
Systematic Understanding of Recent Developments in Bacterial Cellulose Biosynthesis at Genetic, Bioprocess and Product Levels

Abstract
:1. Bacterial Cellulose: What We Know So Far
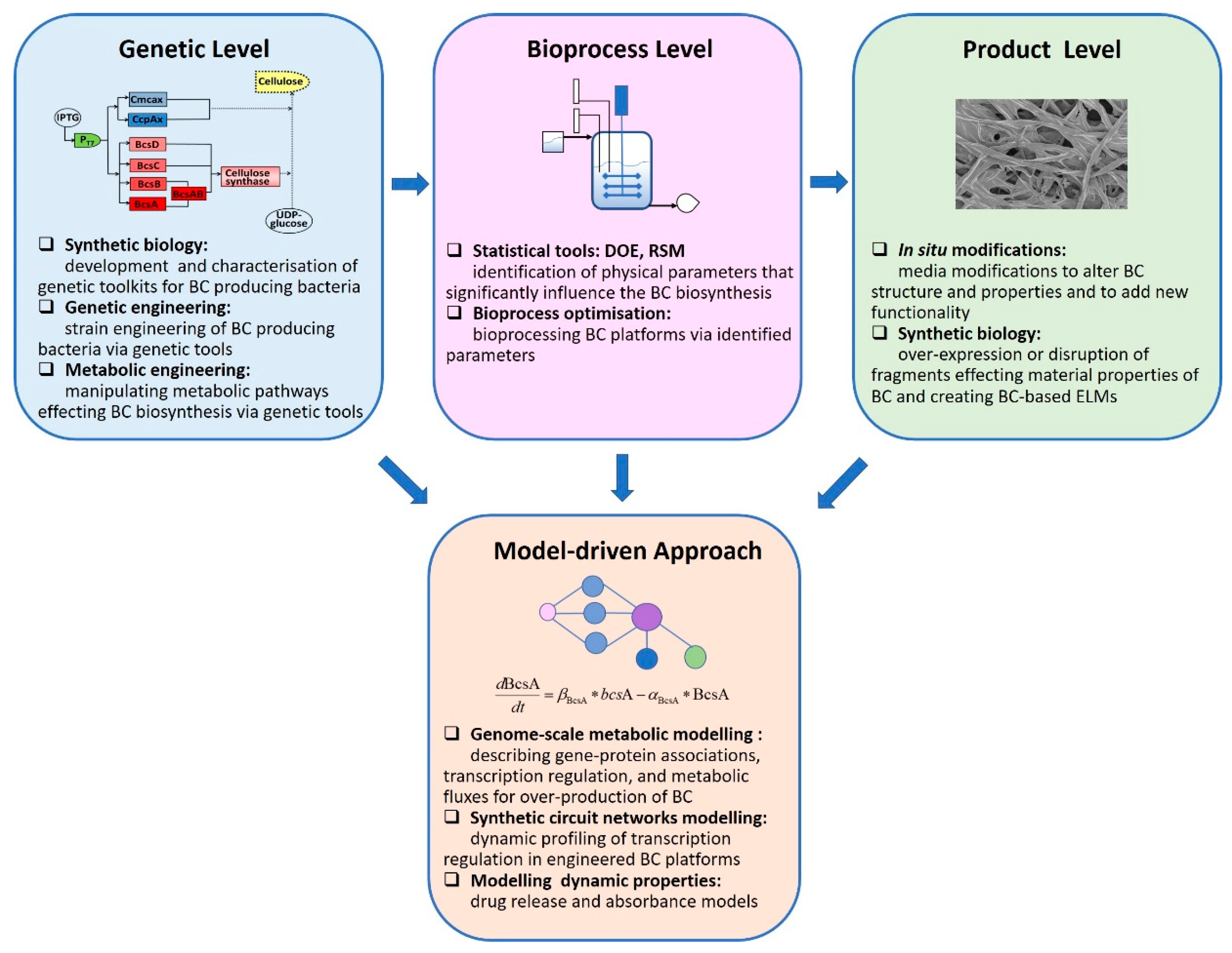
2. Recent Advances
2.1. Genetic Level
2.1.1. Genetic Engineering and Synthetic Biology Approaches
2.1.2. Metabolic Engineering and Synthetic Biology Approaches
2.2. Bioprocess Level
2.3. Product Level
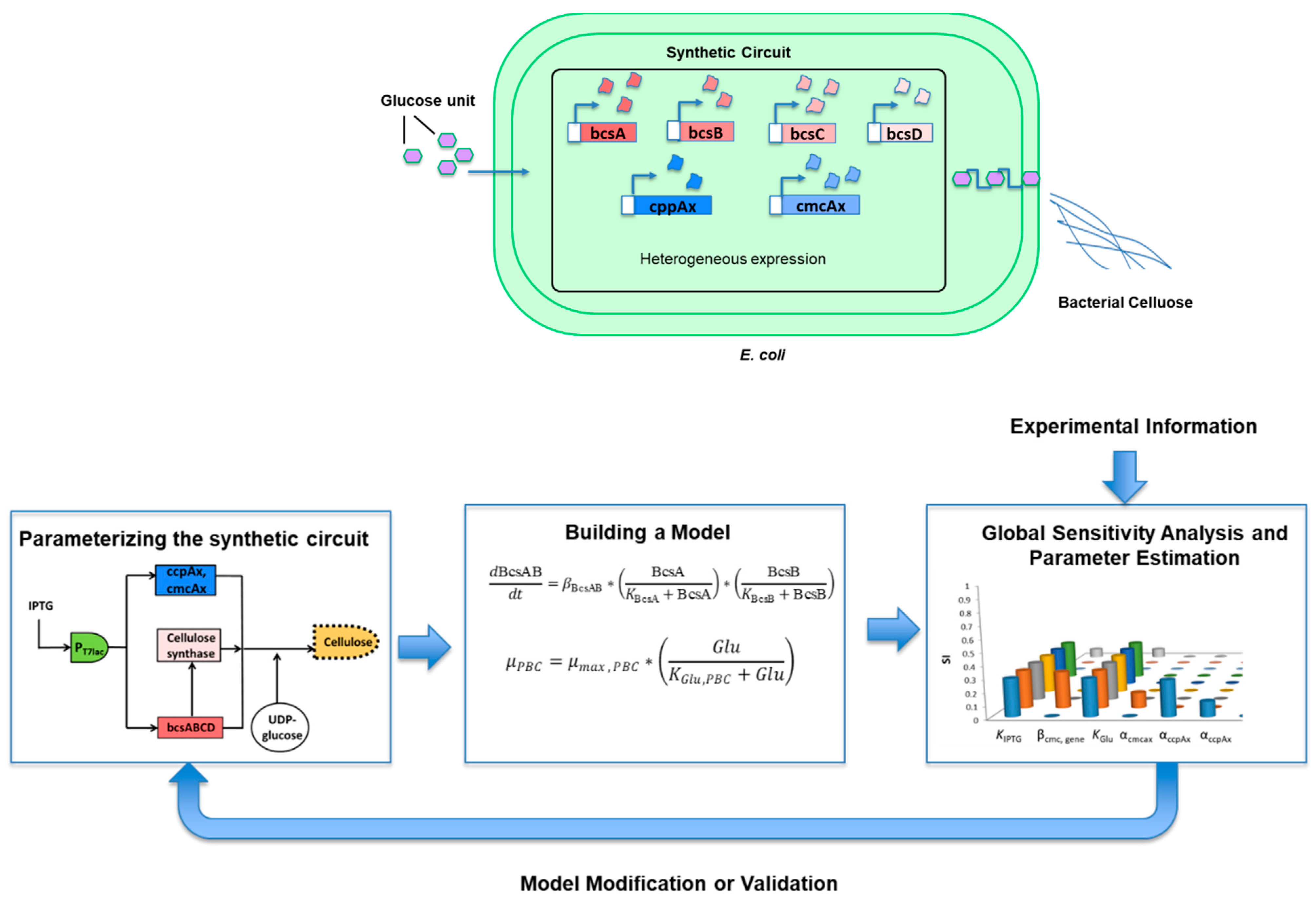
2.4. Model-Driven Approaches
2.4.1. Genome-Scale Metabolic Models
2.4.2. Synthetic Circuit Modeling
2.4.3. Modeling Dynamic Properties
3. Future Perspectives: What Is Next?
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Ryngajłło, M.; Jędrzejczak-Krzepkowska, M.; Kubiak, K.; Ludwicka, K.; Bielecki, S. Towards control of cellulose biosynthesis by Komagataeibacter using systems-level and strain engineering strategies: Current progress and perspectives. Appl. Microbiol. Biotechnol. 2020, 104, 6565–6585. [Google Scholar] [CrossRef]
- Ross, P.; Mayer, R.; Benziman, M. Cellulose biosynthesis and function in bacteria. Microbiol. Rev. 1991, 55, 35–58. [Google Scholar] [CrossRef]
- Lee, K.Y.; Buldum, G.; Mantalaris, A.; Bismarck, A. More than meets the eye in bacterial cellulose: Biosynthesis, bioprocessing, and applications in advanced fiber composites. Macromol. Biosci. 2014, 14, 10–32. [Google Scholar] [CrossRef] [Green Version]
- Andriani, D.; Apriyana, A.Y.; Karina, M. The optimization of bacterial cellulose production and its applications: A review. Cellulose 2020, 27, 6747–6766. [Google Scholar] [CrossRef]
- Zhong, C. Industrial-Scale Production and Applications of Bacterial Cellulose. Front. Bioeng. Biotechnol. 2020, 8, 1425. [Google Scholar] [CrossRef]
- Seddiqi, H.; Oliaei, E.; Honarkar, H.; Jin, J.; Geonzon, L.C.; Bacabac, R.G.; Klein-Nulend, J. Cellulose and its derivatives: Towards biomedical applications. Cellulose 2021, 28, 1893–1931. [Google Scholar] [CrossRef]
- Moniri, M.; Boroumand Moghaddam, A.; Azizi, S.; Abdul Rahim, R.; Bin Ariff, A.; Zuhainis Saad, W.; Navaderi, M.; Mohamad, R. Production and Status of Bacterial Cellulose in Biomedical Engineering. Nanomaterials 2017, 7, 257. [Google Scholar] [CrossRef] [Green Version]
- Campano, C.; Balea, A.; Blanco, A.; Negro, C. Enhancement of the fermentation process and properties of bacterial cellulose: A review. Cellulose 2016, 23, 57–91. [Google Scholar] [CrossRef]
- Dou, C.; Li, Z.; Gong, J.; Li, Q.; Qiao, C.; Zhang, J. Bio-based poly (γ-glutamic acid) hydrogels reinforced with bacterial cellulose nanofibers exhibiting superior mechanical properties and cytocompatibility. Int. J. Biol. Macromol. 2021, 170, 354–365. [Google Scholar] [CrossRef]
- Jian-Yi Wu, F.-M.L. Manufacturing Method of Bio-Fiber Membrane Speaker 2015. Univ Dayeh, Patent No. TW201414326A, 1 April 2014. [Google Scholar]
- Azimi, B.; Milazzo, M.; Danti, S. Cellulose-Based Fibrous Materials From Bacteria to Repair Tympanic Membrane Perforations. Front. Bioeng. Biotechnol. 2021, 9, 474. [Google Scholar] [CrossRef]
- Gautam, S.P.; Bundela, P.S.; Pandey, A.K.; Jamaluddin, J.; Awasthi, M.K.; Sarsaiya, S. A review on systematic study of cellulose. J. Appl. Nat. Sci. 2010, 2, 330–343. [Google Scholar] [CrossRef] [Green Version]
- Saxena, I.M.; Kudlicka, K.; Okuda, K.; Brown, R.M. Characterization of genes in the cellulose-synthesizing operon (acs operon) of acetobacter-xylinum–implications for cellulose crystallization. J. Bacteriol. 1994, 176, 5735–5752. [Google Scholar] [CrossRef] [Green Version]
- Morgan, J.L.W.; Strumillo, J.; Zimmer, J. Crystallographic snapshot of cellulose synthesis and membrane translocation. Nature 2013, 493, 181–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valla, S.; Coucheron, D.H.; Fjaervik, E.; Kjosbakken, J.; Weinhouse, H.; Ross, P.; Amikam, D.; Benziman, M. Cloning of a gene involved in cellulose biosynthesis in acetobacter-xylinum–complementation of cellulose-negative mutants by the udpg pyrophosphorylase structural gene. Mol. Gen. Genet. 1989, 217, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Römling, U.; Galperin, M.Y.; Gomelsky, M. Cyclic di-GMP: The first 25 years of a universal bacterial second messenger. Microbiol. Mol. Biol. Rev. 2013, 77, 1–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujiwara, T.; Komoda, K.; Sakurai, N.; Tajima, K.; Tanaka, I.; Yao, M. The c-di-GMP recognition mechanism of the PilZ domain of bacterial cellulose synthase subunit A. Biochem. Biophys. Res. Commun. 2013, 431, 802–807. [Google Scholar] [CrossRef] [Green Version]
- Iguchi, M.; Yamanaka, S.; Budhiono, A. Bacterial cellulose—A masterpiece of nature’s arts. J. Mater. Sci. 2000, 35, 261–270. [Google Scholar] [CrossRef]
- Jacek, P.; Dourado, F.; Gama, M.; Bielecki, S. Molecular aspects of bacterial nanocellulose biosynthesis. Microb. Biotechnol. 2019, 12, 633–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salgado, L.; Blank, S.; Esfahani, R.A.M.; Strap, J.L.; Bonetta, D. Missense mutations in a transmembrane domain of the Komagataeibacter xylinus BcsA lead to changes in cellulose synthesis. BMC Microbiol. 2019, 19, 216. [Google Scholar] [CrossRef] [Green Version]
- McManus, J.B.; Deng, Y.; Nagachar, N.; Kao, T.-H.; Tien, M. AcsA-AcsB: The core of the cellulose synthase complex from Gluconacetobacter hansenii ATCC23769. Enzyme Microb. Technol. 2016, 82, 58–65. [Google Scholar] [CrossRef] [Green Version]
- Römling, U.; Galperin, M.Y. Bacterial cellulose biosynthesis: Diversity of operons, subunits, products, and functions. Trends Microbiol. 2015, 23, 545–557. [Google Scholar] [CrossRef] [Green Version]
- Mehta, K.; Pfeffer, S.; Brown, R.M. Characterization of an acsD disruption mutant provides additional evidence for the hierarchical cell-directed self-assembly of cellulose in Gluconacetobacter xylinus. Cellulose 2015, 22, 119–137. [Google Scholar] [CrossRef]
- Romling, U. Molecular biology of cellulose production in bacteria. Res. Microbiol. 2002, 153, 205–212. [Google Scholar] [CrossRef]
- Cámara, E.; Lenitz, I.; Nygård, Y. A CRISPR activation and interference toolkit for industrial Saccharomyces cerevisiae strain KE6-12. Sci. Rep. 2020, 10, 14605. [Google Scholar] [CrossRef]
- Rosano, G.L.; Ceccarelli, E.A. Recombinant protein expression in Escherichia coli: Advances and challenges. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonouchi, N.; Tsuchida, T.; Yoshinaga, F.; Horinouchi, S.; Beppu, T. A Host–Vector System for a Cellulose-Producing Acetobacter Strain. Biosci. Biotechnol. Biochem. 1994, 58, 1899–1901. [Google Scholar] [CrossRef] [Green Version]
- Nakai, T.; Tonouchi, N.; Konishi, T.; Kojima, Y.; Tsuchida, T.; Yoshinaga, F.; Sakai, F.; Hayashi, T. Enhancement of cellulose production by expression of sucrose synthase in Acetobacter xylinum. Proc. Natl. Acad. Sci. USA 1999, 96, 14–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonouchi, N.; Horinouchi, S.; Tsuchida, T.; Yoshinaga, F. Increased Cellulose Production from Sucrose by Acetobacter after Introducing the Sucrose Phosphorylase Gene. Biosci. Biotechnol. Biochem. 1998, 62, 1778–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonouchi, N.; Yanase, H.; Kojima, Y.; Tsuchida, T.; Yoshinaga, F.; Horinouchi, S. Increased Cellulose Production from Sucrose with Reduced Levan Accumulation by an Acetobacter Strain Harboring a Recombinant Plasmid. Biosci. Biotechnol. Biochem. 1998, 62, 833–836. [Google Scholar] [CrossRef]
- Battad-Bernardo, E.; McCrindle, S.L.; Couperwhite, I.; Neilan, B.A. Insertion of an E. coli lacZ gene in Acetobacter xylinus for the production of cellulose in whey. FEMS Microbiol. Lett. 2004, 231, 253–260. [Google Scholar] [CrossRef] [Green Version]
- Chien, L.-J.; Chen, H.-T.; Yang, P.-F.; Lee, C.-K. Enhancement of Cellulose Pellicle Production by Constitutively Expressing Vitreoscilla Hemoglobin in Acetobacter xylinum. Biotechnol. Prog. 2006, 22, 1598–1603. [Google Scholar] [CrossRef]
- Setyawati, M.I.; Chien, L.-J.; Lee, C.-K. Self-immobilized recombinant Acetobacter xylinum for biotransformation. Biochem. Eng. J. 2009, 43, 78–84. [Google Scholar] [CrossRef]
- Yadav, V.; Paniliatis, B.J.; Shi, H.; Lee, K.; Cebe, P.; Kaplan, D.L. Novel In Vivo-Degradable Cellulose-Chitin Copolymer from Metabolically Engineered Gluconacetobacter xylinus. Appl. Environ. Microbiol. 2010, 76, 6257–6265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawano, S.; Tajima, K.; Uemori, Y.; Yamashita, H.; Erata, T.; Munekata, M.; Takai, M. Cloning of cellulose synthesis related genes from Acetobacter xylinum ATCC23769 and ATCC53582: Comparison of cellulose synthetic ability between strains. DNA Res. 2002, 9, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Imai, T.; Sun, S.J.; Horikawa, Y.; Wada, M.; Sugiyama, J. Functional reconstitution of cellulose synthase in Escherichia coli. Biomacromolecules 2014, 15, 4206–4213. [Google Scholar] [CrossRef]
- Buldum, G.; Bismarck, A.; Mantalaris, A. Recombinant biosynthesis of bacterial cellulose in genetically modified Escherichia coli. Bioprocess. Biosyst. Eng. 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Florea, M.; Hagemann, H.; Santosa, G.; Abbott, J.; Micklem, C.N.; Spencer-Milnes, X.; de Arroyo Garcia, L.; Paschou, D.; Lazenbatt, C.; Kong, D.; et al. Engineering control of bacterial cellulose production using a genetic toolkit and a new cellulose-producing strain. Proc. Natl. Acad. Sci. USA 2016, 113, E3431–E3440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teh, M.Y.; Ooi, K.H.; Danny Teo, S.X.; Bin Mansoor, M.E.; Shaun Lim, W.Z.; Tan, M.H. An Expanded Synthetic Biology Toolkit for Gene Expression Control in Acetobacteraceae. ACS Synth. Biol. 2019, 8, 708–723. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.-H.; Liu, Q.-J.; Sun, X.-W.; Li, X.-J.; Liu, M.; Jia, S.-R.; Xie, Y.-Y.; Zhong, C. Tailoring bacterial cellulose structure through CRISPR interference-mediated downregulation of galU in Komagataeibacter xylinus CGMCC 2955. Biotechnol. Bioeng. 2020, 117, 2165–2176. [Google Scholar] [CrossRef]
- Kuo, C.-H.; Teng, H.-Y.; Lee, C.-K. Knock-out of glucose dehydrogenase gene in Gluconacetobacter xylinus for bacterial cellulose production enhancement. Biotechnol. Bioprocess. Eng. 2015, 20, 18–25. [Google Scholar] [CrossRef]
- Gwon, H.; Park, K.; Chung, S.-C.; Kim, R.-H.; Kang, J.K.; Ji, S.M.; Kim, N.-J.; Lee, S.; Ku, J.-H.; Do, E.C.; et al. A safe and sustainable bacterial cellulose nanofiber separator for lithium rechargeable batteries. Proc. Natl. Acad. Sci. USA 2019, 116, 19288–19293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Li, S.; Xie, Y.; Jia, S.; Hou, Y.; Zou, Y.; Zhong, C. Enhanced bacterial cellulose production by Gluconacetobacter xylinus via expression of Vitreoscilla hemoglobin and oxygen tension regulation. Appl. Microbiol. Biotechnol. 2018, 102, 1155–1165. [Google Scholar] [CrossRef]
- Liu, D.; Cao, Y.; Qu, R.; Gao, G.; Chen, S.; Zhang, Y.; Wu, M.; Ma, T.; Li, G. Production of bacterial cellulose hydrogels with tailored crystallinity from Enterobacter sp. FY-07 by the controlled expression of colanic acid synthetic genes. Carbohydr. Polym. 2019, 207, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, C.; Tang, T.-C.; Ott, W.; Dorr, B.A.; Shaw, W.M.; Sun, G.L.; Lu, T.K.; Ellis, T. Living materials with programmable functionalities grown from engineered microbial co-cultures. Nat. Mater. 2021, 20, 691–700. [Google Scholar] [CrossRef]
- Jacek, P.; Ryngajłło, M.; Bielecki, S. Structural changes of bacterial nanocellulose pellicles induced by genetic modification of Komagataeibacter hansenii ATCC 23769. Appl. Microbiol. Biotechnol. 2019, 103, 5339–5353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacek, P.; Kubiak, K.; Ryngajłło, M.; Rytczak, P.; Paluch, P.; Bielecki, S. Modification of bacterial nanocellulose properties through mutation of motility related genes in Komagataeibacter hansenii ATCC 53582. New Biotechnol. 2019, 52, 60–68. [Google Scholar] [CrossRef]
- García-Granados, R.; Lerma-Escalera, J.A.; Morones-Ramírez, J.R. Metabolic Engineering and Synthetic Biology: Synergies, Future, and Challenges. Front. Bioeng. Biotechnol. 2019, 7, 36. [Google Scholar] [CrossRef]
- Khan, T.; Khan, S.; Park, J.K. Simple fed-batch cultivation strategy for the enhanced production of a single-sugar glucuronic acid-based oligosaccharides by a cellulose-producing Gluconacetobacter hansenii strain. Biotechnol. Bioprocess. Eng. 2008, 13, 240–247. [Google Scholar] [CrossRef]
- Li, Y.; Tian, C.; Tian, H.; Zhang, J.; He, X.; Ping, W.; Lei, H. Improvement of bacterial cellulose production by manipulating the metabolic pathways in which ethanol and sodium citrate involved. Appl. Microbiol. Biotechnol. 2012, 96, 1479–1487. [Google Scholar] [CrossRef]
- Shigematsu, T.; Takamine, K.; Kitazato, M.; Morita, T.; Naritomi, T.; Morimura, S.; Kida, K. Cellulose production from glucose using a glucose dehydrogenase gene (gdh)-deficient mutant of Gluconacetobacter xylinus and its use for bioconversion of sweet potato pulp. J. Biosci. Bioeng. 2005, 99, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Cerrutti, P.; Roldán, P.; García, R.M.; Galvagno, M.A.; Vázquez, A.; Foresti, M.L. Production of bacterial nanocellulose from wine industry residues: Importance of fermentation time on pellicle characteristics. J. Appl. Polym. Sci. 2016, 133. [Google Scholar] [CrossRef]
- Du, R.; Zhao, F.; Peng, Q.; Zhou, Z.; Han, Y. Production and characterization of bacterial cellulose produced by Gluconacetobacter xylinus isolated from Chinese persimmon vinegar. Carbohydr. Polym. 2018, 194, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Abdelraof, M.; Hasanin, M.S.; El -Saied, H. Ecofriendly green conversion of potato peel wastes to high productivity bacterial cellulose. Carbohydr. Polym. 2019, 211, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Bilgi, E.; Bayır, E.; Sendemir, A.; Hameş Tuna, E.E. Optimization of Bacterial Cellulose Production by Gluconacetobacter xylinus using Carob and Haricot Bean. Int. J. Biol. Macromol. 2016, 90. [Google Scholar] [CrossRef]
- Basu, A.; Vadanan, S.V.; Lim, S. Rational design of a scalable bioprocess platform for bacterial cellulose production. Carbohydr. Polym. 2019, 207, 684–693. [Google Scholar] [CrossRef]
- Qiu, Z.; Wang, M.; Zhang, T.; Yang, D.; Qiu, F. In-situ fabrication of dynamic and recyclable TiO2 coated bacterial cellulose membranes as an efficient hybrid absorbent for tellurium extraction. Cellulose 2020, 27, 4591–4608. [Google Scholar] [CrossRef]
- Orlando, I.; Basnett, P.; Nigmatullin, R.; Wang, W.; Knowles, J.C.; Roy, I. Chemical Modification of Bacterial Cellulose for the Development of an Antibacterial Wound Dressing. Front. Bioeng. Biotechnol. 2020, 8, 557885. [Google Scholar] [CrossRef]
- Liu, K.; Catchmark, J.M. Enhanced mechanical properties of bacterial cellulose nanocomposites produced by co-culturing Gluconacetobacter hansenii and Escherichia coli under static conditions. Carbohydr. Polym. 2019, 219, 12–20. [Google Scholar] [CrossRef]
- Molina-Ramírez, C.; Castro, M.; Osorio, M.; Torres-Taborda, M.; Gómez, B.; Zuluaga, R.; Gómez, C.; Gañán, P.; Rojas, O.J.; Castro, C. Effect of Different Carbon Sources on Bacterial Nanocellulose Production and Structure Using the Low pH Resistant Strain Komagataeibacter Medellinensis. Materails 2017, 10, 639. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, C.; Ellis, T. Biological Engineered Living Materials: Growing Functional Materials with Genetically Programmable Properties. ACS Synth. Biol. 2019, 8, 1–15. [Google Scholar] [CrossRef]
- Sruba, W.V., III. Engineered Living Materials: Taxonomies and Emerging Trends. Trends Biotechnol. 2021, 39, 574–583. [Google Scholar] [CrossRef]
- Liu, S.; Xu, W. Engineered Living Materials-Based Sensing and Actuation. Front. Sens. 2020, 1, 6. [Google Scholar] [CrossRef]
- Gao, M.; Li, J.; Bao, Z.; Hu, M.; Nian, R.; Feng, D.; An, D.; Li, X.; Xian, M.; Zhang, H. A natural in situ fabrication method of functional bacterial cellulose using a microorganism. Nat. Commun. 2019, 10, 437. [Google Scholar] [CrossRef] [Green Version]
- Castro, C.; Cordeiro, N.; Faria, M.; Zuluaga, R.; Putaux, J.-L.; Filpponen, I.; Velez, L.; Rojas, O.J.; Gañán, P. In-situ glyoxalization during biosynthesis of bacterial cellulose. Carbohydr. Polym. 2015, 126, 32–39. [Google Scholar] [CrossRef]
- Yin, N.; Chen, S.; Cao, Y.; Wang, H.; Wu, Q. Improvement in mechanical properties and biocompatibility of biosynthetic bacterial cellulose/lotus root starch composites. Chin. J. Polym. Sci. 2017, 35, 354–364. [Google Scholar] [CrossRef]
- de Lima Fontes, M.; Meneguin, A.B.; Tercjak, A.; Gutierrez, J.; Cury, B.S.F.; dos Santos, A.M.; Ribeiro, S.J.L.; Barud, H.S. Effect of in situ modification of bacterial cellulose with carboxymethylcellulose on its nano/microstructure and methotrexate release properties. Carbohydr. Polym. 2018, 179, 126–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hornung, M.; Biener, R.; Schmauder, H.-P. Dynamic modelling of bacterial cellulose formation. Eng. Life Sci. 2009, 9, 342–347. [Google Scholar] [CrossRef]
- Ismail, F.; Dissanayake, D. Mathematical Modeling of Bacterial Cellulose Production by Acetobacter xylinum Using Rotating Biological Fermentor. In Proceedings of the 27th European Conference on Modelling and Simulation, Alesund, Norway, 27–30 May 2013. [Google Scholar]
- Mendoza, S.N.; Olivier, B.G.; Molenaar, D.; Teusink, B. A systematic assessment of current genome-scale metabolic reconstruction tools. Genome Biol. 2019, 20, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, C.; Kim, G.B.; Kim, W.J.; Kim, H.U.; Lee, S.Y. Current status and applications of genome-scale metabolic models. Genome Biol. 2019, 20, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Ye, C.; Xu, N.; Chen, C.; Chen, X.; Yuan, F.; Xu, Y.; Yang, J.; Sun, D. Reconstruction of a Genome-scale Metabolic Network of Komagataeibacter nataicola RZS01 for Cellulose Production. Sci. Rep. 2017, 7, 7911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Souza, S.S.; de Castro, J.V.; Porto, L.M. Modeling the core metabolism of komagataeibacter hansenii atcc 23769 to evaluate nanocellulose biosynthesis. Braz. J. Chem. Eng. 2018, 35, 869–886. [Google Scholar] [CrossRef]
- Rezazadeh, M.; Babaeipour, V.; Motamedian, E. Reconstruction, verification and in-silico analysis of a genome-scale metabolic model of bacterial cellulose producing Komagataeibacter xylinus. Bioprocess. Biosyst. Eng. 2020, 43, 1017–1026. [Google Scholar] [CrossRef]
- Jang, W.D.; Kim, T.Y.; Kim, H.U.; Shim, W.Y.; Ryu, J.Y.; Park, J.H.; Lee, S.Y. Genomic and metabolic analysis of Komagataeibacter xylinus DSM 2325 producing bacterial cellulose nanofiber. Biotechnol. Bioeng. 2019, 116, 3372–3381. [Google Scholar] [CrossRef] [PubMed]
- Tsipa, A.; Koutinas, M.; Usaku, C.; Mantalaris, A. Optimal bioprocess design through a gene regulatory network–Growth kinetic hybrid model: Towards replacing Monod kinetics. Metab. Eng. 2018, 48, 129–137. [Google Scholar] [CrossRef]
- Buldum, G.; Tsipa, A.; Mantalaris, A. Linking Engineered Gene Circuit Kinetic Modeling to Cellulose Biosynthesis Prediction in Escherichia coli: Toward Bioprocessing of Microbial Cell Factories. Ind. Eng. Chem. Res. 2020, 59, 4659–4669. [Google Scholar] [CrossRef] [Green Version]
- Juncu, G.; Stoica-Guzun, A.; Stroescu, M.; Isopencu, G.; Jinga, S.I. Drug release kinetics from carboxymethylcellulose-bacterial cellulose composite films. Int. J. Pharm. 2016, 510, 485–492. [Google Scholar] [CrossRef]
- George, J.; Kumar, R.; Sajeevkumar, V.A.; Ramana, K.V.; Rajamanickam, R.; Abhishek, V.; Nadanasabapathy, S. Siddaramaiah Hybrid HPMC nanocomposites containing bacterial cellulose nanocrystals and silver nanoparticles. Carbohydr. Polym. 2014, 105, 285–292. [Google Scholar] [CrossRef]
- Ye, S.; Jiang, L.; Su, C.; Zhu, Z.; Wen, Y.; Shao, W. Development of gelatin/bacterial cellulose composite sponges as potential natural wound dressings. Int. J. Biol. Macromol. 2019, 133, 148–155. [Google Scholar] [CrossRef]
- Adepu, S.; Khandelwal, M. Ex-situ modification of bacterial cellulose for immediate and sustained drug release with insights into release mechanism. Carbohydr. Polym. 2020, 249, 116816. [Google Scholar] [CrossRef] [PubMed]
- Ciecholewska-Juśko, D.; Żywicka, A.; Junka, A.; Drozd, R.; Sobolewski, P.; Migdał, P.; Kowalska, U.; Toporkiewicz, M.; Fijałkowski, K. Superabsorbent crosslinked bacterial cellulose biomaterials for chronic wound dressings. Carbohydr. Polym. 2021, 253, 117247. [Google Scholar] [CrossRef] [PubMed]
- Cacicedo, M.L.; Pacheco, G.; Islan, G.A.; Alvarez, V.A.; Barud, H.S.; Castro, G.R. Chitosan-bacterial cellulose patch of ciprofloxacin for wound dressing: Preparation and characterization studies. Int. J. Biol. Macromol. 2020, 147, 1136–1145. [Google Scholar] [CrossRef]
- Bernardelli de Mattos, I.; Nischwitz, S.P.; Tuca, A.-C.; Groeber-Becker, F.; Funk, M.; Birngruber, T.; Mautner, S.I.; Kamolz, L.-P.; Holzer, J.C.J. Delivery of antiseptic solutions by a bacterial cellulose wound dressing: Uptake, release and antibacterial efficacy of octenidine and povidone-iodine. Burns 2020, 46, 918–927. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Briffa, S.M.; Swingler, S.; Gibson, H.; Kannappan, V.; Adamus, G.; Kowalczuk, M.; Martin, C.; Radecka, I. Synthesis of Silver Nanoparticles Using Curcumin-Cyclodextrins Loaded into Bacterial Cellulose-Based Hydrogels for Wound Dressing Applications. Biomacromolecules 2020. [Google Scholar] [CrossRef]
- Shao, W.; Liu, H.; Wang, S.; Wu, J.; Huang, M.; Min, H.; Liu, X. Controlled release and antibacterial activity of tetracycline hydrochloride-loaded bacterial cellulose composite membranes. Carbohydr. Polym. 2016, 145, 114–120. [Google Scholar] [CrossRef]
- Beekmann, U.; Schmölz, L.; Lorkowski, S.; Werz, O.; Thamm, J.; Fischer, D.; Kralisch, D. Process control and scale-up of modified bacterial cellulose production for tailor-made anti-inflammatory drug delivery systems. Carbohydr. Polym. 2020, 236, 116062. [Google Scholar] [CrossRef] [PubMed]
- Lopez, K.M.; Ravula, S.; Pérez, R.L.; Ayala, C.E.; Losso, J.N.; Janes, M.E.; Warner, I.M. Hyaluronic Acid–Cellulose Composites as Patches for Minimizing Bacterial Infections. ACS Omega 2020, 5, 4125–4132. [Google Scholar] [CrossRef] [Green Version]
- Solomevich, S.O.; Dmitruk, E.I.; Bychkovsky, P.M.; Nebytov, A.E.; Yurkshtovich, T.L.; Golub, N. V Fabrication of oxidized bacterial cellulose by nitrogen dioxide in chloroform/cyclohexane as a highly loaded drug carrier for sustained release of cisplatin. Carbohydr. Polym. 2020, 248, 116745. [Google Scholar] [CrossRef] [PubMed]
- Meneguin, A.B.; da Silva Barud, H.; Sábio, R.M.; de Sousa, P.Z.; Manieri, K.F.; de Freitas, L.A.P.; Pacheco, G.; Alonso, J.D.; Chorilli, M. Spray-dried bacterial cellulose nanofibers: A new generation of pharmaceutical excipient intended for intestinal drug delivery. Carbohydr. Polym. 2020, 249, 116838. [Google Scholar] [CrossRef]
- Coelho, F.; Cavicchioli, M.; Specian, S.S.; Scarel-Caminaga, R.M.; de Penteado, L.A.; de Medeiros, A.I.; de Ribeiro, S.J.L.; de Capote, T.S.O. Bacterial cellulose membrane functionalized with hydroxiapatite and anti-bone morphogenetic protein 2: A promising material for bone regeneration. PLoS ONE 2019, 14, e0221286. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wang, X.; Li, X.; Zhang, L.; Jiang, F. A 3D porous microsphere with multistage structure and component based on bacterial cellulose and collagen for bone tissue engineering. Carbohydr. Polym. 2020, 236, 116043. [Google Scholar] [CrossRef] [PubMed]
- Lamboni, L.; Xu, C.; Clasohm, J.; Yang, J.; Saumer, M.; Schäfer, K.-H.; Yang, G. Silk sericin-enhanced microstructured bacterial cellulose as tissue engineering scaffold towards prospective gut repair. Mater. Sci. Eng. C 2019, 102, 502–510. [Google Scholar] [CrossRef] [PubMed]
- Sajjad, W.; Khan, T.; Ul-Islam, M.; Khan, R.; Hussain, Z.; Khalid, A.; Wahid, F. Development of modified montmorillonite-bacterial cellulose nanocomposites as a novel substitute for burn skin and tissue regeneration. Carbohydr. Polym. 2019, 206, 548–556. [Google Scholar] [CrossRef] [PubMed]
- Arias, S.L.; Shetty, A.; Devorkin, J.; Allain, J.-P. Magnetic targeting of smooth muscle cells in vitro using a magnetic bacterial cellulose to improve cell retention in tissue-engineering vascular grafts. Acta Biomater. 2018, 77, 172–181. [Google Scholar] [CrossRef]
- Li, Y.; Jiang, K.; Feng, J.; Liu, J.; Huang, R.; Chen, Z.; Yang, J.; Dai, Z.; Chen, Y.; Wang, N.; et al. Construction of Small-Diameter Vascular Graft by Shape-Memory and Self-Rolling Bacterial Cellulose Membrane. Adv. Healthc. Mater. 2017, 6, 1601343. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Strain | Aim | Approach | Outcome | Reference |
|---|---|---|---|---|
| K. sucrofermentans BPR 2001 | sucrose synthase expression to enable sucrose metabolism | overexpression by pSA-SD derived from pSA19 | 2 times increased BC yield (8 g/L) | [28] |
| Acetobacter ITDI 2.1 | β-galactosidase expression to enable lactose metabolism | lacZ genome integration | 28-fold increased BC yield ability to use lactose and whey as carbon source | [31] |
| A. xylinum BCRC12334 | Vitreoscilla hemoglobin (VHb) expression to promote O2 utilisation | constitutive expression of VHb by pBla-VHb-122 derived from pBBR122 | 2-fold increased BC production 50% increased growth rate | [32] |
| A. xylinum BCRC12334 | d-Amino acid oxidase (DAAO) expression and immobilization | inducible expression of DAAO by pLacDAAO-122 | self-immobilization of DAAO+ cells (10% activity) | [33] |
| G. xylinus 10245 | to incorporate N-acetylglucosamine (GlcNAc) sugar residues into glucan chains | overexpression of AGM1, NAG5 and UAP1 via pBBR-GlcNAc | cellulose-chitin copolymer synthesis | [34] |
| E.coli XL1-Blue | BC biosynthesis in E. coli | heterogeneous co-expression of BcsA, BcsB and DGC (diguanyl cyclase) | reconstitution of cellulose synthase no BC crystallization | [36] |
| E.coli C41 (DE3) E.coli HMS174(DE3) | BC biosynthesis in E. coli | heterogeneous expression of bcsABCD operon and upstream operon (cmcax, ccpAx) via inducible pCMP and pBCS | large fibres with diameters ranging from 10 to 20 μm rapid BC production and short culturing period | [37] |
| K. rhaeticus iGEM | building genetic toolkit for Acetobacteraceae | identification of plasmid backbones, characterisation, and engineering of constitutive and inducible promoters | toolkit achieved biosynthesis of patterned cellulose, functionalization of the cellulose surface with proteins, and tunable control over cellulose production | [38] |
| G. xylinus ATCC 700178 G. hansenii ATCC 53582 K. rhaeticus iGEM | building an expanded genetic toolkit for Acetobacteraceae | characterisation multiple natural and synthetic promoters, ribosome binding sites, terminators, and degradation tags by expressing RFP1 reporter gene CRISPRi targeting endogenous acs operon (acsAB and acsD) | expanded toolkit readily mix-and match for expression modified cellulose with variable chitin content via high or low expression plasmids | [39] |
| K. xylinus CMCC 2955 | structural characterisation of BC under various galU expression | CRISPRi to downregulate galU | porosity increased by 0.5-fold with galU repression crystallinity increased with the rise in galU expression | [40] |
| G. xylinus BCRC12334 | reducing gluconic acid production by eliminating the membrane-bound glucose dehydrogenase (GDH) activity | GDH knock-out | 40 and 230% increased BC production in static and shaken culture | [41] |
| K. xylinus DSM 2325 | identifying the effect of pfkA gene in glucose metabolism | pfkA genome integration CRP overexpression by pIN01-crp | increased cellulose production (from 3.5 to 4.5 g/L) sharp decrease in the yield of gluconic acid (from 64.8 to 39.2%) | [42] |
| K. xylinus CGMCC 2955 | exploring the effect of oxygen tension on BC production | constitutive expression of VHb by pBla-VHb-122 derived from pBBR122 | increased BC yield 26.5 and 58.6% at oxygen tensions of 10 and 15% | [43] |
| Enterobacter sp. FY-07 | production of colanic acid to improve water holding capacity of BC | overexpression of wca operon (encoding colanic acid) via inducible pTSK1-tac | water holding capacity enhanced slightly by 1.7-fold | [44] |
| K. rhaeticus and S. cerevisiae co-cultures | developing ELM system programmed for dedicated tasks | co-culturing of K. rhaeticus with engineered strains of S. cerevisiae to secrete enzymes into BC or creating living materials that can sense and respond to environmental stimuli | enzyme-functionalized BC, altered physical properties and produced BC-based ELMs that can sense and respond to chemical and optical stimuli | [45] |
| K. hansenii ATCC 23769 | investigating the effect of motility genes (motA and motB) on BC structure | overexpression of MotA and MotB disruption of motA and motB | substantial loosening of intra-membrane structure overexpression of motility proteins, compact BC structure achieved via disruption of these genes | [46,47] |
| Application Area | Properties of BC | Reference |
|---|---|---|
| Wound dressing | non-toxic, non-carcinogenic and biocompatible, capacity to retain moisture, allows for oxygen exchange | [82,83,84,85] |
| Drug delivery | nanofibrillar structure represent a suitable macromolecular support for inclusion of drugs and therefore modulation of the drug release | [86,87,88,89,90] |
| Tissue regeneration/ scaffolds | allows cellular adhesion and proliferation, customizable to control its features | [91,92,93,94] |
| Vascular grafts | represents high mechanical strength and microporosity | [95,96] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buldum, G.; Mantalaris, A. Systematic Understanding of Recent Developments in Bacterial Cellulose Biosynthesis at Genetic, Bioprocess and Product Levels. Int. J. Mol. Sci. 2021, 22, 7192. https://doi.org/10.3390/ijms22137192
Buldum G, Mantalaris A. Systematic Understanding of Recent Developments in Bacterial Cellulose Biosynthesis at Genetic, Bioprocess and Product Levels. International Journal of Molecular Sciences. 2021; 22(13):7192. https://doi.org/10.3390/ijms22137192
Chicago/Turabian StyleBuldum, Gizem, and Athanasios Mantalaris. 2021. "Systematic Understanding of Recent Developments in Bacterial Cellulose Biosynthesis at Genetic, Bioprocess and Product Levels" International Journal of Molecular Sciences 22, no. 13: 7192. https://doi.org/10.3390/ijms22137192
APA StyleBuldum, G., & Mantalaris, A. (2021). Systematic Understanding of Recent Developments in Bacterial Cellulose Biosynthesis at Genetic, Bioprocess and Product Levels. International Journal of Molecular Sciences, 22(13), 7192. https://doi.org/10.3390/ijms22137192