vesicle-incorporated miRNAs and cell-free miRNAs in whole plasma of prostate cancer patients. BMC Cancer 2017, 17, 730. [Google Scholar] [CrossRef]
- Popēna, I.; Ābols, A.; Saulīte, L.; Pleiko, K.; Zandberga, E.; Jēkabsons, K.; Endzeliņš, E.; Llorente, A.; Linē, A.; Riekstiņa, U. Effect of colorectal cancer-derived extracellular vesicles on the immunophenotype and cytokine secretion profile of monocytes and macrophages. Cell Commun. Signal. 2018, 16, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Simionescu, N.; Sima, A.; Dobrian, A.; Tirziu, D. Pathobiochemical Changes of the Arterial Wall at the Inception of Atherosclerosis. In Glomerulonephritis; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 1993; Volume 87, pp. 1–45. [Google Scholar]
- Schwartz, C.J.; Valente, A.J.; Sprague, E.A. A modern view of atherogenesis. Am. J. Cardiol. 1993, 71, B9–B14. [Google Scholar] [CrossRef]
- Manduteanu, I.; Simionescu, M. Inflammation in atherosclerosis: A cause or a result of vascular disorders? J. Cell. Mol. Med. 2012, 16, 1978–1990. [Google Scholar] [CrossRef] [Green Version]
- Arraud, N.; Linares, R.; Tan, S.; Gounou, C.; Pasquet, J.-M.; Mornet, S.; Brisson, A.R. Extracellular vesicles from blood plasma: Determination of their morphology, size, phenotype and concentration. J. Thromb. Haemost. 2014, 12, 614–627. [Google Scholar] [CrossRef] [PubMed]
- Zonneveld, M.; Brisson, A.R.; van Herwijnen, M.; Tan, S.; Van De Lest, C.H.A.; Redegeld, F.A.; Garssen, J.; Wauben, M.; Nolte-’t Hoen, E.N.M. Recovery of extracellular vesicles from human breast milk is influenced by sample collection and vesicle isolation procedures. J. Extracell. Vesicles 2014, 3, 24215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Höög, J.L.; Lötvall, J. Diversity of extracellular vesicles in human ejaculates revealed by cryo-electron microscopy. J. Extracell. Vesicles 2015, 4, 28680. [Google Scholar] [CrossRef] [Green Version]
- Iwai, K.; Minamisawa, T.; Suga, K.; Yajima, Y.; Shiba, K. Isolation of human salivary extracellular vesicles by iodixanol density gradient ultracentrifugation and their characterizations. J. Extracell. Vesicles 2016, 5, 30829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merchant, M.L.; Rood, I.M.; Deegens, J.K.J.; Klein, J.B. Isolation and characterization of urinary extracellular vesicles: Implications for biomarker discovery. Nat. Rev. Nephrol. 2017, 13, 731–749. [Google Scholar] [CrossRef] [Green Version]
- Battistelli, M.; Falcieri, E. Apoptotic Bodies: Particular Extracellular Vesicles Involved in Intercellular Communication. Biology 2020, 9, 21. [Google Scholar] [CrossRef] [Green Version]
- Trams, E.G.; Lauter, C.J.; Salem, J.N.; Heine, U. Exfoliation of membrane ecto-enzymes in the form of micro-vesicles. Biochim. Biophys. Acta (BBA) Biomembr. 1981, 645, 63–70. [Google Scholar] [CrossRef]
- Pan, B.T.; Teng, K.; Wu, C.; Adam, M.; Johnstone, R.M. Electron microscopic evidence for externalization of the transferrin receptor in vesicular form in sheep reticulocytes. J. Cell Biol. 1985, 101, 942–948. [Google Scholar] [CrossRef] [Green Version]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef]
- Ciardiello, C.; Cavallini, L.; Spinelli, C.; Yang, J.; Reis-Sobreiro, M.; De Candia, P.; Minciacchi, V.R.; Di Vizio, D. Focus on Extracellular Vesicles: New Frontiers of Cell-to-Cell Communication in Cancer. Int. J. Mol. Sci. 2016, 17, 175. [Google Scholar] [CrossRef] [Green Version]
- Jansen, F.; Li, Q.; Pfeifer, A.; Werner, N. Endothelial- and Immune Cell-Derived Extracellular Vesicles in the Regulation of Cardiovascular Health and Disease. JACC Basic Transl. Sci. 2017, 2, 790–807. [Google Scholar] [CrossRef] [PubMed]
- Bebelman, M.; Smit, M.J.; Pegtel, D.M.; Baglio, S.R. Biogenesis and function of extracellular vesicles in cancer. Pharmacol. Ther. 2018, 188, 1–11. [Google Scholar] [CrossRef]
- Mathieu, M.; Martin-Jaular, L.; Lavieu, G.; Théry, C. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat. Cell Biol. 2019, 21, 9–17. [Google Scholar] [CrossRef]
- Takahashi, A.; Okada, R.; Nagao, K.; Kawamata, Y.; Hanyu, A.; Yoshimoto, S.; Takasugi, M.; Watanabe, S.; Kanemaki, M.T.; Obuse, C.; et al. Exosomes maintain cellular homeostasis by excreting harmful DNA from cells. Nat. Commun. 2017, 8, 15287. [Google Scholar] [CrossRef] [Green Version]
- Wolf, P. The Nature and Significance of Platelet Products in Human Plasma. Br. J. Haematol. 1967, 13, 269–288. [Google Scholar] [CrossRef]
- Van Der Pol, E.; Böing, A.N.; Harrison, P.; Sturk, A.; Nieuwland, R. Classification, Functions, and Clinical Relevance of Extracellular Vesicles. Pharmacol. Rev. 2012, 64, 676–705. [Google Scholar] [CrossRef] [Green Version]
- Stępień, E.Ł.; Durak-Kozica, M.; Kamińska, A.; Targosz-Korecka, M.; Libera, M.; Tylko, G.; Opalińska, A.; Kapusta, M.; Solnica, B.; Georgescu, A.; et al. Circulating ectosomes: Determination of angiogenic microRNAs in type 2 diabetes. Theranostics 2018, 8, 3874–3890. [Google Scholar] [CrossRef]
- Cocucci, E.; Racchetti, G.; Meldolesi, J. Shedding microvesicles: Artefacts no more. Trends Cell Biol. 2009, 19, 43–51. [Google Scholar] [CrossRef]
- Dolis, D.; Moreau, C.; Zachowski, A.; Devaux, P.F. Aminophospholipid translocase and proteins involved in transmembrane phospholipid traffic. Biophys. Chem. 1997, 68, 221–231. [Google Scholar] [CrossRef]
- Yu, S.-L.; Gan, X.-G.; Huang, J.-M.; Cao, Y.; Wang, Y.-Q.; Pan, S.-H.; Ma, L.-Y.; Teng, Y.-Q.; An, R.-H. Oxalate Impairs Aminophospholipid Translocase Activity in Renal Epithelial Cells Via Oxidative Stress: Implications for Calcium Oxalate Urolithiasis. J. Urol. 2011, 186, 1114–1120. [Google Scholar] [CrossRef]
- Hugel, B.; Martínez, M.C.; Kunzelmann, C.; Freyssinet, J.-M. Membrane Microparticles: Two Sides of the Coin. Physiology 2005, 20, 22–27. [Google Scholar] [CrossRef]
- Crawford, S.; Diamond, D.; Brustolon, L.; Penarreta, R. Effect of Increased Extracellular Ca++ on Microvesicle Production and Tumor Spheroid Formation. Cancer Microenviron. 2010, 4, 93–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muralidharan-Chari, V.; Clancy, J.; Plou, C.; Romao, M.; Chavrier, P.; Raposo, G.; D’Souza-Schorey, C. ARF6-Regulated Shedding of Tumor Cell-Derived Plasma Membrane Microvesicles. Curr. Biol. 2009, 19, 1875–1885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Souza-Schorey, C.; Chavrier, P. ARF proteins: Roles in membrane traffic and beyond. Nat. Rev. Mol. Cell Biol. 2006, 7, 347–358. [Google Scholar] [CrossRef]
- Birge, R.B.; Boeltz, S.; Kumar, S.; Carlson, J.; Wanderley, J.; Calianese, D.; Barcinski, M.; Brekken, R.A.; Huang, X.; Hutchins, J.T.; et al. Phosphatidylserine is a global immunosuppressive signal in efferocytosis, infectious disease, and cancer. Cell Death Differ. 2016, 23, 962–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samanta, S.; Rajasingh, S.; Drosos, N.; Zhou, Z.; Dawn, B.; Rajasingh, J. Exosomes: New molecular targets of diseases. Acta Pharmacol. Sin. 2018, 39, 501–513. [Google Scholar] [CrossRef]
- Valadi, H.; Ekstrom, K.; Bossios, A.; Sjostrand, M.; Lee, J.J.; Lotvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Cocucci, E.; Meldolesi, J. Ectosomes and exosomes: Shedding the confusion between extracellular vesicles. Trends Cell Biol. 2015, 25, 364–372. [Google Scholar] [CrossRef]
- Simons, M.; Raposo, G. Exosomes—Vesicular carriers for intercellular communication. Curr. Opin. Cell Biol. 2009, 21, 575–581. [Google Scholar] [CrossRef]
- Guescini, M.; Guidolin, D.; Vallorani, L.; Casadei, L.; Gioacchini, A.M.; Tibollo, P.; Battistelli, M.; Falcieri, E.; Battistin, L.; Agnati, L.F.; et al. C2C12 myoblasts release micro-vesicles containing mtDNA and proteins involved in signal transduction. Exp. Cell Res. 2010, 316, 1977–1984. [Google Scholar] [CrossRef]
- Ludwig, A.-K.; Giebel, B. Exosomes: Small vesicles participating in intercellular communication. Int. J. Biochem. Cell Biol. 2012, 44, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Ermini, L.; Ausman, J.; Melland-Smith, M.; Yeganeh, B.; Rolfo, A.; Litvack, M.L.; Todros, T.; Letarte, M.; Post, M.; Caniggia, I. A Single Sphingomyelin Species Promotes Exosomal Release of Endoglin into the Maternal Circulation in Preeclampsia. Sci. Rep. 2017, 7, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Marcos, S.; Albiñana, V.; Recio-Poveda, L.; Tarazona, B.; Verde-González, M.; Ojeda-Fernández, L.; Botella, L.-M. SARS-CoV-2 Infection in Hereditary Hemorrhagic Telangiectasia Patients Suggests Less Clinical Impact Than in the General Population. J. Clin. Med. 2021, 10, 1884. [Google Scholar] [CrossRef]
- Meldolesi, J. Exosomes and Ectosomes in Intercellular Communication. Curr. Biol. 2018, 28, R435–R444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keshtkar, S.; Azarpira, N.; Ghahremani, M.H. Mesenchymal stem cell-derived extracellular vesicles: Novel frontiers in regenerative medicine. Stem Cell Res. Ther. 2018, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Su, H.; Li, J.; Lyon, C.; Tang, W.; Wan, M.; Hu, T.Y. Clinical applications of exosome membrane proteins. Precis. Clin. Med. 2020, 3, 54–66. [Google Scholar] [CrossRef]
- Choi, D.-S.; Kim, D.-K.; Kim, Y.-K.; Gho, Y.S. Proteomics of extracellular vesicles: Exosomes and ectosomes. Mass Spectrom. Rev. 2015, 34, 474–490. [Google Scholar] [CrossRef] [PubMed]
- Ravanat, C.; Archipoff, G.; Beretz, A.; Freund, G.; Cazenave, J.P.; Freyssinet, J.M. Use of annexin-V to demonstrate the role of phosphatidylserine exposure in the maintenance of haemostatic balance by endothelial cells. Biochem. J. 1992, 282, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Chahed, S.; Leroyer, A.S.; Benzerroug, M.; Gaucher, D.; Georgescu, A.; Picaud, S.; Silvestre, J.-S.; Gaudric, A.; Tedgui, A.; Massin, P.; et al. Increased Vitreous Shedding of Microparticles in Proliferative Diabetic Retinopathy Stimulates Endothelial Proliferation. Diabetes 2009, 59, 694–701. [Google Scholar] [CrossRef] [Green Version]
- Otzen, D.E.; Blans, K.; Wang, H.; Gilbert, G.E.; Rasmussen, J.T. Lactadherin binds to phosphatidylserine-containing vesicles in a two-step mechanism sensitive to vesicle size and composition. Biochim. Biophys. Acta (BBA) Biomembr. 2012, 1818, 1019–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belik, D.; Tsang, H.; Wharton, J.; Howard, L.; Bernabeu, C.; Wojciak-Stothard, B. Endothelium-derived microparticles from chronically thromboembolic pulmonary hypertensive patients facilitate endothelial angiogenesis. J. Biomed. Sci. 2016, 23, 4. [Google Scholar] [CrossRef] [Green Version]
- Norling, L.V.; Dalli, J. Microparticles are novel effectors of immunity. Curr. Opin. Pharmacol. 2013, 13, 570–575. [Google Scholar] [CrossRef]
- Lannan, K.L.; Esahler, J.; Ekim, N.; Spinelli, S.L.; Maggirwar, S.B.; Egarraud, O.; Cognasse, F.; Eblumberg, N.; Phipps, R.P. Breaking the Mold: Transcription Factors in the Anucleate Platelet and Platelet-Derived Microparticles. Front. Immunol. 2015, 6, 48. [Google Scholar] [CrossRef] [Green Version]
- Georgescu, A.; Alexandru, N.; Nemecz, M.; Titorencu, I.; Popov, D. Irbesartan administration therapeutically influences circulating endothelial progenitor cell and microparticle mobilization by involvement of pro-inflammatory cytokines. Eur. J. Pharmacol. 2013, 711, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Alexandru, N.; Andrei, E.; Dragan, E.; Georgescu, A. Interaction of platelets with endothelial progenitor cells in the experimental atherosclerosis: Role of transplanted endothelial progenitor cells and platelet microparticles. Biol. Cell 2015, 107, 189–204. [Google Scholar] [CrossRef]
- Chong, S.Y.; Lee, C.K.; Huang, C.; Ou, Y.H.; Charles, C.J.; Richards, A.M.; Neupane, Y.R.; Pavon, M.V.; Zharkova, O.; Pastorin, G.; et al. Extracellular Vesicles in Cardiovascular Diseases: Alternative Biomarker Sources, Therapeutic Agents, and Drug Delivery Carriers. Int. J. Mol. Sci. 2019, 20, 3272. [Google Scholar] [CrossRef] [Green Version]
- Lv, Y.; Tan, J.; Miao, Y.; Zhang, Q. The role of microvesicles and its active molecules in regulating cellular biology. J. Cell. Mol. Med. 2019, 23, 7894–7904. [Google Scholar] [CrossRef]
- Alexandru, N.; Costa, A.; Constantin, A.; Cochior, D.; Georgescu, A. Microparticles: From Biogenesis to Biomarkers and Diagnostic Tools in Cardiovascular Disease. Curr. Stem Cell Res. Ther. 2016, 12, 89–102. [Google Scholar] [CrossRef]
- Simeone, P.; Bologna, G.; Lanuti, P.; Pierdomenico, L.; Guagnano, M.T.; Pieragostino, D.; Del Boccio, P.; Vergara, D.; Marchisio, M.; Miscia, S.; et al. Extracellular Vesicles as Signaling Mediators and Disease Biomarkers across Biological Barriers. Int. J. Mol. Sci. 2020, 21, 2514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simionescu, N.; Vasile, E.; Lupu, F.; Popescu, G.; Simionescu, M. Prelesional events in atherogenesis. accumulation of extracellular cholesterol-rich liposomes in the arterial intima and cardiac valves of the hyper-lipidemic rabbit. Am. J. Pathol. 1986, 123, 109–125. [Google Scholar]
- Simionescu, M.; Sima, A.V. Morphology of atherosclerotic lesions. In Inflammation and Atherosclerosis; Wick, G., Grundtman, C., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 19–37. [Google Scholar]
- Lupu, F.; Danaricu, I.; Simionescu, N. Development of intracellular lipid deposits in the lipid-laden cells of atherosclerotic lesions: A cytochemical and ultrastructural study. Atherosclerosis 1987, 67, 127–142. [Google Scholar] [CrossRef]
- Tîrziu, D.; Dobrian, A.; Tasca, C.; Simionescu, M.; Simionescu, N. Intimal thickenings of human aorta contain modified reassembled lipoproteins. Atherosclerosis 1995, 112, 101–114. [Google Scholar] [CrossRef]
- Rocha, V.Z.; Libby, P. Obesity, inflammation, and atherosclerosis. Nat. Rev. Cardiol. 2009, 6, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Getz, G.S. T Cells in Atherosclerosis in Ldlr−/− and Apoe−/− Mice. J. Immunol. Sci. 2018, 2, 69–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rognoni, A.; Cavallino, C.; Veia, A.; Bacchini, S.; Rosso, R.; Facchini, M.; Secco, G.G.; Lupi, A.; Nardi, F.; Rametta, F.; et al. Pathophysiology of Atherosclerotic Plaque Development. Cardiovasc. Hematol. Agents Med. Chem. 2015, 13, 10–13. [Google Scholar] [CrossRef] [PubMed]
- Georgescu, A.; Alexandru, N.; Andrei, E.; Dragan, E.; Cochior, D.; Dias, S. Effects of transplanted circulating endothelial progenitor cells and platelet microparticles in atherosclerosis development. Biol. Cell 2016, 108, 219–243. [Google Scholar] [CrossRef] [PubMed]
- Badimon, L.; Vilahur, G. Thrombosis formation on atherosclerotic lesions and plaque rupture. J. Intern. Med. 2014, 276, 618–632. [Google Scholar] [CrossRef]
- Linton, M.R.F.; Yancey, P.G.; Davies, S.S.; Jerome, W.G.; Linton, E.F.; Song, W.L.; Doran, A.C.; Vickers, K.C. The role of lipids and lipoproteins in atherosclerosis. In Endotext [Internet]; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., Dungan, K., Grossman, A., Hershman, J.M., Kaltsas, G., Koch, C., Kopp, P., et al., Eds.; MDText.com Inc.: South Dartmouth, MA, USA, 2000; Bookshelf ID: NBK343489. [Google Scholar]
- Mach, F. Inflammation Is a Crucial Feature of Atherosclerosis and a Potential Target to Reduce Cardiovascular Events. In Organotypic Models in Drug Development; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2005; pp. 697–722. [Google Scholar]
- Ross, R. Atherosclerosis—An Inflammatory Disease. N. Engl. J. Med. 1999, 340, 115–126. [Google Scholar] [CrossRef]
- Glass, C.K.; Witztum, J.L. Atherosclerosis. Cell 2001, 104, 503–516. [Google Scholar] [CrossRef] [Green Version]
- Lusis, A.J. Atherosclerosis. Nat. Cell Biol. 2000, 407, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Libby, P. Inflammation in Atherosclerosis. Arter. Thromb. Vasc. Biol. 2012, 32, 2045–2051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagareddy, P.; Smyth, S.S. Inflammation and thrombosis in cardiovascular disease. Curr. Opin. Hematol. 2013, 20, 457–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karasawa, T.; Takahashi, M. The crystal-induced activation of NLRP3 inflammasomes in atherosclerosis. Inflamm. Regen. 2017, 37, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tousoulis, D.; Kampoli, A.-M.; Papageorgiou, N.; Androulakis, E.; Antoniades, C.; Toutouzas, K.; Stefanadis, C. Pathophysiology of Atherosclerosis: The Role of Inflammation. Curr. Pharm. Des. 2011, 17, 4089–4110. [Google Scholar] [CrossRef]
- Riksen, N.P. Trained immunity and atherosclerotic cardiovascular disease. Curr. Opin. Lipidol. 2019, 30, 395–400. [Google Scholar] [CrossRef]
- Kumar, P.; Kopecky, S.L.; Yang, E.H.; Oren, O. Clonal Hematopoiesis of Indeterminate Potential and Cardiovascular Disease. Curr. Oncol. Rep. 2020, 22, 1–6. [Google Scholar] [CrossRef]
- Leroyer, A.S.; Isobe, H.; Lesèche, G.; Castier, Y.; Wassef, M.; Mallat, Z.; Binder, B.R.; Tedgui, A.; Boulanger, C.M. Cellular Origins and Thrombogenic Activity of Microparticles Isolated From Human Atherosclerotic Plaques. J. Am. Coll. Cardiol. 2007, 49, 772–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, W.; Tang, T.; Hou, Y.; Zeng, Q.; Wang, Y.; Fan, W.; Qu, S. Extracellular vesicles in atherosclerosis. Clin. Chim. Acta 2019, 495, 109–117. [Google Scholar] [CrossRef]
- Barry, O.P.; Praticò, D.; Savani, R.C.; Fitzgerald, G.A. Modulation of monocyte-endothelial cell interactions by platelet microparticles. J. Clin. Investig. 1998, 102, 136–144. [Google Scholar] [CrossRef]
- Peng, M.; Liu, X.; Xu, G. Extracellular Vesicles as Messengers in Atherosclerosis. J. Cardiovasc. Transl. Res. 2020, 13, 121–130. [Google Scholar] [CrossRef]
- Alexandru, N.; Popov, D.; Dragan, E.; Andrei, E.; Georgescu, A. Circulating Endothelial Progenitor Cell and Platelet Microparticle Impact on Platelet Activation in Hypertension Associated with Hypercholesterolemia. PLoS ONE 2013, 8, e52058. [Google Scholar] [CrossRef]
- Kapustin, A.N.; Schoppet, M.; Schurgers, L.J.; Reynolds, J.L.; McNair, R.; Heiss, A.; Jahnen-Dechent, W.; Hackeng, T.M.; Schlieper, G.; Harrison, P.; et al. Prothrombin Loading of Vascular Smooth Muscle Cell–Derived Exosomes Regulates Coagulation and Calcification. Arter. Thromb. Vasc. Biol. 2017, 37, e22–e32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.-T.; Yuan, H.-X.; Ou, Z.-J.; Ou, J.-S. Microparticles (Exosomes) and Atherosclerosis. Curr. Atheroscler. Rep. 2020, 22, 1–9. [Google Scholar] [CrossRef]
- Charla, E.; Mercer, J.; Maffia, P.; Nicklin, S. Extracellular vesicle signalling in atherosclerosis. Cell. Signal. 2020, 75, 109751. [Google Scholar] [CrossRef]
- Konkoth, A.; Saraswat, R.; Dubrou, C.; Sabatier, F.; Leroyer, A.S.; Lacroix, R.; Duchez, A.-C.; Dignat-George, F. Multifaceted role of extracellular vesicles in atherosclerosis. Atherosclerosis 2021, 319, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Niu, C.; Wang, X.; Zhao, M.; Cai, T.; Liu, P.; Li, J.; Willard, B.; Zu, L.; Zhou, E.; Li, Y.; et al. Macrophage Foam Cell–Derived Extracellular Vesicles Promote Vascular Smooth Muscle Cell Migration and Adhesion. J. Am. Hear. Assoc. 2016, 5, e004099. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Liu, B.; Wang, Z.; Wang, D.; Ni, H.; Zhang, L.; Wang, Y. Exosomes from nicotine-stimulated macrophages accelerate atherosclerosis through miR-21-3p/PTEN-mediated VSMC migration and proliferation. Theranostics 2019, 9, 6901–6919. [Google Scholar] [CrossRef] [PubMed]
- Todorova, D.; Simoncini, S.; Lacroix, R.; Sabatier, F.; Dignat-George, F. Extracellular Vesicles in Angiogenesis. Circ. Res. 2017, 120, 1658–1673. [Google Scholar] [CrossRef] [PubMed]
- Merino-González, C.; Zuñiga, F.A.; Escudero, C.A.; Eormazabanl, V.; Ereyes, C.; Nova-Lamperti, E.; Esalomon, C.; Eaguayo, C. Mesenchymal Stem Cell-Derived Extracellular Vesicles Promote Angiogenesis: Potencial Clinical Application. Front. Physiol. 2016, 7, 24. [Google Scholar] [CrossRef] [Green Version]
- Wadey, R.M.; Connolly, K.D.; Mathew, D.; Walters, G.; Rees, D.A.; James, P.E. Inflammatory adipocyte-derived extracellular vesicles promote leukocyte attachment to vascular endothelial cells. Atherosclerosis 2019, 283, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Feng, C.; Chen, Q.; Fan, M.; Guo, J.; Liu, Y.; Ji, T.; Zhu, J.; Zhao, X. Platelet-derived microparticles promote phagocytosis of oxidized low-density lipoprotein by macrophages, potentially enhancing foam cell formation. Ann. Transl. Med. 2019, 7, 477. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, J.; Zhang, S.; Yan, S.; Wang, Z.; Wang, C.; Zhang, X. MiR-30e and miR-92a are related to atherosclerosis by targeting ABCA1. Mol. Med. Rep. 2019, 19, 3298–3304. [Google Scholar] [CrossRef] [Green Version]
- Huber, H.J.; Holvoet, P. Exosomes. Curr. Opin. Lipidol. 2015, 26, 412–419. [Google Scholar] [CrossRef]
- Sheldon, H.; Heikamp, E.; Turley, H.; Dragovic, R.; Thomas, P.; Oon, C.E.; Leek, R.; Edelmann, M.; Kessler, B.; Sainson, R.C.A.; et al. New mechanism for Notch signaling to endothelium at a distance by Delta-like 4 incorporation into exosomes. Blood 2010, 116, 2385–2394. [Google Scholar] [CrossRef] [Green Version]
- Pironti, G.; Strachan, R.T.; Abraham, D.; Yu, S.M.-W.; Chen, M.; Chen, W.; Hanada, K.; Mao, L.; Watson, L.J.; Rockman, H.A. Circulating Exosomes Induced by Cardiac Pressure Overload Contain Functional Angiotensin II Type 1 Receptors. Circulation 2015, 131, 2120–2130. [Google Scholar] [CrossRef]
- Forlow, S.B.; McEver, R.P.; Nollert, M.U. Leukocyte-leukocyte interactions mediated by platelet microparticles under flow. Blood 2000, 95, 1317–1323. [Google Scholar] [CrossRef] [PubMed]
- Mause, S.F.; Von Hundelshausen, P.; Zernecke, A.; Koenen, R.R.; Weber, C. Platelet Microparticles. Arter. Thromb. Vasc. Biol. 2005, 25, 1512–1518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasina, E.M.; Cauwenberghs, S.; Staudt, M.; Feijge, M.A.; Weber, C.; Koenen, R.R.; Wm Heemskerk, J. Aging-and activation-induced platelet microparticles suppress apoptosis in monocytic cells and diferentially signal to proinflammatory mediator release. Am. J. Blood Res. 2013, 3, 107–123. [Google Scholar] [PubMed]
- Nagy, B.; Fulop, T.; Varga, Z.; Karányi, Z.; Magyar, M.T.; Oláh, L.; Papp, M.; Facskó, A.; Kappelmayer, J.; Paragh, G.; et al. Increased levels of platelet activation markers are positively associated with carotid wall thickness and other atherosclerotic risk factors in obese patients. Thromb. Haemost. 2011, 106, 683–692. [Google Scholar] [CrossRef] [Green Version]
- Gomez, I.; Ward, B.; Souilhol, C.; Recarti, C.; Ariaans, M.; Johnston, J.; Burnett, A.; Mahmoud, M.; Luong, L.A.; West, L.; et al. Neutrophil microvesicles drive atherosclerosis by delivering miR-155 to atheroprone endothelium. Nat. Commun. 2020, 11, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Hussein, M.N.A.; Nieuwland, R.; Hau, C.M.; Evers, L.M.; Meesters, E.W.; Sturk, A. Cell-derived microparticles contain caspase 3 in vitro and in vivo. J. Thromb. Haemost. 2005, 3, 888–896. [Google Scholar] [CrossRef]
- Nozaki, T.; Sugiyama, S.; Koga, H.; Sugamura, K.; Ohba, K.; Matsuzawa, Y.; Sumida, H.; Matsui, K.; Jinnouchi, H.; Ogawa, H. Significance of a Multiple Biomarkers Strategy Including Endothelial Dysfunction to Improve Risk Stratification for Cardiovascular Events in Patients at High Risk for Coronary Heart Disease. J. Am. Coll. Cardiol. 2009, 54, 601–608. [Google Scholar] [CrossRef] [Green Version]
- Bernal-Mizrachi, L.; Jy, W.; Jimenez, J.J.; Pastor, J.; Mauro, L.M.; Horstman, L.L.; de Marchena, E.; Ahn, Y.S. High levels of circulating endothelial microparticles in patients with acute coronary syndromes. Am. Hear. J. 2003, 145, 962–970. [Google Scholar] [CrossRef]
- Sinning, J.-M.; Losch, J.; Walenta, K.; Böhm, M.; Nickenig, G.; Werner, N. Circulating CD31+/Annexin V+ microparticles correlate with cardiovascular outcomes. Eur. Hear. J. 2010, 32, 2034–2041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stępień, E.; Stankiewicz, E.; Zalewski, J.; Godlewski, J.; Zmudka, K.; Wybrańska, I.; Steogonekpień, E.; Stankiewicz, E.; Zalewski, J.; Godlewski, J.; et al. Number of Microparticles Generated During Acute Myocardial Infarction and Stable Angina Correlates with Platelet Activation. Arch. Med Res. 2012, 43, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Angelillo-Scherrer, A. Leukocyte-Derived Microparticles in Vascular Homeostasis. Circ. Res. 2012, 110, 356–369. [Google Scholar] [CrossRef] [Green Version]
- Sarlon-Bartoli, G.; Bennis, Y.; Lacroix, R.; Piercecchi-Marti, M.D.; Bartoli, M.A.; Arnaud, L.; Mancini, J.; Boudes, A.; Sarlon, E.; Thevenin, B.; et al. Plasmatic Level of Leukocyte-Derived Microparticles Is Associated with Unstable Plaque in Asymptomatic Patients With High-Grade Carotid Stenosis. J. Am. Coll. Cardiol. 2013, 62, 1436–1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goetzl, E.J.; Schwartz, J.B.; Mustapic, M.; Lobach, I.V.; Daneman, R.; Abner, E.L.; Jicha, G.A. Altered cargo proteins of human plasma endothelial cell–derived exosomes in atherosclerotic cerebrovascular disease. FASEB J. 2017, 31, 3689–3694. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.; Yuan, S.; Li, S.; Li, L.; Liu, M.; Wan, S. The Exosome-Derived Biomarker in Atherosclerosis and Its Clinical Application. J. Cardiovasc. Transl. Res. 2019, 12, 68–74. [Google Scholar] [CrossRef]
- Yao, Z.-Y.; Chen, W.-B.; Shao, S.-S.; Ma, S.-Z.; Yang, C.-B.; Li, M.-Z.; Zhao, J.; Gao, L. Role of exosome-associated microRNA in diagnostic and therapeutic applications to metabolic disorders. J. Zhejiang Univ. Sci. B 2018, 19, 183–198. [Google Scholar] [CrossRef]
- Ramachandran, S.; Lowenthal, A.; Ritner, C.; Lowenthal, S.; Bernstein, H.S. Plasma microvesicle analysis identifies microRNA 129-5p as a biomarker of heart failure in univentricular heart disease. PLoS ONE 2017, 12, e0183624. [Google Scholar] [CrossRef] [PubMed]
- Jansen, F.; Yang, X.; Proebsting, S.; Hoelscher, M.; Przybilla, D.; Baumann, K.; Schmitz, T.; Dolf, A.; Endl, E.; Franklin, B.S.; et al. MicroRNA Expression in Circulating Microvesicles Predicts Cardiovascular Events in Patients with Coronary Artery Disease. J. Am. Hear. Assoc. 2014, 3, e001249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexandru, N.; Constantin, A.; Nemecz, M.; Comariţa, I.K.; Vîlcu, A.; Procopciuc, A.; Tanko, G.; Georgescu, A. Hypertension Associated with Hyperlipidemia Induced Different MicroRNA Expression Profiles in Plasma, Platelets, and Platelet-Derived Microvesicles; Effects of Endothelial Progenitor Cell Therapy. Front. Med. 2019, 6, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardiner, C.; Di Vizio, D.; Sahoo, S.; Théry, C.; Witwer, K.; Wauben, M.; Hill, A.F. Techniques used for the isolation and characterization of extracellular vesicles: Results of a worldwide survey. J. Extracell. Vesicles 2016, 5, 32945. [Google Scholar] [CrossRef]
- Wu, M.; Ouyang, Y.; Wang, Z.; Zhang, R.; Huang, P.-H.; Chen, C.; Li, H.; Li, P.; Quinn, D.; Dao, M.; et al. Isolation of exosomes from whole blood by integrating acoustics and microfluidics. Proc. Natl. Acad. Sci. USA 2017, 114, 10584–10589. [Google Scholar] [CrossRef] [Green Version]
- Campanella, C.; Bavisotto, C.C.; Logozzi, M.; Gammazza, A.M.; Mizzoni, D.; Cappello, F.; Fais, S. On the Choice of the Extracellular Vesicles for Therapeutic Purposes. Int. J. Mol. Sci. 2019, 20, 236. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Li, S.; Li, L.; Li, M.; Guo, C.; Yao, J.; Mi, S. Exosome and Exosomal MicroRNA: Trafficking, Sorting, and Function. Genom. Proteom. Bioinform. 2015, 13, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Ong, S.-G.; Lee, W.H.; Huang, M.; Dey, D.; Kodo, K.; Sanchez-Freire, V.; Gold, J.D.; Wu, J.C. Cross Talk of Combined Gene and Cell Therapy in Ischemic Heart Disease: Role of Exosomal MicroRNA Transfer. Circulation 2014, 130, S60–S69. [Google Scholar] [CrossRef] [Green Version]
- Jansen, F.; Yang, X.; Baumann, K.; Przybilla, D.; Schmitz, T.; Flender, A.; Paul, K.; Alhusseiny, A.; Nickenig, G.; Werner, N. Endothelial microparticles reduce ICAM-1 expression in a microRNA-222-dependent mechanism. J. Cell. Mol. Med. 2015, 19, 2202–2214. [Google Scholar] [CrossRef]
- Hergenreider, E.; Heydt, S.; Tréguer, K.; Boettger, T.; Horrevoets, A.J.G.; Zeiher, A.M.; Scheffer, M.P.; Frangakis, A.S.; Yin, X.; Mayr, M.; et al. Atheroprotective communication between endothelial cells and smooth muscle cells through miRNAs. Nat. Cell Biol. 2012, 14, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Alexandru, N.; Andrei, E.; Safciuc, F.; Dragan, E.; Balahura, A.M.; Badila, E.; Georgescu, A. Intravenous Administration of Allogenic Cell-Derived Microvesicles of Healthy Origins Defends Against Atherosclerotic Cardiovascular Disease Development by a Direct Action on Endothelial Progenitor Cells. Cells 2020, 9, 423. [Google Scholar] [CrossRef] [Green Version]
- Myers, D.D.; Hawley, A.E.; Farris, D.M.; Wrobleski, S.K.; Thanaporn, P.; Schaub, R.G.; Wagner, D.D.; Kumar, A.; Wakefield, T.W. P-selectin and leukocyte microparticles are associated with venous thrombogenesis. J. Vasc. Surg. 2003, 38, 1075–1089. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Chen, S.; Liu, M.-L. Pathogenic roles of microvesicles in diabetic retinopathy. Acta Pharmacol. Sin. 2017, 39, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berezin, A.E.; Kremzer, A.A.; Samura, T.A.; Martovitskaya, Y.V.; Malinovskiy, Y.V.; Oleshko, S.V.; Berezina, T.A. Predictive value of apoptotic microparticles to mononuclear progenitor cells ratio in advanced chronic heart failure patients. J. Cardiol. 2015, 65, 403–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sterzenbach, U.; Putz, U.; Low, L.-H.; Silke, J.; Tan, S.-S.; Howitt, J. Engineered Exosomes as Vehicles for Biologically Active Proteins. Mol. Ther. 2017, 25, 1269–1278. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Zhang, J.; Zhao, Q.; Zhuang, W.; Ding, J.; Zhang, C.; Gao, H.; Pang, D.-W.; Pu, K.; Xie, H.-Y. Molecularly Engineered Macrophage-Derived Exosomes with Inflammation Tropism and Intrinsic Heme Biosynthesis for Atherosclerosis Treatment. Angew. Chem. Int. Ed. 2020, 59, 4068–4074. [Google Scholar] [CrossRef]
- Gibson, P.G.; Qin, L.; Puah, S.H. COVID-19 acute respiratory distress syndrome (ARDS): Clinical features and differences from typical pre-COVID-19 ARDS. Med. J. Aust. 2020, 213, 54–56.e1. [Google Scholar] [CrossRef]
- Zou, X.; Chen, K.; Zou, J.; Han, P.; Hao, J.; Han, Z. Single-cell RNA-seq data analysis on the receptor ACE2 expression reveals the potential risk of different human organs vulnerable to 2019-nCoV infection. Front. Med. 2020, 14, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Fox, S.E.; Akmatbekov, A.; Harbert, J.L.; Li, G.; Brown, J.Q.; Heide, R.S.V. Pulmonary and cardiac pathology in African American patients with COVID-19: An autopsy series from New Orleans. Lancet Respir. Med. 2020, 8, 681–686. [Google Scholar] [CrossRef]
- Tian, S.; Hu, W.; Niu, L.; Liu, H.; Xu, H.; Xiao, S.-Y. Pulmonary Pathology of Early-Phase 2019 Novel Coronavirus (COVID-19) Pneumonia in Two Patients with Lung Cancer. J. Thorac. Oncol. 2020, 15, 700–704. [Google Scholar] [CrossRef]
- Long, B.; Brady, W.J.; Koyfman, A.; Gottlieb, M. Cardiovascular complications in COVID-19. Am. J. Emerg. Med. 2020, 38, 1504–1507. [Google Scholar] [CrossRef]
- Lee, L.Y.W.; Cazier, J.-B.; Angelis, V.; Arnold, R.; Bisht, V.; Campton, N.A.; Chackathayil, J.; Cheng, V.W.; Curley, H.M.; Fittall, M.W.; et al. COVID-19 mortality in patients with cancer on chemotherapy or other anticancer treatments: A prospective cohort study. Lancet 2020, 395, 1919–1926. [Google Scholar] [CrossRef]
- Ali, M.A.; Spinler, S.A. COVID-19 and thrombosis: From bench to bedside. Trends Cardiovasc. Med. 2021, 31, 143–160. [Google Scholar] [CrossRef] [PubMed]
- Rand, J.H.; Glanville, R.W.; Wu, X.-X.; Ross, J.M.; Zangari, M.; Gordon, R.E.; Schwartz, E.; Potter, B.J. The Significance of Subendothelial von Willebrand Factor. Thromb. Haemost. 1997, 78, 445–450. [Google Scholar] [CrossRef]
- Leebeek, F.W.; Eikenboom, J.C. Von Willebrand’s Disease. N. Engl. J. Med. 2016, 375, 2067–2080. [Google Scholar] [CrossRef]
- Catanzaro, M.; Fagiani, F.; Racchi, M.; Corsini, E.; Govoni, S.; Lanni, C. Immune response in COVID-19: Addressing a pharmacological challenge by targeting pathways triggered by SARS-CoV-2. Signal Transduct. Target. Ther. 2020, 5, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Cao, J.; Wang, Q.; Shi, Q.; Liu, K.; Luo, Z.; Chen, X.; Chen, S.; Yu, K.; Huang, Z.; et al. D-dimer as a biomarker for disease severity and mortality in COVID-19 patients: A case control study. J. Intensiv. Care 2020, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Mackman, N.; Antoniak, S.; Wolberg, A.S.; Kasthuri, R.; Key, N.S. Coagulation Abnormalities and Thrombosis in Patients Infected With SARS-CoV-2 and Other Pandemic Viruses. Arter. Thromb. Vasc. Biol. 2020, 40, 2033–2044. [Google Scholar] [CrossRef] [PubMed]
- Nahand, J.S.; Moghoofei, M.; Salmaninejad, A.; Bahmanpour, Z.; Karimzadeh, M.; Nasiri, M.; Mirzaei, H.R.; Pourhanifeh, M.H.; Bokharaei-Salim, F.; Mirzaei, H.; et al. Pathogenic role of exosomes and microRNAs in HPV-mediated inflammation and cervical cancer: A review. Int. J. Cancer 2020, 146, 305–320. [Google Scholar] [CrossRef]
- Hassanpour, M.; Rezaie, J.; Nouri, M.; Panahi, Y. The role of extracellular vesicles in COVID-19 virus infection. Infect. Genet. Evol. 2020, 85, 104422. [Google Scholar] [CrossRef]
- Wang, J.; Chen, S.; Bihl, J. Exosome-Mediated Transfer of ACE2 (Angiotensin-Converting Enzyme 2) from Endothelial Progenitor Cells Promotes Survival and Function of Endothelial Cell. Oxidative Med. Cell. Longev. 2020, 2020, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujita, Y.; Hoshina, T.; Matsuzaki, J.; Kadota, T.; Fujimoto, S.; Kawamoto, H.; Watanabe, N.; Sawaki, K.; Sakomoto, Y.; Miyajima, M.; et al. Early prediction of COVID-19 severity using extracellular vesicles and extracellular RNAs. medRxiv 2020. [Google Scholar] [CrossRef]
- Rosell, A.; Havervall, S.; Von Meijenfeldt, F.; Hisada, Y.; Aguilera, K.; Grover, S.P.; Lisman, T.; Mackman, N.; Thålin, C. Patients With COVID-19 Have Elevated Levels of Circulating Extracellular Vesicle Tissue Factor Activity That Is Associated With Severity and Mortality—Brief Report. Arterioscler. Thromb. Vasc. Biol. 2021, 41, 878–882. [Google Scholar] [CrossRef]
- Guervilly, C.; Bonifay, A.; Burtey, S.; Sabatier, F.; Cauchois, R.; Abdili, E.; Arnaud, L.; Lano, G.; Pietri, L.; Robert, T.; et al. Dissemination of extreme levels of extracellular vesicles: Tissue factor activity in patients with severe COVID-19. Blood Adv. 2021, 5, 628–634. [Google Scholar] [CrossRef] [PubMed]
- Cappellano, G.; Raineri, D.; Rolla, R.; Giordano, M.; Puricelli, C.; Vilardo, B.; Manfredi, M.; Cantaluppi, V.; Sainaghi, P.P.; Castello, L.; et al. Circulating Platelet-Derived Extracellular Vesicles Are a Hallmark of Sars-Cov-2 Infection. Cells 2021, 10, 85. [Google Scholar] [CrossRef] [PubMed]
- Boilard, E. Thematic Review Series: Exosomes and Microvesicles: Lipids as Key Components of their Biogenesis and Functions Extracellular vesicles and their content in bioactive lipid mediators: More than a sack of microRNA. J. Lipid Res. 2018, 59, 2037–2046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnamachary, B.; Cook, C.; Spikes, L.; Chalise, P.; Dhillon, N.K. The Potential Role of Extracellular Vesicles in COVID-19 Associated Endothelial injury and Pro-inflammation. medRxiv 2020. [Google Scholar] [CrossRef]
- Barberis, E.; Vanella, V.V.; Falasca, M.; Caneapero, V.; Cappellano, G.; Raineri, D.; Ghirimoldi, M.; De Giorgis, V.; Puricelli, C.; Vaschetto, R.; et al. Circulating Exosomes Are Strongly Involved in SARS-CoV-2 Infection. Front. Mol. Biosci. 2021, 8, 916. [Google Scholar] [CrossRef]
- Shah, A. Novel Coronavirus-Induced NLRP3 Inflammasome Activation: A Potential Drug Target in the Treatment of COVID-19. Front. Immunol. 2020, 11, 1021. [Google Scholar] [CrossRef]
- Gedefaw, L.; Ullah, S.; Leung, P.; Cai, Y.; Yip, S.-P.; Huang, C.-L. Inflammasome Activation-Induced Hypercoagulopathy: Impact on Cardiovascular Dysfunction Triggered in COVID-19 Patients. Cells 2021, 10, 916. [Google Scholar] [CrossRef] [PubMed]
- Codo, A.C.; DaVanzo, G.G.; Monteiro, L.D.B.; De Souza, G.F.; Muraro, S.P.; Virgilio-Da-Silva, J.V.; Prodonoff, J.S.; Carregari, V.C.; Junior, C.A.O.D.B.; Crunfli, F.; et al. Elevated Glucose Levels Favor SARS-CoV-2 Infection and Monocyte Response through a HIF-1α/Glycolysis-Dependent Axis. Cell Metab. 2020, 32, 498–499. [Google Scholar] [CrossRef] [PubMed]
- Machhi, J.; Shahjin, F.; Das, S.; Patel, M.; Abdelmoaty, M.M.; Cohen, J.D.; Singh, P.A.; Baldi, A.; Bajwa, N.; Kumar, R.; et al. A Role for Extracellular Vesicles in SARS-CoV-2 Therapeutics and Prevention. J. Neuroimmune Pharmacol. 2021, 16, 270–288. [Google Scholar] [CrossRef] [PubMed]
- Khalaj, K.; Figueira, R.L.; Antounians, L.; Lauriti, G.; Zani, A. Systematic review of extracellular vesicle-based treatments for lung injury: Are EVs a potential therapy for COVID-19? J. Extracell. Vesicles 2020, 9, 517–520. [Google Scholar] [CrossRef] [PubMed]
- Brauer, R.; Ge, L.; Schlesinger, S.Y.; Birkland, T.P.; Huang, Y.; Parimon, T.; Lee, V.; McKinney, B.L.; McGuire, J.K.; Parks, W.C.; et al. Syndecan-1 Attenuates Lung Injury during Influenza Infection by Potentiating c-Met Signaling to Suppress Epithelial Apoptosis. Am. J. Respir. Crit. Care Med. 2016, 194, 333–344. [Google Scholar] [CrossRef] [Green Version]
- Mason, R.J. Hepatocyte growth factor: The key to alveolar septation? Am. J. Respir. Cell Mol. Biol. 2002, 26, 517–520. [Google Scholar] [CrossRef]
- McCarter, S.D.; Mei, S.; Lai, P.F.; Zhang, Q.W.; Parker, C.H.; Suen, R.S.; Hood, R.D.; Zhao, Y.D.; Deng, Y.; Han, R.N.N.; et al. Cell-based Angiopoietin-1 Gene Therapy for Acute Lung Injury. Am. J. Respir. Crit. Care Med. 2007, 175, 1014–1026. [Google Scholar] [CrossRef] [Green Version]
- Gutbier, B.; Neuhauß, A.-K.; Reppe, K.; Ehrler, C.; Santel, A.; Kaufmann, J.; Scholz, M.; Weissmann, N.; Morawietz, L.; Mitchell, T.J.; et al. Prognostic and Pathogenic Role of Angiopoietin-1 and -2 in Pneumonia. Am. J. Respir. Crit. Care Med. 2018, 198, 220–231. [Google Scholar] [CrossRef] [Green Version]
- Guzik, T.J.; Mohiddin, S.A.; DiMarco, A.; Patel, V.; Savvatis, K.; Marelli-Berg, F.M.; Madhur, M.S.; Tomaszewski, M.; Maffia, P.; D’Acquisto, F.; et al. COVID-19 and the cardiovascular system: Implications for risk assessment, diagnosis, and treatment options. Cardiovasc. Res. 2020, 116, 1666–1687. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, V.; Sengupta, S.; Lazo, A.; Woods, P.; Nolan, A.; Bremer, N. Exosomes Derived from Bone Marrow Mesenchymal Stem Cells as Treatment for Severe COVID-19. Stem Cells Dev. 2020, 29, 747–754. [Google Scholar] [CrossRef]
- Zhou, F.; Yu, T.; Du, R.; Fan, G.; Liu, Y.; Liu, Z.; Xiang, J.; Wang, Y.; Song, B.; Gu, X.; et al. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: A retrospective cohort study. Lancet 2020, 395, 1054–1062. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
 increase.
increase.
increase.
increase.| Extracellular Vesicles (EVs) as Prognostic and Biomarkers in COVID-19 and Related Thrombosis | |||
|---|---|---|---|
| EV Isolated from Serum or Plasma of COVID-19 Patients | Extracellular Vesicles– Molecular Changes | EVs as Prognostic and Biomarkers in COVID-19 Patients | References |
| EVs from serum (31 COVID-19 patients (22 with mild and 9 with severe symptoms)/10 healthy subjects (control)) | COPB2/KRAS/PRKCB/RHOC expression in patients with mild COVID-19 symptoms CD147/CAPN2/ECM1/FGG expression in patients with severe COVID-19 symptoms exRNA:SNORD33/AL732437.2/RNU2-29P/CDKN2B-AS1/ AL365184.1/miR-122-5p expression in patients with severe COVID-19 symptoms |
| [147] |
| Plasma EVs (100 COVID-19 patients (with mild and severe symptoms)/28 healthy subjects (control)) | TF-positive EV levels in COVID-19 patients EV-TF activity in COVID-19 patients EV-TF activity—positively correlated with plasma markers: D-dimer, PT, INR, prothrombin, fibrinogen, antithrombin, vWF (markers directly associated with thrombosis) |
| [148,149] |
| Blood PMVs (69 COVID-19 patients, 62 patients after COVID-19, 10 healthy subjects) | PMV levels in COVID-19 patients
|
| [150] |
| EVs from plasma (53 COVID-19 patients (with mild and severe symptoms)/healthy subjects (control)) | PMV levels in patients with mild COVID-19 symptoms exosome levels in patients with severe COVID-19 symptoms TF/t-PA/vWF/CD163/EN-RAGE expression (prothrombotic/endothelial injury factors) in COVID-19 patients with severe pathology HGF/MB/REN/prostasin/PRSS8 (factors associated with cardiovascular pathology) in COVID-19 patients with severe pathology TNF-α/IL-6/MCP-1 levels in patients with mild and severe COVID-19 symptoms caspase 3/7 activity in patients with mild and severe COVID-19 symptoms |
| [152] |
increase;  decrease.
increase; decrease.
decrease.
increase; decrease.| Employing Extracellular Vesicles (EVs) as Therapeutic Agents in COVID-19 | |||
|---|---|---|---|
| Administration of MSC-Derived EVs to Patients | Improved Effects | EV Effects in COVID-19 Patients | References |
| MSC-derived exosomes administered i.v. to 24 COVID-19 patients with moderate to severe pathology | CRP/ferritin/D-dimer levels neutrophil number CD3+/CD4+/CD8+ lymphocyte number PaO2/FiO2 ratio |
| [164] |
| MSC-derived exosomes administered by aerosol inhalation to COVID-19 patients: 5 days to 24 patients; twice a day, 10 days to 30 patients | No official results yet | ClinicalTrials.gov: NCT04276987/ NCT04491240/ NCT04602442 | |
| Engineered exosomes overexpressing CD24, aerosolized by inhalation, once a day, 5 days to 35 COVID-19 patients | suppress cytokine storm |
| ClinicalTrials.gov: NCT04747574 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Georgescu, A.; Simionescu, M. Extracellular Vesicles: Versatile Nanomediators, Potential Biomarkers and Therapeutic Agents in Atherosclerosis and COVID-19-Related Thrombosis. Int. J. Mol. Sci. 2021, 22, 5967. https://doi.org/10.3390/ijms22115967
Georgescu A, Simionescu M. Extracellular Vesicles: Versatile Nanomediators, Potential Biomarkers and Therapeutic Agents in Atherosclerosis and COVID-19-Related Thrombosis. International Journal of Molecular Sciences. 2021; 22(11):5967. https://doi.org/10.3390/ijms22115967
Chicago/Turabian StyleGeorgescu, Adriana, and Maya Simionescu. 2021. "Extracellular Vesicles: Versatile Nanomediators, Potential Biomarkers and Therapeutic Agents in Atherosclerosis and COVID-19-Related Thrombosis" International Journal of Molecular Sciences 22, no. 11: 5967. https://doi.org/10.3390/ijms22115967
APA StyleGeorgescu, A., & Simionescu, M. (2021). Extracellular Vesicles: Versatile Nanomediators, Potential Biomarkers and Therapeutic Agents in Atherosclerosis and COVID-19-Related Thrombosis. International Journal of Molecular Sciences, 22(11), 5967. https://doi.org/10.3390/ijms22115967