
Istradefylline Mitigates Age-Related Hearing Loss in C57BL/6J Mice

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
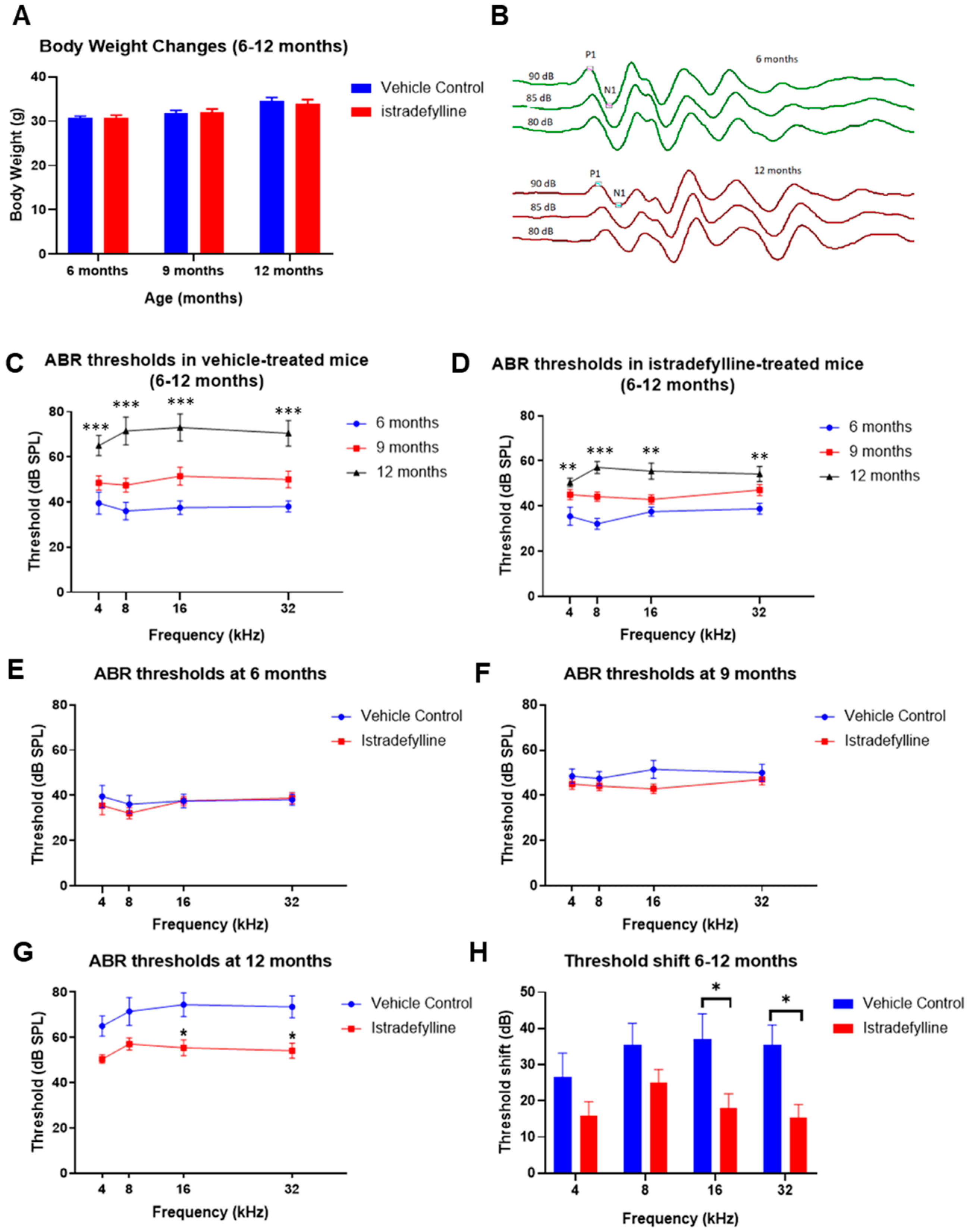
2.1. Body Weights and Auditory Brainstem Responses in Mice Treated with Istradefylline
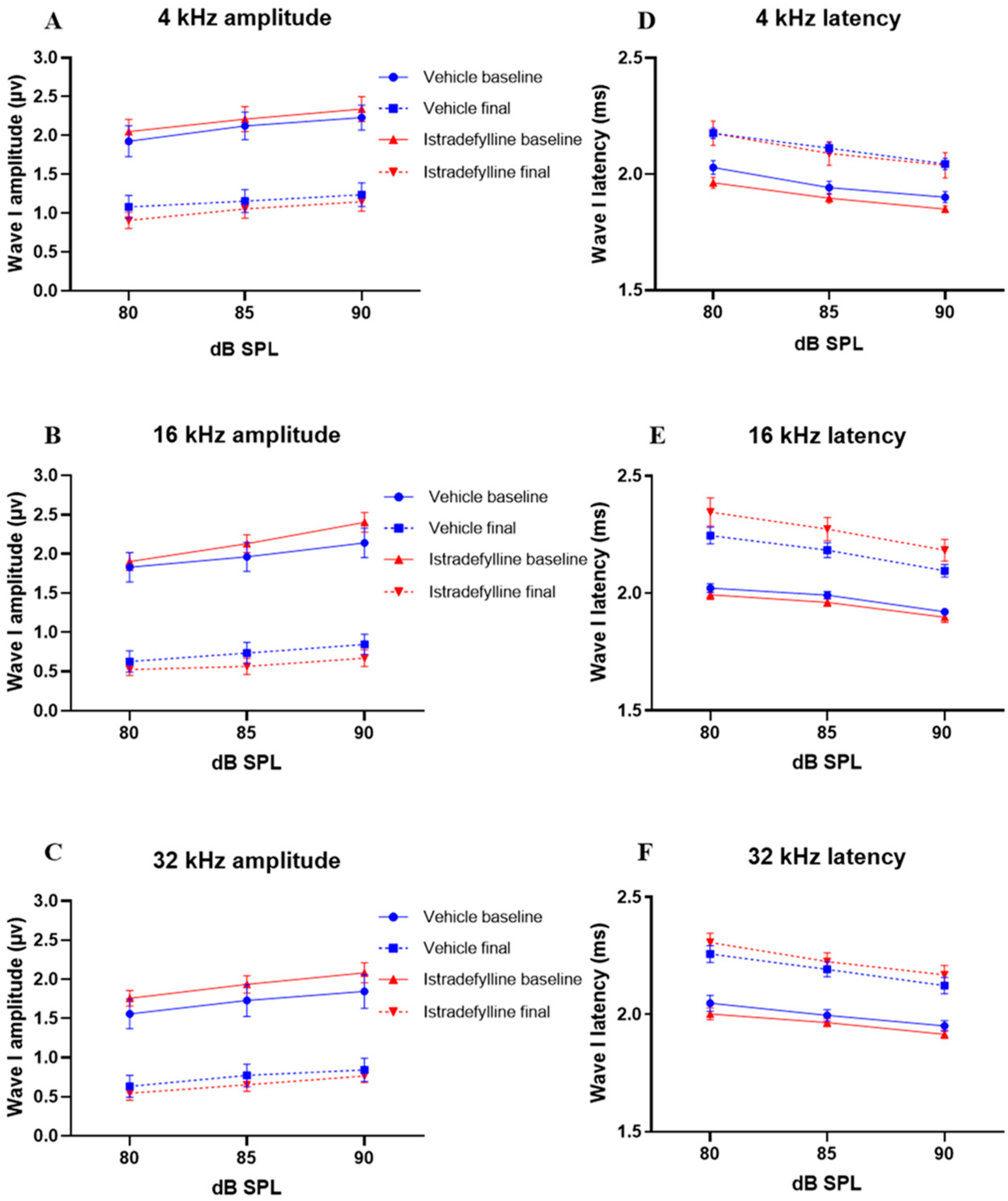
2.2. Input–Output Functions
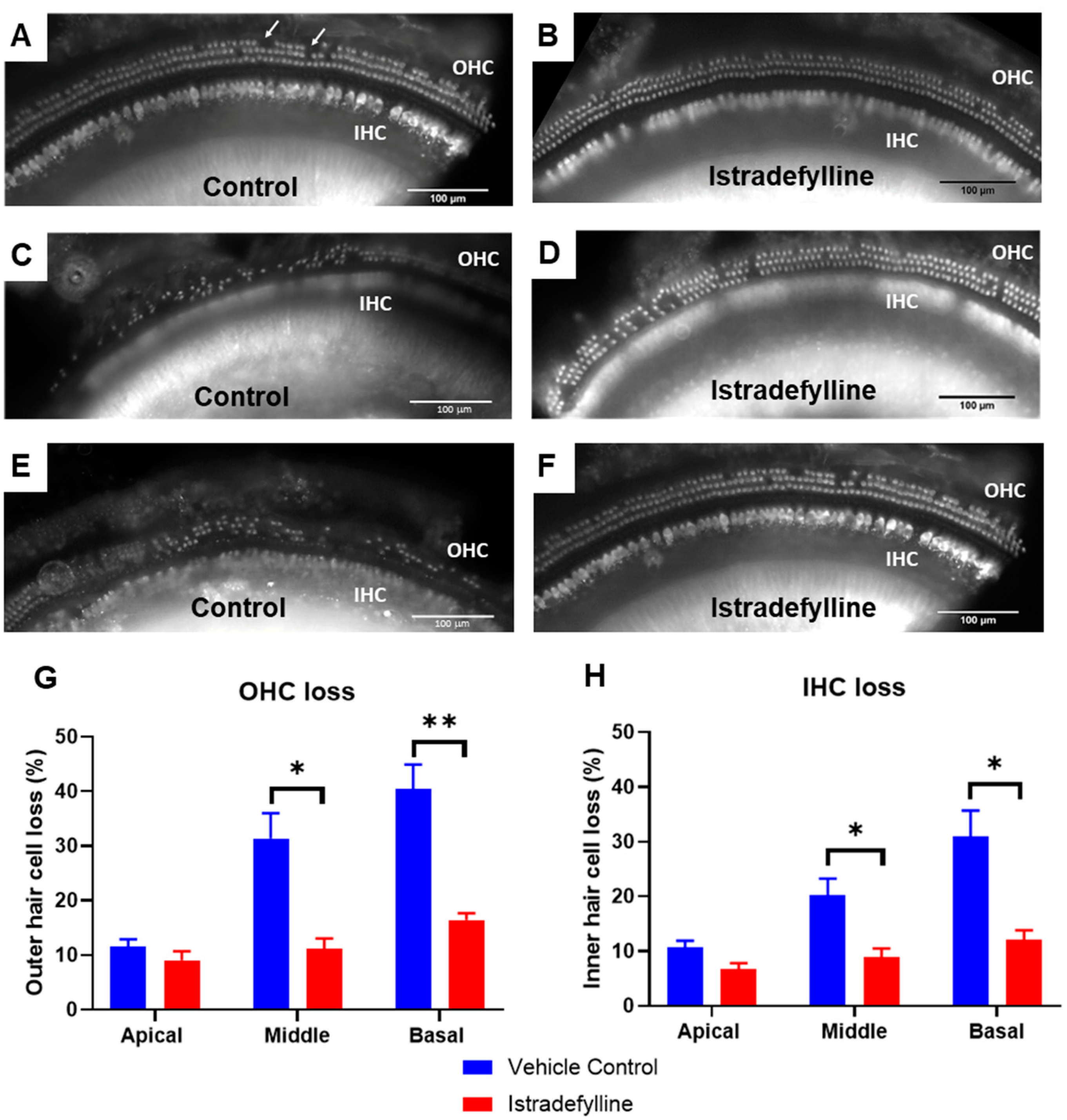
2.3. Hair Cell Survival
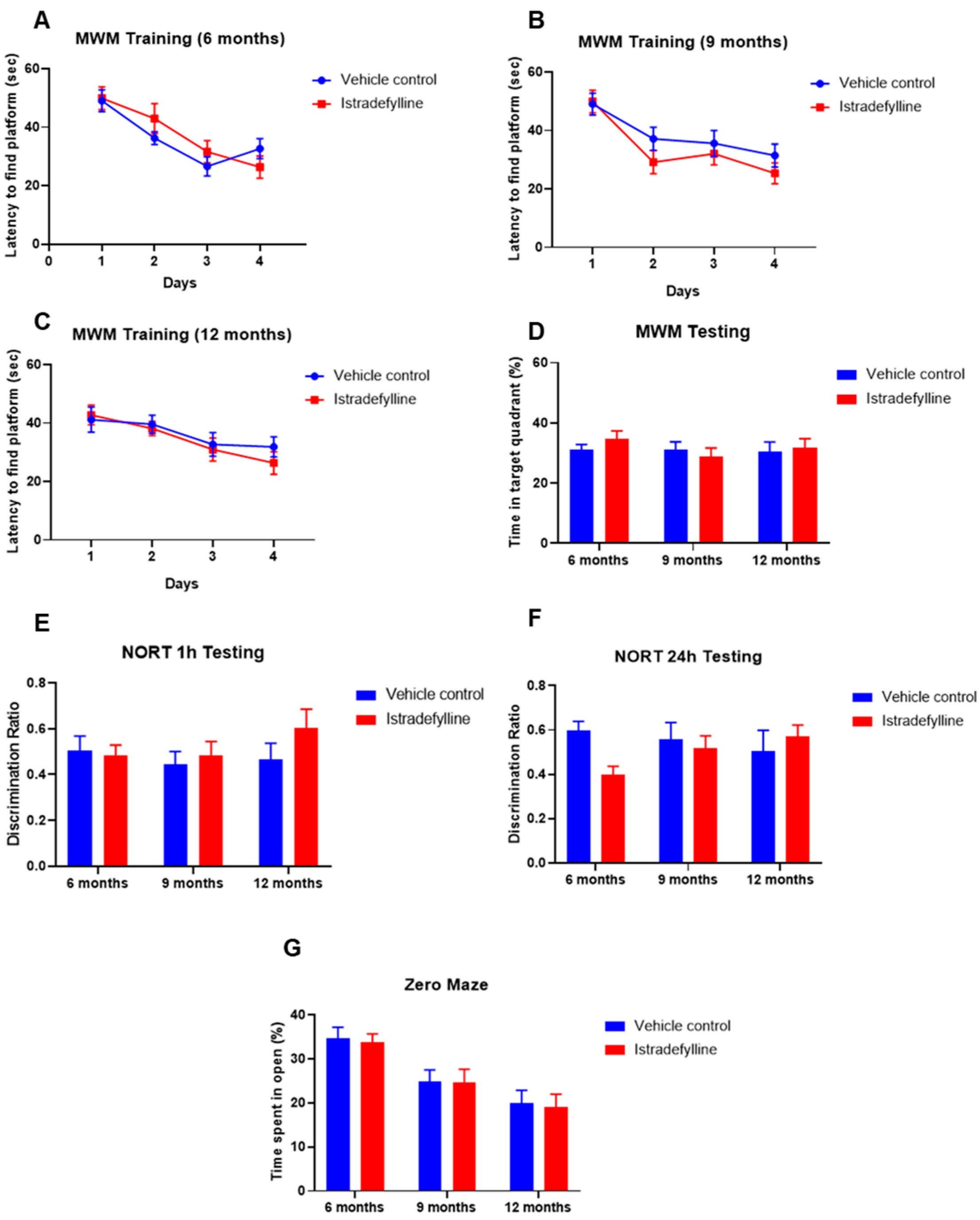
2.4. Behavioural Studies
3. Discussion
3.1. ABR Threshold Shifts and Hair Cell Loss in Istradefylline-Treated Mice
3.2. ABR Suprathreshold Responses
3.3. Cognitive Studies
3.4. Putative Mechanisms of Otoprotection by Istradefylline
4. Materials and Methods
4.1. Animals
4.2. Auditory Brainstem Responses
4.3. Istradefylline Treatment
4.4. Cochlear Tissue Preparation
4.5. Hair Cell Counting
4.6. Morris Water Maze
4.7. Novel Object Recognition Test
4.8. Zero Maze Test
4.9. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. World Report on Hearing. 2021. Available online: https://www.who.int/publications/i/item/world-report-on-hearing (accessed on 3 March 2021).
- Davis, A.; McMahon, C.M.; Pichora-Fuller, K.M.; Russ, S.; Lin, F.; Olusanya, B.O.; Chadha, S.; Tremblay, K.L. Aging and hearing health: The life-course approach. Gerontologist 2016, 56, S256–S267. [Google Scholar] [CrossRef]
- Yamasoba, T.; Lin, F.R.; Someya, S.; Kashio, A.; Sakamoto, T.; Kondo, K. Current concepts in age related hearing loss: Epidemiology and mechanistic pathways. Hear. Res. 2013, 303, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Jayakody, D.M.; Friedland, P.L.; Martins, R.N.; Sohrabi, H.R. Impact of aging on the auditory system and related cognitive functions: A narrative review. Front. Neurosci. 2018, 12, 125. [Google Scholar] [CrossRef]
- Livingston, G.; Sommerlad, A.; Orgeta, V.; Costafreda, S.G.; Huntley, J.; Ames, D.; Ballard, C.; Banerjee, S.; Burns, A.; Mukadam, N.; et al. Dementia prevention, intervention, and care. Lancet 2017, 390, 2673–2734. [Google Scholar] [CrossRef] [Green Version]
- Schuknecht, H.F.; Gacek, M.R. Cochlear pathology in presbyacusis. Ann. Otol. Rhinol. Laryngol. 1993, 102, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Gates, G.A.; Mills, J.H. Presbycusis. Lancet 2005, 366, 1111–1120. [Google Scholar] [CrossRef]
- Yamasoba, T.; Someya, S.; Yamada, C.; Weindruch, R.; Prolla, T.A.; Tanokura, M. Role of mitochondrial dysfunction and mitochondrial DNA mutations in age-related hearing loss. Hear. Res. 2007, 226, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Puel, J.L. Toward cochlear therapies. Physiol. Rev. 2018, 98, 2477–2522. [Google Scholar] [CrossRef] [PubMed]
- Watson, N.; Ding, B.; Zhu, X.; Frisina, R.D. Chronic inflammation—inflammaging—in the ageing cochlea: A novel target for future presbycusis therapy. Ageing Res. Rev. 2017, 40, 142–148. [Google Scholar] [CrossRef]
- Zheng, Q.Y.; Johnson, K.R.; Erway, L.C. Assessment of hearing in 80 inbred strains of mice by ABR threshold analyses. Hear. Res. 1999, 130, 94–107. [Google Scholar] [CrossRef] [Green Version]
- Ohlemiller, K.K. Contributions of mouse models to understanding of age- and noise-related hearing loss. Brain Res. 2006, 1091, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Prosen, C.A.; Dore, D.J.; May, B.J. The functional age of hearing loss in a mouse model of presbyacusis. I. Behavioral assessments. Hear. Res. 2003, 183, 44–56. [Google Scholar] [CrossRef]
- Shoji, H.; Takao, K.; Hattori, S.; Miyakawa, T. Age-related changes in behaviour in C57BL/6J mice from young adulthood to middle age. Mol. Brain 2016, 9, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, A.C.Y.; Guo, C.X.; Gupta, R.; Housley, G.D.; Thorne, P.R.; Vlajkovic, S.M. Post exposure administration of A1 adenosine receptor agonists attenuates noise-induced hearing loss. Hear. Res. 2010, 260, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Vlajkovic, S.M.; Lee, K.H.; Wong, A.C.Y.; Guo, C.X.; Gupta, R.; Housley, G.D.; Thorne, P.R. Adenosine amine congener mitigates noise-induced cochlear injury. Purinergic Signal. 2010, 6, 273–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunewardene, N.; Guo, C.X.; Wong, A.C.Y.; Thorne, P.R.; Vlajkovic, S.M. Adenosine Amine Congener ameliorates cisplatin-induced hearing loss. World J. Otorhinolaryngol. 2013, 3, 100–107. [Google Scholar] [CrossRef]
- Lin, S.C.Y.; Thorne, P.R.; Housley, G.D.; Vlajkovic, S.M. Purinergic signalling and aminoglycoside ototoxicity: The opposing roles of P1 (adenosine) and P2 (ATP) receptors on cochlear hair cell survival. Front. Cell. Neurosci. 2019, 13, 207. [Google Scholar] [CrossRef] [Green Version]
- Vlajkovic, S.M.; Housley, G.D.; Thorne, P.R. Adenosine and the auditory system. Curr. Neuropharmacol. 2009, 7, 246–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.F.; Eltzschig, H.K.; Fredholm, B.B. Adenosine receptors as drug targets—What are the challenges? Nat. Rev. 2013, 12, 265–286. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.F. Adenosine receptor control of cognition in normal and disease. Int. Rev. Neurobiol. 2014, 119, 257–307. [Google Scholar]
- Woods, L.T.; Ajit, D.; Camden, J.M.; Erb, L.; Weisman, G.A. Purinergic receptors as potential therapeutic targets in Alzheimer’s disease. Neuropharmacology 2016, 104, 169–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlajkovic, S.M.; Ambepitiya, K.; Barclay, M.; Boison, D.; Housley, G.D.; Thorne, P.R. Adenosine receptors regulate susceptibility to noise-induced neural injury in the mouse cochlea and hearing loss. Hear. Res. 2017, 345, 43–51. [Google Scholar] [CrossRef]
- Han, B.R.X.; Lin, S.C.Y.; Espinosa, K.; Thorne, P.R.; Vlajkovic, S.M. Inhibition of the adenosine A2A receptor mitigates excitotoxic injury in organotypic tissue cultures of the rat cochlea. Cells 2019, 8, 877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sako, W.; Murakami, N.; Motohama, K.; Izumi, Y.; Kaji, R. The effect of istradefylline for Parkinson’s disease: A meta-analysis. Sci. Rep. 2017, 7, 18018. [Google Scholar] [CrossRef] [Green Version]
- Jenner, P. Istradefylline, a novel adenosine A2A receptor antagonist, for the treatment of Parkinson’s disease. Expert Opin. Investig. Drugs 2005, 14, 729–738. [Google Scholar] [CrossRef]
- Pinna, A. Adenosine A2A receptor antagonists in Parkinson’s disease: Progress in clinical trials from the newly approved istradefylline to drugs in early development and those already discontinued. CNS Drugs 2014, 28, 455–474. [Google Scholar] [CrossRef] [PubMed]
- Oishi, N.; Schacht, J. Emerging treatments for noise-induced hearing loss. Expert Opin. Emerg. Drugs 2011, 16, 235–245. [Google Scholar] [CrossRef] [Green Version]
- Sha, S.H.; Taylor, R.; Forge, A.; Schacht, J. Differential vulnerability of basal and apical hair cells is based on intrinsic susceptibility to free radicals. Hear. Res. 2001, 155, 1–8. [Google Scholar] [CrossRef]
- Yang, C.H.; Schrepfer, T.; Schacht, J. Age-related hearing impairment and the triad of acquired hearing loss. Front. Cell. Neurosci. 2015, 9, 276. [Google Scholar] [CrossRef]
- Schuknecht, H.F.; Woellner, R.C. An experimental and clinical study of deafness from lesions of the cochlear nerve. J. Laryngol. Otol. 1955, 69, 75–97. [Google Scholar] [CrossRef]
- Kujawa, S.G.; Liberman, M.C. Adding insult to injury: Cochlear nerve degeneration after “temporary” noise-induced hearing loss. J. Neurosci. 2009, 29, 14077–14085. [Google Scholar] [CrossRef] [Green Version]
- Wan, G.; Corfas, G. No longer falling on deaf ears: Mechanisms of degeneration and regeneration of cochlear ribbon synapses. Hear. Res. 2015, 329, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sergeyenko, Y.; Lall, K.; Liberman, M.C.; Kujawa, S.G. Age-related cochlear synaptopathy: An early-onset contributor to auditory functional decline. J. Neurosci. 2013, 33, 13686–13694. [Google Scholar] [CrossRef]
- Kennard, J.A.; Woodruff-Pak, D.S. Age sensitivity of behavioral tests and brain substrates of normal aging in mice. Front. Aging Neurosci. 2011, 3, 9. [Google Scholar] [CrossRef] [Green Version]
- Morris, R.G.M. Development of a water-maze procedure for studying spatial learning in the rat. J. Neurosci. Methods. 1984, 11, 47–60. [Google Scholar] [CrossRef]
- Antunes, M.; Biala, G. The novel object recognition memory: Neurobiology, test procedure, and its modifications. Cogn. Process. 2012, 13, 93–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heredia, L.; Torrente, M.; Colomina, M.T.; Domingo, J.L. Assessing anxiety in C57BL/6J mice: A pharmacological characterization of the zero maze test. J. Pharmacol. Toxicol. Methods 2013, 68, 275–283. [Google Scholar] [CrossRef]
- Cunha, R.A. Neuroprotection by adenosine in the brain: From A1 receptor activation to A2A receptor blockade. Purinerg. Signal. 2005, 1, 111–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leite, M.R.; Wilhelm, E.A.; Jesse, C.R.; Brandao, R.; Nogueira, C.W. Protective effect of caffeine and a selective A2A receptor antagonist on impairment of memory and oxidative stress of aged rats. Exp. Gerontol. 2011, 46, 309–315. [Google Scholar] [CrossRef] [Green Version]
- Dall’Igna, O.P.; Fett, P.; Gomes, M.W.; Souza, D.O.; Cunha, R.A.; Lara, D.R. Caffeine and adenosine A2A receptor antagonists prevent beta-amyloid (25–35)-induced cognitive deficits in mice. Exp. Neurol. 2007, 203, 241–245. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhao, Y.; Ning, Y.; Yang, N.; Peng, Y.; Li, P.; Chen, X.Y.; Liu, D.; Wang, H.; Chen, X.; et al. Adenosine A2A receptor inactivation alleviates early-onset cognitive dysfunction after traumatic brain injury involving an inhibition of tau hyperphosphorylation. Transl. Psychiatry 2017, 7, e1123. [Google Scholar] [CrossRef] [Green Version]
- Müller, T. The safety of istradefylline for the treatment of Parkinson’s disease. Expert Opin. Drug Saf. 2015, 14, 769–775. [Google Scholar] [CrossRef]
- Fuxe, K.; Guidolin, D.; Agnati, L.F.; Borroto-Escuela, D.O. Dopamine heteroreceptor complexes as therapeutic targets in Parkinson’s disease. Expert Opin. Ther. Targets 2015, 19, 377–398. [Google Scholar] [CrossRef]
- Vlajkovic, S.M.; Abi, S.; Wang, C.J.H.; Housley, G.D.; Thorne, P.R. Differential distribution of adenosine receptors in rat cochlea. Cell Tissue Res. 2007, 328, 461–471. [Google Scholar] [CrossRef]
- Whitworth, C.A.; Ramkumar, V.; Jones, B.; Tsukasaki, N.; Rybak, L.P. Protection against cisplatin ototoxicity by adenosine agonists. Biochem. Pharmacol. 2004, 67, 1801–1807. [Google Scholar] [CrossRef]
- Borea, P.A.; Gessi, S.; Merighi, S.; Vincenzi, F.; Varani, K. Pharmacology of adenosine receptors: The state of the art. Physiol. Rev. 2018, 98, 1591–1625. [Google Scholar] [CrossRef] [PubMed]
- Ford, M.S.; Maggirwar, S.B.; Rybak, L.P.; Whitworth, C.; Ramkumar, V. Expression and function of adenosine receptors in the chinchilla cochlea. Hear. Res. 1997, 105, 130–140. [Google Scholar] [CrossRef]
- Vlajkovic, S.M.; Chang, H.; Paek, S.Y.; Chi, H.H.T.; Sreebhavan, S.; Telang, R.S.; Tingle, M.D.; Housley, G.D.; Thorne, P.R. Adenosine amine congener as a cochlear rescue agent. Biomed. Res. Int. 2014, 2014, 841489. [Google Scholar] [CrossRef]
- Viberg, A.; Canlon, B. The guide to plotting a cochleogram. Hear. Res. 2004, 197, 1–10. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, M.; Pandya, M.; Espinosa, K.; Telang, R.; Boix, J.; Thorne, P.R.; Vlajkovic, S.M. Istradefylline Mitigates Age-Related Hearing Loss in C57BL/6J Mice. Int. J. Mol. Sci. 2021, 22, 8000. https://doi.org/10.3390/ijms22158000
Shin M, Pandya M, Espinosa K, Telang R, Boix J, Thorne PR, Vlajkovic SM. Istradefylline Mitigates Age-Related Hearing Loss in C57BL/6J Mice. International Journal of Molecular Sciences. 2021; 22(15):8000. https://doi.org/10.3390/ijms22158000
Chicago/Turabian StyleShin, Min, Madhavi Pandya, Kristan Espinosa, Ravindra Telang, Jordi Boix, Peter R. Thorne, and Srdjan M. Vlajkovic. 2021. "Istradefylline Mitigates Age-Related Hearing Loss in C57BL/6J Mice" International Journal of Molecular Sciences 22, no. 15: 8000. https://doi.org/10.3390/ijms22158000
APA StyleShin, M., Pandya, M., Espinosa, K., Telang, R., Boix, J., Thorne, P. R., & Vlajkovic, S. M. (2021). Istradefylline Mitigates Age-Related Hearing Loss in C57BL/6J Mice. International Journal of Molecular Sciences, 22(15), 8000. https://doi.org/10.3390/ijms22158000