
Silica Particles Trigger the Exopolysaccharide Production of Harsh Environment Isolates of Growth-Promoting Rhizobacteria and Increase Their Ability to Enhance Wheat Biomass in Drought-Stressed Soils


Abstract
:1. Introduction
2. Results
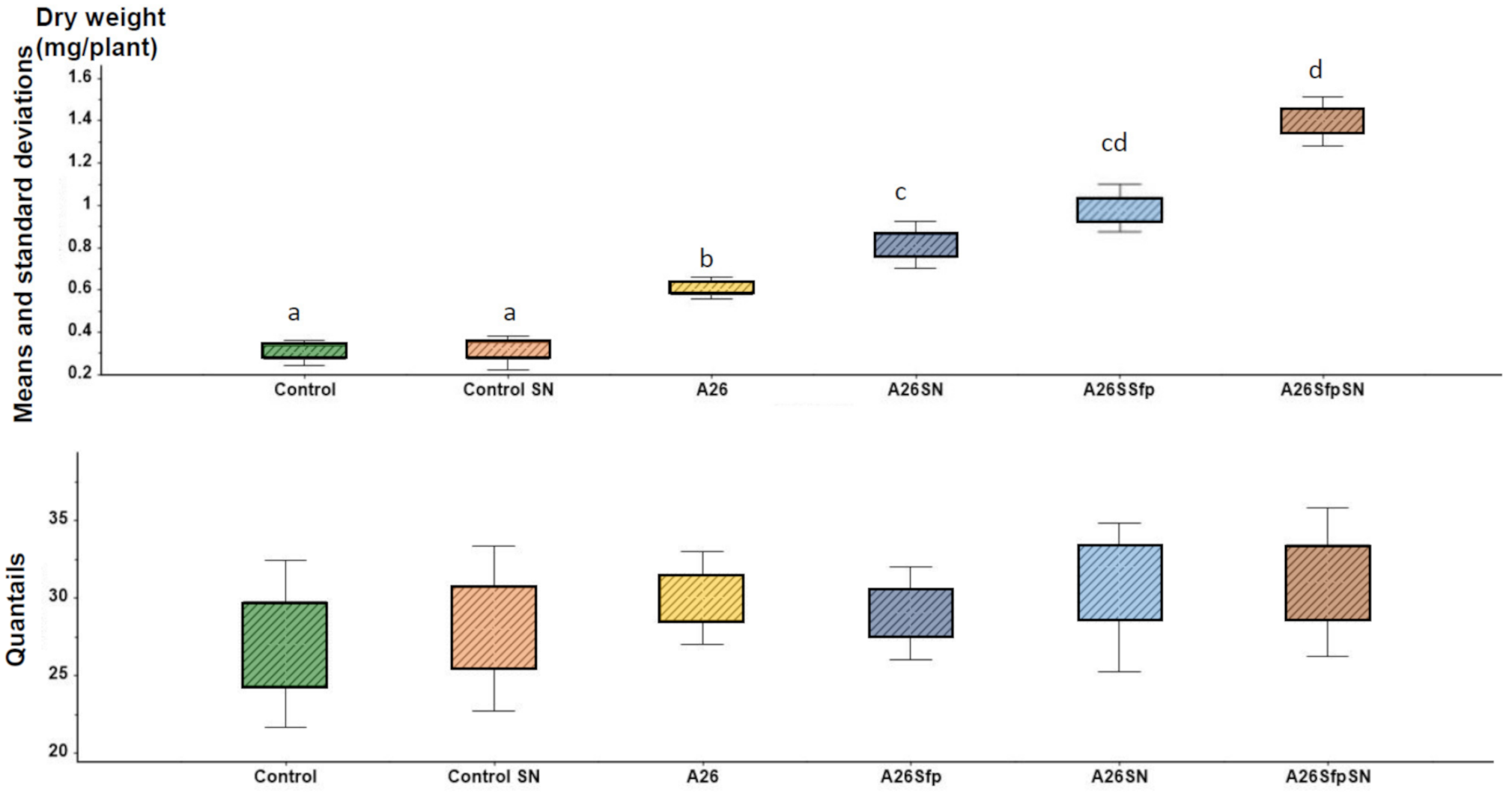
2.1. Ludox® TM50 Reassessment and Enhancement of Plant Biomass by PGPR with and without SN Treatment under Drought Stress
2.2. EPS Production and EPS D-GA Content of the PGPR Grown with and without SNs
2.2.1. Two Sets of Experiments Were Performed, and Both Were Culture Media Experiments
2.2.2. Hydroponic Culture Experiments
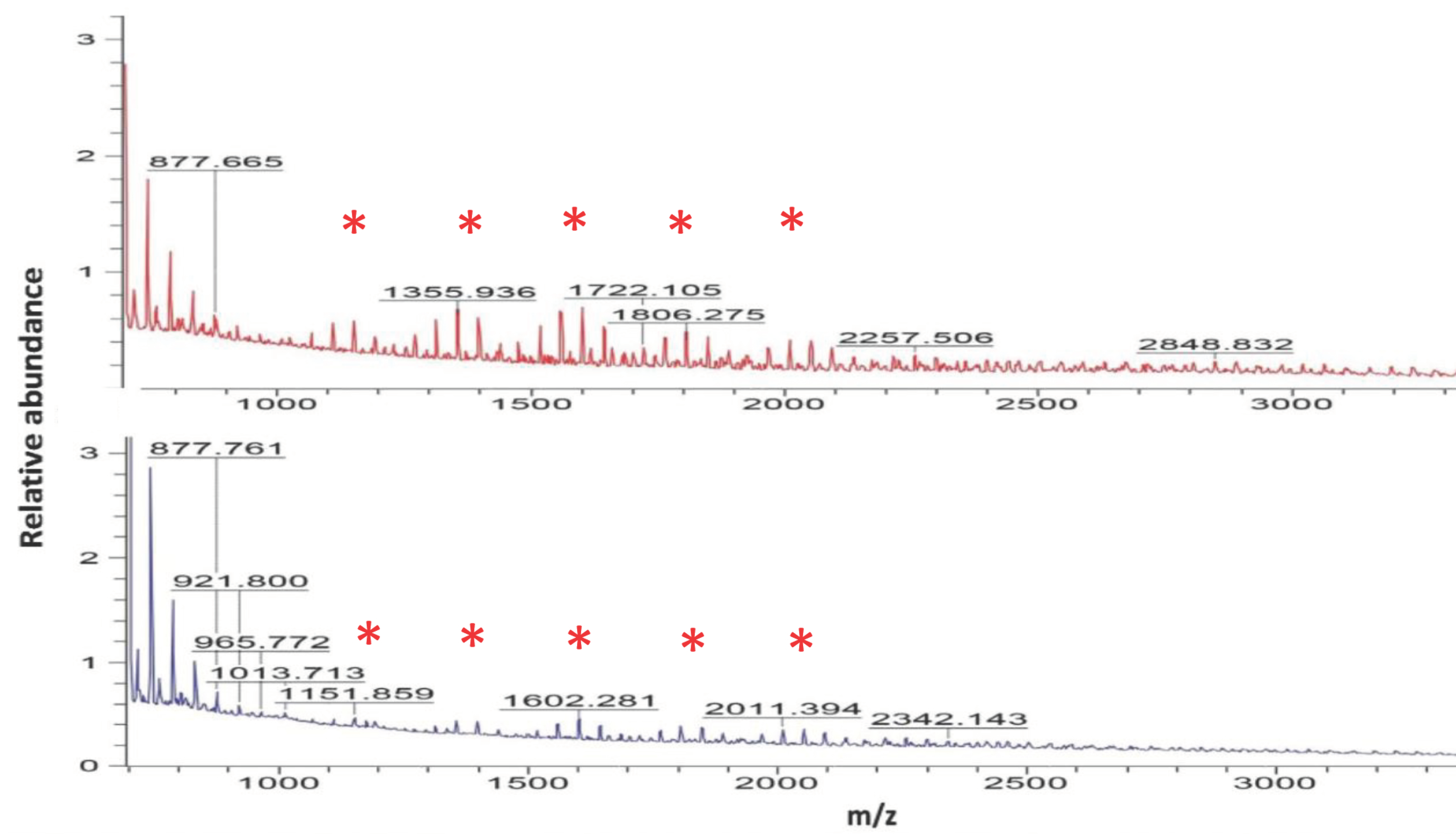
2.3. A26Sfp, A26SfpSN EPS MALDI Mass Spectrometry
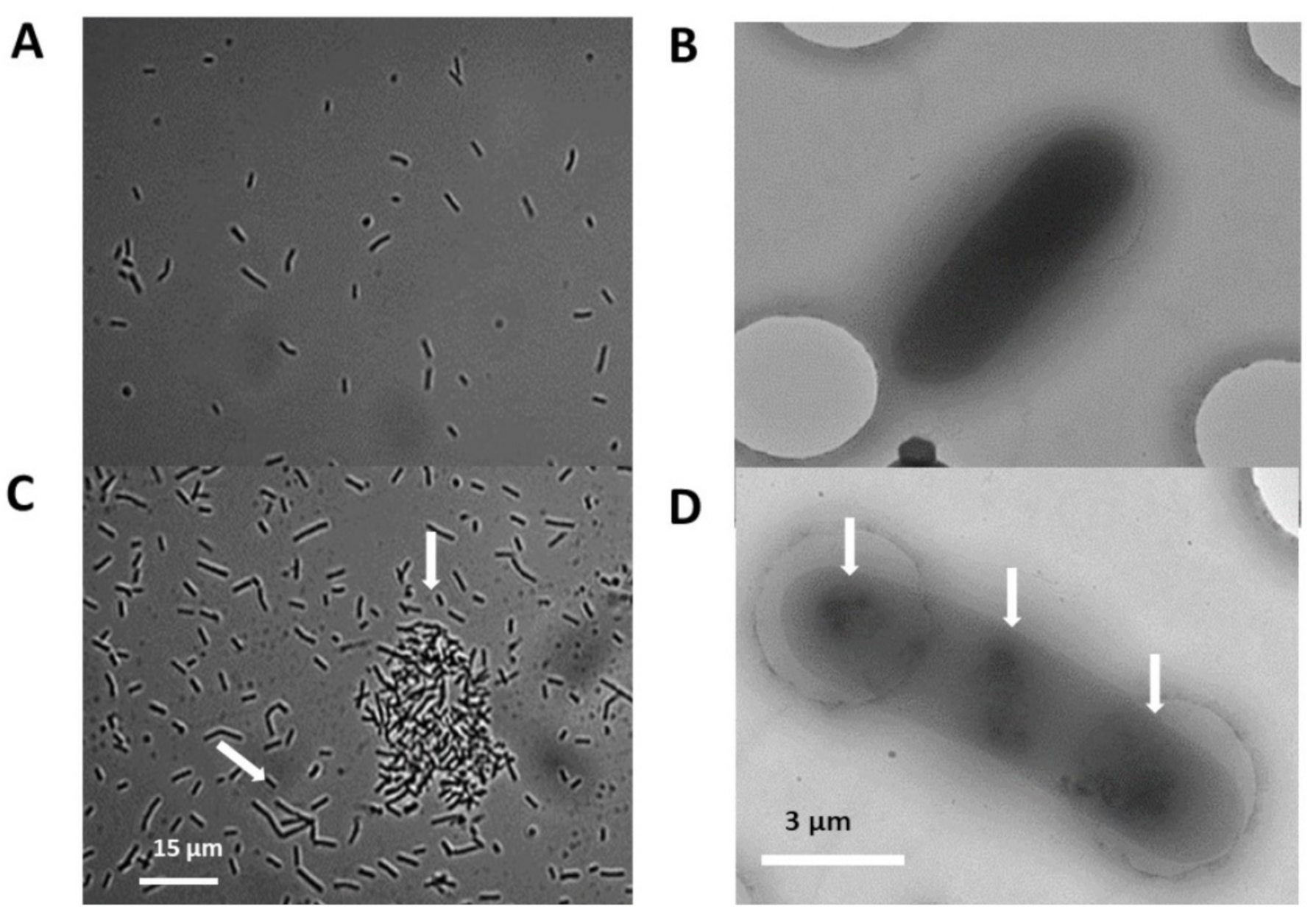
2.4. Silica Particles Induce Bacterial Elongation and Cell Aggregate Formation
2.5. EPS WHC and Osmotic Properties
3. Discussion
4. Material and Methods
4.1. Bacterial Growth and Culture Conditions
4.2. Bacterial Growth in the Presence of Nanoparticles
4.3. Plant Treatment
4.4. EPS and D-GA Content Evaluation
4.5. Light Microscopy
4.6. Cryo-Electron Microscopy
4.7. A26Sfp, A26SfpSN EPS MALDI Mass Spectrometry
4.8. Osmolarity Assay
4.9. Evaluation of A26 and A26Sfp EPS Water Holding Capacity (WHC)
4.10. Data Confirmation and Validation
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dai, A. Increasing drought under global warming in observations and models. Nat. Clim. Change 2012, 3, 52–58. [Google Scholar] [CrossRef]
- Xu, L.; Naylor, D.; Dong, Z.; Simmons, T.; Pierroz, G.; Hixson, K.K.; Kim, Y.M.; Zink, E.M.; Engbrecht, K.M.; Wang, Y.; et al. Drought delays development of the sorghum root microbiome and enriches for monoderm bacteria. Proc. Natl. Acad. Sci. USA 2018, 115, E4284–E4293. [Google Scholar] [CrossRef] [Green Version]
- Fitzpatrick, C.R.; Mustafa, Z.; Viliunas, J. Soil microbes alter plant fitness under competition and drought. J. Evol. Biol. 2019, 32, 438–450. [Google Scholar] [CrossRef] [PubMed]
- Timmusk, S.; Wagner, E.G.H. The Plant-Growth-Promoting Rhizobacterium Paenibacillus polymyxa Induces Changes in Arabidopsis thaliana Gene Expression: A Possible Connection Between Biotic and Abiotic Stress Responses. Mol. Plant-Microbe Interact. 1999, 12, 951–959. [Google Scholar] [CrossRef] [Green Version]
- Shimaila, A.; Glick, B.R. Plant-bacterial interactions in management of plant growth under abiotic stress. In New and Future Developments in Microbial Biotechnology and Bioendineering; Glick, B.R., Ed.; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Timmusk, S.; Copolovici, D.; Copolovici, L.; Teder, T.; Nevo, E.; Behers, L. Paenibacillus polymyxa biofilm polysaccharides antagonise Fusarium graminearum. Sci. Rep. 2019, 9, 662. [Google Scholar] [CrossRef] [PubMed]
- Sukweenadhi, J.; Kim, Y.-J.; Choi, E.-S.; Koh, S.-C.; Lee, S.-W.; Kim, Y.-J.; Yang, D.C. Paenibacillus yonginensis DCY84T induces changes in Arabidopsis thaliana gene expression against aluminum, drought, and salt stress. Microbiol. Res. 2015, 172, 7–15. [Google Scholar] [CrossRef]
- Naylor, D.; Coleman-Derr, D. Drought Stress and Root-Associated Bacterial Communities. Front. Plant Sci. 2018, 8, 2223. [Google Scholar] [CrossRef]
- Bandeppa, S.; Paul, S.; Thakur, J.K.; Chandrashekar, N.; Umesh, D.K.; Aggarwal, C.; Asha, A. Antioxidant, physiological and biochemical responses of drought susceptible and drought tolerant mustard (Brassica juncea L) genotypes to rhizobacterial inoculation under water deficit stress. Plant Physiol. Biochem. 2019, 143, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Zolla, G.; Badri, D.V.; Bakker, M.G.; Daniel, K.M.; Vivancoa, J.M. Soil microbiomes vary in their ability to confer drought tolerance to Arabidopsis. Appl. Soil Ecol. 2012, 68, 1–9. [Google Scholar] [CrossRef]
- Fitzpatrick, C.R.; Copeland, J.; Wang, P.W.; Guttman, D.S.; Kotanen, P.M.; Johnson, M.T.J. Assembly and ecological function of the root microbiome across angiosperm plant species. Proc. Natl. Acad. Sci. USA 2018, 115, E1157–E1165. [Google Scholar] [CrossRef] [Green Version]
- Liepe, K.J.; Hamann, A.; Smets, P.; Fitzpatrick, C.R.; Aitken, S.N. Adaptation of lodgepole pine and interior spruce to climate: Implications for reforestation in a warming world. Evol. Appl. 2016, 9, 409–419. [Google Scholar] [CrossRef] [Green Version]
- Timmusk, S.; Paalme, V.; Pavlicek, T.; Bergquist, J.; Vangala, A.; Danilas, T.; Nevo, E. Bacterial Distribution in the Rhizosphere of Wild Barley under Contrasting Microclimates. PLoS ONE 2011, 6, e17968. [Google Scholar] [CrossRef] [PubMed]
- Timmusk, S.; El-Daim, I.A.A.; Copolovici, L.; Tanilas, T.; Kännaste, A.; Behers, L.; Nevo, E.; Seisenbaeva, G.; Stenström, E.; Niinemets, U. Drought-Tolerance of Wheat Improved by Rhizosphere Bacteria from Harsh Environments: Enhanced Biomass Production and Reduced Emissions of Stress Volatiles. PLoS ONE 2014, 9, e96086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timmusk, S.; Kim, S.-B.; Nevo, E.; El Daim, I.A.; Ek, B.; Bergquist, J.; Behers, L. Sfp-type PPTase inactivation promotes bacterial biofilm formation and ability to enhance wheat drought tolerance. Front. Microbiol. 2015, 6, 387. [Google Scholar] [CrossRef] [Green Version]
- Timmusk, S.; Nevo, E.; Ayele, F.; Noe, S.; Niinemets, Y. Fighting Fusarium Pathogens in the Era of Climate Change: A Conceptual Approach. Pathogens 2020, 9, 419. [Google Scholar] [CrossRef] [PubMed]
- Timmusk, S.; Seisenbaeva, G.; Behers, L. Titania (TiO2) nanoparticles enhance the performance of growth-promoting rhizobacteria. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Timmusk, S.; Conrad, J.; Niinemets, Y.; Nevo, E.; Behers, L.; Bergqvist, J.; Noe, S. Managing Plant Stress in the Era of Climate Change: Realising the Global Sustainable Development Goals. 2020. Available online: http://www.global-engage.com/agricultural-biotechnology/managing-plant-stress-in-the-era-of-climate-change-realising-global-sustainable-development-goals/ (accessed on 7 June 2021).
- Nevo, E. “Evolution Canyon,” a potential microscale monitor of global warming across life. Proc. Natl. Acad. Sci. USA 2012, 109, 2960–2965. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, S.F. Developmental symbiosis facilitates the multiple origins of herbivory. Evol. Dev. 2020, 22, 154–164. [Google Scholar] [CrossRef] [Green Version]
- Rice, R.; Kallonen, A.; Cebra-Thomas, J.; Gilbert, S.F. Development of the turtle plastron, the order-defining skeletal structure. Proc. Natl. Acad. Sci. USA 2016, 113, 5317–5322. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, S.F. Developmental Plasticity and Developmental Symbiosis: The Return of Eco-Devo. Curr. Top. Dev. Biol. 2016, 116, 415–433. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, S.F.; Bosch, T.C.G.; Ledón-Rettig, C. Eco-Evo-Devo: Developmental symbiosis and developmental plasticity as evolutionary agents. Nat. Rev. Genet. 2015, 16, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Vega, N.M. Experimental evolution reveals microbial traits for association with the host gut. PLoS Biol. 2019, 17, e3000129. [Google Scholar] [CrossRef] [PubMed]
- Robinson, C.D.; Klein, H.S.; Murphy, K.D.; Parthasarathy, R.; Guillemin, K.; Bohannan, B.J.M. Experimental bacterial adaptation to the zebrafish gut reveals a primary role for immigration. PLoS Biol. 2018, 16, e2006893. [Google Scholar] [CrossRef] [Green Version]
- De-Bashan, L.E.; Trejo, A.; Huss, V.A.; Hernandez, J.; Bashan, Y. Chlorella sorokiniana UTEX 2805, a heat and intense, sunlight-tolerant microalga with potential for removing ammonium from wastewater. Bioresour. Technol. 2008, 99, 4980–4989. [Google Scholar] [CrossRef]
- Luyckx, M.; Hausman, J.-F.; Lutts, S.; Guerriero, G. Silicon and Plants: Current Knowledge and Technological Perspectives. Front. Plant Sci. 2017, 8, 411. [Google Scholar] [CrossRef] [Green Version]
- Rangaraj, S.; Gopalu, K.; Rathinam, Y.; Periasamy, P.; Venkatachalam, R.; Narayanasamy, K.; Suriyaprabha, R.; Karunakaran, G.; Yuvakkumar, R.; Prabu, P.; et al. Effect of silica nanoparticles on microbial biomass and silica availability in maize rhizosphere. Biotechnol. Appl. Biochem. 2014, 61, 668–675. [Google Scholar] [CrossRef] [PubMed]
- Karunakaran, G.; Suriyaprabha, R.; Manivasakan, P.; Yuvakkumar, R.; Rajendran, V.; Prabu, P.; Kannan, N. Effect of nanosilica and silicon sources on plant growth promoting rhizobacteria, soil nutrients and maize seed germination. IET Nanobiotechnol. 2013, 7, 70–77. [Google Scholar] [CrossRef]
- Various Synthetic Amorphous Silica; ECETOC JACC No 51; European Centre for Ecotoxicology and Toxicology for Chemicals: 2006. Available online: https://www.ecetoc.org/publication/jacc-report-51-synthetic-amorphous-silica/ (accessed on 7 June 2021).
- Orts-Gil, G.; Natte, K.; Drescher, D.; Bresch, H.; Mantion, A.; Kneipp, J.; Osterle, W. Characterization of silica nanoparticles prior to in vitro studies. J. Nanopart. Res. 2011, 13, 1593–1604. [Google Scholar] [CrossRef]
- El-Daim, I.A.; Haggblom, P.; Karlsson, M.; Stenstrom, E.; Timmusk, S. Paenibacillus polymyxa A26 Sfp-type PPTase inactivation limits bacterial antagonism against Fusarium graminearum but not of F. culmorum in kernel assay. Front. Plant. Sci. 2015, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Oberdörster, G.; Elder, A.; Gelein, R.; Mercer, P.; Biswas, P. Does nanoparticle activity depend upon size and crystal phase? Nanotoxicology 2008, 2, 33–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timmusk, S.; Behers, L. Nanotechnology, is it something useful for future agriculture? Atlas Sci. 2019. Available online: http://atlasofscience.org (accessed on 7 June 2021).
- Hristozov, D.; Pizzol, L.; Basei, G.; Zabeo, A.; Mackevica, A.; Hansen, S.F.; Gosens, I.; Cassee, F.R.; De Jong, W.; Koivisto, A.J.; et al. Quantitative human health risk assessment along the lifecycle of nano-scale copper-based wood preservatives. Nanotoxicology 2018, 12, 747–765. [Google Scholar] [CrossRef] [Green Version]
- Mojica, K.; Elsey, D.; Cooney, M.J. Quantitative analysis of biofilm EPS uronic acid content. J. Microbiol. Methods 2007, 71, 61–65. [Google Scholar] [CrossRef]
- Ludovic, L.; Dubessay, P.; Pierre, G.; Delattre, C.; Michaud, P. Biosourced Polysaccharide-Based Superabsorbents. Polysaccharides 2020, 10, 5. [Google Scholar] [CrossRef]
- Schmid, J.; Sieber, V.; Rehm, B.H.A. Bacterial exopolysaccharides: Biosynthesis pathways and engineering strategies. Front. Microbiol. 2015, 6, 496. [Google Scholar] [CrossRef] [Green Version]
- Liang, T.-W.; Wang, S.-L. Recent Advances in Exopolysaccharides from Paenibacillus spp.: Production, Isolation, Structure, and Bioactivities. Mar. Drugs 2015, 13, 1847. [Google Scholar] [CrossRef]
- Xu, X.; Bi, D.; Wu, X.; Wang, Q.; Wei, G.; Chi, L.; Jiang, Z.; Oda, T.; Wan, M. Unsaturated guluronate oligosaccharide enhances the antibacterial activities of macrophages. FASEB J. 2014, 28, 2645–2654. [Google Scholar] [CrossRef]
- López-García, M.; García, M.S.D.; Vilariño, J.M.L.; Rodríguez, M.V.G. MALDI-TOF to compare polysaccharide profiles from commercial health supplements of different mushroom species. Food Chem. 2016, 199, 597–604. [Google Scholar] [CrossRef]
- Kailemia, M.J.; Ruhaak, L.R.; Lebrilla, C.B.; Amster, I.J. Oligosaccharide Analysis by Mass Spectrometry: A Review of Recent Developments. Anal. Chem. 2014, 86, 196–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geoghegan, M.; Andrews, J.S.; Biggs, C.A.; Eboigbodin, K.; Elliott, D.R.; Rolfe, S.; Scholes, J.; Ojeda, J.J.; Romero-González, M.E.; Edyvean, R.G.J.; et al. The polymer physics and chemistry of microbial cell attachment and adhesion. Faraday Discuss. 2008, 139, 85–103. [Google Scholar] [CrossRef] [PubMed]
- Costa, O.Y.A.; Raaijmakers, J.M.; Kuramae, E.E. Microbial Extracellular Polymeric Substances: Ecological Function and Impact on Soil Aggregation. Front. Microbiol. 2018, 9, 1636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timmusk, S.; Zucca, C. The plant microbiome as a resource to increase crop productivity and soil resilience: A systems approach. J. Cameroon Acad. Sci. 2019, 14. [Google Scholar] [CrossRef] [Green Version]
- Douzou, P. Osmotic regulation of gene action. Proc. Natl. Acad. Sci. USA 1994, 91, 1657–1661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falghoush, A.; Beyenal, H.; Besser, T.; Osmland, A.; Call, D. Osmotic componds enhance antibiotic efficacy against Acinetobacter baumannii biofilm communities. Appl. Environ. Microbiol. 2017, 83, e9717. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Seminara, A.; Suaris, M.; Brenner, M.P.; Weitz, D.A.; Angelini, T.E. Nutrient depletion in Bacillus subtilis biofilms triggers matrix production. New J. Phys. 2014, 16, 015028. [Google Scholar] [CrossRef]
- Srinivasan, S.; Vladescu, I.D.; Koehler, S.A.; Wang, X.; Mani, M.; Rubinstein, S.M. Matrix production and sporulation in Bacilllus subtilis biofilms localize to propagating wave fronts. Biophys. J. 2018, 114, 1490–1498. [Google Scholar] [CrossRef] [Green Version]
- Morbach, S.; Krämer, R. Body Shaping under Water Stress: Osmosensing and Osmoregulation of Solute Transport in Bacteria. ChemBioChem 2002, 3, 384–397. [Google Scholar] [CrossRef]
- Timmusk, S.; Behers, L.; Muthoni, J.; Muraya, A.; Aronsson, A.-C. Perspectives and Challenges of Microbial Application for Crop Improvement. Front. Plant Sci. 2017, 8, 49. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-B.; Timmusk, S. A Simplified Method for Gene Knockout and Direct Screening of Recombinant Clones for Application in Paenibacillus polymyxa. PLoS ONE 2013, 8, e68092. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.; Wen, X.; Qiao, L.; Zhao, J.; Zhang, X.; Li, X.; Zhang, S.; Yang, Z.; Chang, Z.; Chen, J.; et al. A novel QTL QTrl.saw-2D.2 associated with the total root length identified by linkage and association analyses in wheat (Triticum aestivum L.). Planta 2019, 250, 129–143. [Google Scholar] [CrossRef]
- Rütering, M.; Schmid, J.; Rühmann, B.; Schilling, M.; Sieber, V. Controlled production of polysaccharides–exploiting nutrient supply for levan and heteropolysaccharide formation in Paenibacillus sp. Carbohydr. Polym. 2016, 148, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Niedermeyer, T.H.J.; Strohalm, M. mMass as a Software Tool for the Annotation of Cyclic Peptide Tandem Mass Spectra. PLoS ONE 2012, 7, e44913. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Time (Days) | Peat Soil | Hydroponic System | Bacterial Cell Culture |
|---|---|---|---|
| 1–5 | Seedlings growth | ||
| 5 | Inoculation with A26, A26SN, A26Sfp and A26SfpSN | EPS, D-GA assay | |
| 8 | Rhizosphere population assay | Rhizosphere population assay/ EPS, D-GA assay | |
| 8–15 | Drought stress treatment | ||
| 15 | Harvest/plant biomass analysis | ||
| Bacterial Population Log CFU/mL 1 | EPS (µg/mL) | D-GA (10−3 µg/mL) | |
|---|---|---|---|
| 1/2 TSB cultures | |||
| A26 | 9.00 ± 0.4 a | 11 ± 2 a | 0.3 ± 0.03 a |
| A26SN | 8.69 ± 0.37 a | 14 ± 2.1 b | 0.4 ± 0.04 b |
| A26Sfp | 9.0 ± 0.4 a | 14.5 ± 2.3 b | 0.7 ± 0.07 c |
| A26SfpSN | 8.89 ± 0.4 a | 18 ± 2.4 c | 0.9 ± 0.08 d |
| Hydroponic culture | |||
| A26 | 3.15 ± 0.15 b | 10 ± 1.5 a | 0.3 ± 0.07 a |
| A26SN | 3.09 ± 0.17 b | 14.8 ± 1.1 b | 0.5 ± 0.08 b |
| A26Sfp | 3.08 ± 0.13 b | 14.6 ± 1.3 b | 0.8 ± 0.8 c |
| A26SfpSN | 3.02 ± 0.15 b | 18 ± 1.9 c | 1.17 ± 0.2 d |
| Root wash Control | 1.19 ± 0.11 a | 0.08 ± 0.16 d | 0.05 ± 0.01 e |
| Peat soil | |||
| A26 | 3.02 ± 0.15 b | ND | ND |
| A26SN | 3.07 ± 0.13 b | ND | ND |
| A26Sfp | 3.09 ± 0.17 b | ND | ND |
| A26SfpSN | 3.11 ± 0.13 b | ND | ND |
| № | m/z | Intensity | Percentage Increase | |
|---|---|---|---|---|
| A26 | A26SN | |||
| 1. | 789 | 141,907.458 | 90,814.536 | 55 |
| 2. | 833 | 88,808.125 | 63,286.470 | 40 |
| 3. | 877 | 72,049.000 | 63,445.000 | 14 |
| 4. | 1109 | 45,547.000 | 56,170.000 | 19 |
| 5. | 1151 | 46,583.000 | 51,814.284 | 10 |
| 6. | 1194 | 43,345.000 | 44,568.000 | 3 |
| 7. | 1272 | 36,336.000 | 46,047.000 | 21 |
| 8. | 1356 | 44,012.000 | 60,857.115 | 28 |
| 9. | 1398 | 41,128.571 | 53,392.158 | 23 |
| 10. | 1440 | 35,220.000 | 38,714.000 | 9 |
| 11. | 1476 | 30,765.000 | 40,044.000 | 23 |
| 12. | 1518 | 34,532.000 | 48,457.729 | 29 |
| 13. | 1560 | 39,376.080 | 59,106.113 | 39 |
| 14. | 1602 | 41,044.232 | 59,972.852 | 32 |
| 15. | 1644 | 37,606.941 | 48,760.800 | 23 |
| 16. | 1806 | 32,269.259 | 38,433.907 | 16 |
| 17. | 1848 | 30,604.477 | 33,294.678 | 8 |
| 18. | 2052 | 27,555.650 | 32,217.000 | 14 |
| 19. | 2173 | 25,381.000 | 24,539.000 | 5 |
| 20. | 2215 | 26,874.000 | 28,241.000 | 5 |
| Fold Change | Strain | Osmolarity mOsm/kg | WHC% | EPS D-GA ** (E-03 µg/mL) | |
|---|---|---|---|---|---|
| Plant Dry Weight Improvement under Drought Stress * | 2 | A26 | 285 ± 13 a | 42 ± 3 a | 0.3 ± 0.07 a |
| 2.7 | A26SN | 299 ± 10 b | 48 ± 3 b | 0.5 ± 0.08 b | |
| 3.3 | A26Sfp | 325 ± 15 c | 59 ± 3 c | 0.8 ± 0.07 c | |
| 4.6 | A26SfpSN | 347 ± 13 d | 71 ± 3 d | 1.17 ± 0.2 d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fetsiukh, A.; Conrad, J.; Bergquist, J.; Timmusk, S. Silica Particles Trigger the Exopolysaccharide Production of Harsh Environment Isolates of Growth-Promoting Rhizobacteria and Increase Their Ability to Enhance Wheat Biomass in Drought-Stressed Soils. Int. J. Mol. Sci. 2021, 22, 6201. https://doi.org/10.3390/ijms22126201
Fetsiukh A, Conrad J, Bergquist J, Timmusk S. Silica Particles Trigger the Exopolysaccharide Production of Harsh Environment Isolates of Growth-Promoting Rhizobacteria and Increase Their Ability to Enhance Wheat Biomass in Drought-Stressed Soils. International Journal of Molecular Sciences. 2021; 22(12):6201. https://doi.org/10.3390/ijms22126201
Chicago/Turabian StyleFetsiukh, Anastasiia, Julian Conrad, Jonas Bergquist, and Salme Timmusk. 2021. "Silica Particles Trigger the Exopolysaccharide Production of Harsh Environment Isolates of Growth-Promoting Rhizobacteria and Increase Their Ability to Enhance Wheat Biomass in Drought-Stressed Soils" International Journal of Molecular Sciences 22, no. 12: 6201. https://doi.org/10.3390/ijms22126201
APA StyleFetsiukh, A., Conrad, J., Bergquist, J., & Timmusk, S. (2021). Silica Particles Trigger the Exopolysaccharide Production of Harsh Environment Isolates of Growth-Promoting Rhizobacteria and Increase Their Ability to Enhance Wheat Biomass in Drought-Stressed Soils. International Journal of Molecular Sciences, 22(12), 6201. https://doi.org/10.3390/ijms22126201