
THE MAIN CYTOTOXIC EFFECTS OF METHYLSELENINIC ACID ON VARIOUS CANCER CELLS


Abstract
:1. Introduction
2. Reasons and Molecular Mechanisms of the Cytotoxic Effect of MSA on Cancer Cells
3. The Role of MSA in the Regulation of ER-Stress
4. Differential Expression of ER-Resident Selenoprotein Genes under ER-Stress Conditions Caused by the MSA
5. Discussion
6. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hawkes, C.V.; Hartley, I.P.; Ineson, P.; Fitter, A.H. Soil temperature affects carbon allocation within arbuscular mycorrhizal networks and carbon transport from plant to fungus. Glob. Chang. Biol. 2007, 14, 1181–1190. [Google Scholar] [CrossRef]
- Fernandes, A.P.; Gandin, V. Selenium compounds as therapeutic agents in cancer. Biochim. Biophys. Acta 2015, 1850, 1642–1660. [Google Scholar] [CrossRef]
- Goltyaev, M.V.; Mal’tseva, V.N.; Varlamova, E.G. Expression of ER-resident selenoproteins and activation of cancer cells apoptosis mechanisms under ER-stress conditions caused by methylseleninic acid. Gene 2020, 755, 144884. [Google Scholar] [CrossRef]
- Goltyaev, M.V.; Varlamova, E.G.; Novoselov, S.V.; Fesenko, E.E. Activation of Signal Pathways of Apoptosis under Conditions of Prolonged ER-Stress Caused by Exposure of Mouse Testicular Teratoma Cells to Selenium-Containing Compounds. Dokl. Biochem. Biophys. 2020, 490, 9–11. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.-H.; Hsia, S.; Shih, M.-Y.; Hsieh, F.-C.; Chen, P.-C. Effects of Selenium Yeast on Oxidative Stress, Growth Inhibition, and Apoptosis in Human Breast Cancer Cells. Int. J. Med. Sci. 2015, 12, 748–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuznetsova, Y.P.; Goltyaev, M.V.; Gorbacheva, O.S.; Novoselov, S.V.; Varlamova, E.G.; Fesenko, E.E. Influence of Sodium Selenite on the mRNA Expression of the Mammalian Selenocysteine-Containing Protein Genes in Testicle and Prostate Cancer Cells. Dokl. Biochem. Biophys. 2018, 480, 131–134. [Google Scholar] [CrossRef]
- Misra, S.; Boylan, M.; Selvam, A.; Spallholz, J.E.; Björnstedt, M. Redox-active selenium compounds––From toxicity and cell death to cancer treatment. Nutrients 2015, 7, 3536–3556. [Google Scholar] [CrossRef] [Green Version]
- Shigemi, Z.; Manabe, K.; Hara, N.; Baba, Y.; Hosokawa, K.; Kagawa, H.; Watanabe, T.; Fujimuro, M. Methylseleninic acid and sodium selenite induce severe ER stress and subsequent apoptosis through UPR activation in PEL cells. Chem. Biol. Interact. 2017, 266, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Suzuki, N.; Ogra, Y. Bioavailability Comparison of Nine Bioselenocompounds In Vitro and In Vivo. Int. J. Mol. 2017, 18, 506. [Google Scholar] [CrossRef] [Green Version]
- Zu, K.; Bihani, T.; Lin, A.; Park, Y.M.; Mori, K.; Ip, C. Enhanced selenium effect on growth arrest by BiP/GRP78 knockdown in p53–null human prostate cancer cells. Oncogene 2006, 25, 546–554. [Google Scholar] [CrossRef] [Green Version]
- Zuazo, A.; Plano, D.; Ansó, E.; Lizarraga, E.; Font, M.; Irujo, J.J.M. Cytotoxic and proapototic activities of imidoselenocarbamate derivatives are dependent on the release of methylselenol. Chem. Res. Toxicol. 2012, 25, 2479–2489. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Durrani, F.A.; Rustum, Y.M. Selective modulation of the therapeutic efficacy of anticancer drugs by selenium containing compounds against human tumor xenografts. Clin. Cancer Res. 2004, 10, 2561–2569. [Google Scholar] [CrossRef] [Green Version]
- Foster, D.G. Selenophenol. Organic Syntheses, Coll. 1955, 3, 771. [Google Scholar] [CrossRef]
- Ip, C.; Dong, Y.; Ganther, H.E. New concepts in selenium chemoprevention. Cancer Metastasis Rev. 2002, 21, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Ip, C.; Zhu, Z.; Thompson, H.J.; Lisk, D.; Ganther, H.E. Chemoprevention of mammary cancer with Se-allylselenocysteine and other selenoamino acids in the rat. Anticancer Res. 1999, 19, 2875–2880. [Google Scholar] [PubMed]
- Weinshilboum, R.M.; Otterness, D.M.; Szumlanski, C.L. Methylation pharmacogenetics: Catechol O-methyltransferase, thiopurine methyltransferase, and histamine N-methyltransferase. Annu Rev. Pharm. Toxicol 1999, 39, 19–52. [Google Scholar] [CrossRef]
- Sunde, R. Molecular biology of selenoproteins. Ann Rev. Nutr. 1990, 10, 451–474. [Google Scholar] [CrossRef] [PubMed]
- Ip, C.; Thompson, H.J.; Zhu, Z.; Ganther, H.E. In vitro and in vivo studies of methylseleninic acid: Evidence that a monomethylated selenium metabolite is critical for cancer chemoprevention. Cancer Res. 2000, 60, 2882–2886. [Google Scholar]
- Dauplais, M.; Bierla, K.; Maizeray, C.; Lestini, R.; Lobinski, R.; Plateau, P.; Szpunar, J.; Lazard, M. Methylselenol Produced In Vivo from Methylseleninic Acid or Dimethyl Diselenide Induces Toxic Protein Aggregation in Saccharomyces cerevisiae. Int J. Mol. Sci. 2021, 22, 2241. [Google Scholar] [CrossRef]
- Dauplais, M.; Mahou, P.; Plateau, P.; Lazard, M. Exposure to the Methylselenol Precursor Dimethyldiselenide Induces a Reductive Endoplasmic Reticulum Stress in Saccharomyces cerevisiae. Int. J. Mol. Sci. 2021, 22, 5467. [Google Scholar] [CrossRef]
- Barrett, C.W.; Short, S.P.; Williams, C.S. Selenoproteins and oxidative stress-induced inflammatory tumorigenesis in the gut. Cell. Mol. Life Sci. 2017, 74, 607–616. [Google Scholar] [CrossRef] [Green Version]
- Peters, K.M.; Carlson, B.A.; Gladyshev, V.N.; Tsuji, P.A. Selenoproteins in colon cancer. Free Radic. Biol. Med. 2018, 127, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Shchedrina, V.A.; Everley, R.A.; Zhang, Y.; Gygi, S.P.; Hatfield, D.L.; Gladyshev, V.N. Selenoprotein K binds multiprotein complexes and is involved in the regulation of endoplasmic reticulum homeostasis. J. Biol. Chem. 2011, 286, 42937–42948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Short, S.P.; Williams, C.S. Selenoproteins in Tumorigenesis and Cancer Progression. Adv. Cancer Res. 2017, 136, 49–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varlamova, E.G. Protein–protein interactions of ER-resident selenoproteins with their physiological partners. Biochimie 2020, 171, 197–204. [Google Scholar] [CrossRef]
- Clark, L.C.; Combs, G.F., Jr.; Turnbull, B.W.; Slate, E.H.; Chalker, D.K.; Chow, J.; Davis, L.S.; Glover, R.A.; Graham, G.F.; Gross, E.G.; et al. Effects of selenium supplementation for patients with carcinoma of the skin. A randomized controlled trial. JAMA 1996, 276, 1957–1963. [Google Scholar] [CrossRef]
- Lipinski, B.; Pretorius, E. Iron-induced fibrin in cardiovascular disease. Curr. Neurovasc. Res. 2013, 10, 269–274. [Google Scholar] [CrossRef] [Green Version]
- Rayman, M.P. The argument for increasing selenium intake. Proc. Nutr. Soc. 2002, 61, 203–215. [Google Scholar] [CrossRef]
- Knock, F.E. Protein–Sulfhydryl Groups in Cancer and Gene Control. JAMA 1966, 197, 151. [Google Scholar] [CrossRef]
- Spallholz, J.E.; Shriver, B.J.; Reid, T.W. Dimethyldiselenide and methylseleninic acid generate superoxide in an in vitro chemiluminescence assay in the presence of glutathione: Implications for the anticarcinogenic activity of L-selenomethionine and L-Se–methylselenocysteine. Nutr. Cancer 2001, 40, 34–41. [Google Scholar] [CrossRef]
- Liu, M.; Hu, C.; Xu, Q.; Chen, L.; Ma, K.; Xu, N.; Zhu, H. Methylseleninic acid activates Keap1/Nrf2 pathway via up-regulating miR-200a in human oesophageal squamous cell carcinoma cells. Biosci. Rep. 2015, 35, e00256. [Google Scholar] [CrossRef]
- Okuno, T.; Honda, E.; Arakawa, T.; Ogino, H.; Ueno, H. Glutathione-dependent cell cycle G1 arrest and apoptosis induction in human lung cancer A549 cells caused by methylseleninic acid: Comparison with sodium selenite. Biol. Pharm. Bull. 2014, 37, 1831–1837. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.-M.; Ding, W.-X.; Ong, C.-N. Intracellular glutathione is a cofactor in methylseleninic acid–induced apoptotic cell death of human hepatoma HEPG2 cells. Free Radic. Biol. Med. 2002, 33, 552–561. [Google Scholar] [CrossRef]
- Lafin, J.T.; Sarsour, E.H.; Kalen, A.L.; Wagner, B.A.; Buettner, G.R.; Goswami, P.C. Methylseleninic Acid Induces Lipid Peroxidation and Radiation Sensitivity in Head and Neck Cancer Cells. Int. J. Mol. Sci. 2019, 20, 225. [Google Scholar] [CrossRef] [Green Version]
- Accioly, M.T.; Pacheco, P.; Maya-Monteiro, C.M.; Carrossini, N.; Robbs, B.K.; Oliveira, S.S.; Kaufmann, C.; Morgado-Diaz, J.A.; Bozza, P.T.; Viola, J.P. Lipid bodies are reservoirs of cyclooxygenase-2 and sites of prostaglandin-E2 synthesis in colon cancer cells. Cancer Res. 2008, 68, 1732–1740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ericsson, J.L.E.; Seljelid, R.; Orrenius, S. Comparative light and electron microscopic observations of the cytoplasmic matrix in renal carcinomas. Virchows Arch. Pathol. Anat. Physiol. Klin. Med. 1966, 341, 204–223. [Google Scholar] [CrossRef]
- Guillaumond, F.; Bidaut, G.; Ouaissi, M.; Servais, S.; Gouirand, V.; Olivares, O.; Lac, S.; Borge, L.; Roques, J.; Gayet, O.; et al. Cholesterol uptake disruption, in association with chemotherapy, is a promising combined metabolic therapy for pancreatic adenocarcinoma. Proc. Natl. Acad. Sci. USA 2015, 112, 2473–2478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toth, K.; Peter, I.; Kremmer, T.; Sugar, J. Lipid–rich cell thyroid adenoma: Histopathology with comparative lipid analysis. Virchows Arch. APathol. Anat. Histopathol. 1990, 417, 273–276. [Google Scholar] [CrossRef] [PubMed]
- Yue, S.; Li, J.; Lee, S.Y.; Lee, H.J.; Shao, T.; Song, B.; Cheng, L.; Masterson, T.A.; Liu, X.; Ratliff, T.L.; et al. Cholesteryl ester accumulation induced by PTEN loss and PI3K/AKT activation underlies human prostate cancer aggressiveness. Cell Metab. 2014, 19, 393–406. [Google Scholar] [CrossRef] [Green Version]
- Lobb, R.J.; Jacobson, G.M.; Cursons, R.T.; Jameson, M.B. The Interaction of Selenium with Chemotherapy and Radiation on Normal and Malignant Human Mononuclear Blood Cells. Int. J. Mol. Sci. 2018, 19, 3167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Z.; Dong, L.; Song, C.; Zhang, Y.; Zhu, C.; Zhang, Y.; Ling, Q.; Hoffmann, P.R.; Li, J.; Huang, Z.; et al. Methylseleninic Acid Provided at Nutritional Selenium Levels Inhibits Angiogenesis by Down-regulating Integrin β3 Signaling. Sci. Rep. 2017, 25, 9445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, S.D.; Hodivala-Dilke, K.M. The role of β3-integrins in tumor angiogenesis: Context is everything. Curr. Opin. Cell Biol. 2011, 23, 630–637. [Google Scholar] [CrossRef] [PubMed]
- Shattil, S.J.; Kim, C.; Ginsberg, M.H. The final steps of integrin activation: The end game. Nature reviews. Mol. Cell Biol. 2010, 11, 288–300. [Google Scholar] [CrossRef] [Green Version]
- Ahn, K.S.; Aggarwal, B.B. Transcription factor NF-kappa B: A sensor for smoke and stress signals. Ann. N. Y. Acad. Sci. 2005, 1056, 218–233. [Google Scholar] [CrossRef]
- Collins, T.; Read, M.A.; Neish, A.S.; Whitley, M.Z.; Thanos, D.; Maniatis, T. Transcriptional regulation of endothelial cell adhesion molecules: NF-kappa B and cytokine-inducible enhancers. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 1995, 9, 899–909. [Google Scholar] [CrossRef] [Green Version]
- Marcu, K.B.; Otero, M.; Olivotto, E.; Borzi, R.M.; Goldring, M.B. NF-κB signaling: Multiple angles to target OA. Curr. Drug Targets 2010, 11, 599–613. [Google Scholar] [CrossRef]
- Shih, R.H.; Wang, C.Y.; Yang, C.M. NF-κB Signaling Pathways in Neurological Inflammation: A Mini Review. Front. Mol. Neurosci. 2015, 8, 77. [Google Scholar] [CrossRef] [Green Version]
- Willett, M.; Pollard, H.J.; Vlasak, M.; Morley, S.J. Localization of ribosomes and translation initiation factors to talin/β3-integrin-enriched adhesion complexes in spreading and migrating mammalian cells. Biol. Cell 2010, 02, 265–276. [Google Scholar] [CrossRef]
- Zaidel–Bar, R.; Ballestrem, C.; Kam, Z.; Geiger, B. Early molecular events in the assembly of matrix adhesions at the leading edge of migrating cells. J. Cell Sci. 2003, 116, 4605–4613. [Google Scholar] [CrossRef] [Green Version]
- Altomare, D.A.; Testa, J.R. Perturbations of the AKT signaling pathway in human cancer. Oncogene 2005, 24, 7455–7464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eijkelenboom, A.; Burgering, B.M.T. FOXOs signalling integrators for homeostasis maintenance. Nat. Rev. Mol. Cell Biol. 2013, 14, 83–97. [Google Scholar] [CrossRef] [PubMed]
- Greer, E.L.; Brunet, A. FOXO transcription factors at the interface between longevity and tumor suppression. Oncogene 2005, 24, 7410–7425. [Google Scholar] [CrossRef] [Green Version]
- Lam, E.W.; Brosens, J.J.; Gomes, A.R.; Koo, C.Y. Forkhead box proteins: Tuning forks for transcriptional harmony. Nat. Rev. Cancer 2013, 13, 482–495. [Google Scholar] [CrossRef]
- Tarrado-Castellarnau, M.; Cortés, R.; Zanuy, M.; Tarragó-Celada, J.; Polat, I.H.; Hill, R.; Fan, T.W.; Link, W.; Cascante, M. Methylseleninic acid promotes antitumour effects via nuclear FOXO3a translocation through Akt inhibition. Pharm. Res. 2015, 102, 218–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, H.; Wu, M. The Inhibitory Efficacy of Methylseleninic Acid Against Colon Cancer Xenografts in C57BL/6 Mice. Nutr. Cancer 2015, 67, 831–838. [Google Scholar] [CrossRef]
- Li, Z.; Carrier, L.; Rowan, B.G. Methylseleninic acid synergizes with tamoxifen to induce caspase-mediated apoptosis in breast cancer cells. Mol. Cancer Ther. 2008, 7, 3056–3063. [Google Scholar] [CrossRef] [Green Version]
- Muecke, R.; Schomburg, L.; Glatzel, M.; Berndt-Skorka, R.; Baaske, D.; Reichl, B.; Buentzel, J.; Kundt, G.; Prott, F.J.; Devries, A.; et al. Multicenter, phase 3 trial comparing selenium supplementation with observation in gynecologic radiation oncology. Int. J. Radiat. Oncol. Biol. Phys. 2010, 78, 828–835. [Google Scholar] [CrossRef] [PubMed]
- Sieja, K.; Talerczyk, M. Selenium as an element in the treatment of ovarian cancer in women receiving chemotherapy. Gynecol. Oncol. 2004, 93, 320–327. [Google Scholar] [CrossRef]
- Qiu, C.; Zhang, T.; Zhu, X.; Qiu, J.; Jiang, K.; Zhao, G.; Wu, H.; Deng, G. Methylseleninic Acid Suppresses Breast Cancer Growth via the JAK2/STAT3 Pathway. Reprod. Sci. 2019, 26, 829–838. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Guo, M.; Liu, Y.; Mu, W.; Deng, G.; Li, C.; Changwei, Q. Selenium induces an anti-tumor effect via inhibiting intra tumoral angiogenesis in a mouse model of transplanted canine mammary tumor cells. Biol. Trace Element Res. 2016, 171, 371–379. [Google Scholar] [CrossRef]
- Baird, L.; Yamamoto, M. The Molecular Mechanisms Regulating the KEAP1-NRF2 Pathway. Mol. Cell Biol. 2020, 40, e00099-20. [Google Scholar] [CrossRef] [PubMed]
- Lennicke, C.; Rahn, J.; Bukur, J.; Hochgräfe, F.; Wessjohann, L.A.; Lichtenfels, R.; Seliger, B. Modulation of MHC class I surface expression in B16F10 melanoma cells by methylseleninic acid. Oncoimmunology 2017, 6, e1259049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Overholtzer, M.; Mailleux, A.A.; Mouneimne, G.; Normand, G.; Schnitt, S.J.; King, R.W.; Cibas, E.S.; Brugge, J.S. A Nonapoptotic Cell Death Process, Entosis, that Occurs by Cell-in-Cell Invasion. Cell 2007, 131, 966–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalkar, P.; Díaz-Argelich, N.; Palop, J.A.; Sanmartín, C.; Fernandes, A.P. Novel Methylselenoesters Induce Programed Cell Death via Entosis in Pancreatic Cancer Cells. Int. J. Mol. Sci. 2018, 19, 2849. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Sun, Y.; Yang, S.; Liang, X. Inhibitory Effect of Selenium on Esophagus Cancer Cells and the Related Mechanism. J. Nutr. Sci. Vitam. 2020, 66, 456–461. [Google Scholar] [CrossRef]
- Marino, M.; Stoilova, T.; Giorgi, C.; Bachi, A.; Cattaneo, A.; Auricchio, A.; Pinton, P.; Zito, E. SEPN1, an endoplasmic reticulum-localized selenoprotein linked to skeletal muscle pathology, counteracts hyperoxidation by means of redox-regulating SERCA2 pump activity, Hum. Mol. Genet. 2015, 24, 1843–1855. [Google Scholar] [CrossRef] [Green Version]
- Davis, C.D.; Uthus, E.O.; Finley, J.W. Dietary selenium and arsenic affect DNA methylation in vitro in Caco-2 cells and in vivo in rat liver and colon. J. Nutr. 2000, 130, 2903–2909. [Google Scholar] [CrossRef]
- Kassam, S.; Goenaga–Infante, H.; Maharaj, L.; Hiley, C.T.; Juliger, S.; Joel, S.P. Methylseleninic acid inhibits HDAC activity in diffuse large B-cell lymphoma cell lines. Cancer Chemother. Pharmacol. 2011, 68, 815–821. [Google Scholar] [CrossRef] [Green Version]
- Yin, M.B.; Li, Z.R.; Tóth, K.; Cao, S.; Durrani, F.A.; Hapke, G.; Bhattacharya, A.; Azrak, R.G.; Frank, C.; Rustum, Y.M. Potentiation of irinotecan sensitivity by Se-methylselenocysteine in an in vivo tumor model is associated with downregulation of cyclooxygenase-2, inducible nitric oxide synthase, and hypoxia-inducible factor 1alpha expression, resulting in reduced angiogenesis. Oncogene 2006, 25, 2509–2519. [Google Scholar]
- Liu, L.; Ning, X.; Sun, L.; Zhang, H.; Shi, Y.; Guo, C.; Han, S.; Liu, J.; Sun, S.; Han, Z.; et al. Hypoxia-inducible factor-1 alpha contributes to hypoxia-induced chemoresistance in gastric cancer. Cancer Sci. 2008, 99, 121–128. [Google Scholar] [PubMed]
- Unruh, A.; Ressel, A.; Mohamed, H.G.; Johnson, R.S.; Nadrowitz, R.; Richter, E.; Katschinski, D.M.; Wenger, R.H. The hypoxia-inducible factor-1 alpha is a negative factor for tumor therapy. Oncogene 2003, 22, 3213–3220. [Google Scholar] [CrossRef] [Green Version]
- Callapina, M.; Zhou, J.; Schmid, T.; Köhl, R.; Brüne, B. NO restores HIF-1alpha hydroxylation during hypoxia: Role of reactive oxygen species. Free Radic. Biol. Med. 2005, 39, 925–936. [Google Scholar] [CrossRef] [PubMed]
- Chintala, S.; Tóth, K.; Cao, S.; Durrani, F.A. Se-methylselenocysteine sensitizes hypoxic tumor cells to irinotecan by targeting hypoxia-inducible factor 1α. Cancer Chemother. Pharmacol. 2010, 66, 899–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finley, L.W.; Carracedo, A.; Lee, J.; Souza, A.; Egia, A.; Zhang, J.; Teruya-Feldstein, J.; Moreira, P.I.; Cardoso, S.M.; Clish, C.B.; et al. SIRT3 opposes reprogramming of cancer cell metabolism through HIF1α destabilization. Cancer Cell. 2011, 19, 416–428. [Google Scholar] [CrossRef] [Green Version]
- Isaacs, J.S.; Jung, Y.J.; Mimnaugh, E.G.; Martinez, A.; Cuttitta, F.; Neckers, L.M. Hsp90 regulates a von Hippel Lindau-independent hypoxia-inducible factor-1 alpha–degradative pathway. J. Biol. Chem. 2002, 277, 29936–29944. [Google Scholar] [CrossRef] [Green Version]
- Koh, M.Y.; Darnay, B.G.; Powis, G. Hypoxia-associated factor, a novel E3-ubiquitin ligase, binds and ubiquitinates hypoxia-inducible factor 1α, leading to its oxygen-independent degradation. Mol. Cell Biol. 2008, 28, 7081–7095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.D.; Kim, Y.P.; Mohammed, K.A.; Jones, D.K.; Muhammad, I.; Dunbar, D.C.; Nagle, D.G. Terpenoid tetrahydroisoquinoline alkaloids emetine, klugine, and isocephaeline inhibit the activation of hypoxia-inducible factor-1 in breast tumor cells. J. Nat. Prod. 2005, 68, 947–950. [Google Scholar] [CrossRef] [Green Version]
- Chintala, S.; Najrana, T.; Toth, K.; Cao, S.; Durrani, F.A.; Pili, R.; Rustum, Y.M. Prolyl hydroxylase 2 dependent and Von-Hippel-Lindau independent degradation of Hypoxia-inducible factor 1 and 2 alpha by selenium in clear cell renal cell carcinoma leads to tumor growth inhibition. BMC Cancer 2012, 12, 293. [Google Scholar] [CrossRef] [Green Version]
- Sinha, I.; Null, K.; Wolter, W.; Suckow, M.A.; King, T.; Pinto, L.T.; Sinha, R. Methylseleninic acid downregulates hypoxia-inducible factor-1α in invasive prostate cancer. Int. J. Cancer 2012, 130, 1430–1439. [Google Scholar] [CrossRef]
- Dery, M.A.; Michaud, M.D.; Richard, D.E. Hypoxia-inducible factor 1: Regulation by hypoxic and non-hypoxic activators. Int. J. Biochem. Cell Biol. 2005, 37, 535–540. [Google Scholar] [CrossRef]
- Kaczmarek, M.; Cachau, R.E.; Topol, I.A.; Kasprzak, K.S.; Ghio, A.; Salnikow, K. Metal ions-stimulated iron oxidation in hydroxylases facilitates stabilization of HIF-1 alpha protein. Toxicol Sci. 2009, 107, 394–403. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Hu, H.; Wang, Z.; Xiong, H.; Cheng, Y.; Liao, J.D.; Deng, Y.; Lü, J. Methylseleninic acid suppresses pancreatic cancer growth involving multiple pathways. Nutr. Cancer. 2014, 66, 295–307. [Google Scholar] [CrossRef]
- Qi, Y.; Fu, X.; Xiong, Z.; Zhang, H.; Hill, S.M.; Rowan, B.G.; Dong, Y. Methylseleninic Acid Enhances Paclitaxel Efficacy for the Treatment of Triple-Negative Breast Cancer. PLoS ONE 2012, 7, e31539. [Google Scholar] [CrossRef] [Green Version]
- Arne’r, E.S.; Holmgren, A. Physiological functions of Trx and Trx reductase. Eur. J. Biochem. 2000, 267, 6102–6109. [Google Scholar] [CrossRef]
- Dikiy, S.A.; Novoselov, V.; Fomenko, D.E.; Sengupta, A.; Carlson, B.A.; Cerny, R.L.; Ginalski, K.; Grishin, N.V.; Hatfield, D.L.; Gladyshev, V.N. SelT, SelW, SelH, and Rdx12: Genomics and molecular insights into the functions of selenoproteins of a novel Trx–like family. J. Biochem. 2007, 46, 6871–6882. [Google Scholar] [CrossRef]
- Ferguson, A.D.; Labunskyy, V.M.; Fomenko, D.E.; Araç, D.; Chelliah, Y.; Amezcua, C.A.; Rizo, J.; Gladyshev, V.N.; Deisenhofer, J. NMR structures of the selenoproteins Sep15 and SelM reveal redox activity of a new Trx–like family. J. Biol. Chem. 2006, 281, 3536–3543. [Google Scholar] [CrossRef] [Green Version]
- Hamieh, A.; Cartier, D.; Abid, H.; Calas, A.; Burel, C.; Bucharles, C.; Jehan, C.; Grumolato, L.; Landry, M.; Lerouge, P.; et al. Selenoprotein T is a novel OST subunit that regulates UPR signaling and hormone secretion. EMBO 2017, 18, 1935–1946. [Google Scholar] [CrossRef]
- Qi, Y.; Grishin, N.V. Structural classification of Trx–like fold proteins. J. Proteins 2005, 58, 376–388. [Google Scholar] [CrossRef] [Green Version]
- Petit, N.; Lescure, A.; Rederstorff, M.; Krol, A.; Moghadaszadeh, B.; Wewer, U.M.; Guicheney, P. Selenoprotein N: An endoplasmic reticulum glycoprotein with an early developmental expression pattern. Hum. Mol. Genet. 2003, 12, 1045–1053. [Google Scholar] [CrossRef] [Green Version]
- Gereben, B.; Zavacki, A.M.; Ribich, S.; Kim, B.W.; Huang, S.A.; Simonides, W.S.; Zeöld, A.; Bianco, A.C. Cellular and Molecular Basis of Deiodinase-Regulated Thyroid Hormone Signaling. Endocr. Rev. 2008, 29, 898–938. [Google Scholar] [CrossRef] [Green Version]
- Sagar, G.D.; Gereben, B.; Callebaut, I.; Mornon, J.P.; Zeöld, A.; da Silva, W.S.; Luongo, C.; Dentice, M.; Tente, S.M.; Freitas, B.C.; et al. Ubiquitination-induced conformational change within the deiodinase dimer is a switch regulating enzyme activity. Mol. Cell. Biol. 2007, 27, 4774–4783. [Google Scholar] [CrossRef] [Green Version]
- Varlamova, E.G.; Goltyaev, M.V.; Fesenko, E.E. Protein-partners of selenoprotein SELM and the role of selenium in regulation of its expression in human cancer cells. Dokl. Biochem. Biophys. 2019, 488, 94–98. [Google Scholar] [CrossRef]
- Davis, T.Z.; Tiwary, A.K.; Stegelmeier, B.L.; Pfister, J.A.; Panter, K.E.; Hall, J.O. Comparative oral dose toxico kinetics of sodium selenite and selenomethionine. J. Appl. Toxicol. 2017, 37, 231–238. [Google Scholar] [CrossRef]
- Han, H.W.; Yang, E.J.; Lee, S.-M. Sodium Selenite Alleviates Breast Cancer-Related Lymphedema Independent of Antioxidant Defense System. Nutrients 2019, 11, 1021. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.B.; Yang, E.Y.; Woo, J.; Kwon, H.; Lim, W.; Moon, B.-I. Sodium Selenite Enhanced the Anti-proliferative Effect of MEK-ERK Inhibitor in Thyroid Cancer Cells. In Vivo 2020, 34, 185–190. [Google Scholar] [CrossRef]
- Bang, J.; Jang, M.; Huh, J.H.; Na, J.W.; Shim, M.; Carlson, B.A.; Tobe, R.; Tsuji, P.A.; Gladyshev, V.N.; Hatfield, D.L.; et al. Deficiency of the 15-kDa selenoprotein led to cytoskeleton remodeling and non-apoptotic membrane blebbing through a RhoA/ROCK pathway. Biochem. Biophys. Res. Commun. 2015, 456, 884–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.L.; Shim, M.S.; Chung, J.; Yoo, H.S.; Ha, J.M.; Kim, J.Y.; Choi, J.; Zang, S.L.; Hou, X.; Carlson, B.A.; et al. G-rich, a Drosophila selenoprotein, is a Golgi-resident type III membrane protein. Biochem. Biophys. Res. Commun. 2006, 348, 1296–1301. [Google Scholar] [CrossRef]
- Shchedrina, V.A.; Zhang, Y.; Labunskyy, V.M.; Hatfield, D.L.; Gladyshev, V.N. Structure-Function Relations, Physiological Roles, and Evolution of Mammalian ER-Resident Selenoproteins. Antioxid. Redox Signal. 2010, 12, 839–849. [Google Scholar] [CrossRef]
- Lilley, B.N.; Ploegh, H.L. Multiprotein complexes that link dislocation, ubiquitination, and extraction of misfolded proteins from the endoplasmic reticulum membrane. Proc. Natl. Acad. Sci. USA 2005, 102, 14296–14301. [Google Scholar] [CrossRef] [Green Version]
- Oda, Y.; Okada, T.; Yoshida, H.; Kaufman, R.J.; Nagata, K.; Mori, K. Derlin-2 and Derlin-3 are regulated by the mammalian unfolded protein response and are required for ER-associated degradation. J. Cell Biol. 2006, 172, 383–393. [Google Scholar] [CrossRef] [Green Version]
- Ballar, P.; Shen, Y.; Yang, H.; Fang, S. The role of a novel p97/valosin-containing protein-interacting motif of gp78 in endoplasmic reticulum-associated degradation. J. Biol. Chem. 2006, 281, 35359–35368. [Google Scholar] [CrossRef] [Green Version]
- Ballar, P.; Pabuccuoglu, A.; Kose, F.A. Different p97/VCP complexes function in retrotranslocation step of mammalian ER-associated degradation (ERAD). Int. J. Biochem. Cell Biol. 2011, 43, 613–621. [Google Scholar] [CrossRef]
- Doroudgar, S.; Vlkers, M.; Thuerauf, D.J.; Khan, M.; Mohsin, S.; Respress, J.L.; Wang, W.; Gude, N.; Meller, O.J.; Wehrens, X.H.; et al. Hrd1 and ER-associated protein degradation, ERAD, are critical elements of the adaptive ER stress response in cardiac myocytes. Circ. Res. 2015, 117, 536–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Kwon, J.H.; Jeon, Y.H.; Ko, K.Y.; Lee, S.-R.; Kim, I.Y. Pro-178 and Pro-183 of selenoprotein S Are essential residues for interaction with p97 (VCP) during endoplasmic reticulum-associated degradation. J. Biol. Chem. 2014, 289, 13758–13768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Park, K.J.; Jang, J.K.; Jeon, Y.H.; Ko, K.Y.; Kwon, J.H.; Lee, S.R.; Kim, I.Y. Selenoprotein S-dependent Selenoprotein K Binding to p97(VCP) Protein Is Essential for Endoplasmic Reticulum-Associated Degradation. J. Biol. Chem. 2015, 290, 29941–29952. [Google Scholar] [CrossRef] [Green Version]
- Chernorudskiy, A.; Varone, E.; Colombo, S.F.; Fumagalli, S.; Cagnotto, A.; Cattaneo, A.; Briens, M.; Baltzinger, M.; Kuhn, L.; Bachi, A.; et al. Selenoprotein N is an endoplasmic reticulum calcium sensor that links luminal calcium levels to a redox activity. Biochemistry 2020, 117, 21288–21298. [Google Scholar] [CrossRef]
- Appenzeller-Herzog, C.; Simmen, T. ER-luminal thiol/selenol-mediated regulation of Ca2+ signaling. Biochem. Soc. Trans. 2016, 44, 452–459. [Google Scholar] [CrossRef]
- Arbogast, S.; Beuvin, M.; Fraysse, B.; Zhou, H.; Muntoni, F.; Ferreiro, A. Oxidative stress in SEPN1-related myopathy: From pathophysiology to treatment. Ann. Neurol. 2009, 65, 677–686. [Google Scholar] [CrossRef]
- Baqui, M.M.; Gereben, B.; Harney, J.W.; Larsen, P.R.; Bianco, A.C. Distinct subcellular localization of transiently expressed types 1 and 2 iodothyronine deiodinases as determined by immunofluorescence confocal microscopy. Endocrinology 2000, 141, 4309–4312. [Google Scholar] [CrossRef]
- Zeöld, A.; Pormüller, L.; Dentice, M.; Harney, J.W.; Curcio-Morelli, C.; Tente, S.M.; Bianco, A.C.; Gereben, B. Metabolic instability of type 2 deiodinase is transferable to stable proteins independently of subcellular localization. J. Biol. Chem. 2006, 281, 31538–31543. [Google Scholar] [CrossRef]
- Arrojo, E.D.R.; Egri, P.; Jo, S.; Gereben, B.; Bianco, A.C. The type II deiodinase is retrotranslocated to the cytoplasm and proteasomes via p97/Atx3 complex. Mol. Endocrinol. 2013, 27, 2105–2115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dentice, M.; Bandyopadhyay, A.; Gereben, B.; Callebaut, I.; Christoffolete, M.A.; Kim, B.W.; Nissim, S.; Mornon, J.P.; Zavacki, A.M.; Zeold, A.; et al. The Hedgehog-inducible ubiquitin ligase subunit WSB-1 modulates thyroid hormone activation and PTHrP secretion in the developing growth plate. Nat. Cell Biol. 2005, 7, 698–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zavacki, A.M.; Arrojo, E.D.R.; Freitas, B.C.; Chung, M.; Harney, J.W.; Egri, P.; Wittmann, G.; Fekete, C.; Gereben, B.; Bianco, A.C. The E3 ubiquitin ligase TEB4 mediates degradation of type 2 iodothyronine deiodinase. Mol. Cell. Biol. 2009, 29, 5339–5347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Miranda, J.X.; de Oliveira Andrade, F.; de Conti, A.; Dagli, M.L.Z.; Moreno, F.S.; Ong, T.P. Effects of selenium compounds on proliferation and epigenetic marks of breast cancer cells. J. Trace Elem. Med. Biol. 2014, 28, 486–491. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
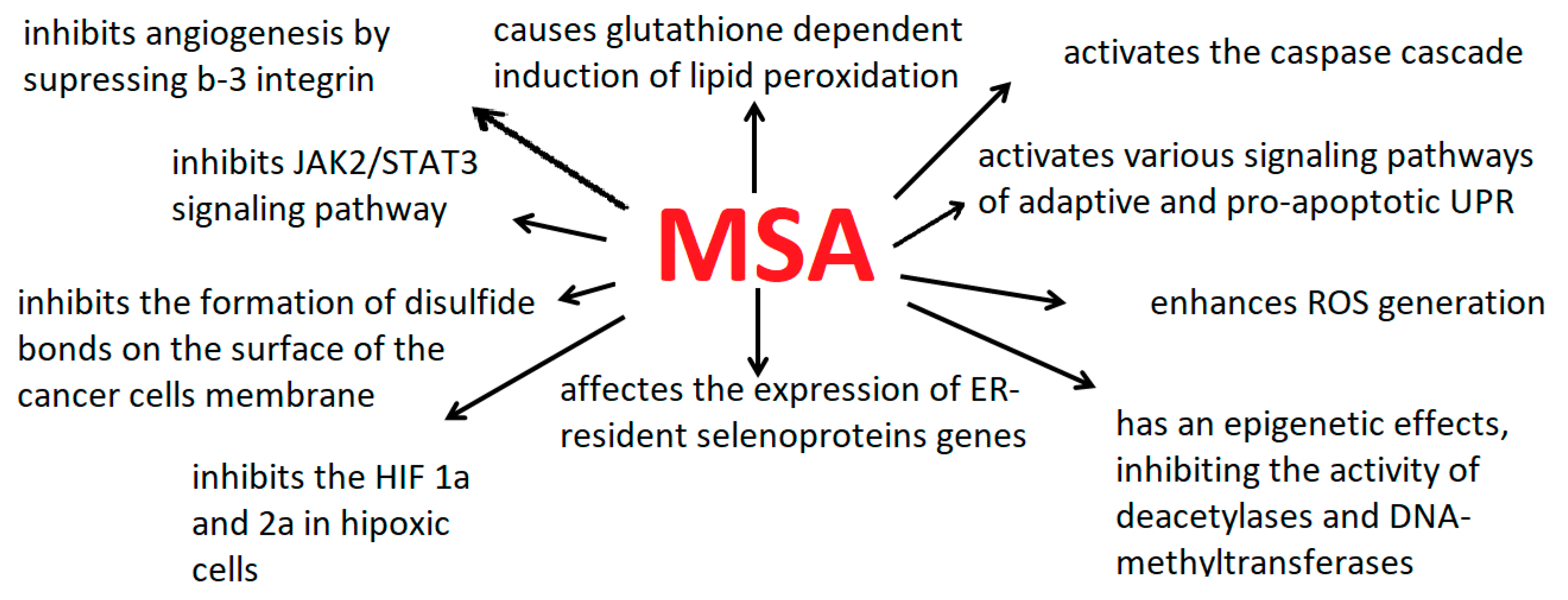{kind=link}
| Cancer Cell Line Name | Molecular Mechanisms of Regulations of MSA Cytotoxicity | Reference |
|---|---|---|
| A-172 (human glioblastoma) | Treatment with 1 μM MSA during 24 h reduces proliferation by 70–80% and increases the expression of mRNA of the transcription factors ATF-4 and ATF-6. Silencing of SELENOT under ER-stress induced by 0.1 μM MSA resulted in an increase in the expression of SELENOM and decreases the expression of AMFR (autocrine motility factor receptor) and RNF5 (ring finger protein 5), which are E3-ubiquitin ligases, important enzymes of the ERAD-system. | [3] |
| A549 (human lung cancer cells) | MSA reduced cell growth by 50% when cells were treated with 2.2 ± 0.3, 1.6 ± 0.2, and 1.3 ± 0.1 μM for 24, 48, and 72 h, respectively. | [32,54] |
| Micromolar concentrations of MSA markedly inhibited the growth of A549 cells. MSA induces G1 arrest by down-regulating cyclin E1 and up-regulating p27Kip1. MSA was simultaneously shown to enhance apoptosis induction in the presence of intracellular GSH. Over 67% of cells were consecutively inhibited at the G0/G1 phase. | ||
| 5 μM MSA attenuates the activity of glycolysis, TCA cycle, PPP, and/or nucleotide biosynthesis. MSA effects are associated with the inhibition of the Akt pathway, leading to dephosphorylation of FOXO proteins and their nuclear translocation, which in turn activate the expression of FOXO target genes. FOXO dephosphorylation and relocalization to the nucleus are early events that activate the antiproliferative response of A549 cells to MSA | ||
| Cal27 (tongue origin, CRL-2095), SCC25 (tongue origin, CRL-1628) (head and neck squamous carcinoma cells) | Treatment with 10 μM MSA during 24 h appears to be more toxic to SCC25 compared to Cal27 cells. MSA inhibits cell proliferation by more than 90%. | [34] |
| Lipid peroxidation (LPO) is an essential step in the MSA-induced toxicity of HNSCC cells. MSA sensitizes HNSCC cells to radiation and exhibits toxicity through a GSH-dependent induction of LPO. Cal27 cells treated with 10 µM MSA for 72 h were found to have 1.16 fmol lipid hydroperoxide per cell, nearly 40 times as much as untreated cells | ||
| DU145 (human prostate carcinoma epithelial cells) | At 1 μM MSA, the viability of these cell lines decreases by 70–80%. MSA promoted the activation of the PERK signaling pathway, increases expression of apoptosis genes, as well as effector caspase-3 and inflammatory caspase-4. When DU 145 cells were treated with 1 μM MSA for 24 h, a significant increase in the expression of the SELENOF and SELENOM genes was observed. | [3] |
| Eca109 (human esophageal carcinoma cell line) | Treatment of cells with 20 and 40 μM MSA for 72 h reduced cell viability by only approximately 50%. | [65] |
| After treatment of Eca109 cells with 20 μM MSA for 48 h cell cycle was arrested in G0/G1 phase, and the cell population was increased in the S phase. MSA significantly reduces the expression of FAL1 (focally amplified lncRNA on chromosome 1), and thus, the level of PTEN (phosphatase and tensin homolog deleted on chromosome 10) is increased. | ||
| HUVEC (human umbilical vein endothelial cells) | A total of 10 μM MSA inhibits cell proliferation by 20% during 24 h. | [41] |
| Effectively increases the adherence to collagen I and inhibits cell migration of HUVECs; down-regulates Integrin β3 and inhibits phosphorylation of AKT; disrupts the clustering of integrin β3 surface localization; inhibits VEGF-induced angiogenesis and the phosphorylation of IκBα and NF-κB, and the nuclear translocation of NF-κB | ||
| KYSE150, KYSE180, KYSE410, and KYSE510 (ESCC-human esophageal squamous cell carcinoma cells): | MSA treatment significantly down-regulated Keap1 (Kelch-like ECH-associated protein 1), induced nuclear accumulation of Nrf2 (nuclear factor E2-related factor 2), and enhance the ARE (antioxidant response element) promoter activity and significantly induce miR-200a expression. | [31] |
| MDA-MB-231(human breast adenocarcinoma cells) | The viability of cells treated with 4 μM MSA for 72 h decreased by more than 40%, while simultaneously treating cells with 4 μM MSA and 10 nM paclitaxel for 72 h-by more than 80%. | [83] |
| MSA synergistically enhances the growth-inhibitory efficacy of paclitaxel in MDA-MB-231 cells. MSA enhances paclitaxel-induced apoptosis. MSA could enhance paclitaxel-mediated G2/M arrest suggests the potential of using MSA to overcome paclitaxel resistance | ||
| PANC-1, PANC-28, Colo357, Bxpc-3, HPAC (human pancreatic cancer cell) lines | PANC-1 treatment with 2.6 μM MSA for 5 d, PANC-28 treatment with 1.2 μM MSA for 3 d, Colo357 treatment with 0.6 μM MSA for 48 h, Bxpc-3 treatment with 1.15 μM MSA for 48 h, HPAC treatment with 3.7 μM MSA for 48 h resulted in a 50% decrease in cells growth. | [64,82] |
| MSA induced G1 arrest and caspase-mediated apoptosis in most pancreatic cancer cell lines and manifested a rapid G2 arrest in the PANC-1 and PANC-28 cell lines. MSA induced G1 arrest in Colo357, Bxpc-3, HPAC cells at 12, 24, and 48 h. A total of 7.5 μM MSA induced a modest 2.2-fold of apoptotic fragmentation in PANC-1 cells compared to control. | ||
| When PANC-1 cells were treated with 1 μM MSA for 72 h, a decrease in cell viability was observed only by 20%, while when treated with 50 μM MSA, by more than 90%. | ||
| MSA induced entosis by cell detachment through downregulation of cell division control protein 42 homologs (CDC42) and its downstream effector β1-integrin (CD29). Treatment with MSA led to a unique phenotype, characterized by changes in morphology and cell detachment from the culture plate prior to cell death. | ||
| PEL (primary effusion lymphoma) | Treatment with 30 μM MSA during 24 h reduces proliferation by 70–80%. | [8] |
| MSA induces pro-apoptotic UPR through transcriptional activation of pro-apoptotic genes, CHOP, Bim, and Puma, via the activation of caspases, induces oxidative stress but not lytic replication | ||
| 4T1 (mouse malignant breast cancer cells) | MSA significantly induces apoptosis of these cancer cells by activating Bax, caspase-3, PARP. In addition, MSA is able to inhibit tumor angiogenesis by reducing the expression of vascular endothelial growth factors VEGF and Ang-2 in mammary cells of dogs and mouse models. In addition, this series of experiments showed that MSA inhibited the JAK2/STAT3 signaling pathway. | [59,60] |
| WM1552c, UKRV, Colo875 (human melanoma cell lines), SK-BR-3, BT-474 (human mammary carcinoma cell lines), B16F10 (mouse skin melanoma cells) | Treatment with MSA increased the MHC class I (major histocompatibility complex surface) expression levels in all the tested tumor cell lines. MSA partially mimics IFNγ signaling, such as the upregulation of STAT1(Stat1 signal transducer and activator of transcription 1), JAK1 (janus kinase 1), IRF1, IRF5, IRF7, and IRF9 (interferon-regulated factor1, 5, 7 and 9) on the mRNA and/or protein expression levels. In addition, MSA treatment leads to activation of the transcription factor Nrf2. | [62] |
| HNSCC (human head and neck squamous cell carcinoma), | MSA effectively inhibits the HIF-1α in hypoxic cells, while PHD 2 and PHD 3, on the contrary, were activated, which was demonstrated in HNSCC cells. | [73] |
| RC2 and 786-0 (clear cell renal cell carcinoma) | The resistance of these cancer cells, overexpressing the HIF-1α under hypoxia, to SN38-the active metabolite of irinotecan, was reduced after MSA treatment. Similar synergistic activity of MSA in combination with docetaxel was shown in a model of prostate cancer cells, which can also be explained by the inhibition of HIF-1α by the activation of PHDs. | [74,75,76,77] |
| (PC-3 and PC-3M, PAIII and DU145 (human prostate cancer cells) | Treatment of clear cell renal cell carcinoma (RC2 and 786-0) with a pharmacological dose of MSA (10 μM) promoted inhibition of constitutively expressed transcription factors HIF-1α and HIF-2α in RC2 and 786-0 cells, respectively. | [78] |
| MSA inhibits the expression and activity of HIF-1α in invasive rat and human prostate cancer cells [79]. Thus, the treatment of highly aggressive human prostate cancer cell lines (PC-3 and PC-3M) with MSA led to significant inhibition of growth and induction of apoptosis, and MSA has a stronger effect on cells under hypoxia than under normoxia, especially at a physiological dose of MSA (5 μM). | [79] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Varlamova, E.G.; Turovsky, E.A. THE MAIN CYTOTOXIC EFFECTS OF METHYLSELENINIC ACID ON VARIOUS CANCER CELLS. Int. J. Mol. Sci. 2021, 22, 6614. https://doi.org/10.3390/ijms22126614
Varlamova EG, Turovsky EA. THE MAIN CYTOTOXIC EFFECTS OF METHYLSELENINIC ACID ON VARIOUS CANCER CELLS. International Journal of Molecular Sciences. 2021; 22(12):6614. https://doi.org/10.3390/ijms22126614
Chicago/Turabian StyleVarlamova, Elena G., and Egor A. Turovsky. 2021. "THE MAIN CYTOTOXIC EFFECTS OF METHYLSELENINIC ACID ON VARIOUS CANCER CELLS" International Journal of Molecular Sciences 22, no. 12: 6614. https://doi.org/10.3390/ijms22126614
APA StyleVarlamova, E. G., & Turovsky, E. A. (2021). THE MAIN CYTOTOXIC EFFECTS OF METHYLSELENINIC ACID ON VARIOUS CANCER CELLS. International Journal of Molecular Sciences, 22(12), 6614. https://doi.org/10.3390/ijms22126614