
Genome-Wide Approach to Identify Quantitative Trait Loci for Drought Tolerance in Tetraploid Potato (Solanum tuberosum L.)


 , , , , ,
, , , , , 
Abstract
:1. Introduction
2. Results
2.1. Informative Markers for Defining Linkage Groups in Starch Potato Cultivars
2.2. QTL Mapping Links Drought Tolerance Index DRYM to Starch Yield Parameters
2.3. Carbohydrate Metabolism Candidate Genes in QTL Regions
2.4. Ethylene Synthesis and Other Stress-Related Factors Co-Localize with DRYM QTL
2.5. Standalone QTL on LG3 Includes Cytochrome P450, Cell Wall Remodeling Genes and Phytohormone Signaling Factors
2.6. Expression QTL for Drought Transcript Markers Overlap with DRYM QTL
2.7. QTL of Drought-Responsive Metabolites Overlap with DRYM QTL
3. Discussion
3.1. Abiotic vs. Biotic Stress Response under Drought
3.2. Cell Wall Remodeling Genes under DRYM QTL
3.3. Nonsense Mutations in Genes under DRYM QTL on LG3
3.4. Co-Localization of Drought Tolerance and Candidate Genes for Starch Metabolism
3.5. Improvement of Drought Tolerance Requires Identification of Stress-Related Mechanisms that Do Not Affect Yield-Relevant Metabolism
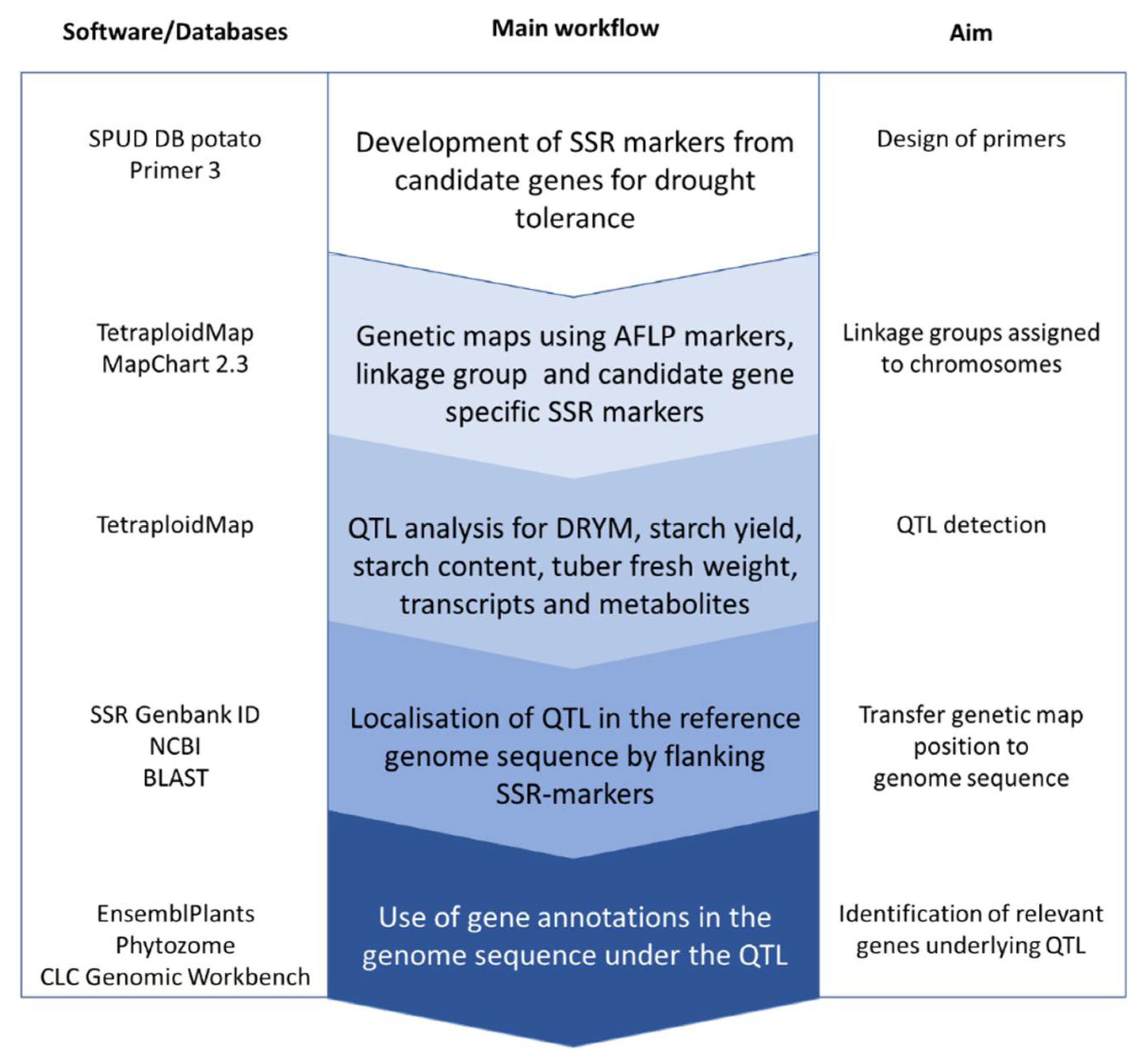
4. Materials and Methods
4.1. Plant Material and Experimental Design
4.2. Control and Drought Stress Treatment
4.3. SSR and AFLP Analyses
4.4. Linkage Mapping Using Tetraploid Map
4.5. QTL Analysis of Drought Tolerance and Yield-Associated Traits
4.6. Search for Candidate Genes in Databases
4.7. Whole-Genome Sequencing in Tetraploid Potato
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Obidiegwu, J.E.; Bryan, G.J.; Jones, H.G.; Prashar, A. Coping with drought: Stress and adaptive responses in potato and perspectives for improvement. Front. Plant Sci. 2015, 6, 542. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, J.R.; Kossmann, J. Starch Trek: The search for yield. Front. Plant Sci. 2019, 9, 1930. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.S.; Amorim, E.P.; Ferreira, C.F.; Pirovani, C.P. Water stress in Musa spp.: A systematic review. PLoS ONE 2018, 13, e0208052. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Shi, S.; Liu, Z.; Yang, F.; Yin, G. Drought tolerance in alfalfa (Medicago sativa L.) varieties is associated with enhanced antioxidative protection and declined lipid peroxidation. J. Plant Physiol. 2019, 232, 226–240. [Google Scholar] [CrossRef] [PubMed]
- Marshall, A.; Aalen, R.B.; Audenaert, D.; Beeckman, T.; Broadley, M.R.; Butenko, M.A.; Cano-Delgado, A.I.; de Vries, S.; Dresselhaus, T.; Felix, G.; et al. Tackling drought stress: Receptor-like kinases present new approaches. Plant Cell. 2012, 24, 2262–2278. [Google Scholar] [CrossRef]
- Joshi, R.; Wani, S.H.; Singh, B.; Bohra, A.; Dar, Z.A.; Lone, A.A.; Pareek, A.; Singla-Pareek, S.L. Transcription factors and plants response to drought stress: Current understanding and future directions. Front. Plant Sci. 2016, 7, 1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potato Genome Sequencing Consortium. Genome sequence and analysis of the tuber crop potato. Nature 2011, 475, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Bourke, P.M.; Voorrips, R.E.; Visser, R.G.; Maliepaard, C. The double-reduction landscape in tetraploid potato as revealed by a high-density linkage map. Genetics 2015, 201, 853–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pieczynski, M.; Wyrzykowska, A.; Milanowska, K.; Boguszewska-Mankowska, D.; Zagdanska, B.; Karlowski, W.; Jarmolowski, A.; Szweykowska-Kulinska, Z. Genomewide identification of genes involved in the potato response to drought indicates functional evolutionary conservation with Arabidopsis plants. Plant. Biotechnol. J. 2017, 16, 603–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, L.; Zhang, H.; Gan, X.; Zhang, L.; Chen, Y.; Nie, F.; Shi, L.; Li, M.; Guo, Z.; Zhang, G.; et al. Transcriptome profiling of the potato (Solanum tuberosum L.) plant under drought stress and water-stimulus conditions. PLoS ONE 2015, 10, e0128041. [Google Scholar] [CrossRef]
- Moon, K.B.; Ahn, D.J.; Park, J.S.; Jung, W.Y.; Cho, H.S.; Kim, H.R.; Jeon, J.H.; Park, Y.I.; Kim, H.S. Transcriptome profiling and characterization of drought-tolerant potato plant (Solanum tuberosum L.). Mol. Cells 2018, 41, 979–992. [Google Scholar] [CrossRef] [PubMed]
- Schafleitner, R.; Gutierrez Rosales, R.O.; Gaudin, A.; Alvarado Aliaga, C.A.; Martinez, G.N.; Tincopa Marca, L.R.; Bolivar, L.A.; Delgado, F.M.; Simon, R.; Bonierbale, M. Capturing candidate drought tolerance traits in two native Andean potato clones by transcription profiling of field grown plants under water stress. Plant Physiol. Biochem. 2007, 45, 673–690. [Google Scholar] [CrossRef]
- Sprenger, H.; Kurowsky, C.; Horn, R.; Erban, A.; Seddig, S.; Rudack, K.; Fischer, A.; Walther, D.; Zuther, E.; Köhl, K.; et al. The drought response of potato reference cultivars with contrasting tolerance. Plant Cell Environ. 2016, 39, 2370–2389. [Google Scholar] [CrossRef] [PubMed]
- Sprenger, H.; Erban, A.; Seddig, S.; Rudack, K.; Thalhammer, A.; Le, M.Q.; Walther, D.; Zuther, E.; Köhl, K.I.; Kopka, J.; et al. Metabolite and transcript markers for the prediction of potato drought tolerance. Plant Biotechnol. J. 2018, 16, 939–950. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Zhao, P.; Kong, N.; Lu, R.; Pei, Y.; Huang, C.; Ma, H.; Chen, Q. Genome-wide identification and characterization of the potato bHLH transcription factor family. Genes 2018, 9, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, P.; Wang, D.; Wang, R.; Kong, N.; Zhang, C.; Yang, C.; Wu, W.; Ma, H.; Chen, Q. Genome-wide analysis of the potato Hsp20 gene family: Identification, genomic organization and expression profiles in response to heat stress. BMC Genom. 2018, 19, 61. [Google Scholar] [CrossRef]
- Anithakumari, A.M.; Nataraja, K.N.; Visser, R.G.; van der Linden, C.G. Genetic dissection of drought tolerance and recovery potential by quantitative trait locus mapping of a diploid potato population. Mol. Breed. 2012, 30, 1413–1429. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A.; Saravia, D.; Munive, S.; Lozano, F.; Farfan, E.; Eyzaguirre, R.; Bonierbale, M. Multiple QTLs linked to agro-morphological and physiological traits related to drought tolerance in potato. Plant Mol. Biol. Rep. 2015, 33, 1286–1298. [Google Scholar] [CrossRef] [Green Version]
- da Silva, W.L.; Ingram, J.; Hackett, C.A.; Coombs, J.J.; Douches, D.; Bryan, G.J.; De Jong, W.; Gray, S. Mapping loci that control tuber and foliar symptoms caused by PVY in autotetraploid potato (Solanum tuberosum L.). Genes Genomes Genet. 2017, 7, 3587–3595. [Google Scholar] [CrossRef]
- Massa, A.N.; Manrique-Carpintero, N.C.; Coombs, J.; Haynes, K.G.; Bethke, P.C.; Brandt, T.L.; Gupta, S.K.; Yencho, G.C.; Novy, R.G.; Douches, D.S. Linkage analysis and QTL mapping in a tetraploid russet mapping population of potato. BMC Genet. 2018, 19, 87. [Google Scholar] [CrossRef] [Green Version]
- Schönhals, E.M.; Ortega, F.; Barandalla, L.; Aragones, A.; Riuz de Galareta, J.I.; Liao, J.-C.; Sanetomo, R.; Walkemeier, B.; Tacke, E.; Ritter, E.; et al. Identification and reproducibility of diagnostic DNA markers for tuber starch and yield optimization in a novel association mapping population of potato (Solanum tuberosum L.). Theor. Appl. Genet. 2016, 129, 767–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santa, J.D.; Berdugo-Cely, J.; Cely-Pardo, L.; Soto-Suarez, M.; Mosquera, T.; Galeano, M.C. QTL analysis reveals quantitative resistant loci for Phytophthora infestans and Tecia solanivora in tetraploid potato (Solanum tuberosum L.). PLoS ONE 2018, 13, e0199716. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, M.; Soolanayakanahally, R.; Ogawa, S.; Uga, Y.; Selvaraj, M.G.; Kagale, S. Drought response in wheat: Key genes and regulatory mechanisms controlling root system architecture and transpiration efficiency. Front. Chem. 2017, 5, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dash, M.; Yordanov, Y.S.; Georgieva, T.; Wei, H.; Busov, V. Gene network analysis of poplar root transcriptome in response to drought stress identifies a PtaJAZ3PtaRAP2.6-centered hierarchical network. PLoS ONE 2018, 13, e0208560. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krannich, C.T.; Maletzki, L.; Kurowsky, C.; Horn, R. Network candidate genes in breeding for drought tolerant crops. Int. J. Mol. Sci. 2015, 16, 16378–16400. [Google Scholar] [CrossRef]
- Zare, M. Evaluation of drought tolerance indices for the selection of Iranian barley Hordeum vulgare cultivars. Afr. J. Biotechnol. 2012, 11, 15975–15981. [Google Scholar] [CrossRef] [Green Version]
- Fischer, R.A.; Maurer, R. Drought resistance in spring wheat cultivars. I Grain yield responses. Aust. J. Agric. Res. 1978, 29, 897–912. [Google Scholar] [CrossRef]
- Fernandez, G.C.J. Effective selection criteria for assessing stress tolerance. In Proceedings of the International Symposium on Adaption of Vegetables and Other Food Crops in Temperature and Water Stress, Tainan, Taiwan, 13–16 August 1992; pp. 257–270. [Google Scholar]
- Schneider, K.A.; Rosales-Serna, R.; Ibarra-Perez, F.; Cazares-Enriquez, B.; Acosta-Gallegos, J.A.; Ramirez-Vallejo, P.; Wassimi, N.; Kelly, J.D. Improving common bean performance under drought stress. Crop Sci. 1997, 37, 43–50. [Google Scholar] [CrossRef]
- Bouslama, M.; Schapaugh, W.T. Stress tolerance in soybean. Part 1. Evaluation of three screening techniques for heat and drought tolerance. Crop Sci. 1984, 24, 933–937. [Google Scholar] [CrossRef]
- Sprenger, H.; Rudack, K.; Schudoma, C.; Neumann, A.; Seddig, S.; Peters, R.; Zuther, E.; Kpka, J.; Hincha, D.K.; Walther, D.; et al. Assessment of drought tolerance and its potential yield penalty in potato. Funct. Plant Biol. 2015, 42, 655–667. [Google Scholar] [CrossRef] [PubMed]
- Ghislain, M.; Núñez, J.; Herrera, M.R.; Pignataro, J.; Guzman, F.; Bonierbale, M.; Spooner, D.M. Robust and highly informative microsatellite-based genetic identity kit for potato. Mol. Breed. 2009, 23, 377–388. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, C.; Krannich, C.T.; Maletzki, L.; Köhl, K.; Kopka, J.; Sprenger, H.; Hincha, D.K.; Seddig, S.; Peters, R.; Hamera, S.; et al. Unravelling differences in candidate genes for drought tolerance in potato (Solanum tuberosum L.) by use of new functional microsatellite markers. Genes 2021, 12, 494. [Google Scholar] [CrossRef]
- Chen, X.; Salamini, F.; Gebhardt, C. A potato molecular-function map for carbohydrate metabolism and transport. Theor. Appl. Genet. 2001, 102, 284–295. [Google Scholar] [CrossRef]
- Helle, S.; Bray, F.; Verbeke, J.; Devassine, S.; Courseaux, A.; Facon, M.; Tokarski, C.; Rolando, C.; Szydlowski, N. Proteome analysis of potato starch reveals the presence of new starch metabolic proteins as well as multiple protease inhibitors. Front. Plant Sci. 2018, 9, 746. [Google Scholar] [CrossRef] [PubMed]
- Schönhals, E.M.; Ding, J.; Ritter, E.; Paulo, M.J.; Cara, N.; Tacke, E.; Hofferbert, H.-R.; Lübeck, J.; Strahwald, J.; Gebhardt, C. Physical mapping of QTL for tuber yield, starch content and starch yield in tetraploid potato (Solanum tuberosum L.) by means of genome wide genotyping by sequencing and the 8.3 K SolCAP SNP array. BMC Genom. 2017, 18, 642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, M.; Sprenger, H.; Zuther, E.; Peter, R.; Seddig, S.; Kopka, J.; Walther, D.; Hincha, D.K.; Köhl, K.I. Can metabolite- and transcript-based selection for drought tolerance in Solanum tubersosum replace selection on yield in arid environments? Front. Plant Sci. 2020, 11, 1071. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.L.C.; Li, H.; Ecker, J.R. Ethylene biosynthesis and signaling networks. Plant Cell 2002, 14, S131–S151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Xiong, Q.; Yin, C.-C.; Ma, B.; Chen, S.-Y.; Zhang, J.-S. Ethylene biosynthesis, signaling, and crosstalk with other hormones in rice. Small Methods 2019, 1900278. [Google Scholar] [CrossRef]
- Potuschak, T.; Lechner, E.; Parmentiert, Y.; Yanagisawa, S.; Grava, S.; Koncz, C.; Genschik, P. EIN3-dependent regulation of plant ethylene hormone signaling by two Arabidopsis F box proteins: EBF1 and EBF2. Cell 2003, 115, 679–689. [Google Scholar] [CrossRef] [Green Version]
- Chao, Q.; Rothenberg, M.; Solano, R.; Roman, G.; Terzaghi, W.; Ecker, J.R. Activation of the ethylene gas response pathway in Arabidopsis by the nuclear protein ETHYLENE-INSENSITIVE3 and related proteins. Cell 1997, 89, 1133–1144. [Google Scholar] [CrossRef] [Green Version]
- An, F.; Zhao, Q.; Ji, Y.; Li, W.; Jiang, Z.; Yu, X.; Zhang, C.; Han, Y.; He, W.; Liu, Y.; et al. Ethylene-induced stabilization of ETHYLENE INSENSITIVE3 and EIN3-LIKE1 is mediated by proteasomal degradation of EIN3 Binding F-Box 1 and 2 that requires EIN2 in Arabidopsis. Plant Cell 2010, 22, 2384–2401. [Google Scholar] [CrossRef] [Green Version]
- Nakano, T.; Suzuki, K.; Fujimura, T.; Shinshi, H. Genome-wide analysis of the ERF gene family in Arabidopsis and rice. Plant Physiol. 2006, 140, 411–432. [Google Scholar] [CrossRef] [Green Version]
- Heyman, J.; Canher, B.; Bisht, A.; Christiaens, F.; De Veylder, L. Emerging role of the plant ERF transcription factors in coordination wound defense responses and repair. J. Cell Sci. 2018, 131, jcs208215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Cadenas, A.; Tadeo, F.R.; Talon, M.; Primo-Millo, E. Leaf abscission induced by ethylene in water-stressed intact seedlings of Cleopatra mandarin requires previous abscisic acid accumulation in roots. Plant Physiol. 1996, 112, 401–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, H.; Chen, S.; Liu, D.; Liesche, J.; Shi, C.; Wang, J.; Ren, M.; Wang, X.; Yang, J.; Shi, W.; et al. Ethylene-induced hydrogen sulfide negatively regulates ethylene biosynthesis by persulfidation of ACO in tomato under osmotic stress. Front. Plant Sci. 2018, 9, 1517. [Google Scholar] [CrossRef] [Green Version]
- Jordan, W.R.; Morgan, P.W.; Davenport, T.L. Water stress enhances ethylene-mediated leaf abscission in cotton. Plant Physiol. 1972, 50, 756–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, C.C.; Zhao, H.; Ma, B.; Chen, S.Y.; Zhang, J.S. Diverse roles of ethylene in regulating agronomic traits in rice. Front. Plant Sci. 2017, 8, 1676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyd, N.S.; Gordon, R.; Martin, R.C. Relationship between leaf area index and ground cover in potato under different management conditions. Potato Res. 2002, 45, 117–129. [Google Scholar] [CrossRef]
- Arc, E.; Sechet, J.; Corbineau, F.; Rajjou, L.; Marion-Poll, A. ABA crosstalk with ethylene and nitric oxide in seed dormancy and germination. Front. Plant. Sci. 2013, 4, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, J.G.; Ellis, C.; Devoto, A. The jasmonate signal pathway. Plant Cell 2002, 14, S153–S164. [Google Scholar] [CrossRef] [Green Version]
- Acharya, B.R.; Assmann, S.M. Hormone interactions in stomatal function. Plant Mol. Biol. 2009, 69, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Wasternack, C.; Hause, B. Jasmonates: Biosynthesis, perception, signal transduction and action in plant stress response; growth and development. Ann. Bot. 2013, 111, 1021–1058. [Google Scholar] [CrossRef] [PubMed]
- Jing, H.; Li, C.; Ma, F.; Ma, J.-H.; Khan, A.; Wang, X.; Zhao, L.-Y.; Gong, Z.-H.; Chen, R.-G. Genome-wide identification, expression diversication of dehydrin gene family and characterization of CaDHN3 in pepper (Capsicum annuum L.). PLoS ONE 2016, 11, e0161073. [Google Scholar] [CrossRef]
- van Muijen, D.; Anithakumari, A.M.; Maliepaard, C.; Visser, R.G.; van der Linden, C.G. Systems genetics reveals key genetic elements of drought induced gene regulation in diploid potato. Plant Cell Environ. 2016, 39, 1895–1908. [Google Scholar] [CrossRef]
- Alexandersson, E.; Mulugeta, T.; Lankinen, A.; Liljeroth, E.; Andreasson, E. Plant resistance inducers against pathogens in Solanaceae species-from molecular mechanisms to field application. Int. J. Mol. Sci. 2016, 17, E1673. [Google Scholar] [CrossRef] [PubMed]
- Shu, K.; Zhou, W.; Chen, F.; Luo, X.; Yang, W. Abscisic acid and gibberellins antagonistically mediate plant development and abiotic stress responses. Front. Plant Sci. 2018, 9, 416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Pascal, J.M.; Ellenberger, T. The rise and fall of poly(ADP-ribose): An enzymatic perspective. DNA Repair 2015, 32, 10–16. [Google Scholar] [CrossRef] [Green Version]
- Hanai, S.; Kanai, M.; Ohashi, S.; Okamoto, K.; Yamada, M.; Takahashi, H.; Miwa, M. Loss of poly(ADP-ribose) glycohydrolase causes progressive neurodegeneration in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2004, 101, 82–86. [Google Scholar] [CrossRef] [Green Version]
- Rouleau, M.; Patel, A.; Hendzel, M.J.; Kaufmann, S.H.; Poirier, G.G. PARP inhibition: PARP1 and beyond. Nat. Rev. Cancer 2010, 10, 293–301. [Google Scholar] [CrossRef] [Green Version]
- Briggs, A.G.; Bent, A.F. Poly(ADP-ribosyl)ation in plants. Trends Plant. Sci. 2011, 16, 372–380. [Google Scholar] [CrossRef]
- Li, G.; Nasar, V.; Yang, Y.; Li, W.; Liu, B.; Sun, L.; Li, D.; Song, F. Arabidopsis poly(ADP-ribose) glycohydrolase 1 is required for drought; osmotic and oxidative stress responses. Plant Sci. 2011, 180, 283–291. [Google Scholar] [CrossRef]
- Vainonen, J.P.; Shapiguzov, A.; Vaattovaara, A.; Kangasjärvi, J. Plant PARPs, PARGs and PARP-like proteins. Curr. Protein Pept. Sci. 2016, 17, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Briggs, A.G.; Adams-Phillips, L.C.; Keppler, B.D.; Zebell, S.G.; Arend, K.C.; Apfelbaum, A.A.; Smith, J.A.; Bent, A.F. A transcriptomics approach uncovers novel roles for poly(ADP-ribosyl)ation in the basal defense response in Arabidopsis thaliana. PLoS ONE 2017, 12, e0190268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novaković, L.; Guo, T.; Bacic, A.; Sampathkumar, A.; Johnson, K.L. Hitting the wall-sensing and signaling pathways involved in plant cell wall remodeling in response to abiotic stress. Plants 2018, 7, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosgrove, D.J. Loosening of plant cell walls by expansins. Nature 2000, 407, 321–326. [Google Scholar] [CrossRef]
- Le Gall, H.; Philippe, F.; Domon, J.M.; Gillet, F.; Pelloux, J.; Rayon, C. Cell wall metabolism in response to abiotic stress. Plants 2015, 4, 112–166. [Google Scholar] [CrossRef] [PubMed]
- Brady, S.M.; Song, S.; Dhugga, K.S.; Rafalski, J.A.; Benfey, P. Combining expression and comparative evolutionary analysis. The COBRA gene family. Plant Physiol. 2007, 143, 172–187. [Google Scholar] [CrossRef] [Green Version]
- Leng, Y.; Yang, Y.; Ren, D.; Huang, L.; Dai, L.; Wang, Y.; Chen, L.; Tu, Z.; Gao, Y.; Li, X.; et al. A rice PECTATE LYASE_LIKE gene is required for plant growth and leaf senescence. Plant Physiol. 2018, 174, 1151–1166. [Google Scholar] [CrossRef] [Green Version]
- Tenhaken, R. Cell wall remodeling under abiotic stress. Front. Plant Sci. 2014, 5, 771. [Google Scholar] [CrossRef] [Green Version]
- Rudack, K.; Seddig, S.; Sprenger, H.; Köhl, K.; Uptmoor, R.; Ordon, F. Drought stress-induced changes in starch yield and physiological traits in potato. J. Agron. Crop Sci. 2017, 203, 494–505. [Google Scholar] [CrossRef]
- Shang, Y.; Dai, C.; Lee, M.M.; Kwak, J.M.; Na, K.H. BRI1-associated receptor kinase 1 regulates guard cell ABA signaling mediated by open stomata 1 in Arabidopsis. Mol. Plant 2016, 9, 447–460. [Google Scholar] [CrossRef] [Green Version]
- Pandian, B.A.; Sathishraj, R.; Djanaguiraman, M.; Prasad, P.V.V.; Jugulam, M. Role of cytochrome P450 enzymes in plant stress response. Antioxidants 2020, 9, 454. [Google Scholar] [CrossRef]
- Höwing, T.; Dann, M.; Müller, B.; Helm, M.; Scholz, S.; Schneitz, K.; Hammes, U.Z.; Gietl, C. The role of KDEL-tailed cysteine endopeptidase of Arabidopsis (AtCEP2 and AtCEP1) in root development. PLoS ONE 2018, 13, e0209407. [Google Scholar] [CrossRef]
- Rivero, R.M.; Kojima, M.; Gepstein, A.; Sabakibara, H.; Mittler, R.; Gepstein, S.; Blumwald, E. Delayed leaf senescence induces extreme drought tolerance in a flowering plant. Proc. Natl. Acad. Sci. USA 2007, 104, 19631–19636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.-L.; Guo, H.; Li, Z. Gene network analysis of senescence-associated genes in annual plants and comparative assessment of aging in perennials and animals. Transl. Med. Aging 2019, 3, 6–13. [Google Scholar] [CrossRef]
- Weaver, L.M.; Gan, S.; Quirino, B.; Amasino, R.M. A comparison of the expression patterns of several senescence-associated genes in response to stress and hormone treatment. Plant Mol. Biol. 1998, 37, 455–469. [Google Scholar] [CrossRef] [PubMed]
- Gregersen, P.L.; Culetic, A.; Boschian, L.; Krupinska, K. Plant senescence and crop productivity. Plant Mol. Biol. 2013, 82, 603–622. [Google Scholar] [CrossRef] [PubMed]
- Kyriakidou, M.; Tai, H.H.; Anglin, N.L.; Ellis, D.; Stromvik, M.V. Current strategies of polyploid plant genome sequence assembly. Front. Plant Sci. 2018, 9, 1660. [Google Scholar] [CrossRef]
- Seung, D.; Boudet, J.; Monroe, J.; Schreier, T.B.; David, L.C.; Abt, M.; Lu, K.J.; Zanella, M.; Zeerman, S.C. Homologs of PROTEIN TARGETING TO STARCH control starch granule initiation in Arabidopsis leaves. Plant Cell 2017, 29, 1657–1677. [Google Scholar] [CrossRef] [Green Version]
- Vikram, P.; Mallikarjuna Swamy, B.P.; Dixit, S.; Singh, R.; Singh, B.P.; Miro, B.; Kohli, A.; Henry, A.; Singh, N.K.; Kumar, A. Drought susceptibility of modern rice varieties: An effect of linkage of drought tolerance with undesirable traits. Sci. Rep. 2015, 5, 14799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wani, S.H.; Singh, N.B.; Haribhushan, A.; Mir, J.I. Compatible solute engineering in plants for abiotic stress tolerance—Role of glycine betaine. Curr. Genom. 2013, 14, 157–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kloostermann, B.; Abelenda, J.A.; Gomez, M.C.; Oortwijn, M.; de Broer, J.M.; Kowitwanich, K.; Horvath, B.M.; van Eck, H.J.; Smaczniak, C.; Prat, S.; et al. Naturally occurring allele diversity allows potato cultivation in northern latitudes. Nature 2013, 495, 246–250. [Google Scholar] [CrossRef] [PubMed]
- Boguszewska-Mańkowska, D.; Pieczyński, M.; Wyrzykowska, A.; Kalaji, H.M.; Sieczko, L.; Szweykowska-Kulińska, Z.; Zagdańska, B. Divergent strategies displayed by potato (Solanum tuberosum L.) cultivars to cope with soil drought. J. Agron. Crop. Sci. 2018, 204, 13–30. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.; Bhardwaj, R.; Thukrai, A.K.; Handa, N.; Kaur, R.; Kumar, V. Osmolyte dynamics: New strategies for crop tolerance to abiotic stress signals. In Emerging Technologies and Management of Crop Stress Tolerance; Ahmad, P., Ed.; Elvesier: Amsterdam, The Netherlands, 2014; Volume 2, pp. 405–430. [Google Scholar] [CrossRef]
- Zhou, M.-L.; Zhang, Q.; Zhou, M.; Sun, Z.-M.; Zhu, X.-M.; Shao, J.-R.; Tang, Y.-X.; Wu, Y.-M. Genome-wide identification of genes involved in raffinose metabolism in maize. Glycobiology 2012, 22, 1775–1785. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, A.; Yabuta, Y.; Shigeoka, S. Galactinol and raffinose constitute a novel function to protect plants from oxidative damage. Plant Physiol. 2008, 147, 1251–1263. [Google Scholar] [CrossRef] [Green Version]
- Taji, T.; Ohsumi, C.; Iuchi, S.; Seki, M.; Kasuga, M.; Kobayashi, M.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Important roles of drought- and cold-inducible genes for galactinol synthase in stress tolerance in Arabidopsis thaliana. Plant J. 2002, 29, 417–426. [Google Scholar] [CrossRef]
- Sengupta, S.; Mukherjee, S.; Basak, P.; Majumder, A.L. Significance of galactinol and raffinose family oligosaccharide synthesis in plants. Front. Plant Sci. 2015, 6, 656. [Google Scholar] [CrossRef] [Green Version]
- Kang, Z.; Babar, M.A.; Khan, N.; Guo, J.; Khan, J.; Islam, S.; Shrestha, S.; Shahi, D. Comparative metabolomic profiling in the roots and leaves in contrasting genotypes reveals complex mechanisms involved in post-anthesis drought tolerance in wheat. PLoS ONE 2019, 14, e0213502. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid total DNA preparation procedure for fresh plant tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Köhl, K.I.; Basler, G.; Luedemann, A.; Selbig, J.; Walther, D. A plant resource and experiment management system based on the Golm Plant Database as a basic tool for omics research. Plant Methods 2008, 4, 11. [Google Scholar] [CrossRef] [Green Version]
- Feingold, S.; Lloyd, J.; Norero, N.; Bonierbale, M.; Lorenzen, J. Mapping and characterization of new EST-derived microsatellites for potato (Solanum tuberosum L.). Theor. Appl. Genet. 2005, 111, 456–466. [Google Scholar] [CrossRef] [PubMed]
- Milbourne, D.; Meyer, R.C.; Collins, A.J.; Ramsay, L.D.; Gebhardt, C.; Waugh, R. Isolation, characterisation and mapping of simple sequence repeat loci in potato. Mol. Gen. Genet. 1998, 259, 233–245. [Google Scholar] [CrossRef]
- Provan, J.; Powell, W.; Waugh, R. Microsatellite analysis of relationships within cultivated potato (Solanum tuberosum). Theor. Appl. Genet. 1996, 92, 1078–1084. [Google Scholar] [CrossRef] [PubMed]
- Veilleux, R.E.; Shen, L.Y.; Paz, M.M. Analysis of the genetic composition of anther-derived potato by randomly amplified polymorphic DNA and simple sequence repeats. Genome 1995, 38, 1153–1162. [Google Scholar] [CrossRef] [PubMed]
- Sajer, O.; Scorza, R.; Dardick, C.; Zhebentyayeva, T.; Abbott, A.G.; Horn, R. Development of sequence-tagged site markers linked to the pillar growth type in peach (Prunus persica). Plant Breed. 2012, 131, 186–192. [Google Scholar] [CrossRef]
- Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; van de Lee, T.; Hornes, M.; Frijters, A.; Pot, J.; Peleman, J.; Kuiper, M. AFLP: A new technique for DNA fingerprinting. Nucleic Acids Res. 1995, 23, 4407–4414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hackett, C.A. TetraploidMap: Construction of a linkage map in autotetraploid species. J. Hered. 2003, 94, 358–359. [Google Scholar] [CrossRef] [Green Version]
- Hackett, C.A.; Milne, I.; Bradshaw, J.E.; Luo, Z. TetraploidMap for Windows: Linkage map construction and QTL mapping in autotetraploid species. J. Hered. 2007, 98, 727–729. [Google Scholar] [CrossRef] [PubMed]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DRAGEN Pipeline. Available online: https://www.illumina.com/products/by-type/informatics-products/dragen-bio-it-platform.html (accessed on 3 June 2021).
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyriakidou, M.; Anglin, N.L.; Ellis, D.; Tai, H.H.; Strömvik, M.V. Genome assembly of six polyploid potato genomes. Sci. Data 2020, 7, 88. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| LG | QTL | LOD Threshold (95%) | Max LOD Score | Position (cM) | 1 LOD Region (cM) | 2 LOD Region (cM) | Explained Variance (%) |
|---|---|---|---|---|---|---|---|
| 1A | 72292_2015_Ds_drym | 3.86 | 6.22 | 8 | 0–12 | 0–14 | 63.9 |
| 1A | 68015_2014_Ds_drym | 3.65 | 3.97 | 8 | 0–14 | 0–24 | 19.1 |
| 1A | 76354_2016_Ds_tuber_FW_kg_per_plant | 4.45 | 4.74 | 46 | 28–62 | 6–64 | 59.7 |
| 1A | 68015_2014_Co_starch_g_per_kg | 3.37 | 4.13 | 64 | 62–66 | 62–68 | 20.1 |
| 1A | 67518_2014_Co_starch_g_per_kg | 3.37 | 3.76 | 128 | 122–132 | 74–132 | 17.2 |
| 1A | 76354_2016_Ds_starch_g_per_kg | 4.49 | 5.48 | 94 | 86–132 | 84–132 | 79.4 |
| 1A | 67199_2014_Ds_tuber_FW_kg_per_plant | 3.54 | 5.69 | 124 | 118–132 | 114–132 | 25.8 |
| 1R | 76219_2016_Ds_drym | 3.84 | 4.29 | 84 | 4–40 | 2–46 | 43.0 |
| 1R | 67518_2014_Co_starch_yield_g_per_plant | 3.33 | 3.78 | 84 | 4–16 | 2–22 | 14.1 |
| 1R | 72292_2015_Ds_drym | 3.88 | 4.60 | 66 | 26–38 | 24–40 | 53.5 |
| 1R | 67518_2014_Co_tuber_FW_kg_per_plant | 3.27 | 3.42 | 84 | 2–16 | 2–30 | 12.4 |
| 2R | 76240_2016_Ds_drym | 3.87 | 4.01 | 56 | 42–74 | 34–80 | 52.6 |
| 2R | 76528_2016_Ds_starch_yield_g_per_plant | 4.06 | 4.36 | 88 | 48–96 | 28–100 | 50.3 |
| 2R | MW_Ds_tuber_FW_kg_per_plant | 4.34 | 4.17 | 48 | 36–100 | 32–106 | 58.9 |
| 2R | norm_Ds_starch_yield_g_per_plant | 4.99 | 4.75 | 58 | 40–74 | 36–82 | 58.5 |
| 2R | 67518_2014_Co_starch_yield_g_per_plant | 3.54 | 4.43 | 74 | 46–80 | 38–90 | 20.8 |
| 2R | 67518_2014_Co_tuber_FW_kg_per_plant | 3.31 | 4.23 | 58 | 46–82 | 38–100 | 20.4 |
| 2R | 76528_2016_Ds_drym | 7.27 | 8.01 | 88 | 52–94 | 42–102 | 74.4 |
| 2R | 76528_2016_Co_starch_yield_g_per_plant | 4.35 | 6.54 | 92 | 86–96 | 84–100 | 69.3 |
| 2R | 72275_2015_Co_tuber_FW_kg_per_plant | 4.76 | 4.77 | 88 | 84–94 | 84–100 | 52.7 |
| 2R | 72482_2015_Co_tuber_FW_kg_per_plant | 4.11 | 4.92 | 88 | 86–94 | 84–104 | 53.7 |
| 2R | 76528_2016_Co_tuber_FW_kg_per_plant | 4.55 | 6.08 | 92 | 86–96 | 86–102 | 66.2 |
| 2R | MW_Co_tuber_FW_kg_per_plant | 4.39 | 4.38 | 88 | 84–98 | 84–100 | 48.6 |
| 3A | 76354_2016_Ds_drym | 4.41 | 4.56 | 64 | 48–74 | 42–78 | 53.0 |
| 4A | 68015_2014_Ds_drym | 3.47 | 4.63 | 0 | 0–10 | 0–24 | 18.0 |
| 4A | 67199_2014_Ds_starch_g_per_kg | 3.50 | 3.83 | 8 | 0–12 | 0–28 | 13.8 |
| 4A | 67199_2014_Co_starch_g_per_kg | 3.26 | 5.71 | 8 | 0–20 | 0–32 | 23.9 |
| 4A | 68015_2014_Co_starch_g_per_kg | 3.42 | 3.97 | 0 | 0–18 | 0–40 | 15.2 |
| 4A | 68015_2014_Co_starch_yield_g_per_plant | 3.37 | 5.09 | 8 | 0–24 | 0–36 | 19.6 |
| 4A | MW_Co_starch_g_per_kg | 5.26 | 5.29 | 88 | 64–94 | 58–96 | 60.1 |
| 4A | 72482_2015_Co_starch_g_per_kg | 4.03 | 4.77 | 88 | 70–122 | 60–122 | 53.4 |
| 4R | 76529_2016_Ds_drym | 4.52 | 5.25 | 54 | 44–62 | 42–64 | 72.7 |
| 4R | 68015_2014_Co_starch_g_per_kg | 3.29 | 4.71 | 32 | 28–56 | 22–66 | 17.9 |
| 4R | 68015_2014_Co_tuber_FW_kg_per_plant | 3.47 | 5.35 | 28 | 20–52 | 16–58 | 22.3 |
| 4R | 67199_2014_Ds_tuber_FW_kg_per_plant | 3.57 | 3.94 | 0 | 0–2 | 0–4 | 14.0 |
| 4R | 76528_2016_Ds_tuber_FW_kg_per_plant | 4.23 | 5.59 | 4 | 0–6 | 0–8 | 54.5 |
| 6A | 72292_2015_Ds_drym | 3.99 | 4.07 | 36 | 20–44 | 0–52 | 40.0 |
| 6A | 68015_2014_Co_starch_g_per_kg | 3.29 | 3.47 | 14 | 0–22 | 0–22 | 14.8 |
| 6A | 76354_2016_Ds_drym | 4.23 | 5.26 | 66 | 60–80 | 58–88 | 56.8 |
| 6A | 76240_2016_Ds_starch_yield_g_per_plant | 3.86 | 4.22 | 92 | 70–92 | 64–92 | 41.6 |
| 6A | 76354_2016_Ds_tuber_FW_kg_per_plant | 3.90 | 4.66 | 46 | 46–86 | 40–92 | 51.9 |
| 6A | 76240_2016_Ds_tuber_FW_kg_per_plant | 3.86 | 3.90 | 90 | 68–92 | 62–92 | 39.0 |
| 7R | 67518_2014_Ds_drym | 3.32 | 3.62 | 48 | 3–49 | 3–51 | 14.0 |
| 7R | 67518_2014_Ds_starch_g_per_kg | 3.51 | 3.94 | 46 | 3–17 | 3–31 | 14.2 |
| 7R | 72247_2015_Ds_starch_g_per_kg | 4.07 | 5.33 | 48 | 3–31 | 3–39 | 49.1 |
| 7R | 72275_2015_Ds_starch_g_per_kg | 4.26 | 7.11 | 48 | 3–7 | 3–41 | 61.9 |
| 7R | MW_Ds_starch_g_per_kg | 4.45 | 4.55 | 30 | 17–37 | 3–45 | 45.2 |
| 7R | 72275_2015_Co_starch_g_per_kg | 4.41 | 6.72 | 48 | 3–19 | 3–33 | 59.6 |
| 7R | 76219_2016_Co_starch_g_per_kg | 4.05 | 4.22 | 42 | 3–35 | 3–45 | 42.0 |
| 7R | 72482_2015_Co_starch_g_per_kg | 4.08 | 4.27 | 48 | 3–43 | 3–47 | 40.8 |
| 7R | 68015_2014_Co_starch_g_per_kg | 3.44 | 4.28 | 8 | 25–51 | 3–51 | 16.4 |
| 7R | 76528_2016_Co_starch_g_per_kg | 3.75 | 3.90 | 24 | 15–47 | 3–51 | 42.5 |
| 7R | MW_Co_starch_g_per_kg | 5.16 | 4.60 | 48 | 3–37 | 3–49 | 42.8 |
| 7R | 72292_2015_Co_starch_yield_g_per_plant | 3.82 | 4.21 | 30 | 15–45 | 9–51 | 47.0 |
| 7R | 72482_2015_Ds_starch_yield_g_per_plant | 4.12 | 4.35 | 0 | 47–51 | 43–51 | 46.9 |
| 7R | 72482_2015_Ds_tuber_FW_kg_per_plant | 4.03 | 4.48 | 0 | 47–51 | 43–51 | 47.3 |
| 8R | 76528_2016_Ds_drym | 7.04 | 7.14 | 0 | 0–20 | 0–20 | 72.0 |
| 8R | 76219_2016_Ds_drym | 3.86 | 3.98 | 14 | 0–28 | 0–34 | 53.5 |
| 8R | norm_Ds_starch_yield_g_per_plant | 5.03 | 5.69 | 0 | 0–16 | 0–16 | 62.8 |
| 10R | 72247_2015_Ds_drym | 4.10 | 4.13 | 44 | 40–48 | 38–48 | 46.9 |
| 10R | 76219_2016_Ds_tuber_FW_kg_per_plant | 3.61 | 4.22 | 4 | 2–10 | 2–12 | 39.3 |
| 10R | 76240_2016_Ds_tuber_FW_kg_per_plant | 3.70 | 4.52 | 4 | 2–8 | 0–10 | 42.1 |
| 11R | 67518_2014_Ds_drym | 3.42 | 4.86 | 40 | 26–60 | 22–90 | 23.2 |
| 11R | 72292_2015_Ds_starch_g_per_kg | 4.22 | 5.60 | 90 | 82–90 | 82–90 | 57.5 |
| 11R | 76219_2016_Ds_starch_g_per_kg | 3.76 | 3.85 | 86 | 82–90 | 82–90 | 41.0 |
| 11R | 76240_2016_Ds_starch_g_per_kg | 4.08 | 5.53 | 84 | 82–90 | 82–90 | 55.6 |
| 11R | MW_Ds_starch_g_per_kg | 4.61 | 6.42 | 86 | 82–90 | 82–90 | 61.1 |
| 11R | MW_Co_starch_g_per_kg | 4.85 | 4.62 | 86 | 82–90 | 82–90 | 46.5 |
| 11R | 72247_2015_Co_tuber_FW_kg_per_plant | 3.85 | 4.08 | 90 | 86–90 | 84–90 | 42.2 |
| 11R | 72292_2015_Ds_tuber_FW_kg_per_plant | 4.00 | 4.57 | 90 | 84–90 | 82–90 | 49.2 |
| 11R | 76219_2016_Ds_tuber_FW_kg_per_plant | 4.03 | 4.83 | 86 | 82–90 | 82–90 | 50.8 |
| 12A | 76354_2016_Ds_drym | 4.41 | 5.46 | 66 | 3–9 | 3–11 | 58.3 |
| 12A | 72275_2015_Ds_drym | 3.73 | 3.95 | 64 | 3–23 | 3–33 | 39.9 |
| 12A | 76219_2016_Co_starch_g_per_kg | 4.31 | 4.36 | 64 | 3–9 | 3–17 | 50.4 |
| 12A | 72396_2015_Co_starch_g_per_kg | 3.62 | 4.37 | 64 | 3–11 | 3–19 | 47.4 |
| 12A | 76528_2016_Co_starch_g_per_kg | 3.84 | 4.18 | 64 | 3–11 | 3–23 | 43.8 |
| 12A | 76528_2016_Ds_starch_g_per_kg | 3.94 | 4.04 | 64 | 3–23 | 3–25 | 42.0 |
| 12A | 76219_2016_Ds_starch_g_per_kg | 3.51 | 3.98 | 66 | 3–23 | 3–25 | 42.7 |
| 12A | 68015_2014_Co_tuber_FW_kg_per_plant | 3.22 | 4.03 | 64 | 3–11 | 3–49 | 15.9 |
| 12R | 72292_2015_Ds_drym | 3.92 | 4.51 | 20 | 12–24 | 8–38 | 71.1 |
| 12R | 76219_2016_Ds_starch_g_per_kg | 3.84 | 4.41 | 94 | 72–94 | 68–94 | 56.8 |
| LG | QTL/ Marker Correlation | Position (cM) | Starch Candidate Gene | Location Phytozome Mb | Phytozome ID/ GenBank PGSC0003 | Annotation |
|---|---|---|---|---|---|---|
| 2R | 76240_2016_Ds_drym | 34–80 | ||||
| 76528_2016_Ds_drym | 42–102 | |||||
| E39M60_212 | 30.2 | SS4 | 30.14 | DMG400008322 | Starch synthase IV | |
| HRO_EREBP1_1_a2 | 56.7 | PFP-BETA | 36.84 | DMG400016726 | Pyrophosphate-fructose 6-phosphate 1-phosphotransferase subunit beta | |
| STI0036 | 76.6 | SS3 | 36.38 | DMG400016481 | Soluble starch synthase III; chloroplastic/amyloplastic | |
| STM5114y_b | 93.6 | PTST1 | 41.93 | DMG400030609 | Protein targeting to starch | |
| HRO_ACCS3_A_D | 100.6 | SS5 | 42.10 | DMG400030619 | Starch synthase V | |
| STI0024_d | 105.7 | |||||
| 7R | 67518_2014_Ds_drym | 3–51 | ||||
| STM0031_a_c | 6.3 | SPS | 3.89 | DMG400027936 | Sucrose-phosphate-synthase | |
| STI0033_2 | 6.5 | SUS II | 40.64 | DMG400013546 | Sucrose synthase 2 | |
| STI0025_2 | 7.9 | |||||
| 8R | 76528_2016_Ds_drym | 0–20 | ||||
| 76219_2016_Ds_drym | 0–34 | |||||
| HRO_MRP_ATF_3D_b | 0 | |||||
| STM1104 | 16.3 | WAXY | 56.8 | DMG400012111 | Granule-bound starch synthase | |
| E41M61_162 | 41.6 | |||||
| 11R | 67518_2014_Ds_drym | 22–90 | DBE | 3.95 | A52190.1 | De-branching enzyme |
| HRO_BSDRP4_5C_b | 18.0 | SEX4-like | 4.3 | DMG400027327 | Protein tyrosine phosphatase | |
| STM5130_a_d | 39.6 | SUT1 | 9.05 | DMG400009213 | Sucrose transport protein | |
| STI0028_1 | 65.2 | TAL1 | 19.47 | DMG402028027 | Transaldolase | |
| E41M61_85 | 92.5 | ANT | 34.62 | DMG400013596 | ADP; ATP carrier protein | |
| 12A | 76354_2016_Ds_drym | 3–11 | ||||
| 72275_2015_Ds_drym | 3–33 | |||||
| HRO_ETR1_1A_a_d | 0.0 | AGP | 1.22 | DMG400046891 | Glucose-1-phosphate adenylyltransferase | |
| HRO_EIX_1E_a | 8.4 | |||||
| HRO_JA2_1_B | 11.6 | |||||
| HRO_EBF1_2_b_2 | 24.7 | |||||
| E38M48_140 | 33.7 |
| LG | QTL/Marker Correlation | Position (cM) | Explained Variance in % | Drought Candidate Gene | Annotation | Phytozome PGSC0003 | Location Phytozome (Mb) |
|---|---|---|---|---|---|---|---|
| 1A | 72292_2015_Ds_drym | 0–14 | 63.9 | ||||
| 68015_2014_Ds_drym | 0–24 | 19.1 | |||||
| HRO_LIPOX_1B | 0 | LOX | Lipoxygenase | DMG400032207 | 2.15 | ||
| STI0034_b | 15.2 | FLA14 | Fasciclin-like arabinogalactan protein 14 | DMG400021372 | 2.66 | ||
| STI0043_c | 18.2 | Zinc finger protein | DMG400016379 | 3.5 | |||
| STI0043_b | 23 | Zinc finger protein | DMG400016379 | 3.5 | |||
| 1R | 76219_2016_Ds_drym | 2–46 | 43.0 | ||||
| 72292_2015_Ds_drym | 24–40 | 53.5 | |||||
| HRO_EIL2_1_b | 0 | EIL2 | Ethylene insensitive 3-like2 | DMG400008712 | 6.14 | ||
| STG0016_1_c | 44.6 | LHP1 | Chromo domain protein LHP1 | DMG400031112 | 67.23 | ||
| STG0016_2 | 51.1 | LHP1 | Chromo domain protein LHP1 | DMG400031112 | 67.23 | ||
| 2R | 76240_2016_Ds_drym | 34–80 | 52.6 | ||||
| 76528_2016_Ds_drym | 42–102 | 74.4 | |||||
| E39M60_212 | 30.2 | - | |||||
| HRO_EREBP1_1_a2 | 56.7 | EREBP1 | putative ethylene responsive element binding protein 1 | DMG400029713 | 33.63 | ||
| STI0036 | 76.6 | Transcriptional regulator family protein | DMG400028477 | 31.85 | |||
| STM5114y_b | 93.6 | Disease resistance response protein | DMG403001521 | 38.55 | |||
| HRO_ACCS3_A_D | 100.6 | ACS3 | 1-aminocyclopropane-1-carboxylate synthase 3 | DMG400021426 | 42.42 | ||
| STI0024_d | 105.7 | HRGP | Hydroxyproline-rich glycoprotein family protein | DMG400010074 | 44.53 | ||
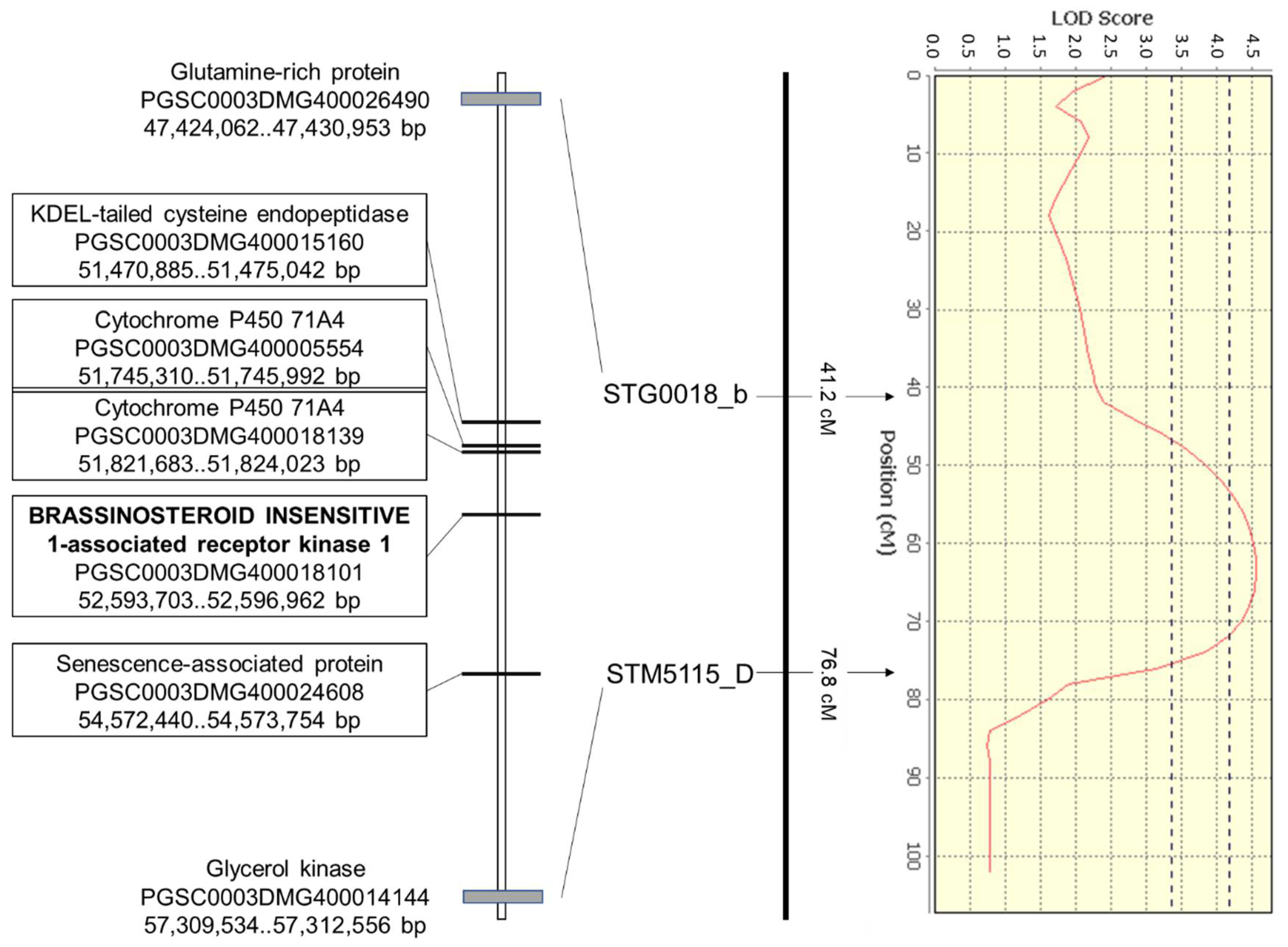
| 3A | 76354_2016_Ds_drym | 42–78 | 53 | ||||
| STG0018_b | 41.2 | Glutamine-rich protein | DMG400026490 | 47.43 | |||
| STM5115_D | 76.8 | GK | Glycerol kinase | DMG400014144 | 57.31 | ||
| 4R | 76529_2016_Ds_drym | 42–64 | 72.7 | ||||
| HRO_ALDH_H | 39.6 | ALDH | Aldehyde dehydrogenase family 7 member | DMG400034597 | 22.59 | ||
| STI0001_1_c | 73 | TSSR | Tuber-specific and sucrose- responsive element binding protein | DMG400007994 | 68.72 | ||
| 6A | 72292_2015_Ds_drym | 0–52 | 40.0 | ||||
| HRO_LEA_1_A_2 | 0 | LEA | Late embryogenesis abundant protein 5 | DMG400017936 | 0.46 | ||
| STI0021_2_c | 21.4 | HSFA6b | Heat stress transcription factor A-6b | DMG400016270 | 40.22 | ||
| STI0021_1 | 28.7 | HSFA6b | Heat stress transcription factor A-6b | DMG400016270 | 40.22 | ||
| STI0021_2_d | 36.4 | HSFA6b | Heat stress transcription factor A-6b | DMG400016270 | 40.22 | ||
| STM5126_1 | 58.1 | Conserved gene of unknown function | DMG400004051 | 50.92 | |||
| 6A | 76354_2016_Ds_drym | 58–88 | 56.8 | ||||
| STM5126_1 | 58.1 | Conserved gene of unknown function | DMG400004051 | 50.92 | |||
| STM5126_3 | 64.2 | Conserved gene of unknown function | DMG400004051 | 50.92 | |||
| HRO_BADH_2_c | 65.6 | BADH | Betaine aldehyde dehydrogenase | DMG400033028 | 52.13 | ||
| HRO_BADH_2_a | 72.1 | BADH | Betaine aldehyde dehydrogenase | DMG400033028 | 52.13 | ||
| STI004_2_a | 82.0 | Nucleic acid binding protein | DMG400003372 | 55.86 | |||
| STI004_1 | 94.2 | Nucleic acid binding protein | DMG400003372 | 55.86 | |||
| 7R | 67518_2014_Ds_drym | 3–51 | 14.4 | ||||
| STM0031_a_c | 6.3 | - | |||||
| STI0033_2 | 6.5 | HSFA9 | Heat stress transcription factor HSFA9 | DMG400032793 | 36.27 | ||
| STI0025_2 | 7.9 | - | |||||
| 8R | 76528_2016_Ds_drym | 0–20 | 72.0 | ||||
| 76219_2016_Ds_drym | 0–34 | 53.5 | |||||
| HRO_MRP_ATF_3D_b | 0 | MRP | Multidrug resistance protein ABC transporter | DMG400012167 | 55.53 | ||
| STM1104 | 16.3 | - | |||||
| E41M61_162 | 41.6 | - | |||||
| 10R | 72247_2015_Ds_drym | 38–48 | 46.9 | ||||
| STG0025 | 38.6 | Oxidoreductase/transition metal ion binding protein | DMG400028767 | 33.54 | |||
| 11R | 67518_2014_Ds_drym | 22–90 | 23.2 | ||||
| HRO_BSDRP4_5C_b | 18.0 | Bs4 | Bacterial spot disease resistance protein 4 | DMG400033334 | 37.64 | ||
| STM5130_a_d | 39.6 | SNRNP | U11/U12 small nuclear ribonucleoprotein | DMG400031069 | 3.78 | ||
| STI0028_1 | 65.2 | Conserved gene unknown function | DMG400007365 | 37.97 | |||
| E41M61_85 | 92.5 | - | |||||
| 12A | 76354_2016_Ds_drym | 3–11 | 58.3 | ||||
| 72275_2015_Ds_drym | 3–33 | 39.9 | |||||
| HRO_ETR1_1A_a_d | 0.0 | ETR1 | Ethylene receptor 1 | DMG400007843 | 1.11 | ||
| HRO_EIX_1E_a | 8.4 | EIX | Ethylene-inducing xylanase | DMG400007876 | 1.81 | ||
| HRO_JA2_1_B | 11.6 | JA2 | Jasmonic acid 2 | DMG400015342 | 0.82 | ||
| HRO_EBF1_2_b_2 | 24.7 | EBF1 | EIN3-binding F-box protein 1 | DMG400002914 | 2.85 | ||
| E38M48_140 | 33.7 | - | |||||
| 12R | 72292_2015_Ds_drym | 8–38 | 71.1 | ||||
| HRO_EBF1_2_a | 1.4 | EBF1 | EIN3-binding F-box protein 1 | DMG400002914 | 2.85 | ||
| STM5121_a | 25 | Conserved gene unknown function | DMG400000292 | 4.0 | |||
| STI0030_1_a | 37.9 | Conserved gene unknown function | DMG400014472 | 49.06 |
| Chromosome | Region | ID | Name |
|---|---|---|---|
| ST4.03ch03 | 49,754,402..49,754,857 | PGSC0003DMG400010135 | Ethylene-responsive element-binding family protein |
| ST4.03ch03 | 50,652,743..50,654,344 | PGSC0003DMG400015255 | DELLA protein RGL1 |
| ST4.03ch03 | 50,902,946..50,905,075 | PGSC0003DMG400015188 | Auxin-independent growth promoter |
| ST4.03ch03 | 51,989,958..51,991,673 | PGSC0003DMG400018128 | Protein phosphatase 2C |
| ST4.03ch03 | 52,593,703..52,596,962 | PGSC0003DMG400018101 | BAK1 |
| ST4.03ch03 | 52,736,187..52,740,429 | PGSC0003DMG400018153 | Gibberellin receptor GID1 |
| ST4.03ch03 | 53,228,956..53,233,993 | PGSC0003DMG400025330 | BAK1 |
| ST4.03ch03 | 54,533,650..54,534,364 | PGSC0003DMG400024606 | ERF transcription factor |
| ST4.03ch03 | 57,039,987..57,041,187 | PGSC0003DMG400014196 | Ethylene response factor |
| Chromosome | Region | ID | Name |
|---|---|---|---|
| ST4.03ch03 | 49,222,889..49,226,202 | PGSC0003DMG402010181 | Xyloglucan endotransglucosylase/hydrolase protein 9 |
| ST4.03ch03 | 51,432,584..51,434,561 | PGSC0003DMG400015230 | Pectate lyase |
| ST4.03ch03 | 52,023,650..52,024,174 | PGSC0003DMG400040957 | Pectinesterase inhibitor |
| ST4.03ch03 | 52,075,082..52,075,546 | PGSC0003DMG400034620 | Pectinesterase inhibitor |
| ST4.03ch03 | 52,079,987..52,080,508 | PGSC0003DMG400018189 | Pectinesterase inhibitor |
| ST4.03ch03 | 52,778,937..52,780,494 | PGSC0003DMG400018093 | Fasciclin-like arabinogalactan protein 9 |
| ST4.03ch03 | 52,906,784..52,909,093 | PGSC0003DMG400018146 | Pectinesterase |
| ST4.03ch03 | 53,068,010..53,070,815 | PGSC0003DMG400018142 | Pectate lyase |
| ST4.03ch03 | 54,961,544..54,963,584 | PGSC0003DMG400024530 | Protein COBRA |
| ST4.03ch03 | 54,963,884..54,966,403 | PGSC0003DMG400024628 | COBRA 3 |
| ST4.03ch03 | 55,316,389..55,316,718 | PGSC0003DMG400024646 | Expansin |
| ST4.03ch03 | 55,320,358..55,324,943 | PGSC0003DMG400024647 | Expansin |
| ST4.03ch03 | 55,333,818..55,338,034 | PGSC0003DMG400024648 | Expansin |
| ST4.03ch03 | 55,854,740..55,856,726 | PGSC0003DMG400019507 | Expansin |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schumacher, C.; Thümecke, S.; Schilling, F.; Köhl, K.; Kopka, J.; Sprenger, H.; Hincha, D.K.; Walther, D.; Seddig, S.; Peters, R.; et al. Genome-Wide Approach to Identify Quantitative Trait Loci for Drought Tolerance in Tetraploid Potato (Solanum tuberosum L.). Int. J. Mol. Sci. 2021, 22, 6123. https://doi.org/10.3390/ijms22116123
Schumacher C, Thümecke S, Schilling F, Köhl K, Kopka J, Sprenger H, Hincha DK, Walther D, Seddig S, Peters R, et al. Genome-Wide Approach to Identify Quantitative Trait Loci for Drought Tolerance in Tetraploid Potato (Solanum tuberosum L.). International Journal of Molecular Sciences. 2021; 22(11):6123. https://doi.org/10.3390/ijms22116123
Chicago/Turabian StyleSchumacher, Christina, Susanne Thümecke, Florian Schilling, Karin Köhl, Joachim Kopka, Heike Sprenger, Dirk Karl Hincha, Dirk Walther, Sylvia Seddig, Rolf Peters, and et al. 2021. "Genome-Wide Approach to Identify Quantitative Trait Loci for Drought Tolerance in Tetraploid Potato (Solanum tuberosum L.)" International Journal of Molecular Sciences 22, no. 11: 6123. https://doi.org/10.3390/ijms22116123
APA StyleSchumacher, C., Thümecke, S., Schilling, F., Köhl, K., Kopka, J., Sprenger, H., Hincha, D. K., Walther, D., Seddig, S., Peters, R., Zuther, E., Haas, M., & Horn, R. (2021). Genome-Wide Approach to Identify Quantitative Trait Loci for Drought Tolerance in Tetraploid Potato (Solanum tuberosum L.). International Journal of Molecular Sciences, 22(11), 6123. https://doi.org/10.3390/ijms22116123