
Toxicity of Jegosaponins A and B from Styrax japonica Siebold et al. Zuccarini in Prostate Cancer Cells and Zebrafish Embryos Resulting from Increased Membrane Permeability

 , , and
, , and
Abstract
:1. Introduction
2. Results
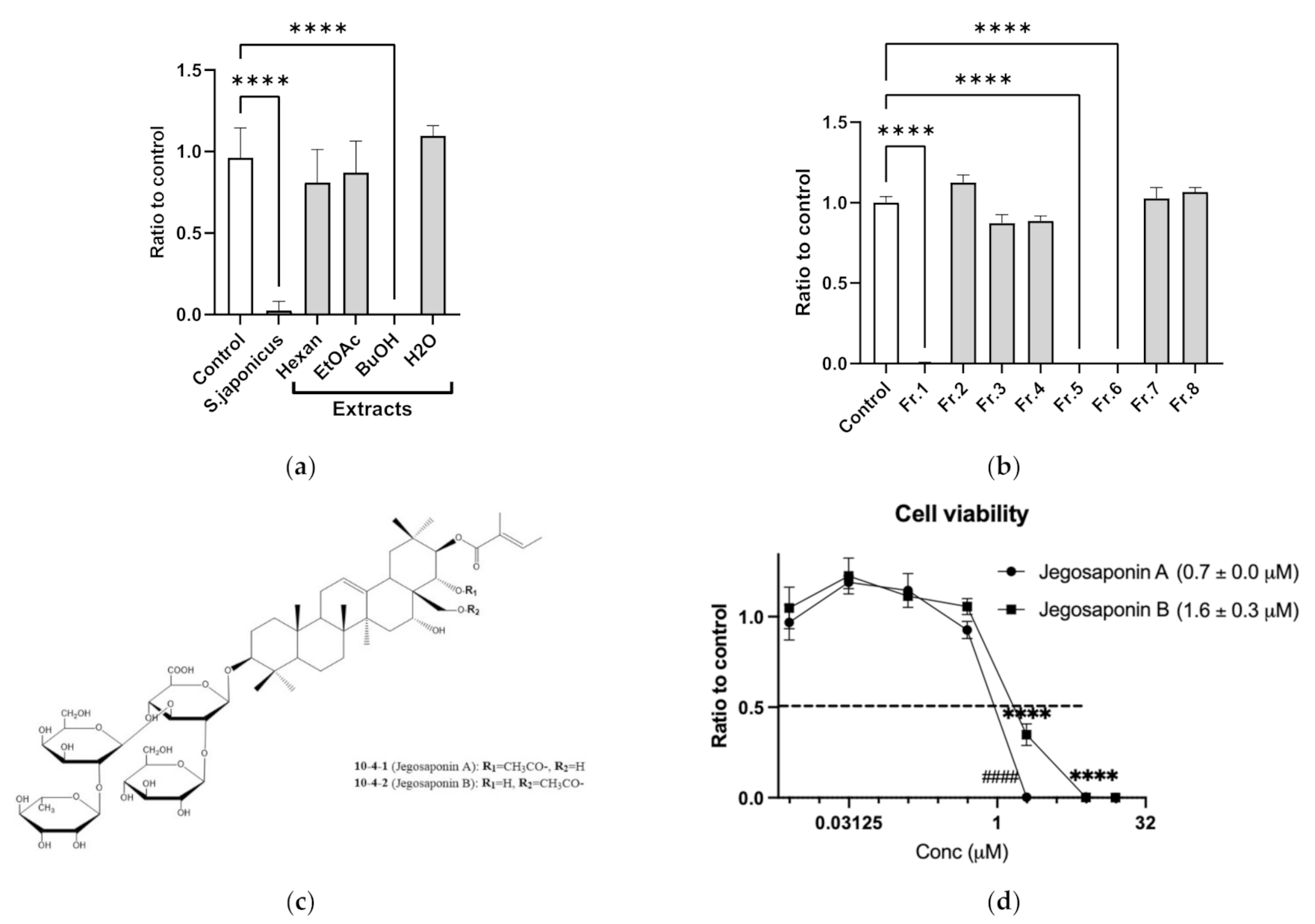
2.1. Jegosaponins from Styrax japonica Siebold et al. Zuccarini (SJSZ) Suppresses Cell Viability in PC-3 Prostate Cancer Cells
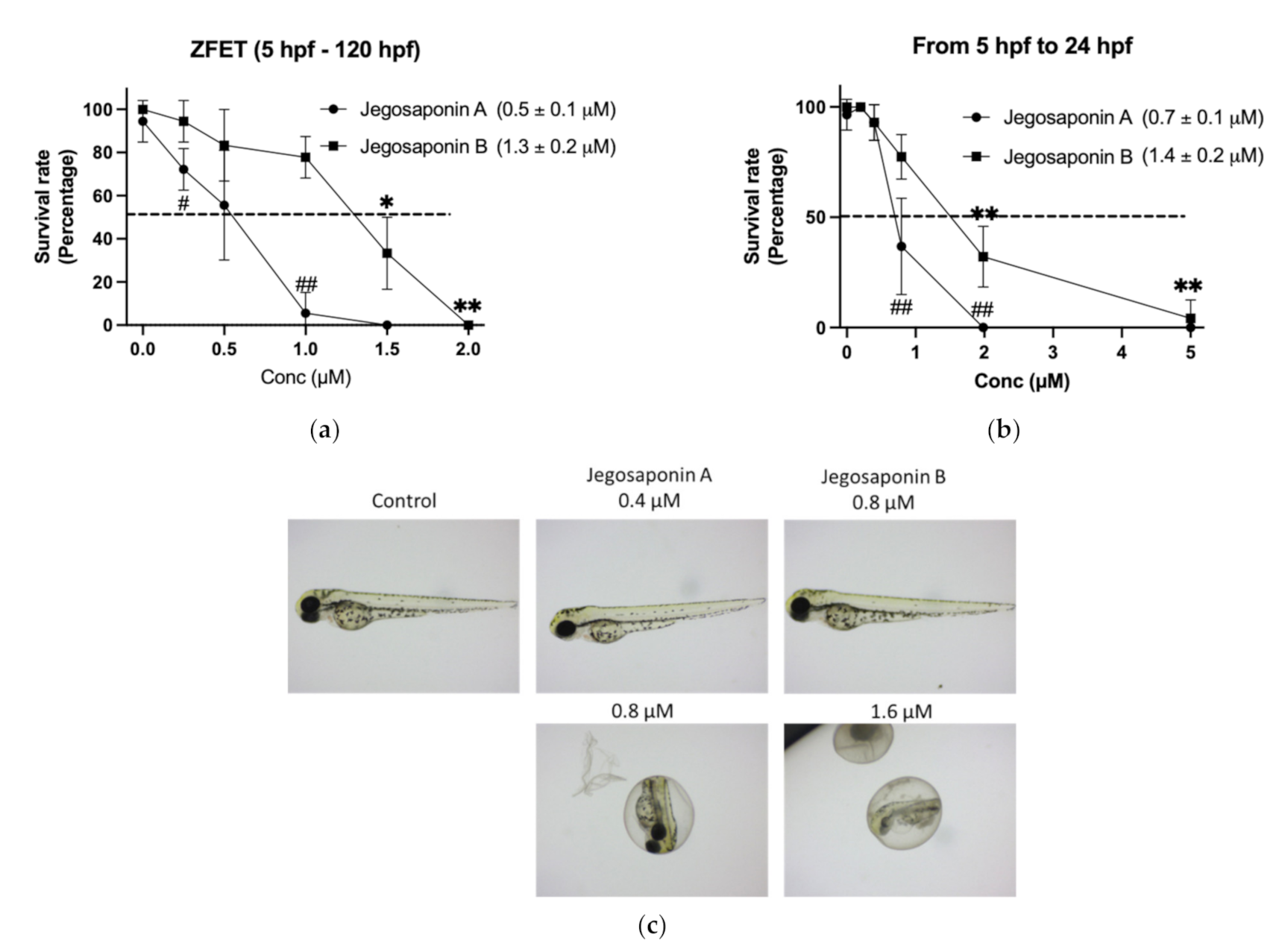
2.2. Acute Toxicity of Jegosaponins in Zebrafish Embryos
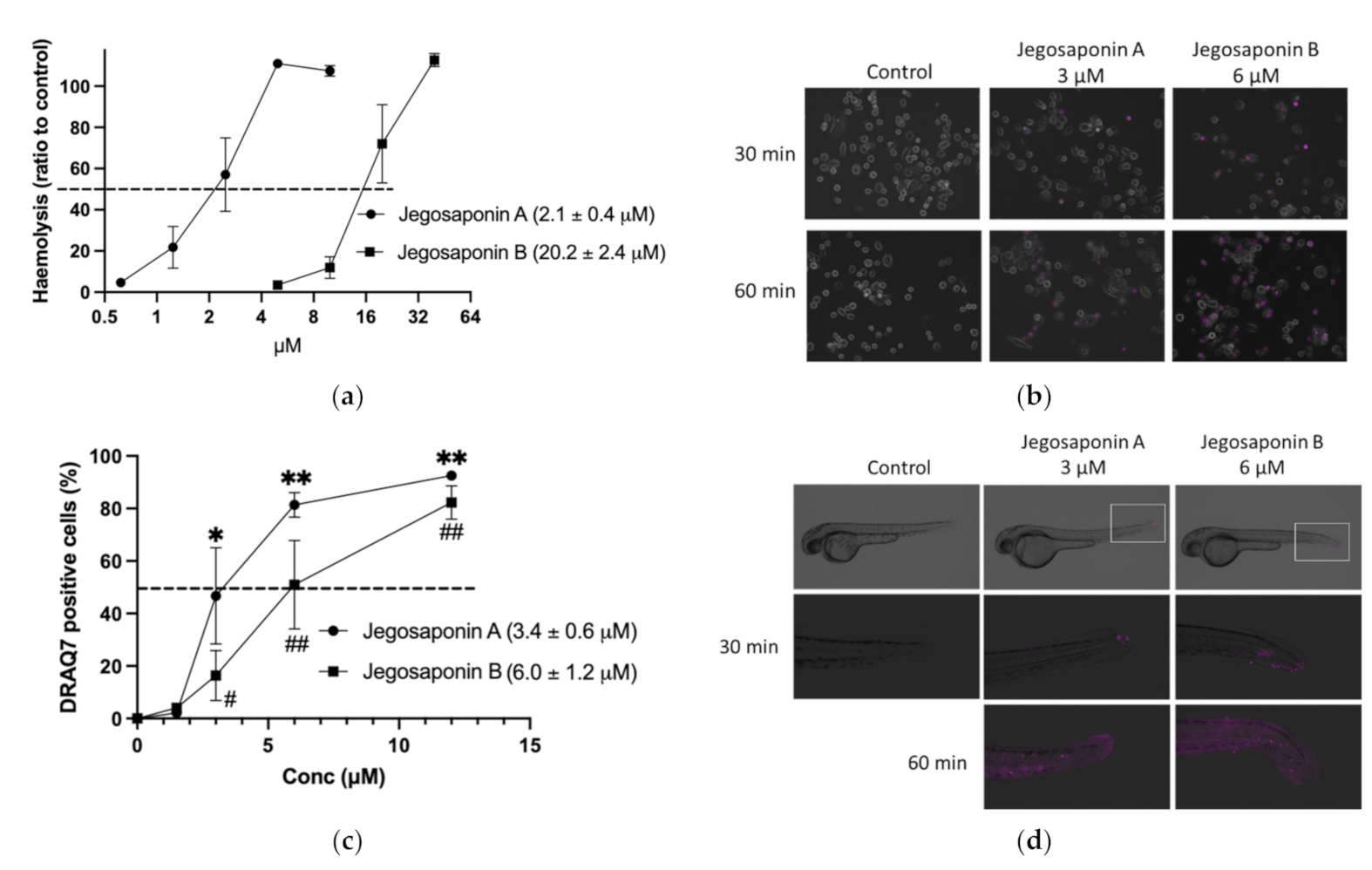
2.3. Cytotoxic Mechanisms of Jegosponin A and B
3. Discussion
4. Materials and Methods
4.1. Instruments
4.2. Sample Materials
4.3. Isolation Procedure
4.4. Cell Viability Assay
4.5. Zebrafish
4.6. Zebrafish Embryo Acute Toxicity Test (ZFET)
4.7. Caspase 3/7 Assay
4.8. Haemolysis Assay
4.9. DRAQ7 Membrane Permeability Assay
4.10. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chin, Y.W.; Balunas, M.J.; Chai, H.B.; Kinghorn, A.D. Drug discovery from natural sources. AAPS J. 2006, 8, E239–E253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cragg, G.M.; Newman, D.J. Natural products: A continuing source of novel drug leads. Biochim. Biophys. Acta 2013, 1830, 3670–3695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chensom, S.; Shimada, Y.; Nakayama, H.; Yoshida, K.; Kondo, T.; Katsuzaki, H.; Hasegawa, S.; Mishima, T. Determination of Anthocyanins and Antioxidants in ‘Titanbicus’ Edible Flowers In Vitro and In Vivo. Plant Foods Hum. Nutr. 2020, 75, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; An, L.; Li, Z.; Wang, X.; Wang, H.; Shi, L.; Bao, J.; Lan, X.; Zhang, E.; Lall, N.; et al. Structural elucidation of an immunological arabinan from the rhizomes of Ligusticum chuanxiong, a traditional Chinese medicine. Int. J. Biol. Macromol. 2021, 170, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Balkrishna, A.; Solleti, S.K.; Verma, S.; Varshney, A. Application of Humanized Zebrafish Model in the Suppression of SARS-CoV-2 Spike Protein Induced Pathology by Tri-Herbal Medicine Coronil via Cytokine Modulation. Molecules 2020, 25, 5091. [Google Scholar] [CrossRef] [PubMed]
- Park, K.R.; Kim, S.; Cho, M.; Kang, S.W.; Yun, H.M. Effects of PIN on Osteoblast Differentiation and Matrix Mineralization through Runt-Related Transcription Factor. Int. J. Mol. Sci. 2020, 21, 9579. [Google Scholar] [CrossRef] [PubMed]
- Min, B.S.; Oh, S.R.; Ahn, K.S.; Kim, J.H.; Lee, J.; Kim, D.Y.; Kim, E.H.; Lee, H.K. Anti-complement activity of norlignans and terpenes from the stem bark of Styrax japonica. Planta Med. 2004, 70, 1210–1215. [Google Scholar] [CrossRef] [PubMed]
- Yun, K.J.; Min, B.S.; Kim, J.Y.; Lee, K.T. Styraxoside A isolated from the stem bark of Styrax japonica inhibits lipopolysaccharide-induced expression of inducible nitric oxide synthase and cyclooxygenase-2 in RAW 264.7 cells by suppressing nuclear factor-kappa B activation. Biol. Pharm. Bull. 2007, 30, 139–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Lim, K.T. Phytoglycoprotein (38 kDa) induces cell cycle (G0/G1) arrest and apoptosis in HepG2 cells. J. Cell Biochem. 2011, 112, 3129–3139. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Lim, K.T. Modulatory effect of phytoglycoprotein (38 kDa) on cyclin D1/CDK4 in BNL CL.2 cells induced by N-methyl-N′-nitro-N-nitrosoguanidine. Naunyn Schmiedebergs Arch. Pharmacol. 2012, 385, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Lim, K.T. SJSZ glycoprotein (38 kDa) inhibits cell cycle and oxidative stress in N-methyl-N′-nitro-N-nitrosoguanidine-induced ICR mice. Anticancer Agents Med. Chem. 2013, 13, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Hill, A.J.; Teraoka, H.; Heideman, W.; Peterson, R.E. Zebrafish as a model vertebrate for investigating chemical toxicity. Toxicol. Sci. 2005, 86, 6–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawada, T.; Kuroyanagi, J.; Okazaki, F.; Taniguchi, M.; Nakayama, H.; Suda, N.; Abiko, S.; Kaneco, S.; Nishimura, N.; Shimada, Y. An Integrative Evaluation Method for the Biological Safety of Down and Feather Materials. Int. J. Mol. Sci. 2019, 20, 1434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busquet, F.; Strecker, R.; Rawlings, J.M.; Belanger, S.E.; Braunbeck, T.; Carr, G.J.; Cenijn, P.; Fochtman, P.; Gourmelon, A.; Hübler, N.; et al. OECD validation study to assess intra- and inter-laboratory reproducibility of the zebrafish embryo toxicity test for acute aquatic toxicity testing. Regul. Toxicol. Pharmacol. 2014, 69, 496–511. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, Y.; Yata, K.; Nomoto, T.; Ogiwara, T.; Watanabe, K.; Shintou, T.; Tsuboyama, A.; Okano, M.; Umemoto, N.; Zhang, Z.; et al. Identification of a novel indoline derivative for in vivo fluorescent imaging of blood-brain barrier disruption in animal models. ACS Chem. Neurosci. 2013, 4, 1183–1193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, K.; Nishimura, Y.; Nomoto, T.; Umemoto, N.; Zhang, Z.; Zhang, B.; Kuroyanagi, J.; Shimada, Y.; Shintou, T.; Okano, M.; et al. In vivo assessment of the permeability of the blood-brain barrier and blood-retinal barrier to fluorescent indoline derivatives in zebrafish. BMC Neurosci. 2012, 13, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshikawa, K.; Hirai, H.; Tanaka, M.; Arihara, S. Antisweet natural products. XV. Structures of Jegosaponins A-D from Styrax japonica Sieb. et Zucc. Chem. Pharm. Bull. 2000, 48, 1093–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waheed, A.; Barker, J.; Barton, S.J.; Owen, C.P.; Ahmed, S.; Carew, M.A. A novel steroidal saponin glycoside from Fagonia indica induces cell-selective apoptosis or necrosis in cancer cells. Eur. J. Pharm. Sci. 2012, 47, 464–473. [Google Scholar] [CrossRef] [PubMed]
- Voutquenne, L.; Lavaud, C.; Massiot, G.; Le Men-Olivier, L. Structure-activity relationships of haemolytic saponins. Pharm. Biol. 2002, 40, 253–262. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| RBC | PC-3 Cells | Zebrafish | |||
|---|---|---|---|---|---|
| Hemolysis HD50 | LC50 | DRAQ7 IC50 | LC50 | LC50 | |
| (1 h) | (48 h) | (1 h) | (120 h) | (24 h) | |
| Jegosaponin A | 2.1 μM | 0.7 μM | 3.4 μM | 0.5 μM | 0.7 μM |
| Jegosaponin B | 20.2 μM | 1.6 μM | 6.0 μM | 1.3 μM | 1.4 μM |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nishimura, M.; Fuchino, H.; Takayanagi, K.; Kawakami, H.; Nakayama, H.; Kawahara, N.; Shimada, Y. Toxicity of Jegosaponins A and B from Styrax japonica Siebold et al. Zuccarini in Prostate Cancer Cells and Zebrafish Embryos Resulting from Increased Membrane Permeability. Int. J. Mol. Sci. 2021, 22, 6354. https://doi.org/10.3390/ijms22126354
Nishimura M, Fuchino H, Takayanagi K, Kawakami H, Nakayama H, Kawahara N, Shimada Y. Toxicity of Jegosaponins A and B from Styrax japonica Siebold et al. Zuccarini in Prostate Cancer Cells and Zebrafish Embryos Resulting from Increased Membrane Permeability. International Journal of Molecular Sciences. 2021; 22(12):6354. https://doi.org/10.3390/ijms22126354
Chicago/Turabian StyleNishimura, Moe, Hiroyuki Fuchino, Kaoru Takayanagi, Hitomi Kawakami, Hiroko Nakayama, Nobuo Kawahara, and Yasuhito Shimada. 2021. "Toxicity of Jegosaponins A and B from Styrax japonica Siebold et al. Zuccarini in Prostate Cancer Cells and Zebrafish Embryos Resulting from Increased Membrane Permeability" International Journal of Molecular Sciences 22, no. 12: 6354. https://doi.org/10.3390/ijms22126354
APA StyleNishimura, M., Fuchino, H., Takayanagi, K., Kawakami, H., Nakayama, H., Kawahara, N., & Shimada, Y. (2021). Toxicity of Jegosaponins A and B from Styrax japonica Siebold et al. Zuccarini in Prostate Cancer Cells and Zebrafish Embryos Resulting from Increased Membrane Permeability. International Journal of Molecular Sciences, 22(12), 6354. https://doi.org/10.3390/ijms22126354