
FcRn as a Transporter for Nasal Delivery of Biologics: A Systematic Review

 , , and
, , and
Abstract
:1. Introduction
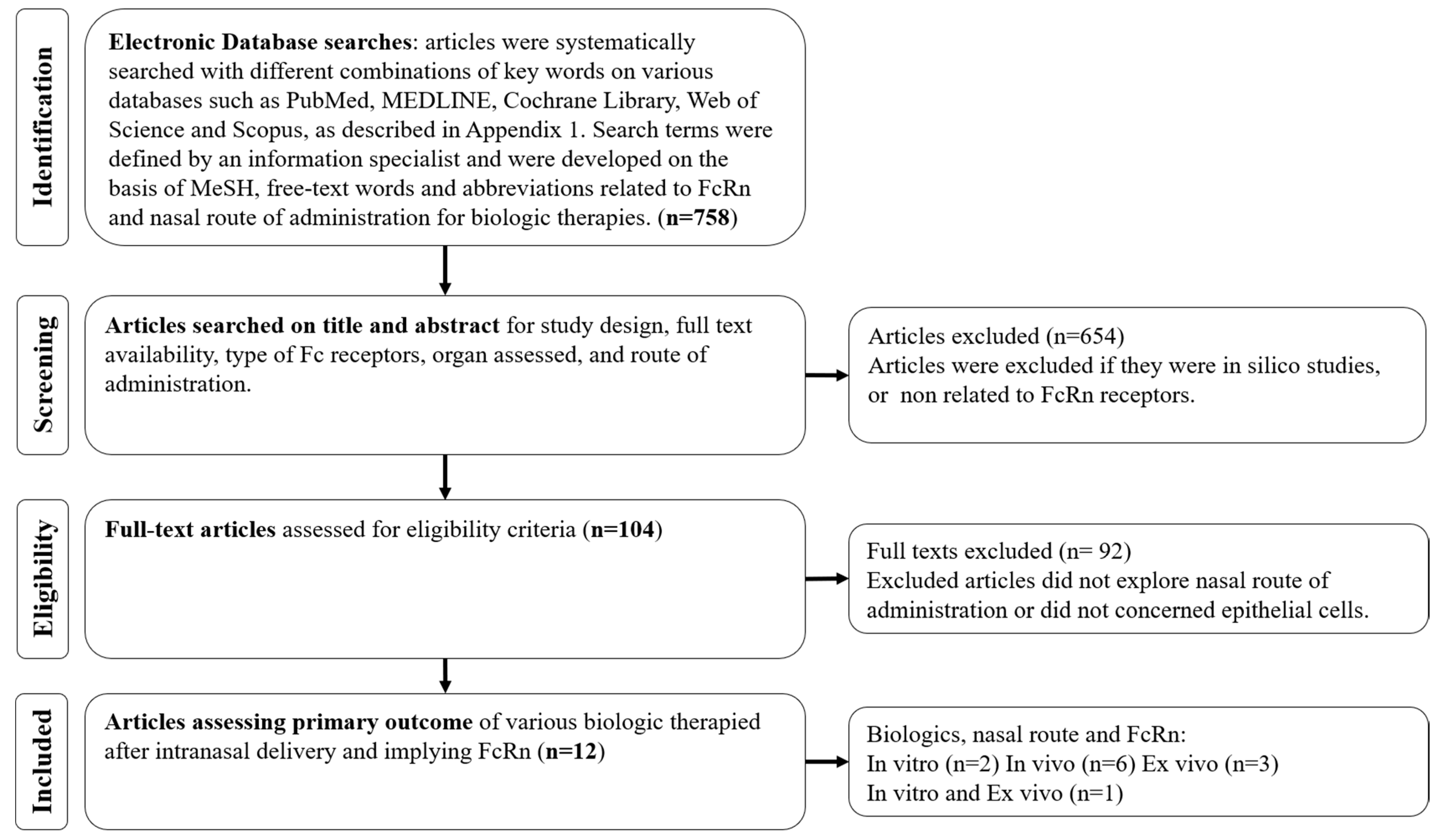
2. Methods
3. Results
3.1. Selection, Characteristics and Risk of Bias of the Included Studies
3.2. Expression of FcRn in the Upper Airway System
3.3. Biologics-Transcytosis Efficiency after Intranasal Delivery
3.4. Factors Influencing Transcytosis
3.5. Toxicity of Intranasal Delivery
3.6. Selected Studies Drawbacks
4. Discussion
4.1. FcRn Expression
4.2. Transcytosis Models
4.3. Factors Influencing Nasal Transcytosis
4.4. Biologics Integrity
5. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Simister, N.E.; Mostov, K.E. An Fc Receptor Structurally Related to MHC Class I Antigens. Nature 1989, 337, 184–187. [Google Scholar] [CrossRef]
- Rodewald, R.; Abrahamson, D.R. Receptor-Mediated Transport of IgG across the Intestinal Epithelium of the Neonatal Rat. Ciba Found. Symp. 1982, 209–232. [Google Scholar] [CrossRef]
- Rodewald, R.; Kraehenbuhl, J.P. Receptor-Mediated Transport of IgG. J. Cell Biol. 1984, 99, 159s–164s. [Google Scholar] [CrossRef]
- Simister, N.E.; Rees, A.R. Isolation and Characterization of an Fc Receptor from Neonatal Rat Small Intestine. Eur. J. Immunol. 1985, 15, 733–738. [Google Scholar] [CrossRef]
- Wallace, K.H.; Rees, A.R. Studies on the Immunoglobulin-G Fc-Fragment Receptor from Neonatal Rat Small Intestine. Biochem. J. 1980, 188, 9–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rath, T.; Kuo, T.T.; Baker, K.; Qiao, S.-W.; Kobayashi, K.; Yoshida, M.; Roopenian, D.; Fiebiger, E.; Lencer, W.I.; Blumberg, R.S. The Immunologic Functions of the Neonatal Fc Receptor for IgG. J. Clin. Immunol. 2013, 33 (Suppl. 1), 9–17. [Google Scholar] [CrossRef] [PubMed]
- Roopenian, D.C.; Akilesh, S. FcRn: The Neonatal Fc Receptor Comes of Age. Nat. Rev. Immunol. 2007, 7, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Ward, E.S.; Ober, R.J. Chapter 4: Multitasking by Exploitation of Intracellular Transport Functions the Many Faces of FcRn. Adv. Immunol. 2009, 103, 77–115. [Google Scholar] [CrossRef] [Green Version]
- Bequignon, E.; Dhommée, C.; Angely, C.; Thomas, L.; Bottier, M.; Escudier, E.; Isabey, D.; Coste, A.; Louis, B.; Papon, J.-F.; et al. FcRn-Dependent Transcytosis of Monoclonal Antibody in Human Nasal Epithelial Cells In Vitro: A Prerequisite for a New Delivery Route for Therapy? Int. J. Mol. Sci. 2019, 20, 1379. [Google Scholar] [CrossRef] [Green Version]
- Heidl, S.; Ellinger, I.; Niederberger, V.; Waltl, E.E.; Fuchs, R. Localization of the Human Neonatal Fc Receptor (FcRn) in Human Nasal Epithelium. Protoplasma 2016, 253, 1557–1564. [Google Scholar] [CrossRef]
- Spiekermann, G.M.; Finn, P.W.; Ward, E.S.; Dumont, J.; Dickinson, B.L.; Blumberg, R.S.; Lencer, W.I. Receptor-Mediated Immunoglobulin G Transport across Mucosal Barriers in Adult Life: Functional Expression of FcRn in the Mammalian Lung. J. Exp. Med. 2002, 196, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Bitonti, A.J.; Dumont, J.A.; Low, S.C.; Peters, R.T.; Kropp, K.E.; Palombella, V.J.; Stattel, J.M.; Lu, Y.; Tan, C.A.; Song, J.J.; et al. Pulmonary Delivery of an Erythropoietin Fc Fusion Protein in Non-Human Primates through an Immunoglobulin Transport Pathway. Proc. Natl. Acad. Sci. USA 2004, 101, 9763–9768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bitonti, A.J.; Dumont, J.A. Pulmonary Administration of Therapeutic Proteins Using an Immunoglobulin Transport Pathway. Adv. Drug Deliv. Rev. 2006, 58, 1106–1118. [Google Scholar] [CrossRef] [PubMed]
- Dumont, J.A.; Bitonti, A.J.; Clark, D.; Evans, S.; Pickford, M.; Newman, S.P. Delivery of an Erythropoietin-Fc Fusion Protein by Inhalation in Humans through an Immunoglobulin Transport Pathway. J. Aerosol Med. 2005, 18, 294–303. [Google Scholar] [CrossRef]
- Ladel, S.; Flamm, J.; Zadeh, A.S.; Filzwieser, D.; Walter, J.-C.; Schlossbauer, P.; Kinscherf, R.; Lischka, K.; Luksch, H.; Schindowski, K. Allogenic Fc Domain-Facilitated Uptake of IgG in Nasal Lamina Propria: Friend or Foe for Intranasal CNS Delivery? Pharmaceutics 2018, 10, 107. [Google Scholar] [CrossRef] [Green Version]
- Ladel, S.; Schlossbauer, P.; Flamm, J.; Luksch, H.; Mizaikoff, B.; Schindowski, K. Improved In Vitro Model for Intranasal Mucosal Drug Delivery: Primary Olfactory and Respiratory Epithelial Cells Compared with the Permanent Nasal Cell Line RPMI 2650. Pharmaceutics 2019, 11, 367. [Google Scholar] [CrossRef] [Green Version]
- Ghetie, V.; Ward, E.S. FcRn: The MHC Class I-Related Receptor That Is More than an IgG Transporter. Immunol. Today 1997, 18, 592–598. [Google Scholar] [CrossRef]
- Junghans, R.P.; Anderson, C.L. The Protection Receptor for IgG Catabolism Is the Beta2-Microglobulin-Containing Neonatal Intestinal Transport Receptor. Proc. Natl. Acad. Sci. USA 1996, 93, 5512–5516. [Google Scholar] [CrossRef] [Green Version]
- Wani, M.A.; Haynes, L.D.; Kim, J.; Bronson, C.L.; Chaudhury, C.; Mohanty, S.; Waldmann, T.A.; Robinson, J.M.; Anderson, C.L. Familial Hypercatabolic Hypoproteinemia Caused by Deficiency of the Neonatal Fc Receptor, FcRn, Due to a Mutant Beta2-Microglobulin Gene. Proc. Natl. Acad. Sci. USA 2006, 103, 5084–5089. [Google Scholar] [CrossRef] [Green Version]
- Lobo, E.D.; Hansen, R.J.; Balthasar, J.P. Antibody Pharmacokinetics and Pharmacodynamics. J. Pharm. Sci. 2004, 93, 2645–2668. [Google Scholar] [CrossRef]
- Nissim, A.; Chernajovsky, Y. Historical Development of Monoclonal Antibody Therapeutics. Handb. Exp. Pharmacol. 2008, 3–18. [Google Scholar] [CrossRef]
- Claypool, S.M.; Dickinson, B.L.; Wagner, J.S.; Johansen, F.-E.; Venu, N.; Borawski, J.A.; Lencer, W.I.; Blumberg, R.S. Bidirectional Transepithelial IgG Transport by a Strongly Polarized Basolateral Membrane Fcgamma-Receptor. Mol. Biol. Cell 2004, 15, 1746–1759. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, B.L.; Badizadegan, K.; Wu, Z.; Ahouse, J.C.; Zhu, X.; Simister, N.E.; Blumberg, R.S.; Lencer, W.I. Bidirectional FcRn-Dependent IgG Transport in a Polarized Human Intestinal Epithelial Cell Line. J. Clin. Investig. 1999, 104, 903–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firan, M.; Bawdon, R.; Radu, C.; Ober, R.J.; Eaken, D.; Antohe, F.; Ghetie, V.; Ward, E.S. The MHC Class I-Related Receptor, FcRn, Plays an Essential Role in the Maternofetal Transfer of Gamma-Globulin in Humans. Int. Immunol. 2001, 13, 993–1002. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, K.M.; Yoong, Y.; Simister, N.E. Bidirectional Transcytosis of IgG by the Rat Neonatal Fc Receptor Expressed in a Rat Kidney Cell Line: A System to Study Protein Transport across Epithelia. J. Cell Sci. 2000, 113 Pt 7, 1277–1285. [Google Scholar] [CrossRef]
- Bern, M.; Nilsen, J.; Ferrarese, M.; Sand, K.M.K.; Gjølberg, T.T.; Lode, H.E.; Davidson, R.J.; Camire, R.M.; Bækkevold, E.S.; Foss, S.; et al. An Engineered Human Albumin Enhances Half-Life and Transmucosal Delivery When Fused to Protein-Based Biologics. Sci. Transl. Med. 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Rawool, D.B.; Bitsaktsis, C.; Li, Y.; Gosselin, D.R.; Lin, Y.; Kurkure, N.V.; Metzger, D.W.; Gosselin, E.J. Utilization of Fc Receptors as a Mucosal Vaccine Strategy against an Intracellular Bacterium, Francisella Tularensis. J. Immunol. 2008, 180, 5548–5557. [Google Scholar] [CrossRef]
- Lu, L.; Palaniyandi, S.; Zeng, R.; Bai, Y.; Liu, X.; Wang, Y.; Pauza, C.D.; Roopenian, D.C.; Zhu, X. A Neonatal Fc Receptor-Targeted Mucosal Vaccine Strategy Effectively Induces HIV-1 Antigen-Specific Immunity to Genital Infection. J. Virol. 2011, 85, 10542–10553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, L.; Zeng, R.; Bai, Y.; Roopenian, D.C.; Zhu, X. Efficient Mucosal Vaccination Mediated by the Neonatal Fc Receptor. Nat. Biotechnol. 2011, 29, 158–163. [Google Scholar] [CrossRef] [Green Version]
- Ko, S.-Y.; Pegu, A.; Rudicell, R.S.; Yang, Z.; Joyce, M.G.; Chen, X.; Wang, K.; Bao, S.; Kraemer, T.D.; Rath, T.; et al. Enhanced Neonatal Fc Receptor Function Improves Protection against Primate SHIV Infection. Nature 2014, 514, 642–645. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Palaniyandi, S.; Zeng, R.; Tuo, W.; Roopenian, D.C.; Zhu, X. Transfer of IgG in the Female Genital Tract by MHC Class I-Related Neonatal Fc Receptor (FcRn) Confers Protective Immunity to Vaginal Infection. Proc. Natl. Acad. Sci. USA 2011, 108, 4388–4393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, M.; Kobayashi, K.; Kuo, T.T.; Bry, L.; Glickman, J.N.; Claypool, S.M.; Kaser, A.; Nagaishi, T.; Higgins, D.E.; Mizoguchi, E.; et al. Neonatal Fc Receptor for IgG Regulates Mucosal Immune Responses to Luminal Bacteria. J. Clin. Investig. 2006, 116, 2142–2151. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Masuda, A.; Kuo, T.T.; Kobayashi, K.; Claypool, S.M.; Takagawa, T.; Kutsumi, H.; Azuma, T.; Lencer, W.I.; Blumberg, R.S. IgG Transport across Mucosal Barriers by Neonatal Fc Receptor for IgG and Mucosal Immunity. Springer Semin. Immunopathol. 2006, 28, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Ladinsky, M.S.; Huey-Tubman, K.E.; Bjorkman, P.J. Electron Tomography of Late Stages of FcRn-Mediated Antibody Transcytosis in Neonatal Rat Small Intestine. Mol. Biol. Cell 2012, 23, 2537–2545. [Google Scholar] [CrossRef] [PubMed]
- Röhm, M.; Carle, S.; Maigler, F.; Flamm, J.; Kramer, V.; Mavoungou, C.; Schmid, O.; Schindowski, K. A Comprehensive Screening Platform for Aerosolizable Protein Formulations for Intranasal and Pulmonary Drug Delivery. Int. J. Pharm. 2017, 532, 537–546. [Google Scholar] [CrossRef]
- Pridgen, E.M.; Alexis, F.; Kuo, T.T.; Levy-Nissenbaum, E.; Karnik, R.; Blumberg, R.S.; Langer, R.; Farokhzad, O.C. Transepithelial Transport of Fc-Targeted Nanoparticles by the Neonatal Fc Receptor for Oral Delivery. Sci. Transl. Med. 2013, 5, 213ra167. [Google Scholar] [CrossRef] [Green Version]
- Martins, J.P.; Kennedy, P.J.; Santos, H.A.; Barrias, C.; Sarmento, B. A Comprehensive Review of the Neonatal Fc Receptor and Its Application in Drug Delivery. Pharmacol. Ther. 2016, 161, 22–39. [Google Scholar] [CrossRef]
- England, R.J.; Homer, J.J.; Knight, L.C.; Ell, S.R. Nasal PH Measurement: A Reliable and Repeatable Parameter. Clin. Otolaryngol. Allied Sci. 1999, 24, 67–68. [Google Scholar] [CrossRef] [Green Version]
- Chaudhury, C.; Mehnaz, S.; Robinson, J.M.; Hayton, W.L.; Pearl, D.K.; Roopenian, D.C.; Anderson, C.L. The Major Histocompatibility Complex-Related Fc Receptor for IgG (FcRn) Binds Albumin and Prolongs Its Lifespan. J. Exp. Med. 2003, 197, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Andersen, J.T.; Dee Qian, J.; Sandlie, I. The Conserved Histidine 166 Residue of the Human Neonatal Fc Receptor Heavy Chain Is Critical for the PH-Dependent Binding to Albumin. Eur. J. Immunol. 2006, 36, 3044–3051. [Google Scholar] [CrossRef]
- Oganesyan, V.; Damschroder, M.M.; Cook, K.E.; Li, Q.; Gao, C.; Wu, H.; Dall’Acqua, W.F. Structural Insights into Neonatal Fc Receptor-Based Recycling Mechanisms. J. Biol. Chem. 2014, 289, 7812–7824. [Google Scholar] [CrossRef] [Green Version]
- Köhler, G.; Milstein, C. Continuous Cultures of Fused Cells Secreting Antibody of Predefined Specificity. Nature 1975, 256, 495–497. [Google Scholar] [CrossRef]
- Brekke, O.H.; Sandlie, I. Therapeutic Antibodies for Human Diseases at the Dawn of the Twenty-First Century. Nat. Rev. Drug Discov. 2003, 2, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Ward, E.S.; Devanaboyina, S.C.; Ober, R.J. Targeting FcRn for the Modulation of Antibody Dynamics. Mol. Immunol. 2015, 67, 131–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Wang, E.Q.; Balthasar, J.P. Monoclonal Antibody Pharmacokinetics and Pharmacodynamics. Clin. Pharmacol. Ther. 2008, 84, 548–558. [Google Scholar] [CrossRef] [PubMed]
- Samson, G.; García de la Calera, A.; Dupuis-Girod, S.; Faure, F.; Decullier, E.; Paintaud, G.; Vignault, C.; Scoazec, J.-Y.; Pivot, C.; Plauchu, H.; et al. Ex Vivo Study of Bevacizumab Transport through Porcine Nasal Mucosa. Eur. J. Pharm. Biopharm. 2012, 80, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Low, S.C.; Nunes, S.L.; Bitonti, A.J.; Dumont, J.A. Oral and Pulmonary Delivery of FSH-Fc Fusion Proteins via Neonatal Fc Receptor-Mediated Transcytosis. Hum. Reprod. Oxf. Engl. 2005, 20, 1805–1813. [Google Scholar] [CrossRef] [Green Version]
- Benedict, C.; Hallschmid, M.; Schmitz, K.; Schultes, B.; Ratter, F.; Fehm, H.L.; Born, J.; Kern, W. Intranasal Insulin Improves Memory in Humans: Superiority of Insulin Aspart. Neuropsychopharmacology 2007, 32, 239–243. [Google Scholar] [CrossRef] [Green Version]
- Benedict, C.; Hallschmid, M.; Hatke, A.; Schultes, B.; Fehm, H.L.; Born, J.; Kern, W. Intranasal Insulin Improves Memory in Humans. Psychoneuroendocrinology 2004, 29, 1326–1334. [Google Scholar] [CrossRef]
- Benedict, C.; Frey, W.H.; Schiöth, H.B.; Schultes, B.; Born, J.; Hallschmid, M. Intranasal Insulin as a Therapeutic Option in the Treatment of Cognitive Impairments. Exp. Gerontol. 2011, 46, 112–115. [Google Scholar] [CrossRef]
- Heni, M.; Wagner, R.; Kullmann, S.; Veit, R.; Mat Husin, H.; Linder, K.; Benkendorff, C.; Peter, A.; Stefan, N.; Häring, H.-U.; et al. Central Insulin Administration Improves Whole-Body Insulin Sensitivity via Hypothalamus and Parasympathetic Outputs in Men. Diabetes 2014, 63, 4083–4088. [Google Scholar] [CrossRef] [Green Version]
- Kullmann, S.; Frank, S.; Heni, M.; Ketterer, C.; Veit, R.; Häring, H.-U.; Fritsche, A.; Preissl, H. Intranasal Insulin Modulates Intrinsic Reward and Prefrontal Circuitry of the Human Brain in Lean Women. Neuroendocrinology 2013, 97, 176–182. [Google Scholar] [CrossRef]
- Balin, B.J.; Broadwell, R.D.; Salcman, M.; el-Kalliny, M. Avenues for Entry of Peripherally Administered Protein to the Central Nervous System in Mouse, Rat, and Squirrel Monkey. J. Comp. Neurol. 1986, 251, 260–280. [Google Scholar] [CrossRef]
- Warnken, Z.N.; Smyth, H.D.C.; Watts, A.B.; Weitman, S.; Kuhn, J.G.; Williams, R.O. Formulation and Device Design to Increase Nose to Brain Drug Delivery. J. Drug Deliv. Sci. Technol. 2016, 35, 213–222. [Google Scholar] [CrossRef]
- Stote, R.; Miller, M.; Marbury, T.; Shi, L.; Strange, P. Enhanced Absorption of NasulinTM, an Ultrarapid-Acting Intranasal Insulin Formulation, Using Single Nostril Administration in Normal Subjects. J. Diabetes Sci. Technol. 2011, 5, 113–119. [Google Scholar] [CrossRef] [Green Version]
- Guilleminault, L.; Azzopardi, N.; Arnoult, C.; Sobilo, J.; Hervé, V.; Montharu, J.; Guillon, A.; Andres, C.; Herault, O.; Le Pape, A.; et al. Fate of Inhaled Monoclonal Antibodies after the Deposition of Aerosolized Particles in the Respiratory System. J. Control Release 2014, 196, 344–354. [Google Scholar] [CrossRef] [PubMed]
- Engelhardt, L.; Röhm, M.; Mavoungou, C.; Schindowski, K.; Schafmeister, A.; Simon, U. First Steps to Develop and Validate a CFPD Model in Order to Support the Design of Nose-to-Brain Delivered Biopharmaceuticals. Pharm. Res. 2016, 33, 1337–1350. [Google Scholar] [CrossRef] [PubMed]
- Möller, W.; Schuschnig, U.; Bartenstein, P.; Meyer, G.; Häussinger, K.; Schmid, O.; Becker, S. Drug Delivery to Paranasal Sinuses Using Pulsating Aerosols. J. Aerosol Med. Pulm. Drug Deliv. 2014, 27, 255–263. [Google Scholar] [CrossRef]
- Coates, A.L. Guiding Aerosol Deposition in the Lung. N. Engl. J. Med. 2008, 358, 304–305. [Google Scholar] [CrossRef] [PubMed]
- Lochhead, J.J.; Thorne, R.G. Intranasal Delivery of Biologics to the Central Nervous System. Adv. Drug Deliv. Rev. 2012, 64, 614–628. [Google Scholar] [CrossRef]
- Kumar, N.N.; Lochhead, J.J.; Pizzo, M.E.; Nehra, G.; Boroumand, S.; Greene, G.; Thorne, R.G. Delivery of Immunoglobulin G Antibodies to the Rat Nervous System Following Intranasal Administration: Distribution, Dose-Response, and Mechanisms of Delivery. J. Control Release 2018, 286, 467–484. [Google Scholar] [CrossRef]
- Siddaway, A.P.; Wood, A.M.; Hedges, L.V. How to Do a Systematic Review: A Best Practice Guide for Conducting and Reporting Narrative Reviews, Meta-Analyses, and Meta-Syntheses. Annu. Rev. Psychol. 2019, 70, 747–770. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- Moher, D.; Shamseer, L.; Clarke, M.; Ghersi, D.; Liberati, A.; Petticrew, M.; Shekelle, P.; Stewart, L.A. PRISMA-P Group Preferred Reporting Items for Systematic Review and Meta-Analysis Protocols (PRISMA-P) 2015 Statement. Syst. Rev. 2015, 4, 1. [Google Scholar] [CrossRef] [Green Version]
- Higgins, J.P.T.; Altman, D.G.; Gøtzsche, P.C.; Jüni, P.; Moher, D.; Oxman, A.D.; Savovic, J.; Schulz, K.F.; Weeks, L.; Sterne, J.A.C.; et al. The Cochrane Collaboration’s Tool for Assessing Risk of Bias in Randomised Trials. BMJ 2011, 343, d5928. [Google Scholar] [CrossRef] [Green Version]
- Viswanathan, M.; Berkman, N.D. Development of the RTI Item Bank on Risk of Bias and Precision of Observational Studies. J. Clin. Epidemiol. 2012, 65, 163–178. [Google Scholar] [CrossRef] [PubMed]
- Sterne, J.A.C.; Savović, J.; Page, M.J.; Elbers, R.G.; Blencowe, N.S.; Boutron, I.; Cates, C.J.; Cheng, H.-Y.; Corbett, M.S.; Eldridge, S.M.; et al. RoB 2: A Revised Tool for Assessing Risk of Bias in Randomised Trials. BMJ 2019, 366, l4898. [Google Scholar] [CrossRef] [Green Version]
- Ouzzani, M.; Hammady, H.; Fedorowicz, Z.; Elmagarmid, A. Rayyan-a Web and Mobile App for Systematic Reviews. Syst. Rev. 2016, 5, 210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bitsaktsis, C.; Babadjanova, Z.; Gosselin, E.J. In Vivo Mechanisms Involved in Enhanced Protection Utilizing an Fc Receptor-Targeted Mucosal Vaccine Platform in a Bacterial Vaccine and Challenge Model. Infect. Immun. 2015, 83, 77–89. [Google Scholar] [CrossRef] [Green Version]
- de Taeye, S.W.; Rispens, T.; Vidarsson, G. The Ligands for Human IgG and Their Effector Functions. Antibodies Basel Switz. 2019, 8, 30. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.-D.; Wang, W.-H.; Li, S.-X.; Jia, S.; Zhang, X.-F.; Cao, T.-T. Localization of Neonatal Fc Receptor for IgG in Aggregated Lymphoid Nodules Area in Abomasum of Bactrian Camels (Camelus Bactrianus) of Different Ages. BMC Vet. Res. 2016, 12, 237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hervé, V.; Rabbe, N.; Guilleminault, L.; Paul, F.; Schlick, L.; Azzopardi, N.; Duruisseaux, M.; Fouquenet, D.; Montharu, J.; Redini, F.; et al. VEGF Neutralizing Aerosol Therapy in Primary Pulmonary Adenocarcinoma with K-Ras Activating-Mutations. mAbs 2014, 6, 1638–1648. [Google Scholar] [CrossRef] [Green Version]
- Respaud, R.; Marchand, D.; Pelat, T.; Tchou-Wong, K.-M.; Roy, C.J.; Parent, C.; Cabrera, M.; Guillemain, J.; Mac Loughlin, R.; Levacher, E.; et al. Development of a Drug Delivery System for Efficient Alveolar Delivery of a Neutralizing Monoclonal Antibody to Treat Pulmonary Intoxication to Ricin. J. Control Release 2016, 234, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Respaud, R.; Vecellio, L.; Diot, P.; Heuzé-Vourc’h, N. Nebulization as a Delivery Method for MAbs in Respiratory Diseases. Expert Opin. Drug Deliv. 2015, 12, 1027–1039. [Google Scholar] [CrossRef] [PubMed]
- Cianga, C.; Cianga, P.; Plamadeala, P.; Amalinei, C. Nonclassical Major Histocompatibility Complex I-like Fc Neonatal Receptor (FcRn) Expression in Neonatal Human Tissues. Hum. Immunol. 2011, 72, 1176–1187. [Google Scholar] [CrossRef]
- Stirling, C.M.A.; Charleston, B.; Takamatsu, H.; Claypool, S.; Lencer, W.; Blumberg, R.S.; Wileman, T.E. Characterization of the Porcine Neonatal Fc Receptor--Potential Use for Trans-Epithelial Protein Delivery. Immunology 2005, 114, 542–553. [Google Scholar] [CrossRef]
- Sockolosky, J.T.; Szoka, F.C. The Neonatal Fc Receptor, FcRn, as a Target for Drug Delivery and Therapy. Adv. Drug Deliv. Rev. 2015, 91, 109–124. [Google Scholar] [CrossRef] [Green Version]
- Wada, R.; Matsui, M.; Kawasaki, N. Influence of N-Glycosylation on Effector Functions and Thermal Stability of Glycoengineered IgG1 Monoclonal Antibody with Homogeneous Glycoforms. mAbs 2019, 11, 350–372. [Google Scholar] [CrossRef]
- Gao, X.; Ji, J.A.; Veeravalli, K.; Wang, Y.J.; Zhang, T.; Mcgreevy, W.; Zheng, K.; Kelley, R.F.; Laird, M.W.; Liu, J.; et al. Effect of Individual Fc Methionine Oxidation on FcRn Binding: Met252 Oxidation Impairs FcRn Binding More Profoundly than Met428 Oxidation. J. Pharm. Sci. 2015, 104, 368–377. [Google Scholar] [CrossRef]
- Stracke, J.; Emrich, T.; Rueger, P.; Schlothauer, T.; Kling, L.; Knaupp, A.; Hertenberger, H.; Wolfert, A.; Spick, C.; Lau, W.; et al. A Novel Approach to Investigate the Effect of Methionine Oxidation on Pharmacokinetic Properties of Therapeutic Antibodies. mAbs 2014, 6, 1229–1242. [Google Scholar] [CrossRef] [Green Version]
- Bai, S.; Yang, T.; Abbruscato, T.J.; Ahsan, F. Evaluation of Human Nasal RPMI 2650 Cells Grown at an Air-Liquid Interface as a Model for Nasal Drug Transport Studies. J. Pharm. Sci. 2008, 97, 1165–1178. [Google Scholar] [CrossRef] [PubMed]
- Kreft, M.E.; Jerman, U.D.; Lasič, E.; Lanišnik Rižner, T.; Hevir-Kene, N.; Peternel, L.; Kristan, K. The Characterization of the Human Nasal Epithelial Cell Line RPMI 2650 under Different Culture Conditions and Their Optimization for an Appropriate in Vitro Nasal Model. Pharm. Res. 2015, 32, 665–679. [Google Scholar] [CrossRef] [PubMed]
- Franz-Montan, M.; Serpe, L.; Martinelli, C.C.M.; da Silva, C.B.; Santos, C.P.D.; Novaes, P.D.; Volpato, M.C.; de Paula, E.; Lopez, R.F.V.; Groppo, F.C. Evaluation of Different Pig Oral Mucosa Sites as Permeability Barrier Models for Drug Permeation Studies. Eur. J. Pharm. Sci. 2016, 81, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Tulinski, P.; Fluit, A.C.; van Putten, J.P.M.; de Bruin, A.; Glorieux, S.; Wagenaar, J.A.; Duim, B. An Ex Vivo Porcine Nasal Mucosa Explants Model to Study MRSA Colonization. PLoS ONE 2013, 8, e53783. [Google Scholar] [CrossRef] [Green Version]
- Coste, A.; Brugel, L.; Maître, B.; Boussat, S.; Papon, J.F.; Wingerstmann, L.; Peynègre, R.; Escudier, E. Inflammatory Cells as Well as Epithelial Cells in Nasal Polyps Express Vascular Endothelial Growth Factor. Eur. Respir. J. 2000, 15, 367–372. [Google Scholar] [CrossRef] [Green Version]
- Ramanathan, M.; Lee, W.-K.; Dubin, M.G.; Lin, S.; Spannhake, E.W.; Lane, A.P. Sinonasal Epithelial Cell Expression of Toll-like Receptor 9 Is Decreased in Chronic Rhinosinusitis with Polyps. Am. J. Rhinol. 2007, 21, 110–116. [Google Scholar] [CrossRef]
- Igawa, T.; Ishii, S.; Tachibana, T.; Maeda, A.; Higuchi, Y.; Shimaoka, S.; Moriyama, C.; Watanabe, T.; Takubo, R.; Doi, Y.; et al. Antibody Recycling by Engineered PH-Dependent Antigen Binding Improves the Duration of Antigen Neutralization. Nat. Biotechnol. 2010, 28, 1203–1207. [Google Scholar] [CrossRef]
- Datta-Mannan, A.; Wroblewski, V.J. Application of FcRn Binding Assays to Guide MAb Development. Drug Metab. Dispos. Biol. Fate Chem. 2014, 42, 1867–1872. [Google Scholar] [CrossRef] [Green Version]
- Neuber, T.; Frese, K.; Jaehrling, J.; Jäger, S.; Daubert, D.; Felderer, K.; Linnemann, M.; Höhne, A.; Kaden, S.; Kölln, J.; et al. Characterization and Screening of IgG Binding to the Neonatal Fc Receptor. mAbs 2014, 6, 928–942. [Google Scholar] [CrossRef] [Green Version]
- Yeung, Y.A.; Leabman, M.K.; Marvin, J.S.; Qiu, J.; Adams, C.W.; Lien, S.; Starovasnik, M.A.; Lowman, H.B. Engineering Human IgG1 Affinity to Human Neonatal Fc Receptor: Impact of Affinity Improvement on Pharmacokinetics in Primates. J. Immunol. 2009, 182, 7663–7671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egli, J.; Schlothauer, T.; Spick, C.; Seeber, S.; Singer, T.; Odermatt, A.; Iglesias, A. The Binding of Human IgG to Minipig FcγRs—Implications for Preclinical Assessment of Therapeutic Antibodies. Pharm. Res. 2019, 36, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ober, R.J.; Radu, C.G.; Ghetie, V.; Ward, E.S. Differences in Promiscuity for Antibody-FcRn Interactions across Species: Implications for Therapeutic Antibodies. Int. Immunol. 2001, 13, 1551–1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, W.L.; West, A.P.; Gan, L.; Bjorkman, P.J. Crystal Structure at 2.8 A of an FcRn/Heterodimeric Fc Complex: Mechanism of PH-Dependent Binding. Mol. Cell 2001, 7, 867–877. [Google Scholar] [CrossRef]
- Gurbaxani, B.; Dostalek, M.; Gardner, I. Are Endosomal Trafficking Parameters Better Targets for Improving MAb Pharmacokinetics than FcRn Binding Affinity? Mol. Immunol. 2013, 56, 660–674. [Google Scholar] [CrossRef]
- Valente, D.; Mauriac, C.; Schmidt, T.; Focken, I.; Beninga, J.; Mackness, B.; Qiu, H.; Vicat, P.; Kandira, A.; Radošević, K.; et al. Pharmacokinetics of Novel Fc-Engineered Monoclonal and Multispecific Antibodies in Cynomolgus Monkeys and Humanized FcRn Transgenic Mouse Models. mAbs 2020, 12, 1829337. [Google Scholar] [CrossRef]
- Vaccaro, C.; Bawdon, R.; Wanjie, S.; Ober, R.J.; Ward, E.S. Divergent Activities of an Engineered Antibody in Murine and Human Systems Have Implications for Therapeutic Antibodies. Proc. Natl. Acad. Sci. USA 2006, 103, 18709–18714. [Google Scholar] [CrossRef] [Green Version]
- Jacobsen, B.; Hill, M.; Reynaud, L.; Hey, A.; Barrow, P. FcRn Expression on Placenta and Fetal Jejunum during Early, Mid-, and Late Gestation in Minipigs. Toxicol. Pathol. 2016, 44, 486–491. [Google Scholar] [CrossRef] [Green Version]
- Andersen, J.T.; Dalhus, B.; Viuff, D.; Ravn, B.T.; Gunnarsen, K.S.; Plumridge, A.; Bunting, K.; Antunes, F.; Williamson, R.; Athwal, S.; et al. Extending Serum Half-Life of Albumin by Engineering Neonatal Fc Receptor (FcRn) Binding. J. Biol. Chem. 2014, 289, 13492–13502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsen, J.; Bern, M.; Sand, K.M.K.; Grevys, A.; Dalhus, B.; Sandlie, I.; Andersen, J.T. Human and Mouse Albumin Bind Their Respective Neonatal Fc Receptors Differently. Sci. Rep. 2018, 8, 14648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, P.J.; Lazar, G.A. Next Generation Antibody Drugs: Pursuit of the “High-Hanging Fruit”. Nat. Rev. Drug Discov. 2018, 17, 197–223. [Google Scholar] [CrossRef]
- Lee, C.-H.; Kang, T.H.; Godon, O.; Watanabe, M.; Delidakis, G.; Gillis, C.M.; Sterlin, D.; Hardy, D.; Cogné, M.; Macdonald, L.E.; et al. An Engineered Human Fc Domain That Behaves like a PH-Toggle Switch for Ultra-Long Circulation Persistence. Nat. Commun. 2019, 10, 5031. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Balthasar, J.P. FcRn Expression in Wildtype Mice, Transgenic Mice, and in Human Tissues. Biomolecules 2018, 8, 115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proetzel, G.; Wiles, M.V.; Roopenian, D.C. Genetically Engineered Humanized Mouse Models for Preclinical Antibody Studies. BioDrugs Clin. Immunother. Biopharm. Gene Ther. 2014, 28, 171–180. [Google Scholar] [CrossRef] [Green Version]
- Roopenian, D.C.; Christianson, G.J.; Proetzel, G.; Sproule, T.J. Human FcRn Transgenic Mice for Pharmacokinetic Evaluation of Therapeutic Antibodies. Methods Mol. Biol. Clifton NJ 2016, 1438, 103–114. [Google Scholar] [CrossRef]
- Avery, L.B.; Wang, M.; Kavosi, M.S.; Joyce, A.; Kurz, J.C.; Fan, Y.-Y.; Dowty, M.E.; Zhang, M.; Zhang, Y.; Cheng, A.; et al. Utility of a Human FcRn Transgenic Mouse Model in Drug Discovery for Early Assessment and Prediction of Human Pharmacokinetics of Monoclonal Antibodies. mAbs 2016, 8, 1064–1078. [Google Scholar] [CrossRef] [Green Version]
- Jadhav, K.; Gambhire, M.; Shaikh, I.; Kadam, V.; Pisal, S. Nasal Drug Delivery System-Factors Affecting and Applications. Curr. Drug Ther. 2007, 2, 27–38. [Google Scholar] [CrossRef]
- Dellamary, L.; Smith, D.J.; Bloom, A.; Bot, S.; Guo, G.-R.; Deshmuk, H.; Costello, M.; Bot, A. Rational Design of Solid Aerosols for Immunoglobulin Delivery by Modulation of Aerodynamic and Release Characteristics. J. Control Release 2004, 95, 489–500. [Google Scholar] [CrossRef]
- Schüle, S.; Schulz-Fademrecht, T.; Garidel, P.; Bechtold-Peters, K.; Frieb, W. Stabilization of IgG1 in Spray-Dried Powders for Inhalation. Eur. J. Pharm. Biopharm. 2008, 69, 793–807. [Google Scholar] [CrossRef]
- Bertolotti-Ciarlet, A.; Wang, W.; Lownes, R.; Pristatsky, P.; Fang, Y.; McKelvey, T.; Li, Y.; Li, Y.; Drummond, J.; Prueksaritanont, T.; et al. Impact of Methionine Oxidation on the Binding of Human IgG1 to Fc Rn and Fc Gamma Receptors. Mol. Immunol. 2009, 46, 1878–1882. [Google Scholar] [CrossRef]
- Wang, W.; Vlasak, J.; Li, Y.; Pristatsky, P.; Fang, Y.; Pittman, T.; Roman, J.; Wang, Y.; Prueksaritanont, T.; Ionescu, R. Impact of Methionine Oxidation in Human IgG1 Fc on Serum Half-Life of Monoclonal Antibodies. Mol. Immunol. 2011, 48, 860–866. [Google Scholar] [CrossRef]
- Bien-Ly, N.; Boswell, C.A.; Jeet, S.; Beach, T.G.; Hoyte, K.; Luk, W.; Shihadeh, V.; Ulufatu, S.; Foreman, O.; Lu, Y.; et al. Lack of Widespread BBB Disruption in Alzheimer’s Disease Models: Focus on Therapeutic Antibodies. Neuron 2015, 88, 289–297. [Google Scholar] [CrossRef] [Green Version]
- Deli, M.A. Potential Use of Tight Junction Modulators to Reversibly Open Membranous Barriers and Improve Drug Delivery. Biochim. Biophys. Acta 2009, 1788, 892–910. [Google Scholar] [CrossRef] [Green Version]
- Yabluchanskiy, A.; Ma, Y.; Iyer, R.P.; Hall, M.E.; Lindsey, M.L. Matrix Metalloproteinase-9: Many Shades of Function in Cardiovascular Disease. Physiol. Bethesda Md 2013, 28, 391–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hackel, D.; Krug, S.M.; Sauer, R.-S.; Mousa, S.A.; Böcker, A.; Pflücke, D.; Wrede, E.-J.; Kistner, K.; Hoffmann, T.; Niedermirtl, B.; et al. Transient Opening of the Perineurial Barrier for Analgesic Drug Delivery. Proc. Natl. Acad. Sci. USA 2012, 109, E2018–E2027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gueye, Y.; Ferhat, L.; Sbai, O.; Bianco, J.; Ould-Yahoui, A.; Bernard, A.; Charrat, E.; Chauvin, J.-P.; Risso, J.-J.; Féron, F.; et al. Trafficking and Secretion of Matrix Metalloproteinase-2 in Olfactory Ensheathing Glial Cells: A Role in Cell Migration? Glia 2011, 59, 750–770. [Google Scholar] [CrossRef] [PubMed]
- Lochhead, J.J.; Wolak, D.J.; Pizzo, M.E.; Thorne, R.G. Rapid Transport within Cerebral Perivascular Spaces Underlies Widespread Tracer Distribution in the Brain after Intranasal Administration. J. Cereb. Blood Flow Metab. 2015, 35, 371–381. [Google Scholar] [CrossRef]
- Marttin, E.; Verhoef, J.C.; Cullander, C.; Romeijn, S.G.; Nagelkerke, J.F.; Merkus, F.W. Confocal Laser Scanning Microscopic Visualization of the Transport of Dextrans after Nasal Administration to Rats: Effects of Absorption Enhancers. Pharm. Res. 1997, 14, 631–637. [Google Scholar] [CrossRef]


{kind=link}
| First Author Date | Design | Species | Intervention | Application | Biotherapy Tested | Dose Administered | Concentration | Formulation | Primary Outcome | Blocking FcRn |
|---|---|---|---|---|---|---|---|---|---|---|
| Rawool [27] * 2008, India | In vivo | Mice FcRn WT | Assess FcRn-mediated mucosal vaccine delivery with an Ft model | Vaccine delivery against Ft | mAb-iFT (fusion of iFT and IgG) | 2 × 107 iFT/mAB-iFT/F(ab′)2-iFT organisms | 2 × 107 CFU/20 µL | PBS | FcRn-targeted immunogen enhanced immunogen-specific IgA production and protection against subsequent infection. It is a highly effective vaccination strategy against Ft. | Yes |
| Lu [28] * 2011, USA | In vivo | Mice FcRn WT and KO | Assess ability of FcRn to deliver Gag-Fc fusion protein in a HIV model | Vaccine delivery against VIH | HIV Gag-Fc fusion protein | 20 µg | 1 mg/mL | PBS | FcRn-targeted mucosal immunization was effective at inducing Gag specific Ab responses in serum or mucosal secretions, and high levels of stable immune memory were obtained. | Yes |
| Ye [29] * 2011, USA | In vivo | Mice FcRn WT and KO | Assess FcRn-mediated mucosal vaccine delivery with an HSV-2 model | Vaccine delivery against HSV | gD-Fc/wt (HSV-2 gD fused with an IgG Fc fragment) | 20 µg | 1 mg/mL | PBS | Intranasal immunization with an engineered fused protein resulted in complete protection of wild-type, but not FcRn KO, mice that were intravaginally challenged with virulent HSV-2. | Yes |
| Bitsatksis [69] * 2015, USA | In vivo | Mice FcRn WT | Assess FcRn-mediated mucosal vaccine delivery with an Ft model | Vaccine delivery against Ft | mAb-iFT IC (fusion of iFT and IgG) | 2 × 107 CFU mAb-iFT IC organisms | NA | PNS | FcRn targeting increases the frequency and activation status of DCs in the lungs of immunized mice and mediates the generation of Ft-specific effector memory CD4+ T cells. | No |
| Kumar [61] * 2018, USA | In vivo | Rats FcRn WT | Assess CNS IgG distribution after intranasal administration | Intracerebral mAb delivery | Radiolabeled Ab: [125I]-IgG | 50 µg to 2.5 mg for [125I]-IgG | 1-20-50 mg/mL | PBS | [125I]-IgG concentrations in the CNS was higher following intranasal delivery compared to intra-arterial delivery for doses producing similar endpoint blood concentrations. | No |
| Fluorescently labeled Ab: AF488-IgG | 0.7 mg for AF488-IgG | 30 mg/mL | ||||||||
| Bern [26] * 2020, Norway | In vivo | Mice FcRn WT and KO hFcRn | Assess ability of FcRn for delivery of albumin-based biologics | Haemophilia | Biotinylated albumin (WT, KAHQ or QMP) IgG1 and scFv-Alb | 10 to 30 µg for a 10 g mice | 20 µL à la dose de 1 mg/kg to 3.2 mg/kg | PBS | Nasal FcRn enabled efficient transcytosis of albumin fusion proteins. | Yes |
| Röhm [35] * 2017, Germany | In vitro | RPMI cells | Assess IgG quality after transcytosis | Enhancement of IgG permeation rate | mAb HIRMab 83-14 | 4 mg | 1.4 mg/mL for Fab; 4 mg/mL for IgG | L-arginine, HBC, PS20, sorbitol, trehalose | Aggregation of native IgG was reduced and transepithelial permeation rate was enhanced up to 2.8-fold with the used of specific formulations (F1) for intranasal aerosol-cell delivery. | No |
| Bequignon [9] * 2019, France | In vitro | HNEC | Assess mAb transcytosis via FcRn | Anti-cancer immunotherapy | Infliximab | 12.5 to 1250 ng | 50-500-50000 µg/mL | HBSS-MES | Transepithelial passage of therapeutic mAb was dose-dependent. | No |
| Ladel [16] * 2019, Germany | In vitro | OEPC and RPMI cells | Assess permeation rates of IgGs through the nasal mucosae | Anti-cancer immunotherapy | WT pIgG, WT hIgG and DG hIgG (biosimilar of Bevacizumab) | 50 µg | 1.5 mg/mL | PBS | hIgG permeation was faster than pIgGs over the first four in OEPC ALI cultures, but it converges from 8 h to 48 h. DG hIgG showed a higher permeation rate than WT hIgG in the RPMI ALI model. | No |
| Ex vivo | Porcine olfactory mucosa | hIgG permeation was 12 times higher after 5 h than the one of pIgG. The permeation rate of DG hIgG and WT hIgG did not show differences. | ||||||||
| Samson [46] * 2012, France | Ex vivo | Porcine nasal mucosa | Assess the transport of bevacizumab through porcine nasal mucosa | Rendu-Osler Disease | Bevacizumab | 500 µg | 25 mg/mL | trehalose, sodium phosphate, PS20, water | Total recovery of intranasally-delivered bevacizumab was 83% of the initial dose, with 53% localized at the mucosal surface and 11% that had gone through the mucosa. | No |
| Heidl [10] * 2015, Austria | Ex vivo | HNEC | Assess FcRn expression and localization in HNEC | Intranasal administration of mAb | None | NA | NA | NA | FcRn was detected in ciliated and basal cells of the nasal epithelium as well as in vascular endothelial cells and in gland tissue. | No |
| Ladel [15] * 2018, Germany | Ex vivo | Porcine olfactory mucosa | Assess ability of FcRn to transport IgGs through the nasal lamina propria | Anti-cancer immunotherapy | pIgG and hIgG (biosimilar of Bevacizumab) | 8 µg | 8 mg/mL | PBS | FcRn is expressed in the olfactory mucosa and enabled the apical uptake of allogeni, and xenogenic IgG in a species-specific manner. | No |
| Study | Selection Bias | Performance Bias | Attrition Bias | Detection Bias | Selective Reporting | Other Bias |
|---|---|---|---|---|---|---|
| Rawool [27], 2008 | -- | -- | + | + | + | + |
| Lu [28], 2011 | -- | -- | + | + | ++ | + |
| Ye [29], 2011 | -- | -- | ++ | ++ | ++ | + |
| Bitsatksis [69] 2015 | -- | -- | ++ | + | ++ | + |
| Kumar [61] 2018 | -- | -- | + | + | + | - |
| Bern [26] 2020 | -- | -- | ++ | + | ++ | + |
| Röhm [35] 2017 | -- | -- | ++ | + | ++ | + |
| Bequignon [9] 2019 | -- | -- | ++ | + | ++ | + |
| Samson [46] 2012 | -- | -- | ++ | ++ | ++ | + |
| Heidl [10] 2015 | -- | -- | + | - | + | + |
| Ladel [15] 2018 | -- | -- | + | + | ++ | + |
| Ladel [16] 2019 | -- | -- | + | + | ++ | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fieux, M.; Le Quellec, S.; Bartier, S.; Coste, A.; Louis, B.; Giroudon, C.; Nourredine, M.; Bequignon, E. FcRn as a Transporter for Nasal Delivery of Biologics: A Systematic Review. Int. J. Mol. Sci. 2021, 22, 6475. https://doi.org/10.3390/ijms22126475
Fieux M, Le Quellec S, Bartier S, Coste A, Louis B, Giroudon C, Nourredine M, Bequignon E. FcRn as a Transporter for Nasal Delivery of Biologics: A Systematic Review. International Journal of Molecular Sciences. 2021; 22(12):6475. https://doi.org/10.3390/ijms22126475
Chicago/Turabian StyleFieux, Maxime, Sandra Le Quellec, Sophie Bartier, André Coste, Bruno Louis, Caroline Giroudon, Mikail Nourredine, and Emilie Bequignon. 2021. "FcRn as a Transporter for Nasal Delivery of Biologics: A Systematic Review" International Journal of Molecular Sciences 22, no. 12: 6475. https://doi.org/10.3390/ijms22126475
APA StyleFieux, M., Le Quellec, S., Bartier, S., Coste, A., Louis, B., Giroudon, C., Nourredine, M., & Bequignon, E. (2021). FcRn as a Transporter for Nasal Delivery of Biologics: A Systematic Review. International Journal of Molecular Sciences, 22(12), 6475. https://doi.org/10.3390/ijms22126475