
Architecture of the Pancreatic Islets and Endocrine Cell Arrangement in the Embryonic Pancreas of the Grass Snake (Natrix natrix L.). Immunocytochemical Studies and 3D Reconstructions


Abstract
:1. Introduction
2. Results
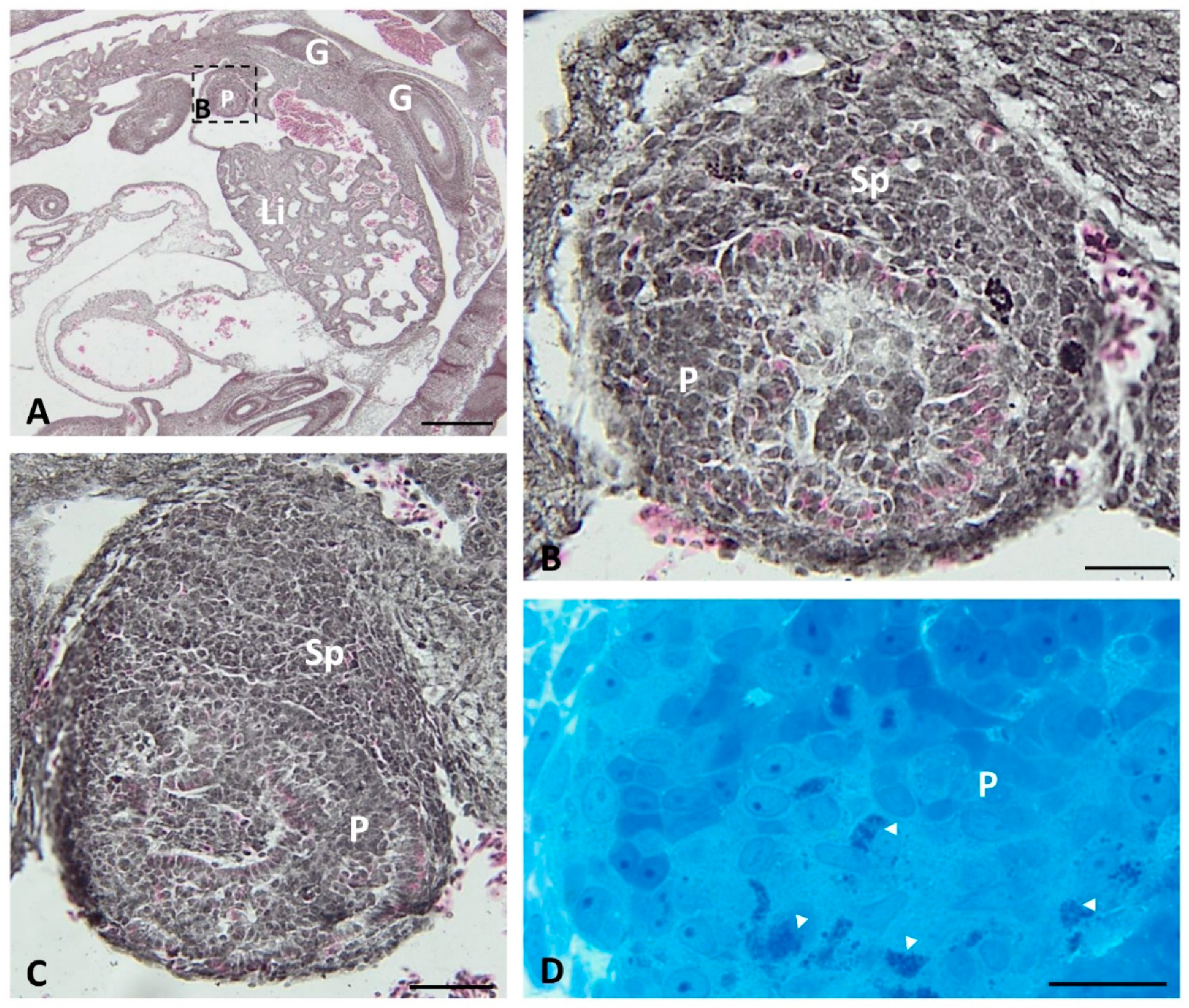
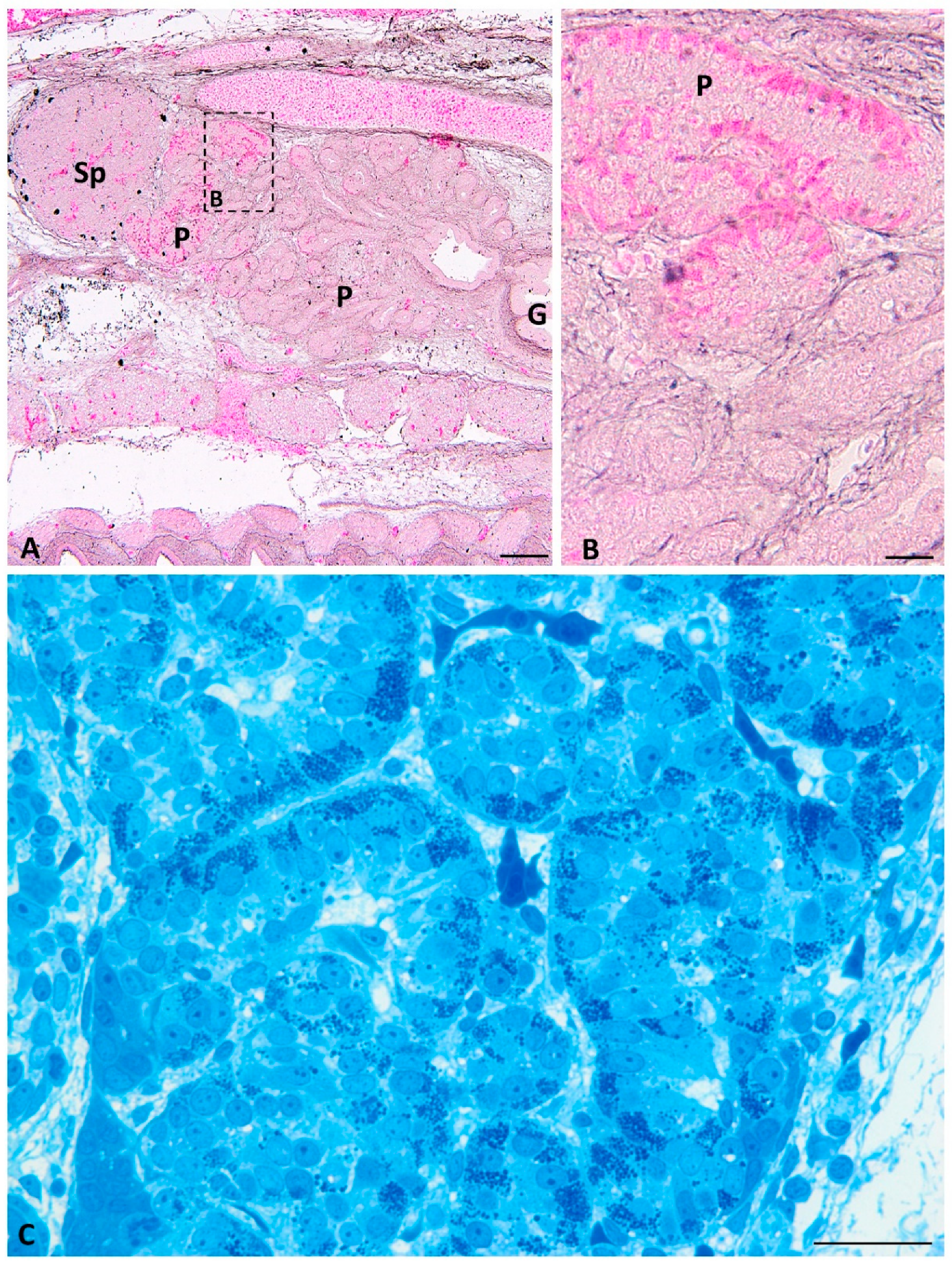
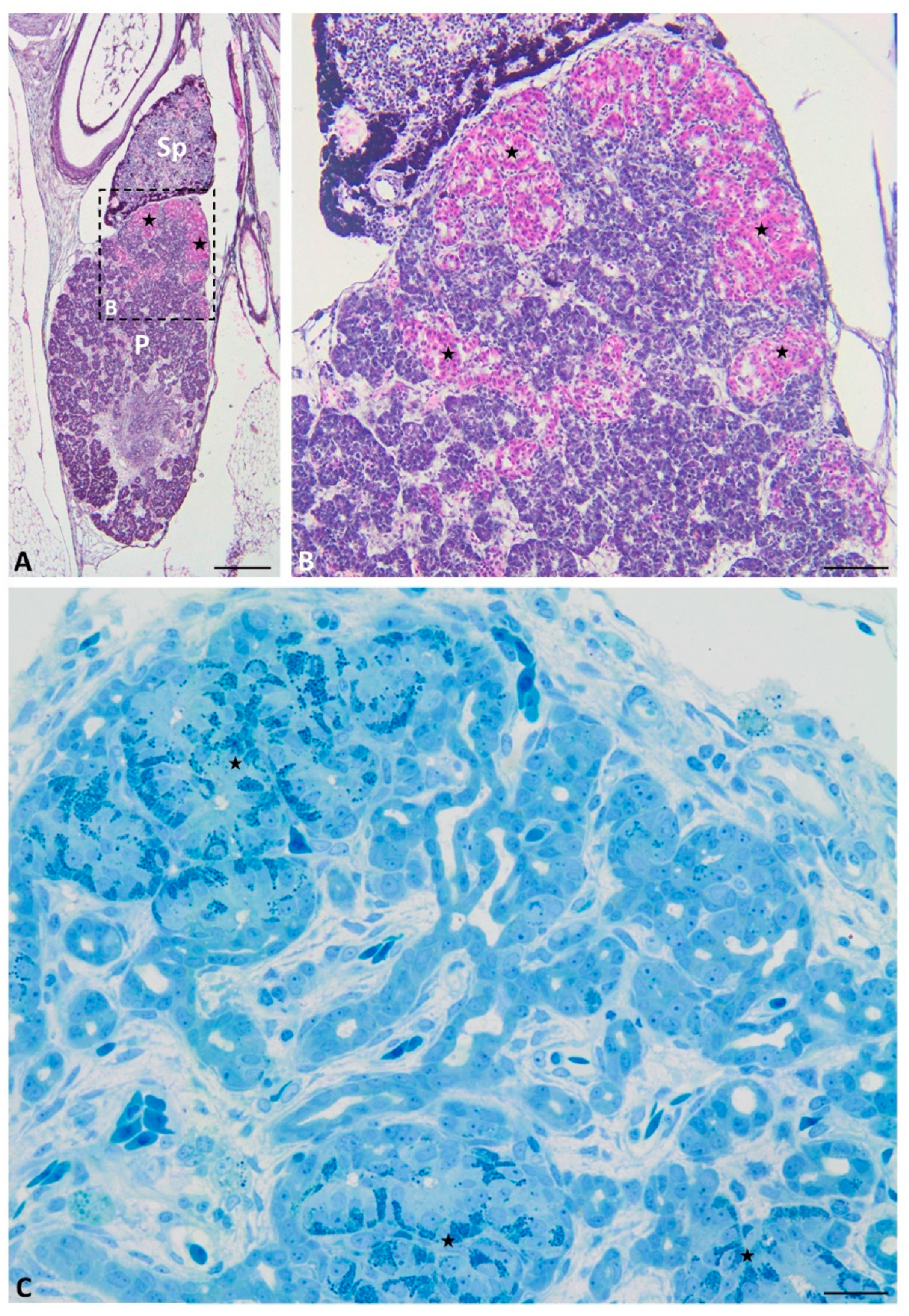
2.1. Light Microscopy
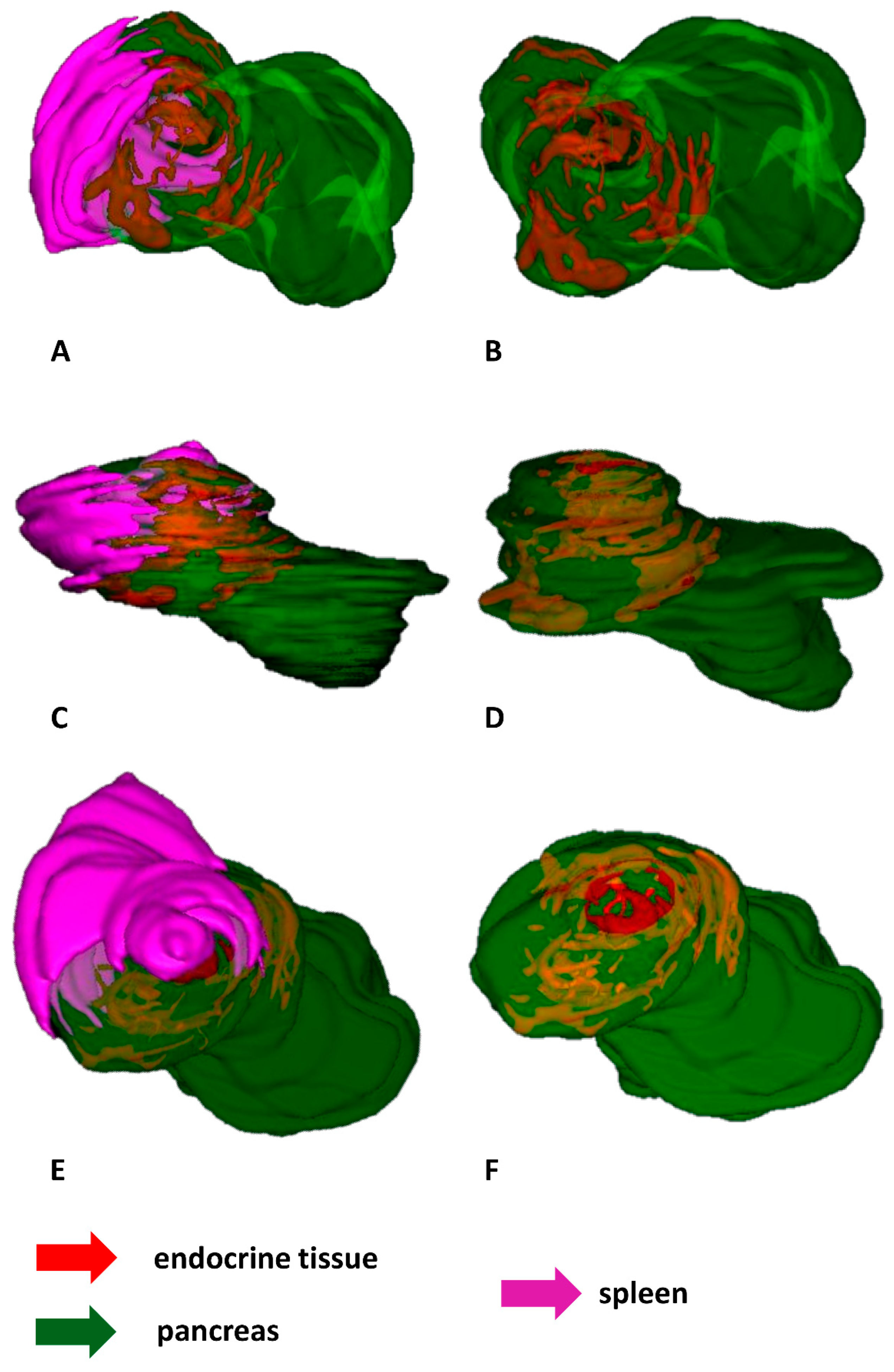
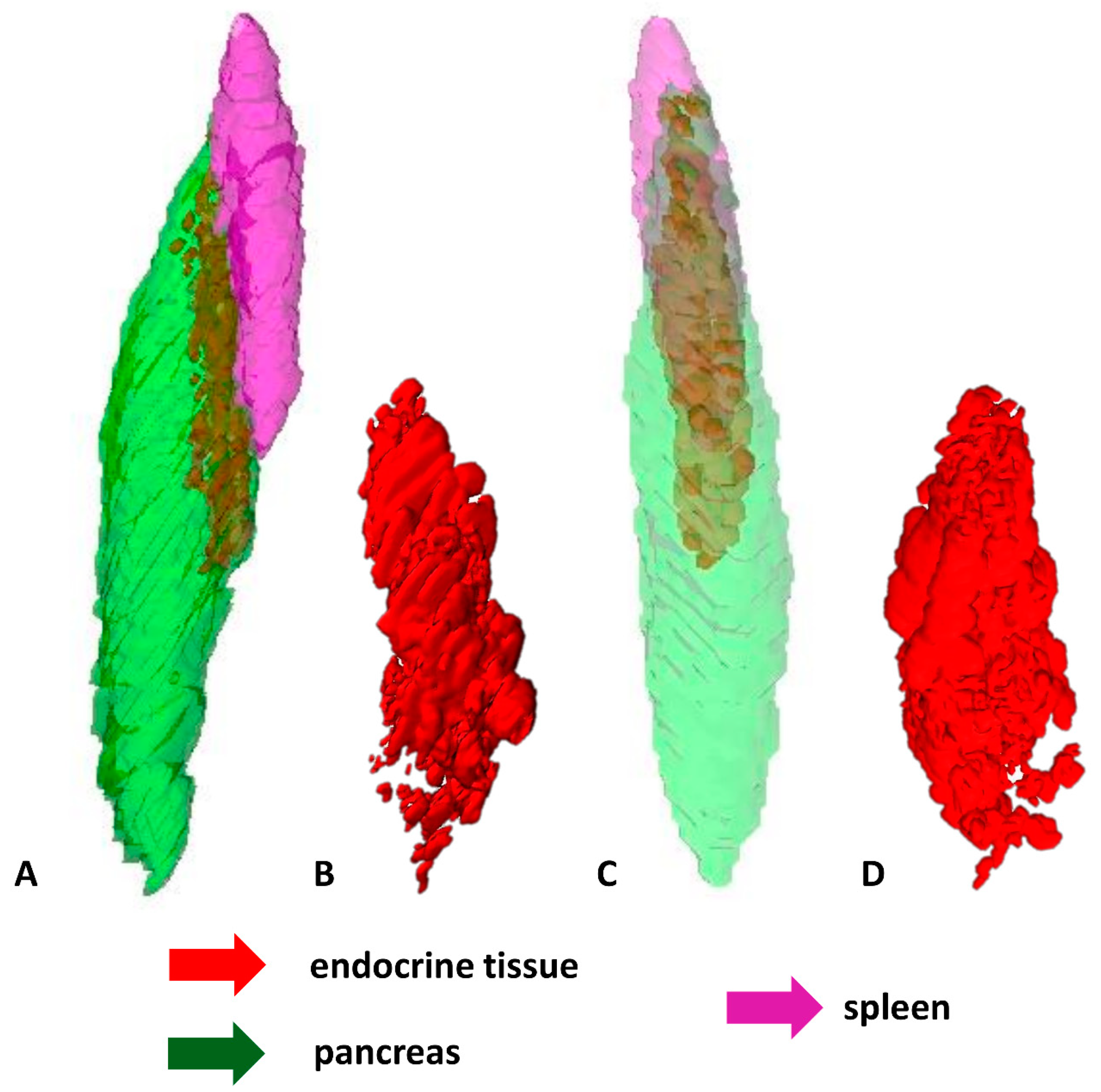
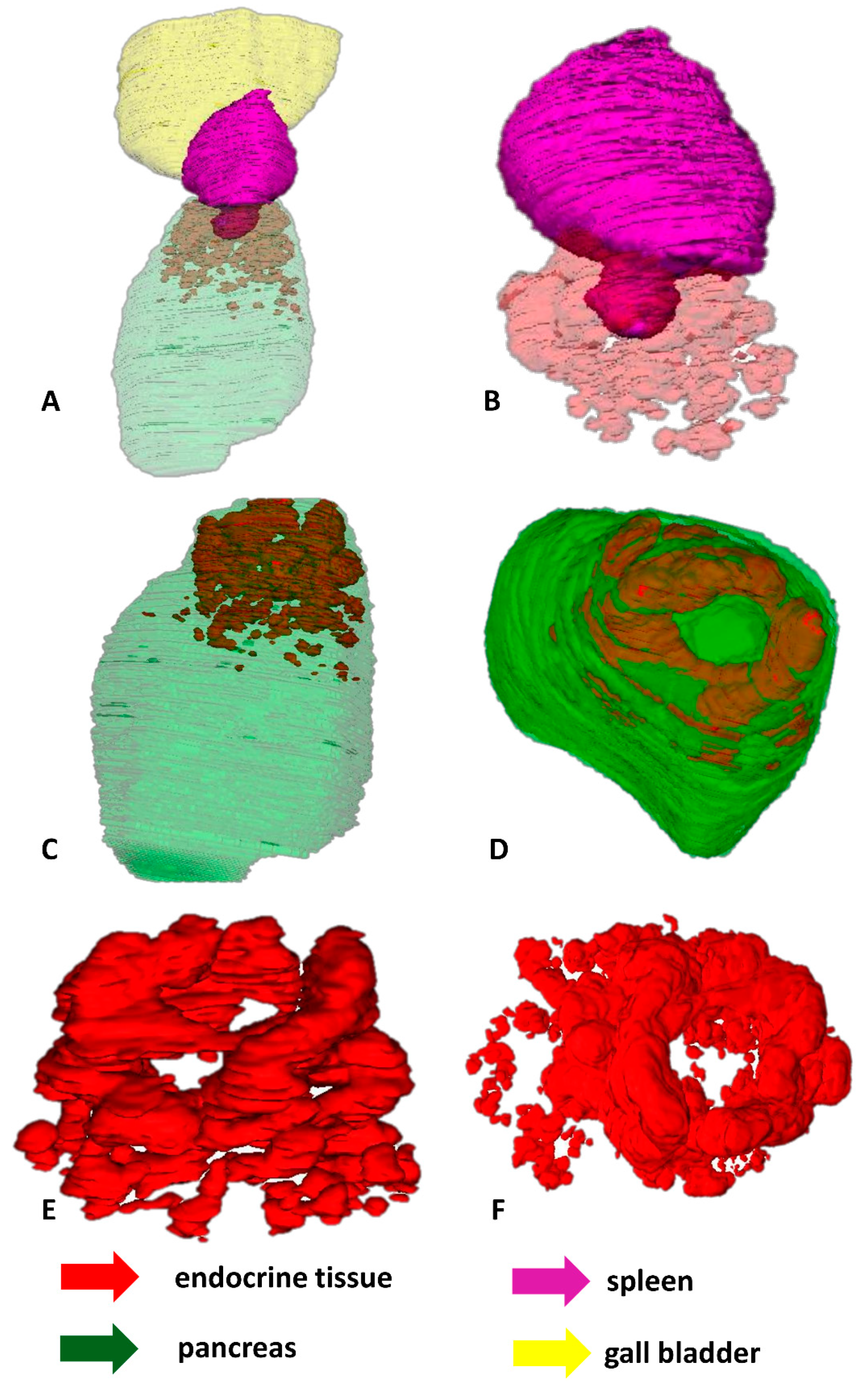
2.2. 3D Reconstructions
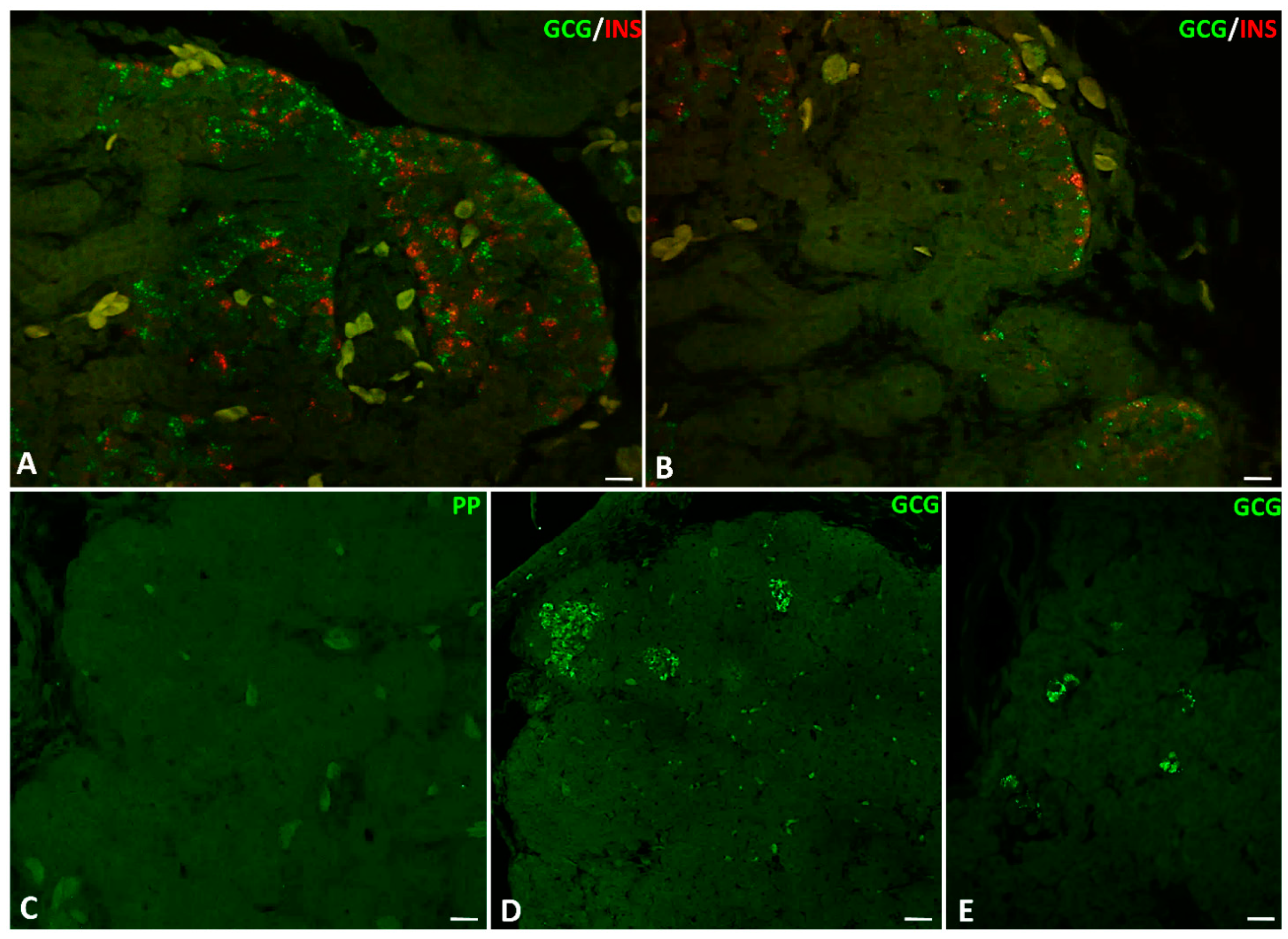
2.3. Immunofluorescence Study
3. Discussion
3.1. Changes of Islet Localization
3.2. Localization of Endocrine Cells within Pancreatic Islets
4. Materials and Methods
4.1. Manipulation of Animals and Embryos
4.2. Light Microscopy
4.3. 3D Reconstructions
4.4. Immunohistochemical Detection of Pancreatic Hormones
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cabrera, O.; Berman-Weinberg, D.M.; Kenyon, N.S.; Ricordi, C.; Berggren, P.-O.; Caicedo, A. The unique cytoarchitecture of human pancreatic islets has implications for islet cell function. Proc. Natl. Acad. Sci. USA 2006, 103, 2334–2339. [Google Scholar] [CrossRef] [Green Version]
- Slack, J.M. Developmental biology of the pancreas. Development 1995, 121, 1569–1580. [Google Scholar] [CrossRef]
- Gittes, G.K. Developmental biology of the pancreas: A comprehensive review. Dev. Biol. 2009, 326, 4–35. [Google Scholar] [CrossRef] [Green Version]
- Wierup, N.; Svensson, H.; Mulder, H.; Sundler, F. The ghrelin cell: A novel developmentally regulated islet cell in the human pancreas. Regul. Pept. 2002, 107, 63–69. [Google Scholar] [CrossRef]
- Napolitano, T.; Silvano, S.; Vieira, A.; Balaji, S.; Garrido-Utrilla, A.; Friano, M.E.; Atlija, J.; Collombat, P. Role of ghrelin in pancreatic development and function. Diabetes Obes. Metab. 2018, 20, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Ionescu-Tirgoviste, C.; Gagniuc, P.A.; Gubceac, E.; Mardare, L.; Popescu, I.; Dima, S.; Militaru, M. A 3D map of the islet routes throughout the healthy human pancreas. Sci. Rep. 2015, 5, 14634. [Google Scholar] [CrossRef] [Green Version]
- Youson, J.H.; Al-Mahrouki, A.A.; Amemiya, Y.; Graham, L.C.; Montpetit, C.J.; Irwin, D.M. The fish endocrine pancreas: Review, new data, and future research directions in ontogeny and phylogeny. Gen. Comp. Endocrinol. 2006, 148, 105–115. [Google Scholar] [CrossRef]
- El-Salhy, M.; Wilander, E.; Abu-Sinna, G. The endocrine pancreas of anuran amphibians: A histological and immunocytochemical study. Biomed. Res. 1982, 3, 579–589. [Google Scholar] [CrossRef] [Green Version]
- Francini, F.; Madsen, O.; Dumm, C.L.G.; Gagliardino, J.J. Topographic Differences in Cell Populations and Insulin Secretion in the Endocrine Pancreas of the Toad Bufo arenarum. Gen. Comp. Endocrinol. 1996, 104, 346–355. [Google Scholar] [CrossRef]
- Dupont, J.; Rideau, N.; Simon, J. Chapter 27—Endocrine Pancreas. In Sturkie’s Avian Physiology, 6th ed.; Scanes, C.G., Ed.; Academic Press: San Diego, CA, USA, 2015; pp. 613–631. [Google Scholar]
- Elayat, A.A.; El-Naggar, M.M.; Tahir, M. An immunocytochemical and morphometric study of the rat pancreatic islets. J. Anat. 1995, 186, 629–637. [Google Scholar]
- Epple, A.; Brinn, J.E. The Comparative Physiology of the Pancreatic Islets, 1st ed.; Springer: Berlin/Heidelberg, Germany, 1987. [Google Scholar]
- Rhoten, W.B. Quantitative immunocytochemical analysis of the endocrine pancreas of the Nile crocodile. Am. J. Anat. 1987, 178, 103–115. [Google Scholar] [CrossRef]
- El-Salhy, M.; Grimelius, L. Histological and immunohistochemical studies of the endocrine pancreas of lizards. Histochemistry 1981, 72, 237–247. [Google Scholar] [CrossRef]
- El-Salhy, M.; Abu-Sinna, G.; Wilander, E. The endocrine pancreas of a squamate reptile, the desert lizard (Chalcides ocellatus). Histochemistry 1983, 78, 391–397. [Google Scholar] [CrossRef]
- Thomas, T.B. The pancreas of snakes. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 1942, 82, 327–345. [Google Scholar] [CrossRef]
- Björkman, N.; Hellerström, C.; Hellman, B.; Petersson, B. The cell types in the endocrine pancreas of the human fetus. Z. Zellforsch. Mikrosk. Anat. 1966, 72, 425–445. [Google Scholar] [CrossRef]
- Maňáková, E.; Titlbach, M. Development of the Chick Pancreas with Regard to Estimation of the Relative Occurrence and Growth of Endocrine Tissue. Anat. Histol. Embryol. 2007, 36, 127–134. [Google Scholar] [CrossRef]
- Kaung, H.-L.C. Immunocytochemical localization of pancreatic endocrine cells in frog embryos and young larvae. Gen. Comp. Endocrinol. 1981, 45, 204–211. [Google Scholar] [CrossRef]
- Mikami, S.-I.; Sudo, S.; Taniguchi, K.; Yamada, S. Immunocytochemical studies on the development of the pancreatic islet cells in the domestic fowl. Jpn. J. Vet. Sci. 1986, 48, 769–780. [Google Scholar] [CrossRef]
- Reddy, S.; Bibby, N.J.; Elliott, R.B. An immunocytochemical study of endocrine cell development in the early fetal guinea pig pancreas. Gen. Comp. Endocrinol. 1992, 86, 275–283. [Google Scholar] [CrossRef]
- Bricout-Neveu, E.; Pechberty, S.; Reynaud, K.; Maenhoudt, C.; José Lecomte, M.; Ravassard, P.; Czernichow, P. Development of the Endocrine Pancreas in the Beagle Dog: From Fetal to Adult Life. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2017, 300, 1429–1438. [Google Scholar] [CrossRef] [Green Version]
- Gupta, D.; Uppal, V.; Bansal, N.; Gupta, A. Differentiation of pancreatic endocrine islets in buffalo fetus. Indian J. Anim. Sci. 2020, 90, 693–697. [Google Scholar]
- Nagaya, M.; Hayashi, A.; Nakano, K.; Honda, M.; Hasegawa, K.; Okamoto, K.; Itazaki, S.; Matsunari, H.; Watanabe, M.; Umeyama, K.; et al. Distributions of endocrine cell clusters during porcine pancreatic development. PLoS ONE 2019, 14, e0216254. [Google Scholar] [CrossRef]
- Da Silva, A.B.S.; Fonseca, C.M.B.; Cavalcante, M.M.A.D.S.; De Oliveira, I.M.; Ferraz, M.S.; Viana, F.J.C.; Fontenele, R.D.; Conde Júnior, A.M. Histomorphometry of pancreas development in hybrid chicken (Galus galus) embryo and fetus. Microsc. Res. Tech. 2018, 81, 614–623. [Google Scholar] [CrossRef]
- Hara, A.; Kadoya, Y.; Kojima, I.; Yamashina, S. Rat pancreatic islet is formed by unification of multiple endocrine cell clusters. Dev. Dyn. 2007, 236, 3451–3458. [Google Scholar] [CrossRef]
- Jeon, J.; Correa-Medina, M.; Ricordi, C.; Edlund, H.; Diez, J.A. Endocrine Cell Clustering During Human Pancreas Development. J. Histochem. Cytochem. 2009, 57, 811–824. [Google Scholar] [CrossRef] [Green Version]
- Miller, K.; Kim, A.; Kilimnik, G.; Jo, J.; Moka, U.; Periwal, V.; Hara, M. Islet Formation during the Neonatal Development in Mice. PLoS ONE 2009, 4, e7739. [Google Scholar] [CrossRef] [Green Version]
- Şimşek, N.; Bayraktaroğlu, A.G.; Altunay, H. Localization of insulin immunpositive cells and histochemical structure of the pancreas in falcons (Falco naumanni). Ankara Üniv. Vet. Fak. Derg. 2009, 56, 241–247. [Google Scholar] [CrossRef]
- Rawdon, B.B. Morphogenesis and differentiation of the avian endocrine pancreas, with particular reference to experimental studies on the chick embryo. Microsc. Res. Tech. 1998, 43, 292–305. [Google Scholar] [CrossRef]
- Buchan, A.M.J. An immunocytochemical study of endocrine pancreas of snakes. Cell Tissue Res. 1984, 235. [Google Scholar] [CrossRef]
- Moscona, A.A. Anatomy of the pancreas and langerhans islets in snakes and lizards. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 1990, 227, 232–244. [Google Scholar] [CrossRef]
- Youson, J.H.; Al-Mahrouki, A.A. Ontogenetic and Phylogenetic Development of the Endocrine Pancreas (Islet Organ) in Fishes. Gen. Comp. Endocrinol. 1999, 116, 303–335. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Ku, S.K.; Park, K.D.; Lee, H.S. Comparative study of endocrine cells in the principal pancreatic islets of two teleosts, Silurus asotus (Siluridae) and Siniperca scherzeri (Centropomidae). J. Vet. Sci. 2001, 2, 75–80. [Google Scholar] [CrossRef]
- Li, Z.; Wen, C.; Peng, J.; Korzh, V.; Gong, Z. Generation of living color transgenic zebrafish to trace somatostatin-expressing cells and endocrine pancreas organization. Differentiation 2009, 77, 128–134. [Google Scholar] [CrossRef]
- Fortin, J.S.; Santamaria-Bouvier, A.; Lair, S.; Dallaire, A.D.; Benoit-Biancamano, M.-O. Anatomic and molecular characterization of the endocrine pancreas of a teleostean fish: Atlantic wolffish (Anarhichas lupus). Zool. Stud. 2015, 54, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Saito, K.; Iwama, N.; Takahashi, T. Morphometrical analysis on topographical difference in size distribution, number and volume of islets in the human pancreas. Tohoku J. Exp. Med. 1978, 124, 177–186. [Google Scholar] [CrossRef] [Green Version]
- Ravi, P.K.; Purkait, S.; Agrawal, U.; Patra, S.; Patnaik, M.; Singh, S.R.; Mishra, P.R. Regional variation of human pancreatic islets dimension and its impact on beta cells in Indian population. Islets 2019, 11, 141–151. [Google Scholar] [CrossRef]
- Miller, M.R. Observations on the comparative histology of the reptilian pancreatic islet. Gen. Comp. Endocrinol. 1962, 2, 407–414. [Google Scholar] [CrossRef]
- Kilimnik, G.; Jo, J.; Periwal, V.; Zielinski, M.; Hara, M. Quantification of islet size and architecture. Islets 2012, 4, 167–172. [Google Scholar] [CrossRef] [Green Version]
- Rhoten, W.B. Immunocytochemical localization of four hormones in the pancreas of the garter snake, Thamnophis sirtalis. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 1984, 208, 233–242. [Google Scholar] [CrossRef]
- Hellerström, C.; Asplund, K. The two types of A-cells in the pancreatic islets of snakes. Z. Zellforsch. Mikrosk. Anat. 1966, 70, 68–80. [Google Scholar] [CrossRef]
- Masini, M.A. Immunocytochemical localization of peptides in the endocrine pancreas of the snakes Vipera aspis and Natrix maura. Acta Histochem. 1988, 84, 111–119. [Google Scholar] [CrossRef]
- Teitelman, G.; Joh, T.H.; Reis, D.J. Transformation of catecholaminergic precursors into glucagon (A) cells in mouse embryonic pancreas. Proc. Natl. Acad. Sci. USA 1981, 78, 5225–5229. [Google Scholar] [CrossRef] [Green Version]
- Proshchina, A.E.; Krivova, Y.; Barabanov, V.M.; Saveliev, S.V. Pancreatic endocrine cell arrangement during human ontogeny. Acta Histochem. 2019, 121, 638–645. [Google Scholar] [CrossRef]
- Rhoten, W.B.; Hall, C.E. An immunocytochemical study of the cytogenesis of pancreatic endocrine cells in the lizard, Anolis carolinensis. Am. J. Anat. 1982, 163, 181–193. [Google Scholar] [CrossRef]
- Jackintell, L.A.; Lance, V.A. Ontogeny and Regional Distribution of Hormone-Producing Cells in the Embryonic Pancreas of Alligator mississippiensis. Gen. Comp. Endocrinol. 1994, 94, 244–260. [Google Scholar] [CrossRef]
- Robb, P. The development of the islets of Langerhans in the human foetus. Q. J. Exp. Physiol. Cogn. Med Sci. 1961, 46, 335–343. [Google Scholar] [CrossRef]
- Steiner, D.J.; Kim, A.; Miller, K.; Hara, M. Pancreatic islet plasticity: Interspecies comparison of islet architecture and composition. Islets 2010, 2, 135–145. [Google Scholar] [CrossRef] [Green Version]
- Dolenšek, J.; Rupnik, M.S.; Stožer, A. Structural similarities and differences between the human and the mouse pancreas. Islets 2015, 7, e1024405. [Google Scholar] [CrossRef] [Green Version]
- Baetens, D.; Malaisse-Lagae, F.; Perrelet, A.; Orci, L. Endocrine pancreas: Three-dimensional reconstruction shows two types of islets of langerhans. Science 1979, 206, 1323–1325. [Google Scholar] [CrossRef]
- Trimble, E.R.; Halban, P.A.; Wollheim, C.B.; Renold, A.E. Functional differences between rat islets of ventral and dorsal pan-creatic origin. J. Clin. Investig. 1982, 69, 405–413. [Google Scholar] [CrossRef]
- Moede, T.; Leibiger, I.B.; Berggren, P.-O. Alpha cell regulation of beta cell function. Diabetologia 2020, 63, 2064–2075. [Google Scholar] [CrossRef]
- Trandaburu, T.; Calugareanu, L. Light and electron microscopic investigation of the endocrine pancreas of the grass-snake [Natrix n. natrix (L.)]. Z. Zellforsch. Mikrosk. Anat. 1969, 97, 212–225. [Google Scholar] [CrossRef]
- Rupik, W. Hollowing or cavitation during follicular lumen formation in the differentiating thyroid of grass snake Natrix natrix L. (Lepidosauria, Serpentes) embryos? An ultrastructural study. Zoology 2012, 115, 389–397. [Google Scholar] [CrossRef]
- Rupik, W. Ultrastructural studies of cilia formation during thyroid gland differentiation in grass snake embryos. Micron 2013, 44, 228–237. [Google Scholar] [CrossRef]
- Kowalska, M.; Rupik, W. Development of pancreatic acini in embryos of the grass snake Natrix natrix (Lepidosauria, Serpentes). J. Morphol. 2020, 281, 110–121. [Google Scholar] [CrossRef]
- Kaczmarek, P.; Rupik, W. Structural and ultrastructural studies on the developing vomeronasal sensory epithelium in the grass snake Natrix natrix (Squamata: Colubroidea). J. Morphol. 2021, 282, 378–407. [Google Scholar] [CrossRef]
- Kaczmarek, P.; Hermyt, M.; Rupik, W. Embryology of the VNO and associated structures in the grass snake Natrix natrix (Squamata: Natricinae): A 3D perspective. Front. Zool. 2017, 14, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Kowalska, M.; Rupik, W. Ultrastructure of endocrine pancreatic granules during pancreatic differentiation in the grass snake, Natrix natrix L. (Lepidosauria, Serpentes). J. Morphol. 2018, 279, 330–348. [Google Scholar] [CrossRef]
- Rupik, W. Early development of the adrenal glands in the grass snake Natrix natrix L. (Lepidosauria, Serpentes). Adv. Anat. Embryol. Cell Biol. 2002, 164. [Google Scholar] [CrossRef]
- Baier, J. Reptiles. In Guidelines for the Euthanasia of Non-Domestic Animals; American Association of Zoo Veterinarians: Yulee, FL, USA, 2006; pp. 42–45. [Google Scholar]
- Conroy, C.J.; Papenfuss, T.; Parker, J.; Hahn, N.E. Use of Tricaine Methanesulfonate (MS222) for Euthanasia of Reptiles. J. Am. Assoc. Lab. Anim. Sci. 2009, 48, 28–32. [Google Scholar]
- Bagiński, S. Technika Mikroskopowa; Państwowe Wydawn: Warszawa, Poland, 1969. [Google Scholar]
- Rupik, W.; Swadźba, E.; Dubińska-Magiera, M.; Jędrzejowska, I.; Daczewska, M. Reptilian myotomal myogenesis—Lessons from the sand lizard Lacerta agilis L. (Reptilia, Lacertidae). Zoology 2012, 115, 330–338. [Google Scholar] [CrossRef]
- Rupik, W.; Kowalska, M.; Swadźba, E.; Maślak, R. Ultrastructural features of the differentiating thyroid primordium in the sand lizard (Lacerta agilis L.) from the differentiation of the cellular cords to the formation of the follicular lumen. Zoology 2016, 119, 97–112. [Google Scholar] [CrossRef]
- Kaczmarek, P.; Janiszewska, K.; Metscher, B.; Rupik, W. Development of the squamate naso-palatal complex: Detailed 3D analysis of the vomeronasal organ and nasal cavity in the brown anole Anolis sagrei (Squamata: Iguania). Front. Zool. 2020, 17, 1–34. [Google Scholar] [CrossRef]
- Kaczmarek, P.; Metscher, B.; Rupik, W. Embryology of the naso-palatal complex in Gekkota based on detailed 3D analysis in Lepidodactylus lugubris and Eublepharis macularius. J. Anat. 2021, 238, 249–287. [Google Scholar] [CrossRef]
- Zadorozhnyĭ, V.V. Modified histological methods using a nuclear dye gallocyanine. Arkh. Patol. 1982, 44, 67–68. [Google Scholar]
- Kowalska, M.; Rupik, W. Development of endocrine pancreatic islets in embryos of the grass snake Natrix natrix (Lepidosauria, Serpentes). J. Morphol. 2019, 280, 103–118. [Google Scholar] [CrossRef] [Green Version]
- Rasband, W.S. ImageJ. U.S.; National Institutes of Health: Bethesda, MD, USA, 1997–2016. Available online: https://imagej.nih.gov/ij/ (accessed on 14 June 2021).
- Kowalska, M.; Hermyt, M.; Rupik, W. Three-dimensional reconstruction of the embryonic pancreas in the grass snake Natrix natrix L. (Lepidosauria, Serpentes) based on histological studies. Zoology 2017, 121, 91–110. [Google Scholar] [CrossRef]
- Kowalska, M.; Rupik, W. Development of the duct system during exocrine pancreas differentiation in the grass snake Natrix natrix (Lepidosauria, Serpentes). J. Morphol. 2018, 279, 724–746. [Google Scholar] [CrossRef]
- Walter, I.; Fleischmann, M.; Klein, D.; Müller, M.; Salmons, B.; Günzburg, W.H.; Renner, M.; Gelbmann, W. Rapid and sensitive detection of enhanced green fluorescent protein expression in paraffin sections by confocal laser scanning microscopy. Histochem. J. 2000, 32, 99–103. [Google Scholar] [CrossRef]
- Rupik, W.; Stawierej, A.; Stolarczyk, I.; Widłak, W. Promoter of the heat shock testis-specific Hsp70.2/Hst70 gene is active in nervous system during embryonic development of mice. Anat. Embryol. 2006, 211, 631–638. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Incubation Time (Days) | Total Length (mm) | Head Length (mm) | Head Width (mm) | Head Height (mm) | Tail Length (mm) | Embryo Age Defined by Stages |
|---|---|---|---|---|---|---|
| 0 | 26.50 | 2.30 | 1.40 | 3.90 | No | I |
| 1 | 27.00 | 2.50 | 1.60 | 4.40 | No | |
| 2 | 30.00 | 3.20 | 1.80 | 4.80 | No | II |
| 3 | 31.50 | 3.70 | 1.80 | 4.90 | 2.00 | |
| 4 | 32.10 | 3.90 | 2.00 | 4.50 | 3.00 | III |
| 5 | 34.90 | 3.90 | 2.10 | 4.30 | 4.30 | IV |
| 6 | 43.80 | 4.10 | 2.20 | 4.20 | 5.20 | |
| 7 | 48.70 | 4.50 | 2.40 | 4.20 | 5.70 | V |
| 8 | 58.50 | 4.80 | 2.40 | 4.00 | 7.80 | |
| 9 | 79.30 | 4.80 | 2.50 | 4.00 | 9.20 | |
| 10 | 82.70 | 5.20 | 2.70 | 3.80 | 11.30 | VI |
| 12 | 87.60 | 6.40 | 2.70 | 3.70 | 11.90 | |
| 14 | 98.20 | 7.10 | 2.90 | 3.70 | 15.80 | VII |
| 17 | 133.70 | 8.00 | 3.30 | 3.50 | 17.10 | VIII |
| 20 | 144.20 | 8.30 | 3.80 | 3.30 | 25.90 | IX |
| 22 | 153.10 | 8.70 | 3.90 | 3.20 | 29.30 | X |
| 25 | 176.30 | 9.40 | 4.00 | 3.00 | 30.70 | XI |
| 27 | 184.70 | 11.30 | 4.20 | 3.00 | 34.20 | |
| 31 | 200.00 | 11.80 | 4.70 | 3.00 | 38.50 | XII |
| 32 | 211.30 | 12.00 | 4.70 | 3.00 | 40.00 | |
| 33 | 211.80 | 12.00 | 4.70 | 3.00 | 40.10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kowalska, M.; Rupik, W. Architecture of the Pancreatic Islets and Endocrine Cell Arrangement in the Embryonic Pancreas of the Grass Snake (Natrix natrix L.). Immunocytochemical Studies and 3D Reconstructions. Int. J. Mol. Sci. 2021, 22, 7601. https://doi.org/10.3390/ijms22147601
Kowalska M, Rupik W. Architecture of the Pancreatic Islets and Endocrine Cell Arrangement in the Embryonic Pancreas of the Grass Snake (Natrix natrix L.). Immunocytochemical Studies and 3D Reconstructions. International Journal of Molecular Sciences. 2021; 22(14):7601. https://doi.org/10.3390/ijms22147601
Chicago/Turabian StyleKowalska, Magdalena, and Weronika Rupik. 2021. "Architecture of the Pancreatic Islets and Endocrine Cell Arrangement in the Embryonic Pancreas of the Grass Snake (Natrix natrix L.). Immunocytochemical Studies and 3D Reconstructions" International Journal of Molecular Sciences 22, no. 14: 7601. https://doi.org/10.3390/ijms22147601
APA StyleKowalska, M., & Rupik, W. (2021). Architecture of the Pancreatic Islets and Endocrine Cell Arrangement in the Embryonic Pancreas of the Grass Snake (Natrix natrix L.). Immunocytochemical Studies and 3D Reconstructions. International Journal of Molecular Sciences, 22(14), 7601. https://doi.org/10.3390/ijms22147601