
Placental Glucose Transporters and Response to Bisphenol A in Pregnancies from of Normal and Overweight Mothers

 ,
,  , , and
, , and 
Abstract
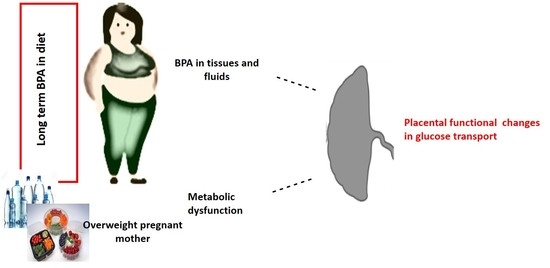
:
1. Introduction
2. Results
2.1. Clinical Features of Study Population
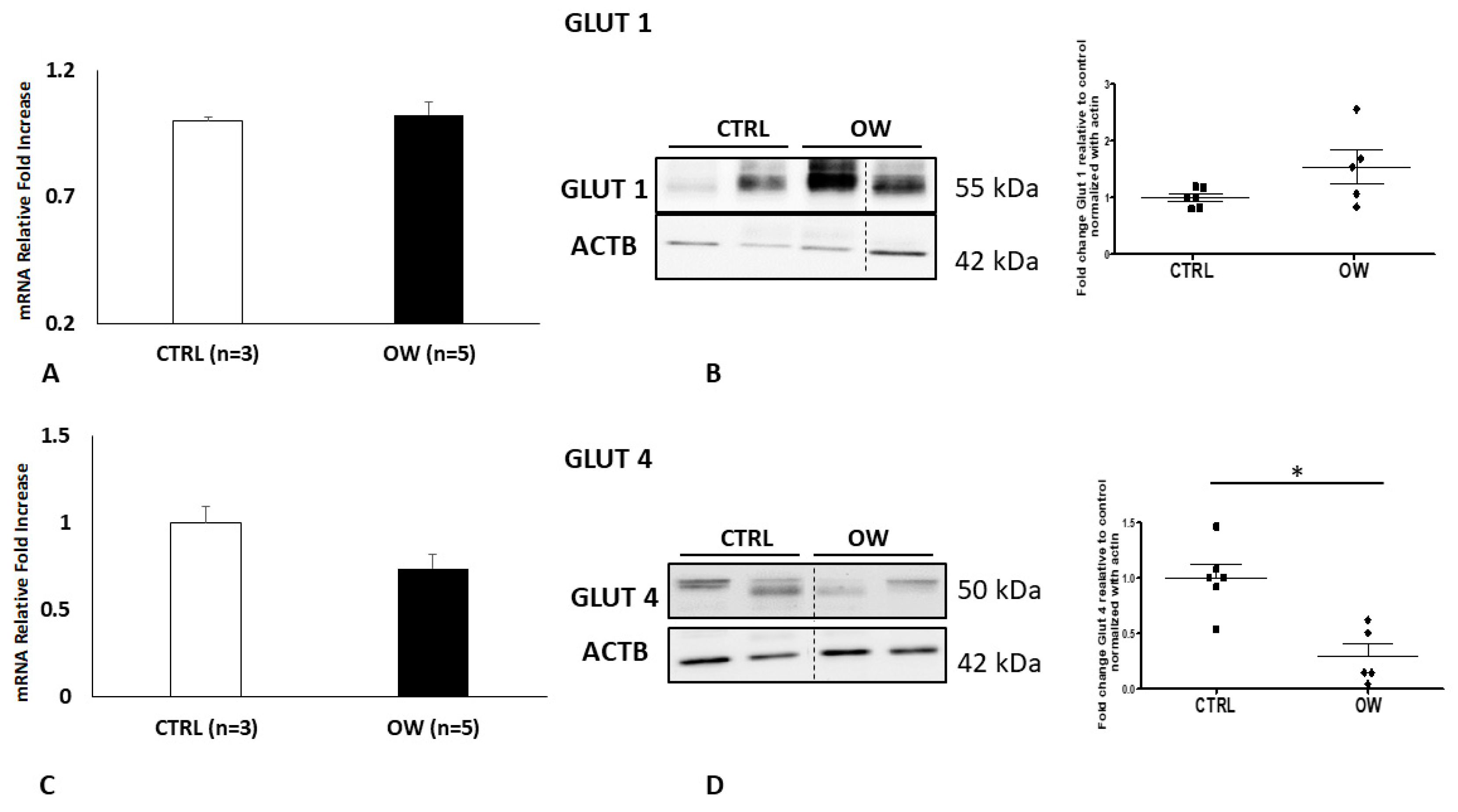
2.2. Placental GLUT1 and GLUT4 Expression in Normal and Overweight Pregnancies
2.3. GLUT1 and GLUT4 Expression in Cultured Placenta Explants from Normal Weight and Overweight Pregnancies
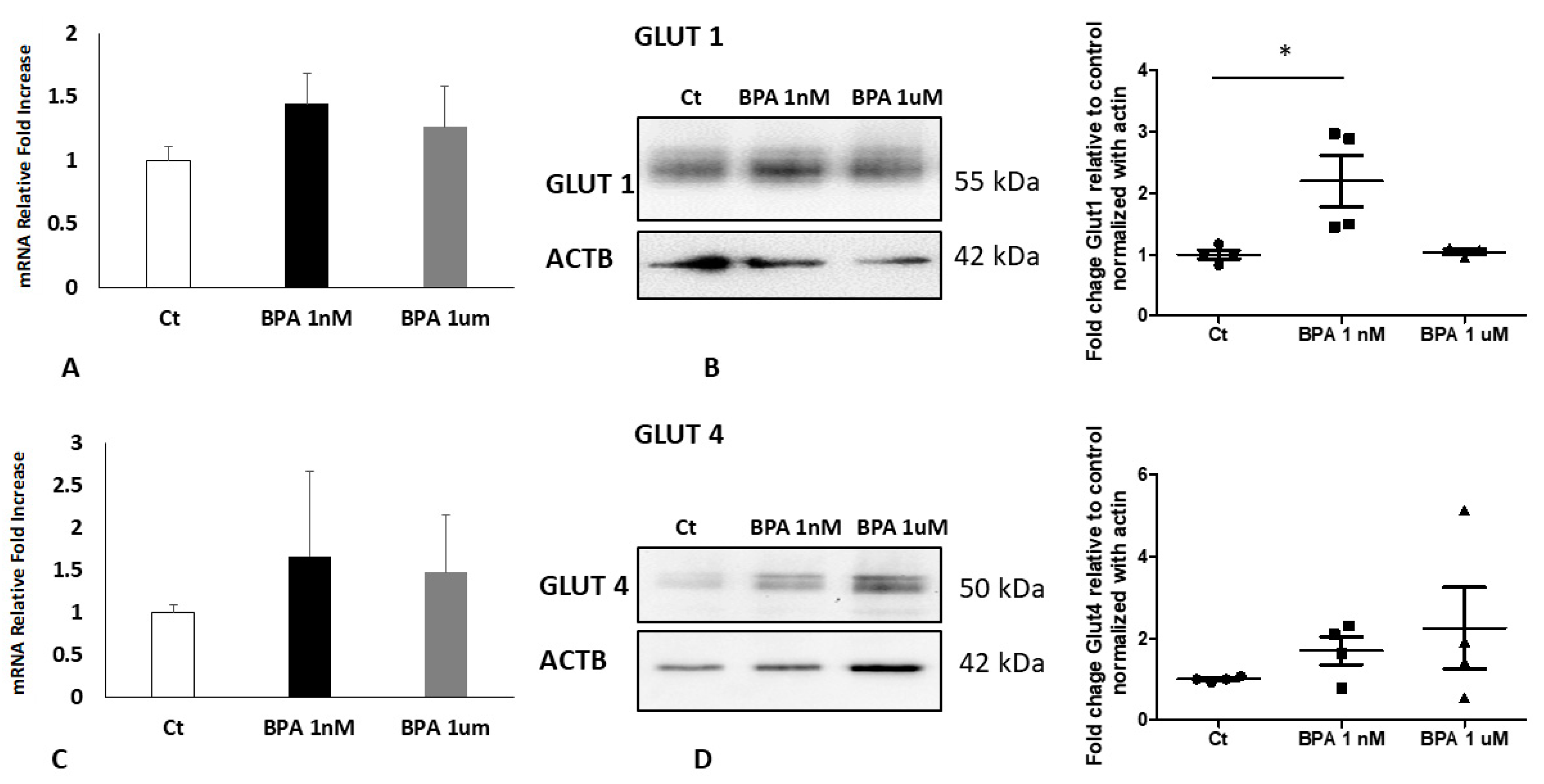
2.4. BPA Impact in Placenta Explants from Normal Weight Pregnancies
2.5. BPA Impact in Placenta Explants from Overweight Pregnancies
3. Discussion
4. Materials and Methods
4.1. Ethics Statement and Place of Recruitment
4.2. Study Population, Definitions and Tissues Collection
4.3. Human Chorionic Villous Explants Cultures and BPA Treatment
4.4. RNA Isolation and Real Time PCR
4.5. Western Blot Analyses
4.6. Antibodies Used in the Research
4.7. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vandenberg, L.N.; Chahoud, I.; Heindel, J.J.; Padmanabhan, V.; Paumgartten, F.J.; Schoenfelder, G. Urinary, circulating, and tissue biomonitoring studies indicate widespread exposure to bisphenol A. Cien Saude Colet 2012, 17, 407–434. [Google Scholar] [CrossRef]
- Cao, X.L.; Perez-Locas, C.; Robichaud, A.; Clement, G.; Popovic, S.; Dufresne, G.; Dabeka, R.W. Levels and temporal trend of bisphenol A in composite food samples from Canadian Total Diet Study 2008–2012. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2015, 32, 2154–2160. [Google Scholar] [CrossRef]
- He, P.J.; Zheng, Z.; Zhang, H.; Shao, L.M.; Tang, Q.Y. PAEs and BPA removal in landfill leachate with Fenton process and its relationship with leachate DOM composition. Sci. Total Environ. 2009, 407, 4928–4933. [Google Scholar] [CrossRef] [PubMed]
- Valentino, R.; D’Esposito, V.; Ariemma, F.; Cimmino, I.; Beguinot, F.; Formisano, P. Bisphenol A environmental exposure and the detrimental effects on human metabolic health: Is it necessary to revise the risk assessment in vulnerable population? J. Endocrinol. Investig. 2016, 39, 259–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolfo, A.; Nuzzo, A.M.; De Amicis, R.; Moretti, L.; Bertoli, S.; Leone, A. Fetal-maternal exposure to endocrine disruptors: Correlation with diet intake and pregnancy outcomes. Nutrients 2020, 12, 1744. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Magdalena, P.; Quesada, I.; Nadal, A. Prenatal exposure to BPA and offspring outcomes: The diabesogenic behavior of BPA. Dose Response 2015, 13, 1559325815590395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somm, E.; Schwitzgebel, V.M.; Toulotte, A.; Cederroth, C.R.; Combescure, C.; Nef, S.; Aubert, M.L.; Huppi, P.S. Perinatal exposure to bisphenol a alters early adipogenesis in the rat. Environ. Health Perspect 2009, 117, 1549–1555. [Google Scholar] [CrossRef] [Green Version]
- Bertoli, S.; Leone, A.; Battezzati, A. Human bisphenol a exposure and the diabesity phenotype. Dose Response 2015, 13, 1559325815599173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Obesity: Preventing and Managing the Global Epidemic. Report of a WHO Consultation; World Health Organisation: Geneva, Switzerland, 2000. [Google Scholar]
- D’Souza, R.; Horyn, I.; Pavalagantharajah, S.; Zaffar, N.; Jacob, C.E. Maternal body mass index and pregnancy outcomes: A systematic review and metaanalysis. Am. J. Obs. Gynecol. MFM 2019, 1, 100041. [Google Scholar] [CrossRef]
- Dag, Z.O.; Dilbaz, B. Impact of obesity on infertility in women. J. Turk. Ger. Gynecol. Assoc. 2015, 16, 111–117. [Google Scholar]
- Han, S.; Middleton, P.; Crowther, C.A. Exercise for pregnant women for preventing gestational diabetes mellitus. Cochrane Database Syst Rev. 2012, 7, CD009021. [Google Scholar] [CrossRef]
- Metzger, B.E.; Gabbe, S.G.; Persson, B.; Buchanan, T.A.; Catalano, P.A.; Damm, P.; Dyer, A.R.; Leiva, A. International association of diabetes and pregnancy study groups recommendations on the diagnosis and classification of hyperglycemia in pregnancy. Diabetes Care 2010, 33, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benincasa, L.; Mandala, M.; Paulesu, L.; Barberio, L.; Ietta, F. Prenatal nutrition containing bisphenol a affects placenta glucose transfer: Evidence in rats and human trophoblast. Nutrients 2020, 12, 1375. [Google Scholar] [CrossRef]
- Illsley, N.P.; Baumann, M.U. Human placental glucose transport in fetoplacental growth and metabolism. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165359. [Google Scholar] [CrossRef] [PubMed]
- Ikezuki, Y.; Tsutsumi, O.; Takai, Y.; Kamei, Y.; Taketani, Y. Determination of bisphenol A concentrations in human biological fluids reveals significant early prenatal exposure. Hum. Reprod. 2002, 17, 2839–2841. [Google Scholar] [CrossRef] [Green Version]
- Myatt, L.; Maloyan, A. Obesity and placental function. Semin. Reprod. Med. 2016, 34, 42–49. [Google Scholar] [PubMed] [Green Version]
- Spagnoletti, A.; Paulesu, L.; Mannelli, C.; Ermini, L.; Romagnoli, R.; Cintorino, M.; Ietta, F. Low concentrations of Bisphenol A and para-Nonylphenol affect extravillous pathway of human trophoblast cells. Mol. Cell Endocrinol. 2015, 412, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Lu, J.; Zhang, Y.Z.; Zhang, M.; Liu, T.; Qu, X.L. Effect of Bisphenol A on invasion ability of human trophoblastic cell line BeWo. Int. J. Clin. Exp. Pathol. 2015, 8, 14355–14364. [Google Scholar]
- Ye, Y.; Tang, Y.; Xiong, Y.; Feng, L.; Li, X. Bisphenol A exposure alters placentation and causes preeclampsia-like features in pregnant mice involved in reprogramming of DNA methylation of WNT2. FASEB J. 2019, 33, 2732–2742. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Arevalo, M.; Alonso-Magdalena, P.; Servitja, J.M.; Boronat-Belda, T.; Merino, B.; Villar-Pazos, S.; Medina-Gomez, G.; Novials, A.; Quesada, I.; Nadal, A. Maternal exposure to Bisphenol-A during pregnancy increases pancreatic beta-cell growth during early life in male mice offspring. Endocrinology 2016, 157, 4158–4171. [Google Scholar] [CrossRef]
- Gaudet, L.; Ferraro, Z.M.; Wen, S.W.; Walker, M. Maternal obesity and occurrence of fetal macrosomia: A systematic review and meta-analysis. Biomed Res. Int. 2014, 2014, 640291. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.M.; Hong, Y.C.; Ha, M.; Kim, Y.; Park, H.; Kim, H.S.; Ha, E.H. Prenatal Bisphenol-A exposure affects fetal length growth by maternal glutathione transferase polymorphisms, and neonatal exposure affects child volume growth by sex: From multiregional prospective birth cohort MOCEH study. Sci. Total Environ. 2018, 612, 1433–1441. [Google Scholar] [CrossRef] [PubMed]
- Muller, J.E.; Meyer, N.; Santamaria, C.G.; Schumacher, A.; Luque, E.H.; Zenclussen, M.L.; Rodriguez, H.A.; Zenclussen, A.C. Bisphenol A exposure during early pregnancy impairs uterine spiral artery remodeling and provokes intrauterine growth restriction in mice. Sci. Rep. 2018, 8, 9196. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.J. The origins of the developmental origins theory. J. Intern. Med. 2007, 261, 412–417. [Google Scholar] [CrossRef]
- Brett, K.E.; Ferraro, Z.M.; Yockell-Lelievre, J.; Gruslin, A.; Adamo, K.B. Maternal-fetal nutrient transport in pregnancy pathologies: The role of the placenta. Int. J. Mol. Sci. 2014, 15, 16153–16185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, C.T. Control of glucose metabolism in the perinatal period. J. Dev. Physiol. 1991, 15, 81–89. [Google Scholar]
- Baumann, M.U.; Deborde, S.; Illsley, N.P. Placental glucose transfer and fetal growth. Endocrine 2002, 19, 13–22. [Google Scholar] [CrossRef]
- Ericsson, A.; Hamark, B.; Jansson, N.; Johansson, B.R.; Powell, T.L.; Jansson, T. Hormonal regulation of glucose and system A amino acid transport in first trimester placental villous fragments. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R656–R662. [Google Scholar] [CrossRef] [Green Version]
- Barros, L.F.; Yudilevich, D.L.; Jarvis, S.M.; Beaumont, N.; Baldwin, S.A. Quantitation and immunolocalization of glucose transporters in the human placenta. Placenta 1995, 16, 623–633. [Google Scholar] [CrossRef]
- Jansson, T.; Wennergren, M.; Illsley, N.P. Glucose transporter protein expression in human placenta throughout gestation and in intrauterine growth retardation. J. Clin. Endocrinol. Metab. 1993, 77, 1554–1562. [Google Scholar]
- Hay, W.W., Jr. Placental-fetal glucose exchange and fetal glucose metabolism. Trans. Am. Clin. Clim. Assoc. 2006, 117, 321–339. [Google Scholar]
- Naser, K.A.; Gruber, A.; Thomson, G.A. The emerging pandemic of obesity and diabetes: Are we doing enough to prevent a disaster? Int. J. Clin. Pract. 2006, 60, 1093–1097. [Google Scholar] [CrossRef] [PubMed]
- Barbour, L.A.; McCurdy, C.E.; Hernandez, T.L.; Kirwan, J.P.; Catalano, P.M.; Friedman, J.E. Cellular mechanisms for insulin resistance in normal pregnancy and gestational diabetes. Diabetes Care 2007, 30 (Suppl 2), S112–S119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roder, M.E.; Porte, D., Jr.; Schwartz, R.S.; Kahn, S.E. Disproportionately elevated proinsulin levels reflect the degree of impaired B cell secretory capacity in patients with noninsulin-dependent diabetes mellitus. J. Clin. Endocrinol. Metab. 1998, 83, 604–608. [Google Scholar] [CrossRef] [PubMed]
- Stanirowski, P.J.; Szukiewicz, D.; Pyzlak, M.; Abdalla, N.; Sawicki, W.; Cendrowski, K. Analysis of correlations between the placental expression of glucose transporters GLUT-1, GLUT-4 and GLUT-9 and selected maternal and fetal parameters in pregnancies complicated by diabetes mellitus. J. Matern. Fetal. Neonatal. Med. 2019, 32, 650–659. [Google Scholar] [CrossRef] [PubMed]
- Balachandiran, M.; Bobby, Z.; Dorairajan, G.; Gladwin, V.; Vinayagam, V.; Packirisamy, R.M. Decreased maternal serum adiponectin and increased insulin-like growth factor-1 levels along with increased placental glucose transporter-1 expression in gestational diabetes mellitus: Possible role in fetal overgrowth. Placenta 2021, 104, 71–80. [Google Scholar] [CrossRef]
- Najafi, F.; Hasani, J.; Izadi, N.; Hashemi-Nazari, S.S.; Namvar, Z.; Mohammadi, S.; Sadeghi, M. The effect of prepregnancy body mass index on the risk of gestational diabetes mellitus: A systematic review and dose-response meta-analysis. Obes. Rev. 2019, 20, 472–486. [Google Scholar] [CrossRef]
- Hutcheon, J.A.; Platt, R.W.; Abrams, B.; Himes, K.P.; Simhan, H.N.; Bodnar, L.M. Pregnancy weight gain charts for obese and overweight women. Obesity 2015, 23, 532–535. [Google Scholar] [CrossRef]
- Black, R.E.; Allen, L.H.; Bhutta, Z.A.; Caulfield, L.E.; de Onis, M.; Ezzati, M.; Mathers, C.; Rivera, J.; Maternal and Child Undernutrition Study Group. Maternal and child undernutrition: Global and regional exposures and health consequences. Lancet 2008, 371, 243–260. [Google Scholar] [CrossRef]
- Black, M.H.; Sacks, D.A.; Xiang, A.H.; Lawrence, J.M. The relative contribution of prepregnancy overweight and obesity, gestational weight gain, and IADPSG-defined gestational diabetes mellitus to fetal overgrowth. Diabetes Care 2013, 36, 56–62. [Google Scholar] [CrossRef] [Green Version]
- Castillo-Castrejon, M.; Powell, T.L. Corrigendum: Placental nutrient transport in gestational diabetic pregnancies. Front. Endocrinol. 2019, 10, 5. [Google Scholar] [CrossRef]
- Acosta, O.; Ramirez, V.I.; Lager, S.; Gaccioli, F.; Dudley, D.J.; Powell, T.L.; Jansson, T. Increased glucose and placental GLUT-1 in large infants of obese nondiabetic mothers. Am. J. Obstet. Gynecol. 2015, 212, 227.e1–227.e7. [Google Scholar] [CrossRef] [PubMed]
- Jones, H.N.; Woollett, L.A.; Barbour, N.; Prasad, P.D.; Powell, T.L.; Jansson, T. High-fat diet before and during pregnancy causes marked up-regulation of placental nutrient transport and fetal overgrowth in C57/BL6 mice. FASEB J. 2009, 23, 271–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coleman, M.A.; McCowan, L.M.; North, R.A. Mid-trimester uterine artery Doppler screening as a predictor of adverse pregnancy outcome in high-risk women. Ultrasound Obstet. Gynecol. 2000, 15, 7–12. [Google Scholar] [CrossRef] [PubMed]
- James-Allan, L.B.; Arbet, J.; Teal, S.B.; Powell, T.L.; Jansson, T. Insulin stimulates GLUT4 trafficking to the syncytiotrophoblast basal plasma membrane in the human placenta. J. Clin. Endocrinol. Metab. 2019, 104, 4225–4238. [Google Scholar] [CrossRef]
- Rajakumar, C.; Guan, H.; Langlois, D.; Cernea, M.; Yang, K. Bisphenol A disrupts gene expression in human placental trophoblast cells. Reprod. Toxicol. 2015, 53, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Gut, P.; Verdin, E. The nexus of chromatin regulation and intermediary metabolism. Nature 2013, 502, 489–498. [Google Scholar] [CrossRef]
- Mitsuya, K.; Parker, A.N.; Liu, L.; Ruan, J.; Vissers, M.C.M.; Myatt, L. Alterations in the placental methylome with maternal obesity and evidence for metabolic regulation. PLoS ONE 2017, 12, e0186115. [Google Scholar] [CrossRef] [Green Version]
- Todros, T.; Piccoli, E.; Rolfo, A.; Cardaropoli, S.; Guiot, C.; Gaglioti, P.; Oberto, M.; Vasario, E.; Caniggia, I. Review: Feto-placental vascularization: A multifaceted approach. Placenta 2011, 32 (Suppl. 2), S165–S169. [Google Scholar] [CrossRef]
- Consultation, W.H.O.E. Appropriate body-mass index for Asian populations and its implications for policy and intervention strategies. Lancet 2004, 363, 157–163. [Google Scholar]
- Nuzzo, A.M.; Giuffrida, D.; Moretti, L.; Re, P.; Grassi, G.; Menato, G.; Rolfo, A. Placental and maternal sFlt1/PlGF expression in gestational diabetes mellitus. Sci. Rep. 2021, 11, 2312. [Google Scholar] [CrossRef] [PubMed]
- Hod, M.; Kapur, A.; Sacks, D.A.; Hadar, E.; Agarwal, M.; Di Renzo, G.C.; Cabero Roura, L.; McIntyre, H.D.; Morris, J.L.; Divakar, H. The International Federation of Gynecology and Obstetrics (FIGO) Initiative on gestational diabetes mellitus: A pragmatic guide for diagnosis, management, and care. Int. J. Gynaecol. Obstet. 2015, 131 (Suppl. 3), S173–S211. [Google Scholar] [CrossRef] [Green Version]
- Bertino, E.; Spada, E.; Occhi, L.; Coscia, A.; Giuliani, F.; Gagliardi, L.; Gilli, G.; Bona, G.; Fabris, C.; De Curtis, M.; et al. Neonatal anthropometric charts: The Italian neonatal study compared with other European studies. J. Pediatr. Gastroenterol. Nutr. 2010, 51, 353–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caniggia, I.; Mostachfi, H.; Winter, J.; Gassmann, M.; Lye, S.J.; Kuliszewski, M.; Post, M. Hypoxia-inducible factor-1 mediates the biological effects of oxygen on human trophoblast differentiation through TGFbeta(3). J. Clin. Investig. 2000, 105, 577–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Ermini, L.; Bhattacharjee, J.; Spagnoletti, A.; Bechi, N.; Aldi, S.; Ferretti, C.; Bianchi, L.; Bini, L.; Rosati, F.; Paulesu, L.; et al. Oxygen governs Galbeta1-3GalNAc epitope in human placenta. Am. J. Physiol. Cell Physiol. 2013, 305, C931–C940. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Normal Weight (n = 32) | Overweight (n = 26) | p Value | |
|---|---|---|---|
| Pre-Pregnancy | |||
| Nulliparae (%) | 46.9 | 22.2 * | p = 0.049 |
| Maternal age (years) | 33.4 ± 0.6 (25–39) | 33 ± 0.7 (24–39) | p > 0.05 |
| Caucasian ethnicity (%) | 100 | 100 | p > 0.05 |
| Pre-pregnancy weight (Kg) | 59 ± 1.3 | 80.3 ± 1.9 * | p < 0.001 |
| Height (cm) | 166 ± 1.3 | 163.6 ± 1.1 | p > 0.05 |
| Pre-pregnancy BMI (kg/m2) | 21.4 ± 0.3 | 30 ± 0.5 * | p < 0.001 |
| Outcomes | |||
| Delivery BMI (kg/m2) | 26.4 ± 0.4 | 33.5 ± 0.6 * | p < 0.001 |
| GWG (kg) | 13.8 ± 0.7 | 9.4 ± 0.9 * | p < 0.001 |
| Plicometry (cm) | 2.5 ± 0.1 | 3.3 ± 0.1 * | p < 0.001 |
| Mid-upper arm circumference (cm) | 25.2 ± 0.4 | 30.2 ± 0.7 * | p < 0.001 |
| Systolic Blood Pressure (mm Hg) | 114.7 ± 2.3 | 122.3 ± 1.5 * | p = 0.012 |
| Diastolic Blood Pressure (mm Hg) | 71.6 ± 1.4 | 72.3 ± 1.6 | p > 0.05 |
| GDM (%) | 12.5 | 37 * | p = 0.027 |
| GDM Therapy (%) | Diet: 100 Insulin: 0 | Diet: 100 Insulin: 0 | p > 0.05 |
| NORMAL WEIGHT ≤ 14 Weeks (n = 32) | OVERWEIGHT ≤ 14 Weeks (n = 26) | NORMAL WEIGHT ≥ 27 Weeks (n = 32) | OVERWEIGHT ≥ 27 Weeks (n = 26) | p Value | |
|---|---|---|---|---|---|
| Gestational age at blood sampling (weeks) | 13.4 ± 0.2 | 13.2 ± 0.2 | 35 ± 0.4 | 34.4 ± 0.5 | p > 0.05 |
| Maternal blood parameters: | |||||
| Glycemia (mg/dL) | 72.8 ± 1.2 | 76.8 ± 2.2 | 68.9 ± 2.1 | 79.2 ± 2.3 § | NW 3tr vs. OW 3tr: p = 0.006 |
| Triglycerides (mg/dL) | 104.5 ± 7.4 | 154.7 ± 20.5 ^ | 214.1 ± 16.4 ^ | 254.3 ± 23.1 ° | NW 2tr vs. OW 2tr: p = 0.018 NW 2tr vs. NW 3tr: p < 0.001 OW 2tr vs. OW 3tr: p = 0.002 |
| Total cholesterol (mg/dL) | 195.2 ± 6.8 | 199.9 ± 5.2 | 259.6 ± 10.3 | 247.3 ± 8.1° | OW 2tr vs. OW 3tr: p < 0.001 |
| HDL cholesterol (mg/dL) | 78.5 ± 2.5 | 71.4 ± 2.3 | 75.7 ± 3.4 | 76.5 ± 3 | p > 0.05 |
| TSH (mU/L) | 1.7 ± 0.2 | 1.6 ± 0.2 | 1.8 ± 0.2 | 1.8 ± 0.2 | p > 0.05 |
| FT4 (ng/dL) | 8.2 ± 1 | 7.2 ± 1 | 6.7 ± 1 | 7.1 ± 0.9 | p > 0.05 |
| Vit. D (ng/mL) | 28.6 ± 6 | 33.7 ± 7.9 | 51.5 ± 14 | 43 ± 10.1 | p > 0.05 |
| Homocysteine (mcmol/L) | 6.1 ± 0.2 | 6.3 ± 0.2 | 6.2 ± 0.3 | 6.8 ± 0.7 | p > 0.05 |
| White Bood Cells (109/L) | 7.95 ± 0.3 | 8.6 ± 0.5 | 9.3 ± 0.3 | 9.9 ± 0.5 | p > 0.05 |
| Red Blood Cells (1012/L) | 4.1 ± 0.1 | 4.1 ± 0.01 | 3.9 ± 0.1 | 4.15 ± 0.1 | p > 0.05 |
| Hemoglobin (g/dL) | 12.2 ± 0.2 | 12.6 ± 0.2 | 11.3 ± 0.2 ^ | 11.8 ± 0.2 | NW 2tr vs. NW 3tr: p = 0.013 |
| Platelet (109/L) | 238 ± 12.5 | 248 ± 9.8 | 222.4 ± 13.4 | 247.1 ± 13.4 | p > 0.05 |
| Normal Weight (n = 32) | Overweight (n = 26) | p Value | |
|---|---|---|---|
| Gestational age at delivery (weeks) | 39.9 ± 0.2 | 39.6 ± 0.2 | p > 0.05 |
| Caesarian section (%) | 31.2 | 22.2 | p > 0.05 |
| Placental weight (g) | 581.6 ± 17.4 | 584.5 ± 12.9 | p > 0.05 |
| Birth weight (g) | 3493.1 ± 58.8 | 3401.8 ± 71.4 | p > 0.05 |
| Fetal to placenta weight ratio | 6.1 ± 0.1 | 5.9 ± 0.1 | p > 0.05 |
| Sex of infant | |||
| Female (%) | 37.5 | 40.7 | p > 0.05 |
| Male (%) | 62.5 | 59.3 | p > 0.05 |
| Large for Gestational Age, AC > 90 centile (%) | 3.1 | 22.2 * | p = 0.024 |
| 5-min Apgar score ≤7 (%) | 0 | 0 | p > 0.05 |
| NICU (%) | 0 | 0 | p > 0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ermini, L.; Nuzzo, A.M.; Ietta, F.; Romagnoli, R.; Moretti, L.; Masturzo, B.; Paulesu, L.; Rolfo, A. Placental Glucose Transporters and Response to Bisphenol A in Pregnancies from of Normal and Overweight Mothers. Int. J. Mol. Sci. 2021, 22, 6625. https://doi.org/10.3390/ijms22126625
Ermini L, Nuzzo AM, Ietta F, Romagnoli R, Moretti L, Masturzo B, Paulesu L, Rolfo A. Placental Glucose Transporters and Response to Bisphenol A in Pregnancies from of Normal and Overweight Mothers. International Journal of Molecular Sciences. 2021; 22(12):6625. https://doi.org/10.3390/ijms22126625
Chicago/Turabian StyleErmini, Leonardo, Anna Maria Nuzzo, Francesca Ietta, Roberta Romagnoli, Laura Moretti, Bianca Masturzo, Luana Paulesu, and Alessandro Rolfo. 2021. "Placental Glucose Transporters and Response to Bisphenol A in Pregnancies from of Normal and Overweight Mothers" International Journal of Molecular Sciences 22, no. 12: 6625. https://doi.org/10.3390/ijms22126625
APA StyleErmini, L., Nuzzo, A. M., Ietta, F., Romagnoli, R., Moretti, L., Masturzo, B., Paulesu, L., & Rolfo, A. (2021). Placental Glucose Transporters and Response to Bisphenol A in Pregnancies from of Normal and Overweight Mothers. International Journal of Molecular Sciences, 22(12), 6625. https://doi.org/10.3390/ijms22126625