
Histamine: A Bacterial Signal Molecule

 ,
,  ,
,  , , and
, , and
Abstract
:1. Introduction
2. Histamine Sensing by Bacteria
2.1. Pseudomonas aeruginosa PAO1
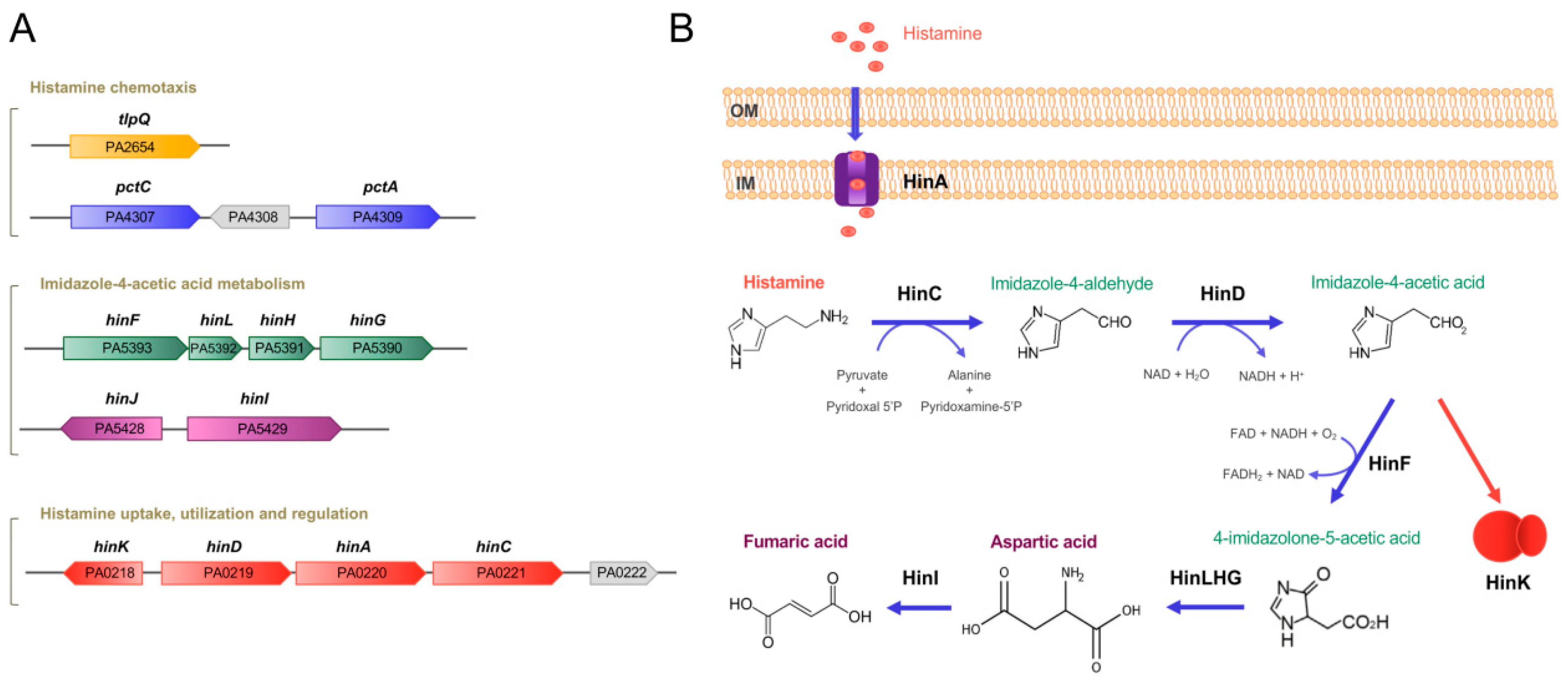
2.1.1. Transcriptional Responses to Histamine Exposure
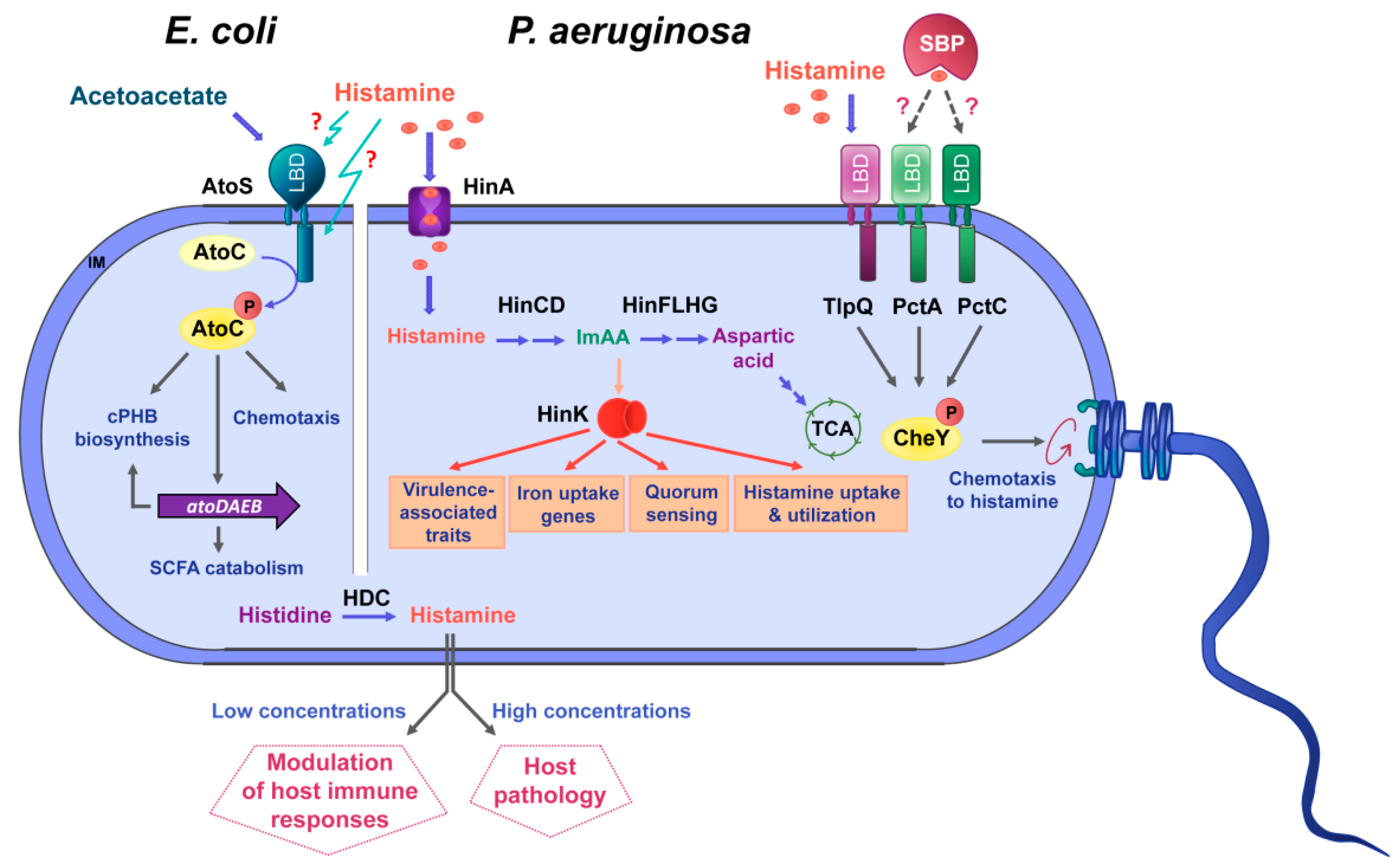
2.1.2. A Large Part of Histamine-Dependent Transcriptional Responses Are Mediated by the Transcriptional Regulator HinK
2.1.3. HinA Is a Histamine Transporter Permitting Histamine Uptake and Sensing by HinK
2.1.4. Histamine and HinK Regulate P. aeruginosa Virulence
2.1.5. Histamine Chemotaxis
2.1.6. The Chemoreceptor TlpQ Binds Histamine at its Ligand-Binding Domain with High Affinity
2.2. Escherichia coli
3. Histamine Release by Bacteria and Its Consequences
4. Outlook
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| cPHB | poly-(R)-3-hydroxybutyrate |
| DEC | diarrheagenic E. coli |
| ECF | extracytoplasmic function |
| HDC | histidine decarboxylase |
| IAA | indole-3-acetic acid |
| IBS | Irritable bowel syndrome |
| ImAA | imidazole-4-acetic acid |
| LBD | ligand binding domain |
| PQS | Pseudomonas quinolone signal |
| TCS | two-component system |
References
- Galperin, M.Y. What bacteria want. Environ. Microbiol. 2018, 20, 4221–4229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krell, T.; Lacal, J.; Busch, A.; Silva-Jimenez, H.; Guazzaroni, M.E.; Ramos, J.L. Bacterial sensor kinases: Diversity in the recognition of environmental signals. Annu. Rev. Microbiol. 2010, 64, 539–559. [Google Scholar] [CrossRef] [PubMed]
- Ortega, A.; Zhulin, I.B.; Krell, T. Sensory Repertoire of Bacterial Chemoreceptors. Microbiol. Mol. Biol. Rev. 2017, 81, e00033-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosier, A.; Bishnoi, U.; Lakshmanan, V.; Sherrier, D.J.; Bais, H.P. A perspective on inter-kingdom signaling in plant-beneficial microbe interactions. Plant Mol. Biol. 2016, 90, 537–548. [Google Scholar] [CrossRef] [PubMed]
- McFall-Ngai, M.; Hadfield, M.G.; Bosch, T.C.; Carey, H.V.; Domazet-Loso, T.; Douglas, A.E.; Dubilier, N.; Eberl, G.; Fukami, T.; Gilbert, S.F.; et al. Animals in a bacterial world, a new imperative for the life sciences. Proc. Natl. Acad. Sci. USA 2013, 110, 3229–3236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casanova-Saez, R.; Mateo-Bonmati, E.; Ljung, K. Auxin Metabolism in Plants. Cold Spring Harb. Perspect Biol. 2021, 13. [Google Scholar] [CrossRef]
- Matilla, M.A.; Daddaoua, A.; Chini, A.; Morel, B.; Krell, T. An auxin controls bacterial antibiotics production. Nucleic Acids Res. 2018, 46, 11229–11238. [Google Scholar] [CrossRef]
- Kunkel, B.N.; Johnson, J.M.B. Auxin Plays Multiple Roles during Plant-Pathogen Interactions. Cold Spring Harb. Perspect Biol. 2021. [Google Scholar] [CrossRef]
- Li, Q.; Ren, Y.; Fu, X. Inter-kingdom signaling between gut microbiota and their host. Cell. Mol. Life Sci. 2019, 76, 2383–2389. [Google Scholar] [CrossRef]
- Shahid, M.; Tripathi, T.; Sobia, F.; Moin, S.; Siddiqui, M.; Khan, R.A. Histamine, Histamine Receptors, and their Role in Immunomodulation: An Updated Systematic Review. Open Immunol. J. 2009, 2, 9–41. [Google Scholar] [CrossRef] [Green Version]
- Patel, R.H.; Mohiuddin, S.S. Biochemistry, Histamine. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Landete, J.M.; De las Rivas, B.; Marcobal, A.; Munoz, R. Updated molecular knowledge about histamine biosynthesis by bacteria. Crit. Rev. Food Sci. Nutr. 2008, 48, 697–714. [Google Scholar] [CrossRef] [Green Version]
- Bjornsdottir-Butler, K.; May, S.; Hayes, M.; Abraham, A.; Benner, R.A., Jr. Characterization of a novel enzyme from Photobacterium phosphoreum with histidine decarboxylase activity. Int. J. Food Microbiol. 2020, 334, 108815. [Google Scholar] [CrossRef] [PubMed]
- Landete, J.M.; Pardo, I.; Ferrer, S. Histamine, histidine, and growth-phase mediated regulation of the histidine decarboxylase gene in lactic acid bacteria isolated from wine. FEMS Microbiol. Lett. 2006, 260, 84–90. [Google Scholar] [CrossRef] [Green Version]
- Ferrario, C.; Borgo, F.; de Las Rivas, B.; Munoz, R.; Ricci, G.; Fortina, M.G. Sequencing, characterization, and gene expression analysis of the histidine decarboxylase gene cluster of Morganella morganii. Curr. Microbiol. 2014, 68, 404–411. [Google Scholar] [CrossRef]
- Diaz, M.; Del Rio, B.; Ladero, V.; Redruello, B.; Fernandez, M.; Martin, M.C.; Alvarez, M.A. Histamine production in Lactobacillus vaginalis improves cell survival at low pH by counteracting the acidification of the cytosol. Int. J. Food Microbiol. 2020, 321, 108548. [Google Scholar] [CrossRef] [PubMed]
- Ramos, J.L.; Timmis, K.N. Experimental evolution of catabolic pathways of bacteria. Microbiol. Sci. 1987, 4, 228–237. [Google Scholar] [PubMed]
- De la Torre, M.; Gomez-Botran, J.L.; Olivera, E.R.; Bermejo, F.; Rodriguez-Moran, J.; Luengo, J.M. Histamine catabolism in Pseudomonas putida U: Identification of the genes, catabolic enzymes and regulators. Environ. Microbiol. 2018, 20, 1828–1841. [Google Scholar] [CrossRef] [PubMed]
- Iglewski, B.H. Chapter 27 Pseudomonas. In Medical Microbiology, 4th ed.; Baron, S., Ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996. [Google Scholar]
- Juhas, M. Pseudomonas aeruginosa essentials: An update on investigation of essential genes. Microbiology 2015, 161, 2053–2060. [Google Scholar] [CrossRef] [PubMed]
- Horino, T.; Chiba, A.; Kawano, S.; Kato, T.; Sato, F.; Maruyama, Y.; Nakazawa, Y.; Yoshikawa, K.; Yoshida, M.; Hori, S. Clinical characteristics and risk factors for mortality in patients with bacteremia caused by Pseudomonas aeruginosa. Intern. Med. 2012, 51, 59–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Global priority list of antibiotic-resistant bacteria to guide research, discovery, and development of new antibiotics. 2017. Available online: http://www.who.int/medicines/publications/global-priority-list-antibiotic-resistant-bacteria/en/ (accessed on 1 June 2021).
- Corral-Lugo, A.; Matilla, M.A.; Martin-Mora, D.; Silva Jimenez, H.; Mesa Torres, N.; Kato, J.; Hida, A.; Oku, S.; Conejero-Muriel, M.; Gavira, J.A.; et al. High-Affinity Chemotaxis to Histamine Mediated by the TlpQ Chemoreceptor of the Human Pathogen Pseudomonas aeruginosa. mBio 2018, 9, e01894-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Cao, Q.; Cao, Q.; Gan, J.; Sun, N.; Yang, C.-G.; Bae, T.; Wuh, M.; Lan, L. Histamine activates HinK to promote the virulence of Pseudomonas aeruginosa. Sci. Bull. 2021, 66, 1101–1118. [Google Scholar] [CrossRef]
- Ghysels, B.; Ochsner, U.; Mollman, U.; Heinisch, L.; Vasil, M.; Cornelis, P.; Matthijs, S. The Pseudomonas aeruginosa pirA gene encodes a second receptor for ferrienterobactin and synthetic catecholate analogues. FEMS Microbiol. Lett. 2005, 246, 167–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillon, L.; Altenburger, S.; Graumann, P.L.; Schalk, I.J. Deciphering protein dynamics of the siderophore pyoverdine pathway in Pseudomonas aeruginosa. PloS ONE 2013, 8, e79111. [Google Scholar] [CrossRef]
- Llamas, M.A.; Sparrius, M.; Kloet, R.; Jimenez, C.R.; Vandenbroucke-Grauls, C.; Bitter, W. The heterologous siderophores ferrioxamine B and ferrichrome activate signaling pathways in Pseudomonas aeruginosa. J. Bacteriol. 2006, 188, 1882–1891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bastiaansen, K.C.; van Ulsen, P.; Wijtmans, M.; Bitter, W.; Llamas, M.A. Self-cleavage of the Pseudomonas aeruginosa Cell-surface Signaling Anti-sigma Factor FoxR Occurs through an N-O Acyl Rearrangement. J. Biol. Chem. 2015, 290, 12237–12246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otero-Asman, J.R.; Garcia-Garcia, A.I.; Civantos, C.; Quesada, J.M.; Llamas, M.A. Pseudomonas aeruginosa possesses three distinct systems for sensing and using the host molecule haem. Environ. Microbiol. 2019, 21, 4629–4647. [Google Scholar] [CrossRef] [PubMed]
- Ghysels, B.; Dieu, B.T.M.; Beatson, S.A.; Pirnay, J.P.; Ochsner, U.A.; Vasil, M.L.; Cornelis, P. FpvB, an alternative type I ferripyoverdine receptor of Pseudomonas aeruginosa. Microbiology (Reading) 2004, 150, 1671–1680. [Google Scholar] [CrossRef] [Green Version]
- Cunrath, O.; Gasser, V.; Hoegy, F.; Reimmann, C.; Guillon, L.; Schalk, I.J. A cell biological view of the siderophore pyochelin iron uptake pathway in Pseudomonas aeruginosa. Environ. Microbiol. 2015, 17, 171–185. [Google Scholar] [CrossRef]
- Adhikari, P.; Kirby, S.D.; Nowalk, A.J.; Veraldi, K.L.; Schryvers, A.B.; Mietzner, T.A. Biochemical characterization of a Haemophilus influenzae periplasmic iron transport operon. J. Biol. Chem. 1995, 270, 25142–25149. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Reyes, S.; Soberon-Chavez, G.; Cocotl-Yanez, M. The third quorum-sensing system of Pseudomonas aeruginosa: Pseudomonas quinolone signal and the enigmatic PqsE protein. J. Med. Microbiol. 2020, 69, 25–34. [Google Scholar] [CrossRef]
- Gallagher, L.A.; McKnight, S.L.; Kuznetsova, M.S.; Pesci, E.C.; Manoil, C. Functions required for extracellular quinolone signaling by Pseudomonas aeruginosa. J. Bacteriol. 2002, 184, 6472–6480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinaud, M.; Chabert, J.; Faudry, E.; Neumann, E.; Lemaire, D.; Pastor, A.; Elsen, S.; Dessen, A.; Attree, I. The PscE-PscF-PscG complex controls type III secretion needle biogenesis in Pseudomonas aeruginosa. J. Biol. Chem. 2005, 280, 36293–36300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wozniak, D.J.; Cram, D.C.; Daniels, C.J.; Galloway, D.R. Nucleotide sequence and characterization of toxR: A gene involved in exotoxin A regulation in Pseudomonas aeruginosa. Nucleic Acids Res. 1987, 15, 2123–2135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Jin, S. PtrB of Pseudomonas aeruginosa suppresses the type III secretion system under the stress of DNA damage. J. Bacteriol. 2005, 187, 6058–6068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rampioni, G.; Schuster, M.; Greenberg, E.P.; Zennaro, E.; Leoni, L. Contribution of the RsaL global regulator to Pseudomonas aeruginosa virulence and biofilm formation. FEMS Microbiol. Lett. 2009, 301, 210–217. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.H.; Zhang, X.F.; Xu, J.L.; Tan, A.T.; Zhang, L.H. VqsM, a novel AraC-type global regulator of quorum-sensing signalling and virulence in Pseudomonas aeruginosa. Mol. Microbiol. 2005, 58, 552–564. [Google Scholar] [CrossRef]
- Dean, C.R.; Neshat, S.; Poole, K. PfeR, an enterobactin-responsive activator of ferric enterobactin receptor gene expression in Pseudomonas aeruginosa. J. Bacteriol. 1996, 178, 5361–5369. [Google Scholar] [CrossRef] [Green Version]
- Shen, D.K.; Filopon, D.; Kuhn, L.; Polack, B.; Toussaint, B. PsrA is a positive transcriptional regulator of the type III secretion system in Pseudomonas aeruginosa. Infect. Immun. 2006, 74, 1121–1129. [Google Scholar] [CrossRef] [Green Version]
- Williams McMackin, E.A.; Marsden, A.E.; Yahr, T.L. H-NS Family Members MvaT and MvaU Regulate the Pseudomonas aeruginosa Type III Secretion System. J. Bacteriol. 2019, 201. [Google Scholar] [CrossRef] [Green Version]
- Alford, M.A.; Baghela, A.; Yeung, A.T.Y.; Pletzer, D.; Hancock, R.E.W. NtrBC Regulates Invasiveness and Virulence of Pseudomonas aeruginosa During High-Density Infection. Front. Microbiol. 2020, 11, 773. [Google Scholar] [CrossRef]
- Zeng, B.; Wang, C.; Zhang, P.; Guo, Z.; Chen, L.; Duan, K. Heat Shock Protein DnaJ in Pseudomonas aeruginosa Affects Biofilm Formation via Pyocyanin Production. Microorganisms 2020, 8, 395. [Google Scholar] [CrossRef] [Green Version]
- Okuda, J.; Yamane, S.; Nagata, S.; Kunikata, C.; Suezawa, C.; Yasuda, M. The Pseudomonas aeruginosa dnaK gene is involved in bacterial translocation across the intestinal epithelial cell barrier. Microbiology (Reading) 2017, 163, 1208–1216. [Google Scholar] [CrossRef]
- Matilla, M.A.; Martin-Mora, D.; Gavira, J.A.; Krell, T. Pseudomonas aeruginosa as a Model To Study Chemosensory Pathway Signaling. Microbiol. Mol. Biol. Rev. 2021, 85. [Google Scholar] [CrossRef] [PubMed]
- Garvis, S.; Munder, A.; Ball, G.; de Bentzmann, S.; Wiehlmann, L.; Ewbank, J.J.; Tummler, B.; Filloux, A. Caenorhabditis elegans semi-automated liquid screen reveals a specialized role for the chemotaxis gene cheB2 in Pseudomonas aeruginosa virulence. PLoS Pathog. 2009, 5, e1000540. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.H.; Zhang, X.F.; Soo, H.M.; Greenberg, E.P.; Zhang, L.H. The two-component response regulator PprB modulates quorum-sensing signal production and global gene expression in Pseudomonas aeruginosa. Mol. Microbiol. 2005, 56, 1287–1301. [Google Scholar] [CrossRef]
- Bardy, S.L.; Maddock, J.R. Polar localization of a soluble methyl-accepting protein of Pseudomonas aeruginosa. J. Bacteriol. 2005, 187, 7840–7844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rico-Jimenez, M.; Reyes-Darias, J.A.; Ortega, A.; Diez Pena, A.I.; Morel, B.; Krell, T. Two different mechanisms mediate chemotaxis to inorganic phosphate in Pseudomonas aeruginosa. Sci. Rep. 2016, 6, 28967. [Google Scholar] [CrossRef] [PubMed]
- Zaborin, A.; Romanowski, K.; Gerdes, S.; Holbrook, C.; Lepine, F.; Long, J.; Poroyko, V.; Diggle, S.P.; Wilke, A.; Righetti, K.; et al. Red death in Caenorhabditis elegans caused by Pseudomonas aeruginosa PAO1. Proc. Natl. Acad. Sci. USA 2009, 106, 6327–6332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Bentzmann, S.; Aurouze, M.; Ball, G.; Filloux, A. FppA, a novel Pseudomonas aeruginosa prepilin peptidase involved in assembly of type IVb pili. J. Bacteriol. 2006, 188, 4851–4860. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.S.; Heo, Y.J.; Lee, J.K.; Cho, Y.H. KatA, the major catalase, is critical for osmoprotection and virulence in Pseudomonas aeruginosa PA14. Infect. Immun. 2005, 73, 4399–4403. [Google Scholar] [CrossRef] [Green Version]
- Scheepers, G.H.; Lycklama, A.N.J.A.; Poolman, B. An updated structural classification of substrate-binding proteins. FEBS. Lett. 2016, 590, 4393–4401. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, M.; Rico-Jimenez, M.; Ortega, A.; Daddaoua, A.; Garcia Garcia, A.I.; Martin-Mora, D.; Torres, N.M.; Tajuelo, A.; Matilla, M.A.; Krell, T. Determination of Ligand Profiles for Pseudomonas aeruginosa Solute Binding Proteins. Int. J. Mol. Sci. 2019, 20, 5156. [Google Scholar] [CrossRef] [Green Version]
- Ortega, D.R.; Fleetwood, A.D.; Krell, T.; Harwood, C.S.; Jensen, G.J.; Zhulin, I.B. Assigning chemoreceptors to chemosensory pathways in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2017, 114, 12809–12814. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Kato, J.; Kuroda, A.; Ikeda, T.; Takiguchi, N.; Ohtake, H. Identification and characterization of two chemotactic transducers for inorganic phosphate in Pseudomonas aeruginosa. J. Bacteriol. 2000, 182, 3400–3404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bains, M.; Fernandez, L.; Hancock, R.E. Phosphate starvation promotes swarming motility and cytotoxicity of Pseudomonas aeruginosa. Appl. Environ. Microbiol. 2012, 78, 6762–6768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gavira, J.A.; Gumerov, V.M.; Rico-Jimenez, M.; Petukh, M.; Upadhyay, A.A.; Ortega, A.; Matilla, M.A.; Zhulin, I.B.; Krell, T. How Bacterial Chemoreceptors Evolve Novel Ligand Specificities. mBio 2020, 11, e03066-19. [Google Scholar] [CrossRef] [Green Version]
- Rico-Jimenez, M.; Munoz-Martinez, F.; Garcia-Fontana, C.; Fernandez, M.; Morel, B.; Ortega, A.; Ramos, J.L.; Krell, T. Paralogous chemoreceptors mediate chemotaxis towards protein amino acids and the non-protein amino acid gamma-aminobutyrate (GABA). Mol. Microbiol. 2013, 88, 1230–1243. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, K.; Fukutomi, H.; Kuroda, A.; Kato, J.; Ohtake, H. Genetic identification of chemotactic transducers for amino acids in Pseudomonas aeruginosa. Microbiology 1997, 143 (Pt 10), 3223–3229. [Google Scholar] [CrossRef] [Green Version]
- Upadhyay, A.A.; Fleetwood, A.D.; Adebali, O.; Finn, R.D.; Zhulin, I.B. Cache Domains That are Homologous to, but Different from PAS Domains Comprise the Largest Superfamily of Extracellular Sensors in Prokaryotes. PLoS Comput. Biol. 2016, 12, e1004862. [Google Scholar] [CrossRef] [Green Version]
- Matilla, M.A.; Ortega, A.; Krell, T. The role of solute binding proteins in signal transduction. Comput. Struct. Biotechnol. J. 2021, 19, 1786–1805. [Google Scholar] [CrossRef]
- Matilla, M.A.; Mora, D.M.; Krell, T. The use of Isothermal Titration Calorimetry to unravel chemotactic signaling mechanisms. Environ. Microbiol. 2020, 22, 3005–3019. [Google Scholar] [CrossRef] [PubMed]
- Gavira, J.A.; Ortega, A.; Martin-Mora, D.; Conejero-Muriel, M.T.; Corral-Lugo, A.; Morel, B.; Matilla, M.A.; Krell, T. Structural Basis for Polyamine Binding at the dCACHE Domain of the McpU Chemoreceptor from Pseudomonas putida. J. Mol. Biol. 2018, 430, 1950–1963. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, M.; Compton, K.K.; Mancl, J.M.; Webb, B.A.; Brown, A.M.; Scharf, B.E.; Schubot, F.D. Structure of the sensory domain of McpX from Sinorhizobium meliloti, the first known bacterial chemotactic sensor for quaternary ammonium compounds. Biochem. J. 2018, 475, 3949–3962. [Google Scholar] [CrossRef]
- Nishiyama, S.; Takahashi, Y.; Yamamoto, K.; Suzuki, D.; Itoh, Y.; Sumita, K.; Uchida, Y.; Homma, M.; Imada, K.; Kawagishi, I. Identification of a Vibrio cholerae chemoreceptor that senses taurine and amino acids as attractants. Sci. Rep. 2016, 6, 20866. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.F.; Machuca, M.A.; Rahman, M.M.; Koc, C.; Norton, R.S.; Smith, B.J.; Roujeinikova, A. Structure-Activity Relationship Study Reveals the Molecular Basis for Specific Sensing of Hydrophobic Amino Acids by the Campylobacter jejuni Chemoreceptor Tlp3. Biomolecules 2020, 10, 744. [Google Scholar] [CrossRef]
- Xia, R.; Wang, N.; Xu, Z.; Lu, Y.; Song, J.; Zhang, A.; Guo, C.; He, Y. Cryo-EM structure of the human histamine H1 receptor/Gq complex. Nat. Comm. 2021, 12, 2086. [Google Scholar] [CrossRef] [PubMed]
- Nakane, T.; Kotecha, A.; Sente, A.; McMullan, G.; Masiulis, S.; Brown, P.; Grigoras, I.T.; Malinauskaite, L.; Malinauskas, T.; Miehling, J.; et al. Single-particle cryo-EM at atomic resolution. Nature 2020, 587, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Jubb, H.C.; Higueruelo, A.P.; Ochoa-Montaño, B.; Pitt, W.R.; Ascher, D.B.; Blundell, T.L. Arpeggio: A Web Server for Calculating and Visualising Interatomic Interactions in Protein Structures. J. Mol. Biol. 2017, 429, 365–371. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef] [PubMed]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J. The Phyre2 web portal for protein modeling, prediction and analysis. Nature Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [Green Version]
- Kyriakidis, D.A.; Tiligada, E. Signal transduction and adaptive regulation through bacterial two-component systems: The Escherichia coli AtoSC paradigm. Amino Acids 2009, 37, 443–458. [Google Scholar] [CrossRef]
- Jenkins, L.S.; Nunn, W.D. Genetic and molecular characterization of the genes involved in short-chain fatty acid degradation in Escherichia coli: The ato system. J. Bacteriol. 1987, 169, 42–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lioliou, E.E.; Mimitou, E.P.; Grigoroudis, A.I.; Panagiotidis, C.H.; Panagiotidis, C.A.; Kyriakidis, D.A. Phosphorylation activity of the response regulator of the two-component signal transduction system AtoS-AtoC in E. coli. Biochim. Biophys. Acta 2005, 1725, 257–268. [Google Scholar] [CrossRef]
- Jenkins, L.S.; Nunn, W.D. Regulation of the ato operon by the atoC gene in Escherichia coli. J. Bacteriol. 1987, 169, 2096–2102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barcik, W.; Wawrzyniak, M.; Akdis, C.A.; O’Mahony, L. Immune regulation by histamine and histamine-secreting bacteria. Curr. Opin. Immunol. 2017, 48, 108–113. [Google Scholar] [CrossRef]
- Thorburn, A.N.; Macia, L.; Mackay, C.R. Diet, metabolites, and “western-lifestyle” inflammatory diseases. Immunity 2014, 40, 833–842. [Google Scholar] [CrossRef] [Green Version]
- Theodorou, M.C.; Panagiotidis, C.A.; Panagiotidis, C.H.; Pantazaki, A.A.; Kyriakidis, D.A. Involvement of the AtoS-AtoC signal transduction system in poly-(R)-3-hydroxybutyrate biosynthesis in Escherichia coli. Biochim. Biophys. Acta 2006, 1760, 896–906. [Google Scholar] [CrossRef]
- Theodorou, M.C.; Theodorou, E.C.; Kyriakidis, D.A. Involvement of AtoSC two-component system in Escherichia coli flagellar regulon. Amino Acids 2012, 43, 833–844. [Google Scholar] [CrossRef] [PubMed]
- Theodorou, M.C.; Theodorou, E.C.; Panagiotidis, C.A.; Kyriakidis, D.A. Spermidine triggering effect to the signal transduction through the AtoS-AtoC/Az two-component system in Escherichia coli. Biochim. Biophys. Acta 2007, 1770, 1104–1114. [Google Scholar] [CrossRef]
- Theodorou, E.C.; Theodorou, M.C.; Kyriakidis, D.A. Regulation of poly-(R)-(3-hydroxybutyrate-co-3-hydroxyvalerate) biosynthesis by the AtoSCDAEB regulon in phaCAB+ Escherichia coli. Appl. Microbiol. Biotechnol. 2013, 97, 5259–5274. [Google Scholar] [CrossRef] [PubMed]
- Theodorou, M.C.; Tiligada, E.; Kyriakidis, D.A. Extracellular Ca2+ transients affect poly-(R)-3-hydroxybutyrate regulation by the AtoS-AtoC system in Escherichia coli. Biochem. J. 2009, 417, 667–672. [Google Scholar] [CrossRef] [Green Version]
- Theodorou, M.C.; Kyriakidis, D.A. Calcium channels blockers inhibit the signal transduction through the AtoSC system in Escherichia coli. Eur. J. Pharm. Sci. 2012, 47, 84–96. [Google Scholar] [CrossRef]
- Kyriakidis, D.A.; Theodorou, M.C.; Filippou, P.S.; Kyriakidis, K.D.; Tiligada, E. Effect of histamine on the signal transduction of the AtoS-AtoC two component system and involvement in poly-(R)-3-hydroxybutyrate biosynthesis in Escherichia coli. Amino Acids 2008, 35, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Visciano, P.; Schirone, M.; Paparella, A. An Overview of Histamine and Other Biogenic Amines in Fish and Fish Products. Foods 2020, 9, 1795. [Google Scholar] [CrossRef] [PubMed]
- Moniente, M.; Garcia-Gonzalo, D.; Ontanon, I.; Pagan, R.; Botello-Morte, L. Histamine accumulation in dairy products: Microbial causes, techniques for the detection of histamine-producing microbiota, and potential solutions. Compr. Rev. Food Sci. Food Saf. 2021, 20, 1481–1523. [Google Scholar] [CrossRef]
- Rossi, F.; Gardini, F.; Rizzotti, L.; La Gioia, F.; Tabanelli, G.; Torriani, S. Quantitative analysis of histidine decarboxylase gene (hdcA) transcription and histamine production by Streptococcus thermophilus PRI60 under conditions relevant to cheese making. Appl. Environ. Microbiol. 2011, 77, 2817–2822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frei, R.; Ferstl, R.; Konieczna, P.; Ziegler, M.; Simon, T.; Rugeles, T.M.; Mailand, S.; Watanabe, T.; Lauener, R.; Akdis, C.A.; et al. Histamine receptor 2 modifies dendritic cell responses to microbial ligands. J. Allergy Clin. Immunol. 2013, 132, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Ferstl, R.; Frei, R.; Schiavi, E.; Konieczna, P.; Barcik, W.; Ziegler, M.; Lauener, R.P.; Chassard, C.; Lacroix, C.; Akdis, C.A.; et al. Histamine receptor 2 is a key influence in immune responses to intestinal histamine-secreting microbes. J. Allergy Clin. Immunol. 2014, 134, 744–746. [Google Scholar] [CrossRef]
- Barcik, W.; Pugin, B.; Westermann, P.; Perez, N.R.; Ferstl, R.; Wawrzyniak, M.; Smolinska, S.; Jutel, M.; Hessel, E.M.; Michalovich, D.; et al. Histamine-secreting microbes are increased in the gut of adult asthma patients. J. Allergy Clin. Immunol. 2016, 138, 1491–1494 e1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallardo, P.; Izquierdo, M.; Vidal, R.M.; Soto, F.; Ossa, J.C.; Farfan, M.J. Gut Microbiota-Metabolome Changes in Children With Diarrhea by Diarrheagenic E. coli. Front. Cell Infect. Microbiol. 2020, 10, 485. [Google Scholar] [CrossRef]
- Barcik, W.; Pugin, B.; Bresco, M.S.; Westermann, P.; Rinaldi, A.; Groeger, D.; Van Elst, D.; Sokolowska, M.; Krawczyk, K.; Frei, R.; et al. Bacterial secretion of histamine within the gut influences immune responses within the lung. Allergy 2019, 74, 899–909. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Nwe, P.K.; Yang, Y.; Rosen, C.E.; Bielecka, A.A.; Kuchroo, M.; Cline, G.W.; Kruse, A.C.; Ring, A.M.; Crawford, J.M.; et al. A Forward Chemical Genetic Screen Reveals Gut Microbiota Metabolites That Modulate Host Physiology. Cell 2019, 177, 1217–1231 e1218. [Google Scholar] [CrossRef] [PubMed]
- Mishima, Y.; Ishihara, S. Molecular Mechanisms of Microbiota-Mediated Pathology in Irritable Bowel Syndrome. Int. J. Mol. Sci. 2020, 21, 8664. [Google Scholar] [CrossRef] [PubMed]
- Barbara, G.; Wang, B.; Stanghellini, V.; de Giorgio, R.; Cremon, C.; Di Nardo, G.; Trevisani, M.; Campi, B.; Geppetti, P.; Tonini, M.; et al. Mast cell-dependent excitation of visceral-nociceptive sensory neurons in irritable bowel syndrome. Gastroenterology 2007, 132, 26–37. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Name | Log2 Fold Change | Description | Function/ Comment | Ref. Function |
|---|---|---|---|---|---|
| Histamine-mediated upregulation | |||||
| Histamine metabolism, transport and regulation | |||||
| PA5390 | hinG | 7.9 | Probable peptidic bond hydrolase | Histamine utilization | [24] |
| PA5391 | hinH | 10.9 | Hypothetical protein | ||
| PA5392 | hinL | 10.7 | Conserved hypothetical protein | ||
| PA5393 | hinF | 11.1 | Conserved hypothetical protein | ||
| PA0219 | hinD | 10.0 | Probable aldehyde dehydrogenase | ||
| PA0221 | hinC | 10.0 | Probable aminotransferase | ||
| PA0220 | hinA | 9.5 | Histamine transporter | Histamine transport | |
| PA0218 | hinK | 4.8 | Transcriptional regulator | Histamine-mediated regulation | |
| PA0222 | 8.7 | Solute-binding protein | Possibly transport | ||
| Iron acquisition | |||||
| PA0931 | pirA | 3.2 | Ferric enterobactin receptor PirA | [25] | |
| PA2385 | pvdQ | 6.3 | 3-oxo-C12-homoserine lactone acylase PvdQ | Siderophore pyoverdin synthesis, secretion, regulation and pyoverdin-Fe uptake | [26] |
| PA2386 | pvdA | 7.5 | L-ornithine N5-oxygenase | ||
| PA2389 | pvdR | 2.6 | PvdR | ||
| PA2390 | pvdT | 2.4 | PvdT | ||
| PA2392 | pvdP | 4.1 | PvdP | ||
| PA2394 | pvdN | 5.9 | PvdN | ||
| PA2395 | pvdO | 6.3 | PvdO | ||
| PA2396 | pvdF | 3.4 | Pyoverdine synthetase F | ||
| PA2397 | pvdE | 6.3 | Pyoverdine biosynthesis protein PvdE | ||
| PA2398 | fpvA | 6.0 | Ferripyoverdine receptor | ||
| PA2399 | pvdD | 2.9 | Pyoverdine synthetase D | ||
| PA2400 | pvdJ | 3.0 | PvdJ | ||
| PA2413 | pvdH | 5.6 | L-2,4-diaminobutyrate:2-ketoglutarate 4-aminotransferase | ||
| PA2424 | pvdL | 5.8 | PvdL | ||
| PA2425 | pvdG | 6.2 | PvdG | ||
| PA2426 | pvdS | 5.7 | Sigma factor PvdS | ||
| PA0472 | fiuI | 3.1 | ECF sigma factor FiuI | Ferrichrome activated | [27] |
| PA2468 | foxI | 2.5 | ECF sigma factor FoxI | Ferrioxamine activated | [28] |
| PA3410 | hasI | 2.9 | ECF sigma factor HasI | Heme activated | [29] |
| PA4168 | fpvB | 3.3 | Second ferric pyoverdine receptor FpvB | Pyoverdine transport | [30] |
| PA4221 | fptA | 1.7 | Fe(III)-pyochelin outer membrane receptor precursor | Siderophore pyochelin synthesis and transport | [31] |
| PA4226 | pchE | 3.1 | Dihydroaeruginoic acid synthetase | ||
| PA4228 | pchD | 4.1 | Pyochelin biosynthesis protein PchD | ||
| PA4229 | pchC | 3.6 | Pyochelin biosynthetic protein PchC | ||
| PA4230 | pchB | 2.7 | Salicylate biosynthesis protein PchB | ||
| PA4231 | pchA | 2.3 | Salicylate biosynthesis isochorismate synthase | ||
| PA4687 | hitA | 3.3 | Ferric iron-binding periplasmic protein HitA | Iron transport | [32] |
| PA4688 | hitB | 3.2 | Iron (III)-transport system permease HitB | ||
| Quorum sensing | |||||
| PA0996 | pqsA | 3.4 | Probable coenzyme A ligase | Pseudomonas quinolone signal (PQS) quorum sensing system | [33] |
| PA0997 | pqsB | 3.8 | PqsB | ||
| PA0998 | pqsC | 3.8 | PqsC | ||
| PA0999 | pqsD | 3.8 | 3-oxoacyl-[acyl-carrier-protein] synthase III | ||
| PA1000 | pqsE | 3.6 | Quinolone signal response protein | ||
| PA1001 | phnA | 3.5 | Anthranilate synthase components I and II (important for PQS synthesis) | PQS synthesis | [34] |
| PA1002 | phnB | 3.0 | |||
| Secretion system | |||||
| PA1718 | pscE | 2.3 | Type III export protein PscE | Type III secretion apparatus | [35] |
| PA1721 | pscH | 1.9 | Type III export protein PscH | ||
| PA1715 | pscB | 1.8 | Type III export apparatus protein | ||
| Regulation | |||||
| PA0707 | toxR | 1.9 | Transcriptional regulator ToxR | Exotoxin A expression | [36] |
| PA0612 | ptrB | 2.0 | Repressor PtrB | Type III secretion system expression | [37] |
| PA1431 | rsaL | 2.0 | Regulatory protein RsaL | Virulence and biofilm formation | [38] |
| PA2227 | vqsM | 2.4 | Transcriptional regulator VqsM | Quorum sensing and virulence | [39] |
| PA2686 | pfeR | 3.1 | PfeR response regulator | Enterobactin receptor | [40] |
| PA2687 | pfeS | 2.6 | PfeS sensor kinase | ||
| PA3006 | psrA | 1.8 | Transcriptional regulator PsrA | Type III secretion system | [41] |
| PA4315 | mvaT | 2.3 | Transcriptional regulator MvaT | Type III secretion system | [42] |
| PA5124 | ntrB | 4.0 | NtrB kinase | Invasiveness and Virulence | [43] |
| PA5125 | ntrC | 3.7 | NtrC response regulator | ||
| Others | |||||
| PA4760 | dnaJ | 3.2 | Heat shock protein | Pyocyanin production | [44] |
| PA4761 | dnaK | 3.7 | Chaperone DnaK | Translocation across the intestinal epithelia cells | [45] |
| Histamine-mediated downregulation | |||||
| Regulation | |||||
| PA0173 | cheB2 | −2.1 | CheB2 methylesterase | Che2 pathway, unknown function, involved in virulence | [46,47] |
| PA0174 | cheD | −2.2 | CheD deamidase | ||
| PA0175 | cheR2 | −2.4 | CheR2 methyltransferase | ||
| PA0176 | mcpB/aer2 | −2.3 | Aer2/McpB chemoreceptor | ||
| PA0177 | cheW | −1.9 | CheW coupling protein | ||
| PA4293 | pprA | −2.5 | Sensor kinase PprA | Quorum sensingregulation | [48] |
| PA4296 | pprB | −1.6 | Response regulator PprB | ||
| Motility | |||||
| PA1930 | mcpS | −2.1 | Chemoreceptor McpS | Chemotaxis | [49] |
| PA2561 | ctpH | −2.4 | Inorganic phosphate (Pi) specific chemoreceptor CtpH | Pi is a major virulence signal | [50,51] |
| Others | |||||
| PA4299-4306 | Flp-tad-rcp locus | −2.3 to −4.8 | Formation of type IVb pili | Aggregation and biofilm formation | [52] |
| PA4236 | katA | −2.1 | Major catalase KatA | Osmoprotection and virulence | [53] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krell, T.; Gavira, J.A.; Velando, F.; Fernández, M.; Roca, A.; Monteagudo-Cascales, E.; Matilla, M.A. Histamine: A Bacterial Signal Molecule. Int. J. Mol. Sci. 2021, 22, 6312. https://doi.org/10.3390/ijms22126312
Krell T, Gavira JA, Velando F, Fernández M, Roca A, Monteagudo-Cascales E, Matilla MA. Histamine: A Bacterial Signal Molecule. International Journal of Molecular Sciences. 2021; 22(12):6312. https://doi.org/10.3390/ijms22126312
Chicago/Turabian StyleKrell, Tino, José A. Gavira, Félix Velando, Matilde Fernández, Amalia Roca, Elizabet Monteagudo-Cascales, and Miguel A. Matilla. 2021. "Histamine: A Bacterial Signal Molecule" International Journal of Molecular Sciences 22, no. 12: 6312. https://doi.org/10.3390/ijms22126312
APA StyleKrell, T., Gavira, J. A., Velando, F., Fernández, M., Roca, A., Monteagudo-Cascales, E., & Matilla, M. A. (2021). Histamine: A Bacterial Signal Molecule. International Journal of Molecular Sciences, 22(12), 6312. https://doi.org/10.3390/ijms22126312