
Evaluation of Cytotoxic and Mutagenic Effects of the Synthetic Cathinones Mexedrone, α-PVP and α-PHP

 , , ,
, , , 

Abstract
:1. Introduction
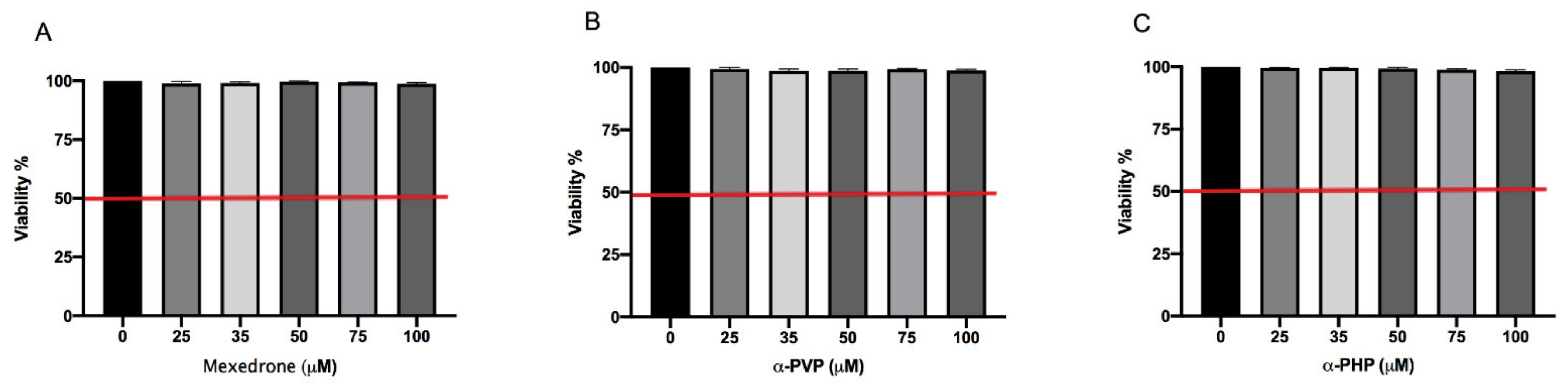
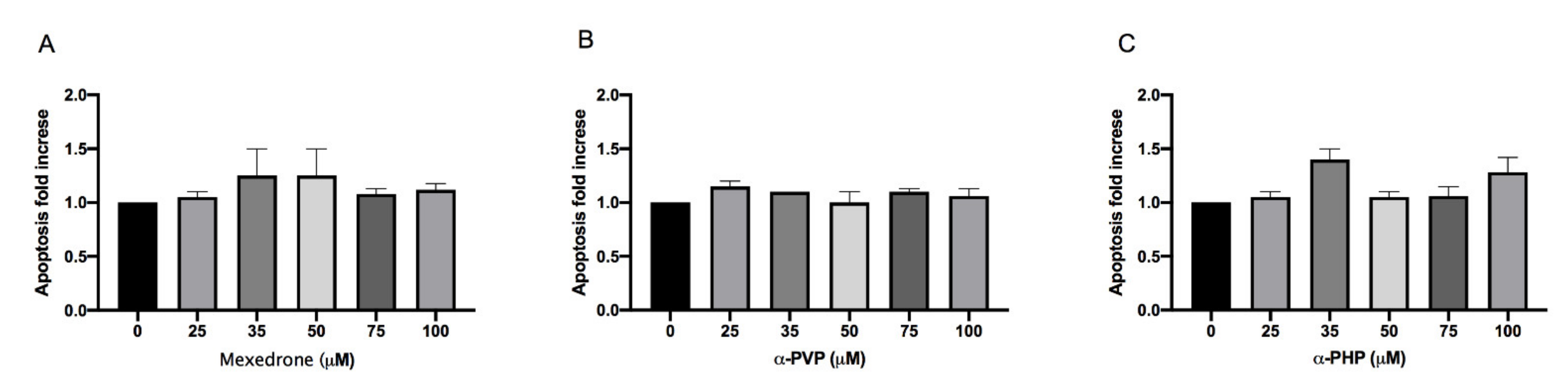
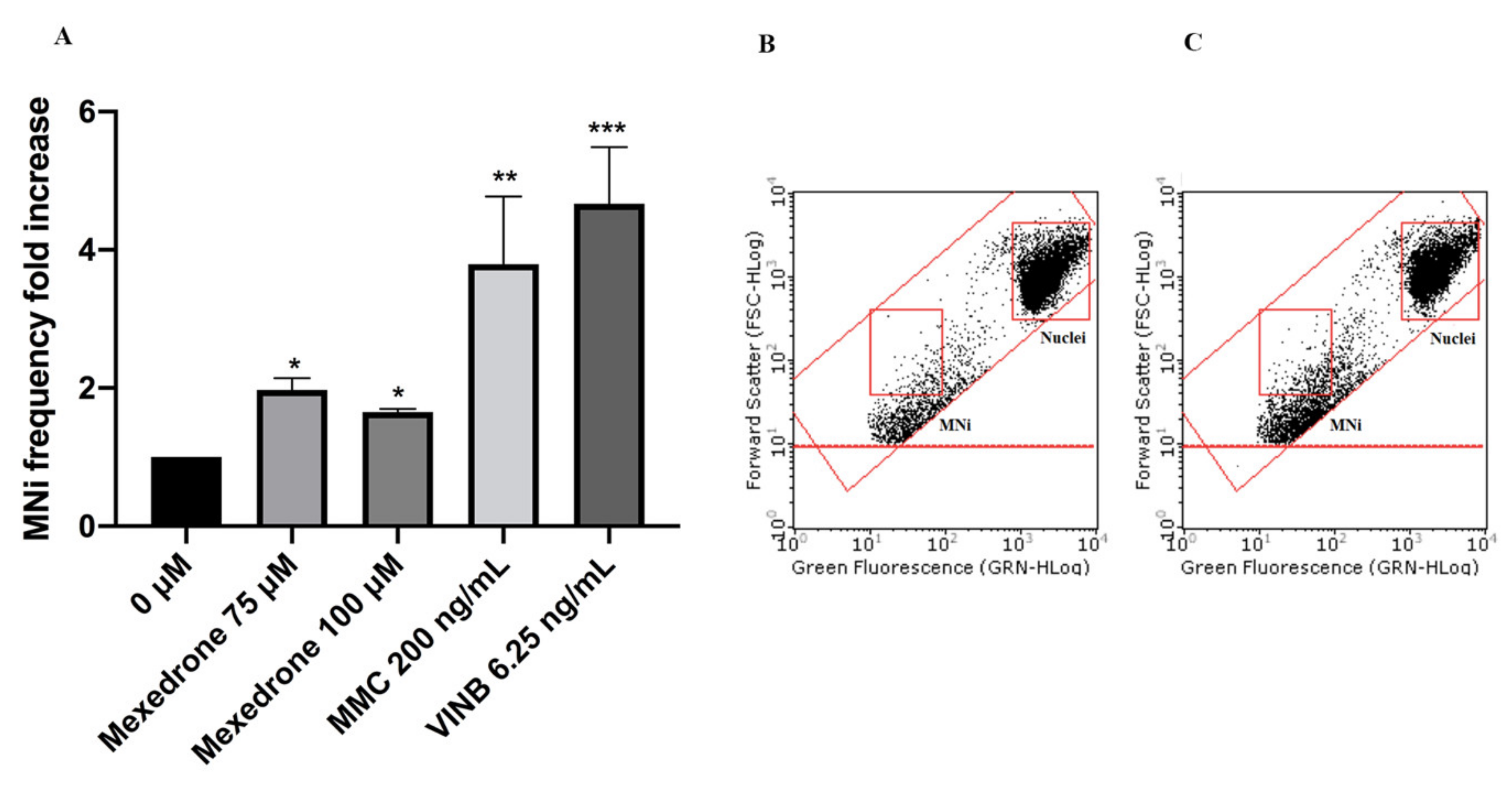
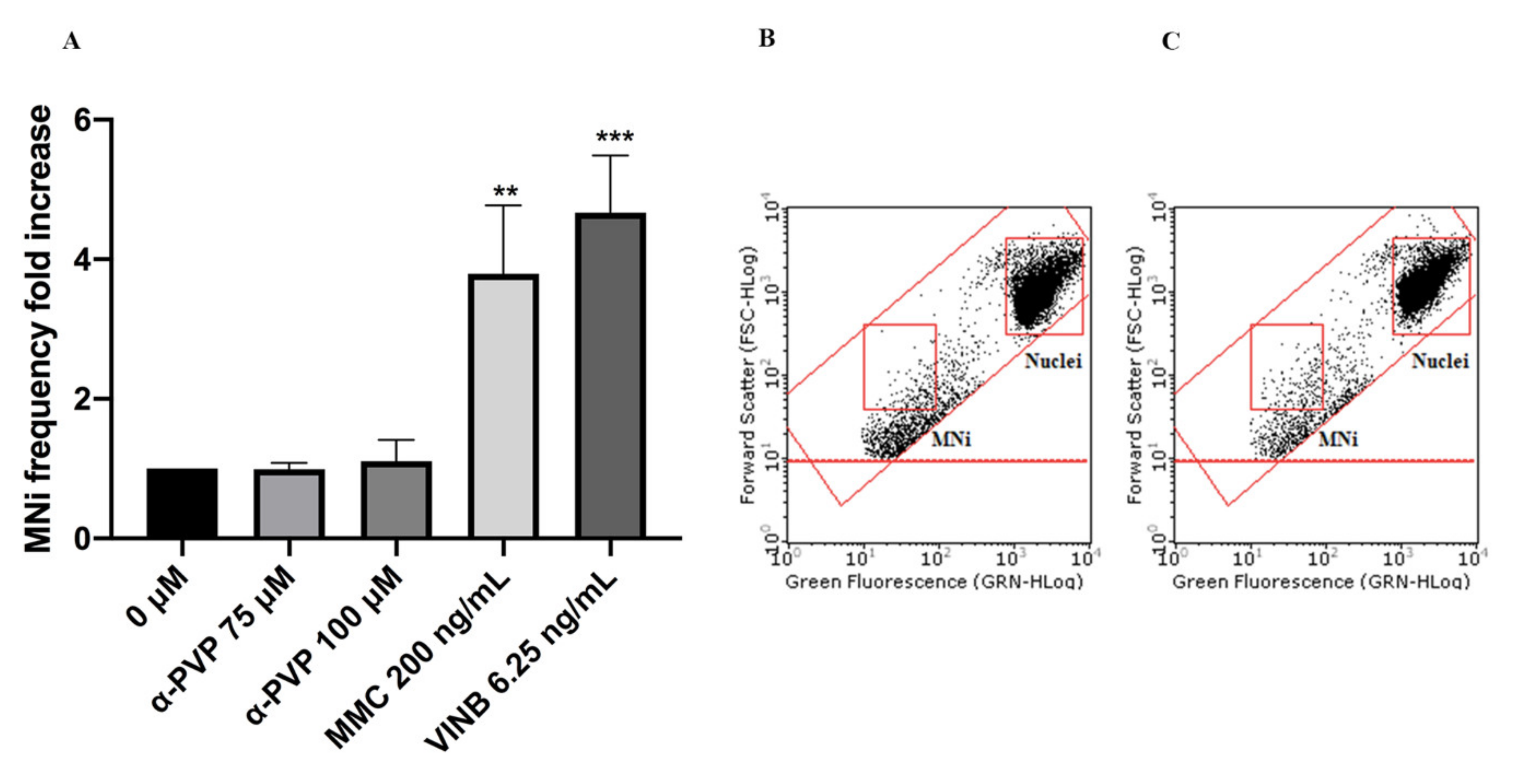
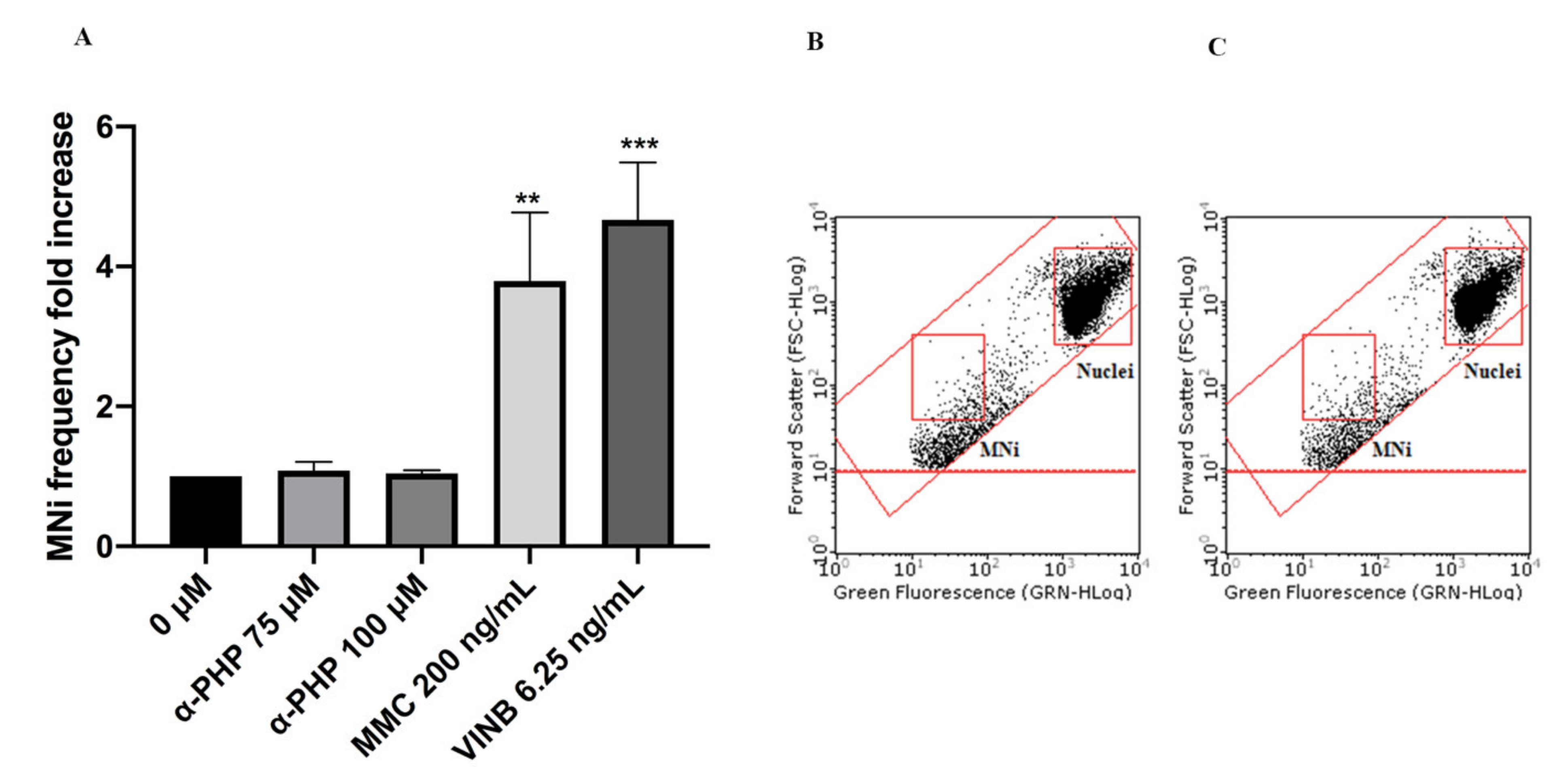
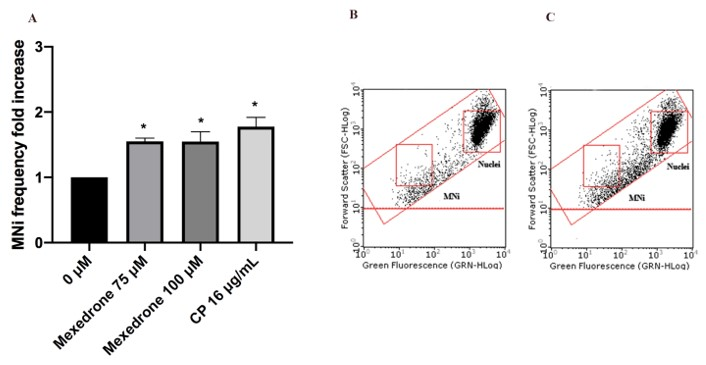
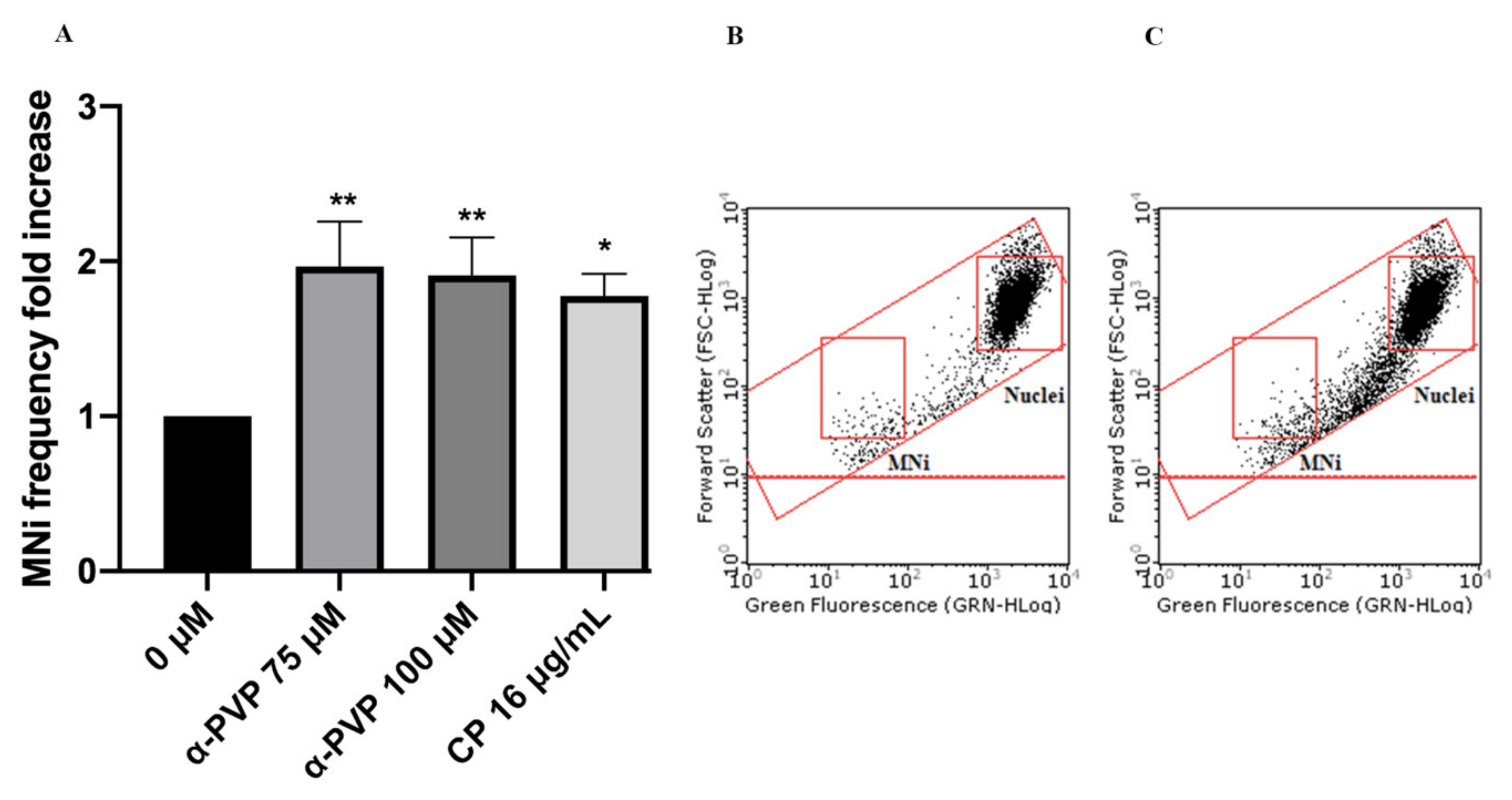
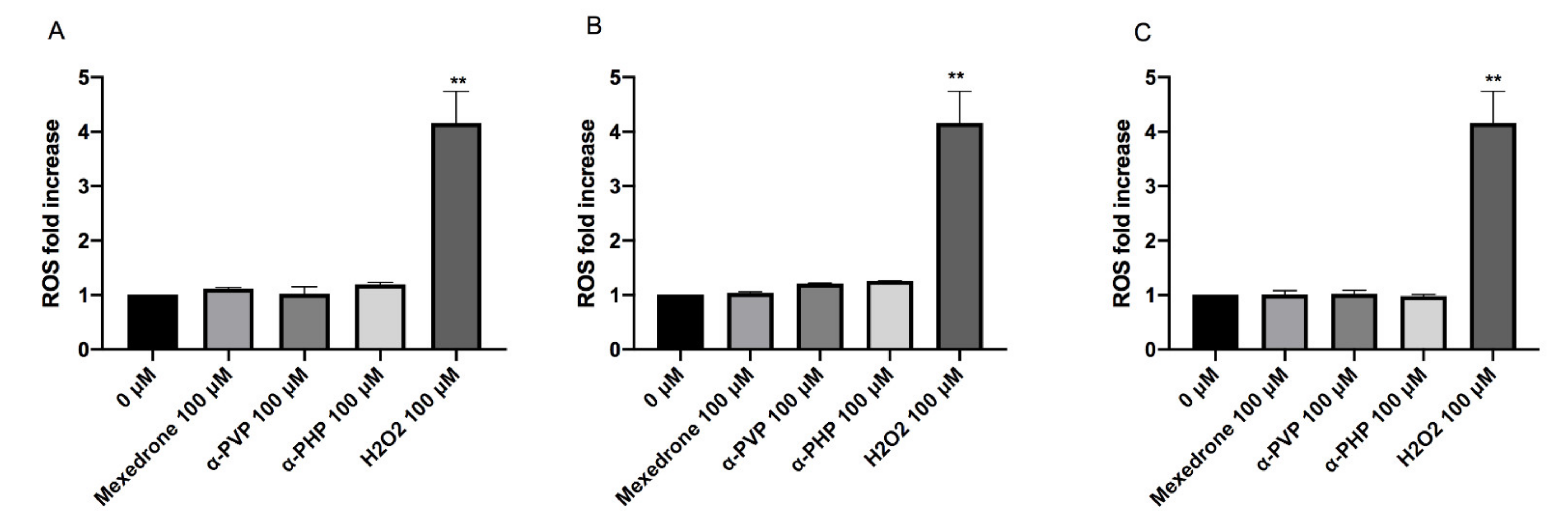
2. Results
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Synthetic Cathinones
4.3. Cell Culture and Treatments
4.4. Flow Cytometry
4.5. Cytotoxicity Analysis
4.6. Cytostasis Analysis
4.7. Apoptosis Analysis
4.8. Genotoxicity Analysis
4.9. ROS Analysis
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availabity Statement
Conflicts of Interest
References
- Lenzi, M.; Cocchi, V.; Cavazza, L.; Bilel, S.; Hrelia, P.; Marti, M. Genotoxic Properties of Synthetic Cannabinoids on TK6 Human Cells by Flow Cytometry. Int. J. Mol. Sci. 2020, 21, 1150. [Google Scholar] [CrossRef] [Green Version]
- Koller, V.J.; Ferk, F.; Al-Serori, H.; Mišík, M.; Nersesyan, A.; Auwärter, V.; Grummt, T.; Knasmüller, S. Genotoxic properties of representatives of alkylindazoles and aminoalkyl-indoles which are consumed as synthetic cannabinoids. Food Chem. Toxicol. 2015, 80, 130–136. [Google Scholar] [CrossRef]
- Koller, V.J.; Zlabinger, G.J.; Auwärter, V.; Fuchs, S.; Knasmueller, S. Toxicological profiles of selected synthetic cannabinoids showing high binding affinities to the cannabinoid receptor subtype CB₁. Arch. Toxicol. 2013, 87, 1287–1297. [Google Scholar] [CrossRef]
- Cocchi, V.; Gasperini, S.; Hrelia, P.; Tirri, M.; Marti, M.; Lenzi, M. Novel Psychoactive Phenethylamines: Impact on Genetic Material. Int. J. Mol. Sci. 2020, 21, 9616. [Google Scholar] [CrossRef] [PubMed]
- European Monitoring Centre for Drugs and Drug Addiction (EMCDDA). European Drug Report 2020: Trends and Developments; Publications Office of the European Union: Lisbon, Portugal, 2020. [Google Scholar]
- European Monitoring Centre for Drugs and Drug Addiction (EMCDDA). European Drug Report 2018: Trends and Developments; Publications Office of the European Union: Luxembourg, 2018. [Google Scholar]
- European Monitoring Centre for Drugs and Drug Addiction (EMCDDA). EMCDDA-Europol Joint Report on a New Psychoactive Substance: 1-phenyl-2-(1-pyrrolidynyl)-1-pentanone (α-PVP); EMCDDA-Europol Joint Publications: Lisbon, Portugal, 2015. [Google Scholar]
- Klavž, J.; Gorenjak, M.; Marinšek, M. Suicide attempt with a mix of synthetic cannabinoids and synthetic cathinones: Case report of non-fatal intoxication with AB-CHMINACA, AB-FUBINACA, alpha-PHP, alpha-PVP and 4-CMC. Forensic Sci. Int. 2016, 265, 121–124. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). 1-Phenyl-2-(pyrrolidine-1-yl)pentan-1-one (α-PVP). Critical Review Report; Expert Committee on Drug Dependence: Geneva, Switzerland, 2015. [Google Scholar]
- Zawilska, B.J.; Wojcieszak, J. α-Pyrrolidinophenones: A new wave of designer cathinones. Forensic Toxicol. 2017, 35, 201–216. [Google Scholar] [CrossRef]
- Roberts, L.; Ford, L.; Patel, N.; Vale, J.A.; Bradberry, S.M. 11 analytically confirmed cases of mexedrone use among polydrug users. Clin. Toxicol. 2017, 55, 181–186. [Google Scholar] [CrossRef]
- Adamowicz, P.; Hydzic, P. Fetal death associated with the use of 3,4-MDPHP and α-PHP. Clin. Toxicol. 2019, 57, 112–116. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Alpha-PHP (α-Pyrrolidinohexanophenone) or PV-7. Critical Review Report; Expert Committee on Drug Dependence: Geneva, Switzerland, 2019. [Google Scholar]
- Zaami, S.; Giorgetti, R.; Pichini, S.; Pantano, F.; Marinelli, E.; Busardò, F.P. Synthetic cathinones related fatalities: An update. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 268–274A. [Google Scholar] [PubMed]
- La Maida, N.; di Trana, A.; Giorgetti, R.; Tagliabracci, A.; Busardò, F.P.; Huestis, M.A. Review of Synthetic Cathinone-Related Fatalities From 2017 to 2020. Ther. Drug Monit. 2021, 43, 52–68. [Google Scholar] [CrossRef]
- Uchiyama, N.; Shimokawa, Y.; Kawamura, M.; Kikura-Hanajiri, R.; Hakamatsuka, T. Chemical analysis of a benzofuran derivative, 2-(2-ethylaminopropyl)benzofuran(2-EAPB), eight synthetic cannabi-noids, five cathinone derivatives, and five other designer drugs newly detected in illegal products. Forensic Toxicol. 2014, 32, 266–281. [Google Scholar] [CrossRef]
- Qian, Z.; Jia, W.; Li, T.; Liu, C.; Hua, Z. Identification and analytical characterization of four synthetic cathinone derivatives iso-4-BMC, β-TH-naphyrone, mexedrone, and 4-MDMC. Drug Test. Anal. 2017, 9, 274–281. [Google Scholar] [CrossRef]
- Pantano, F.; Tittarelli, R.; Mannocchi, G.; Pacifici, R.; di Luca, A.; Busardò, F.P.; Marinelli, E. Neurotoxicity Induced by Mephedrone: An up-to-date Review. Curr. Neuropharmacol. 2017, 15, 738–749. [Google Scholar] [CrossRef] [Green Version]
- Busardò, F.P.; Kyriakou, C.; Napoletano, S.; Marinelli, E.; Zaami, S. Mephedrone related fatalities: A review. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 3777–3790. [Google Scholar] [PubMed]
- Dragogna, F.; Oldani, L.; Buoli, M.; Altamura, A.C. A case of severe psychosis induced by novel recreational drugs. F1000Research 2016, 3, 21. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Mita, T.; Usui, K.; Kamijo, Y.; Kikuchi, S.; Onodera, M.; Fujino, Y.; Inoue, Y. Toxicokinetics of the Synthetic Cathinone α-Pyrrolidinohexa-nophenone. J. Anal. Toxicol. 2018, 42, e1–e5. [Google Scholar] [CrossRef]
- Quesada, L.; Gomila, I.; Yates, C.; Barcelo, C.; Puiguriguer, J.; Barcelo, B. Elimination half-life of alpha-pyrrolidinovalerophenone in an acute non-fatal intoxication. Clin. Toxicol. 2016, 54, 531–532. [Google Scholar] [CrossRef] [PubMed]
- Umebachi, R.; Aoki, H.; Sugita, M.; Taira, T.; Wakai, S.; Saito, T.; Inokuchi, S. Clinical characteristics of α-pyrrolidinovalerophenone (α-PVP) poisoning. Clin. Toxicol. 2016, 54, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Rickli, A.; Hoener, M.C.; Liechti, M.E. Monoamine transporter and receptor interaction profiles of novel psychoactive substances: Para-halogenated amphetamines and pyrovalerone cathinones. Eur. Neuropsychopharmacol. 2015, 25, 365–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eshleman, A.J.; Wolfrum, K.M.; Reed, J.F.; Kim, S.K.; Swanson, T.; Johnson, R.A.; Janowsky, A. Structure-activity relationship of substituted cathinones, with trasporter binding, uptake, and release. J. Pharmacol. Exp. Ther. 2017, 360, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Marusich, J.A.; Antonazzo, K.R.; Wiley, J.L.; Blough, B.E.; Partilla, J.S.; Baumann, M.H. Pharmacology of novel synthetic stimulants structurally related to the “bath salts” constituent 3,4-methylenedioxypyrovalerone (MDPV). Neuropharmacology 2014, 87, 206–213. [Google Scholar] [CrossRef] [Green Version]
- Senior, T.; Botha, M.J.; Kennedy, A.R.; Calvo-Castro, J. Understanding the Contribution of Individual Amino Acid Residues in the Binding of Psychoactive Substances to Monoamine Transporters. ACS Omega 2020, 5, 17223–17231. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, G.; Morris, N.; Kavanagh, P.V.; Power, J.D.; Dowling, G.; Twamley, B.; O’Brien, J.; Talbot, B.; Walther, D.; Partilla, J.S.; et al. Synthesis, characterization and monoamine transporter activity of the new psychoactive substance mexedrone and its N-methoxy positional isomer, N-methoxymephedrone. Drug Test. Anal. 2017, 9, 358–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Serori, H.; Ferk, F.; Angerer, V.; Mišík, M.; Nersesyan, A.; Setayesh, T.; Auwärter, V.; Haslinger, E.; Huber, W.; Knasmüller, S. Investigations of the genotoxic properties of two synthetic cathinones (3-MMC, 4-MEC) which are used as psychoactive drugs. Toxicol. Res. 2016, 5, 1410–1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Test, No. 487: In Vitro Mammalian Cell Micronucleus Test. In OECD Guideline for the Testing of Chemicals; OECD: Paris, France, 2016; Available online: https://www.oecd-ilibrary.org/environment/test-no-487-in-vitro-mammalian-cell-micronucleus-test_9789264264861-en (accessed on 15 January 2020).
- Lenzi, M.; Cocchi, V.; Hrelia, P. Flow cytometry vs. optical microscopy in the evaluation of the genotoxic potential of xenobiotic compounds. Cytom. B Clin. Cytom. 2018, 94, 696–706. [Google Scholar] [CrossRef] [Green Version]
- Fowler, P.; Smith, R.; Smith, K.; Young, J.; Jeffrey, L.; Carmichael, P.; Kirkland, D.; Pfuhler, S. Reduction of misleading (“false”) positive results in mammalian cell genotoxicity assays. III: Sensitivity of human cell types to known genotoxic agents. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2014, 767, 28–36. [Google Scholar] [CrossRef]
- Tyrkkö, E.; Pelander, A.; Ketola, R.A.; Ojanperä, I. In silico and in vitro metabolism studies support identification of designer drugs in human urine by liquid chromatography/quadrupole-time-of-flight mass spectrometry. Anal. Bioanal. Chem. 2013, 405, 6697–6709. [Google Scholar] [CrossRef]
- Negreira, N.; Erratico, C.; Kosjek, T.; van Nuijs, A.L.; Heath, E.; Neels, H.; Covaci, A. In vitro Phase I and Phase II metabolism of α-pyrrolidinovalerophenone (α-PVP), methylenedioxypyrovalerone (MDPV) and methedrone by human liver microsomes and human liver cytosol. Anal. Bioanal. Chem. 2015, 407, 5803–5816. [Google Scholar] [CrossRef]
- Glennon, R.A.; Young, R. Neurobiology of 3,4-methylenedioxypreovalerone (MDPV) and alpha-pyrrolidinovalerophenone (α-PVP). Brain Res. Bull. 2016, 126 Pt 1, 111–126. [Google Scholar] [CrossRef] [Green Version]
- Paul, M.; Bleicher, S.; Guber, S.; Ippisch, J.; Polettini, A.; Schultis, W. Identification of phase I and II metabolites of the new designer drug α-pyrrolidinohexiophenone (α-PHP) in human urine by liquid chromatography quadrupole time-of-flight mass spectrometry (LC-QTOF-MS). J. Mass Spectrom. 2015, 50, 1305–1317. [Google Scholar] [CrossRef]
- Matsuta, S.; Shima, N.; Kakehashi, H.; Kamata, H.; Nakano, S.; Sasaki, K.; Kamata, T.; Nishioka, H.; Miki, A.; Zaitsu, K.; et al. Metabolism of α-PHP and α-PHPP in humans and the effects of alkyl chain lengths on the metabolism of α-pyrrolidinophenone-type designer drugs. Forensic Toxicol. 2018, 36, 486–497. [Google Scholar] [CrossRef] [Green Version]
- Carlier, J.; Diao, X.; Giorgetti, R.; Busardò, F.P.; Huestis, M.A. Pyrrolidinyl Synthetic Cathinones α-PHP and 4F-α-PVP Metabolite Profiling Using Human Hepatocyte Incubations. Int. J. Mol. Sci. 2020, 22, 230. [Google Scholar] [CrossRef]
- Zhou, X.; Luethi, D.; Sanvee, G.M.; Bouitbir, J.; Liechti, M.E.; Krähenbühl, S. Molecular Toxicological Mechanisms of Synthetic Cathinones on C2C12 Myoblasts. Int. J. Mol. Sci. 2019, 20, 1561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valente, M.J.; Amaral, C.; Correia-da-Silva, G.; Duarte, J.A.; Bastos, M.L.; Carvalho, F.; Guedes de Pinho, P.; Carvalho, M. Methylone and MDPV activate autophagy in human dopaminergic SH-SY5Y cells: A new insight into the context of β-keto amphetamines-related neurotoxicity. Arch. Toxicol. 2017, 91, 3663–3676. [Google Scholar] [CrossRef] [PubMed]
- Valente, M.J.; Bastos, M.L.; Fernandes, E.; Carvalho, F.; Guedes de Pinho, P.; Carvalho, M. Neurotoxicity of β-Keto Amphetamines: Deathly Mechanisms Elicited by Methylone and MDPV in Human Dopaminergic SH-SY5Y Cells. ACS Chem. Neurosci. 2017, 8, 850–859. [Google Scholar] [CrossRef]
- Kamińska, K.; Noworyta-Sokołowska, K.; Górska, A.; Rzemieniec, J.; Wnuk, A.; Wojtas, A.; Kreiner, G.; Kajta, M.; Gołembiowska, K. The Effects of Exposure to Mephedrone During Adolescence on Brain Neurotransmission and Neurotoxicity in Adult Rats. Neurotox. Res. 2018, 34, 525–537. [Google Scholar] [CrossRef]
- De Hondt, H.A.; Fahmy, A.M.; Abdelbaset, S.A. Chromosomal and biochemical studies on the effect of kat extract on laboratory rats. Environ. Mutagen. 1984, 6, 851–860. [Google Scholar] [CrossRef]
- Tariq, M.; Parmar, N.S.; Qureshi, S.; El-Feraly, F.S.; Al-Meshal, I.A. Clastogenic evaluation of cathinone and amphetamine in somatic cells of mice. Mutat. Res. 1987, 190, 153–157. [Google Scholar] [CrossRef]
- Qureshi, S.; Tariq, M.; El-Feraly, F.S.; Al-Meshal, I.A. Genetic effects of chronic treatment with cathinone in mice. Mutagenesis 1988, 3, 481–483. [Google Scholar] [CrossRef]
- Chatterjee, N.; Walker, G.C. Mechanisms of DNA damage, repair, and mutagenesis. Environ. Mol. Mutagen. 2017, 58, 235–263. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Relative Population Doubling (RPD) | |||
|---|---|---|---|
| Mexedrone | α-PVP | α-PHP | |
| 0 µM | 100.00% | 100.00% | 100.00% |
| 25 µM | 97.0% ± 3.2 | 91.5% ± 5.9 | 88.3% ± 1.3 |
| 35 µM | 92.0% ± 4.2 | 88.4% ± 6.8 | 94.4% ± 1.7 |
| 50 µM | 88.4% ± 7.9 | 88.7% ± 7.1 | 96.8% ± 3.2 |
| 75 µM | 95.2% ± 3.5 | 89.9% ± 4.6 | 85.2% ± 3.5 |
| 100 µM | 90.1% ± 3.6 | 87.6% ± 6.3 | 85.9% ± 5.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lenzi, M.; Cocchi, V.; Gasperini, S.; Arfè, R.; Marti, M.; Hrelia, P. Evaluation of Cytotoxic and Mutagenic Effects of the Synthetic Cathinones Mexedrone, α-PVP and α-PHP. Int. J. Mol. Sci. 2021, 22, 6320. https://doi.org/10.3390/ijms22126320
Lenzi M, Cocchi V, Gasperini S, Arfè R, Marti M, Hrelia P. Evaluation of Cytotoxic and Mutagenic Effects of the Synthetic Cathinones Mexedrone, α-PVP and α-PHP. International Journal of Molecular Sciences. 2021; 22(12):6320. https://doi.org/10.3390/ijms22126320
Chicago/Turabian StyleLenzi, Monia, Veronica Cocchi, Sofia Gasperini, Raffaella Arfè, Matteo Marti, and Patrizia Hrelia. 2021. "Evaluation of Cytotoxic and Mutagenic Effects of the Synthetic Cathinones Mexedrone, α-PVP and α-PHP" International Journal of Molecular Sciences 22, no. 12: 6320. https://doi.org/10.3390/ijms22126320
APA StyleLenzi, M., Cocchi, V., Gasperini, S., Arfè, R., Marti, M., & Hrelia, P. (2021). Evaluation of Cytotoxic and Mutagenic Effects of the Synthetic Cathinones Mexedrone, α-PVP and α-PHP. International Journal of Molecular Sciences, 22(12), 6320. https://doi.org/10.3390/ijms22126320