
Role of Amine Neurotransmitters and Their Receptors in Skin Pigmentation: Therapeutic Implication


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Skin Pigmentation
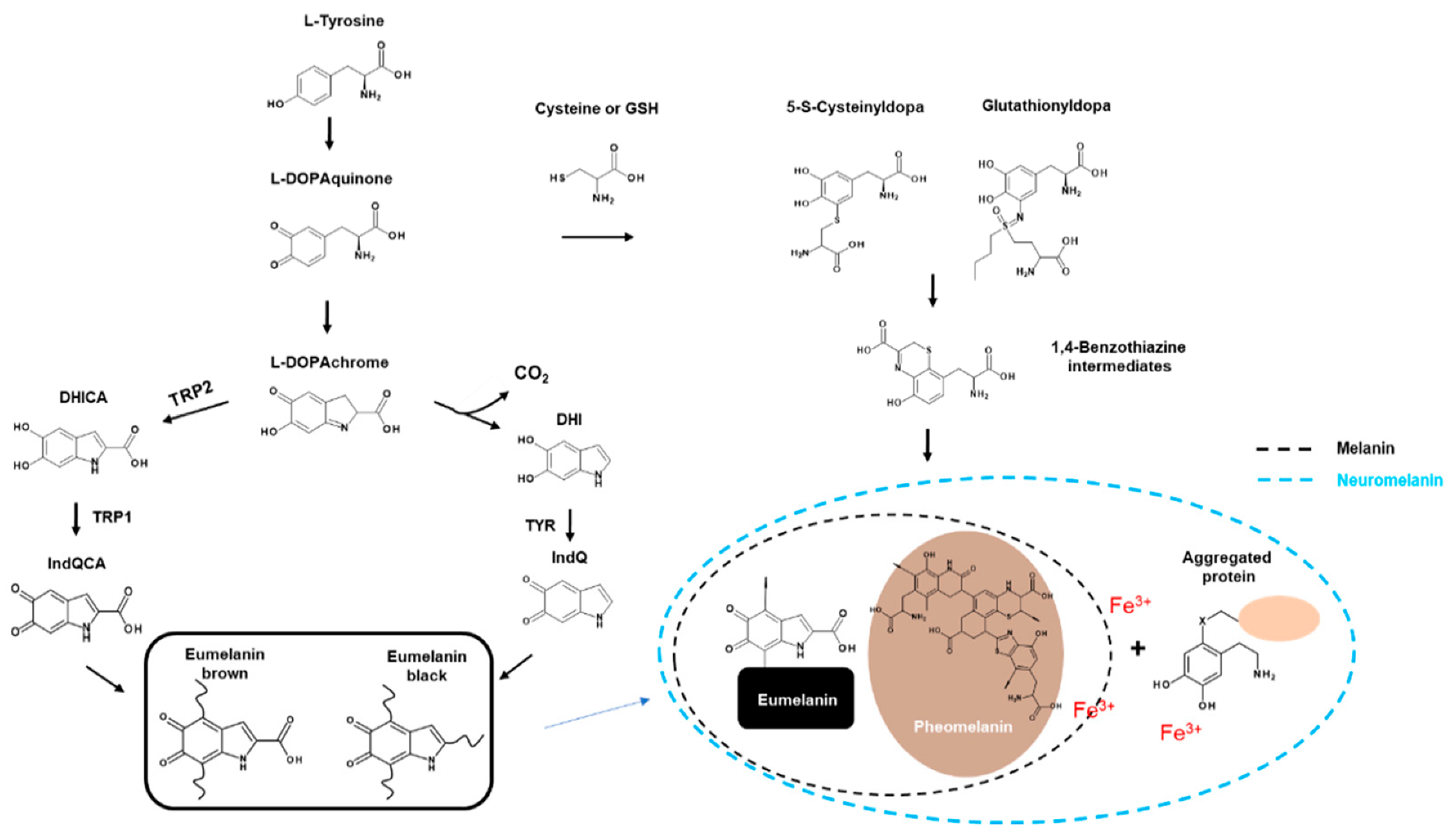
2.1. Types of Melanin
2.1.1. Eumelanin
2.1.2. Pheomelanin
2.1.3. Neuromelanin
2.2. Melanogenesis
2.2.1. Melanocytes
2.2.2. Biosynthesis of Melanin
2.3. Players in Melanogenesis
3. Pigmentation Abnormality in Skin Disease
3.1. Vitiligo and Albinism
3.2. Hyperpigmentation
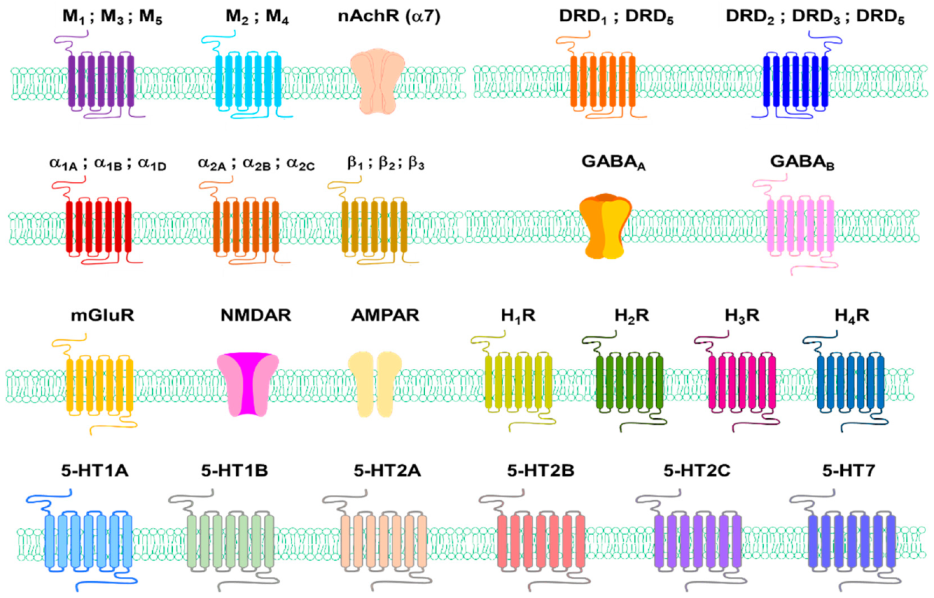
4. Amines Neurotransmitters and Their Receptors in Skin Pigmentation Diseases
4.1. Role of Neurotransmitters and Their Receptors in Melanogenesis
4.1.1. Acetylcholine
4.1.2. Dopamine
4.1.3. Epinephrine and Norepinephrine
4.1.4. Gamma-Aminobutyric Acid
4.1.5. Glutamate
4.1.6. Histamine
4.1.7. Serotonin
4.2. Role of Amine Neurotransmitters and Their Receptors for Skin Pigmentation Abnormality
4.2.1. ACh
4.2.2. DA
4.2.3. EP and NE
4.2.4. Glutamate
4.2.5. HA
4.2.6. 5-HT
5. Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chuong, C.-M.; Nickoloff, B.; Elias, P.; Goldsmith, L.; Macher, E.; Maderson, P.; Sundberg, J.; Tagami, H.; Plonka, P.; Thestrup-Pederson, K. What is the ‘true’ function of skin? Exp. Dermatol. 2002, 11, 159–187. [Google Scholar] [PubMed]
- Slominski, A.; Fischer, T.W.; Zmijewski, M.A.; Wortsman, J.; Semak, I.; Zbytek, B.; Slominski, R.M.; Tobin, D.J. On the role of melatonin in skin physiology and pathology. Endocrine 2005, 27, 137–147. [Google Scholar] [CrossRef] [Green Version]
- Jevtić, M.; Löwa, A.; Nováčková, A.; Kováčik, A.; Kaessmeyer, S.; Erdmann, G.; Vávrová, K.; Hedtrich, S. Impact of intercellular crosstalk between epidermal keratinocytes and dermal fibroblasts on skin homeostasis. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2020, 1867, 118722. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Brenner, M.; Hearing, V.J. The regulation of skin pigmentation. J. Biol. Chem. 2007, 282, 27557–27561. [Google Scholar] [CrossRef] [Green Version]
- Slominski, A.; Tobin, D.J.; Zmijewski, M.A.; Wortsman, J.; Paus, R. Melatonin in the skin: Synthesis, metabolism and functions. Trends Endocrinol. Metab. 2008, 19, 17–24. [Google Scholar] [CrossRef]
- Bertolesi, G.E.; Song, Y.N.; Atkinson-Leadbeater, K.; Yang, J.L.J.; McFarlane, S. Interaction and developmental activation of two neuroendocrine systems that regulate light-mediated skin pigmentation. Pigment Cell Melanoma Res. 2017, 30, 413–423. [Google Scholar] [CrossRef]
- Bertolesi, G.E.; Vazhappilly, S.T.; Hehr, C.L.; McFarlane, S. Pharmacological induction of skin pigmentation unveils the neuroendocrine circuit regulated by light. Pigment Cell Melanoma Res. 2016, 29, 186–198. [Google Scholar] [CrossRef]
- Lin, J.Y.; Fisher, D.E. Melanocyte biology and skin pigmentation. Nature 2007, 445, 843–850. [Google Scholar] [CrossRef]
- Lee, A.Y. Recent progress in melasma pathogenesis. Pigment Cell Melanoma Res. 2015, 28, 648–660. [Google Scholar] [CrossRef]
- Rocha, J. The evolutionary history of human skin pigmentation. J. Mol. Evol. 2020, 88, 77–87. [Google Scholar] [CrossRef]
- Filoni, A.; Mariano, M.; Cameli, N. Melasma: How hormones can modulate skin pigmentation. J. Cosmet. Dermatol. 2019, 18, 458–463. [Google Scholar] [CrossRef]
- Hawthorne, C. Addison’s Disease with Phthisis Pulmonalis and Atypical Pigmentation of the Skin; Melanoderma with Symmetrical Patches of Leucoderma. Glasg. Med. J. 1896, 46, 266. [Google Scholar]
- Yamamoto, T. Latent adrenal insufficiency: Concept, clues to detection, and diagnosis. Endocr. Pract. 2018, 24, 746–755. [Google Scholar] [CrossRef]
- Del Bino, S.; Duval, C.; Bernerd, F. Clinical and biological characterization of skin pigmentation diversity and its consequences on UV impact. Int. J. Mol. Sci. 2018, 19, 2668. [Google Scholar] [CrossRef] [Green Version]
- Ezzedine, K.; Vance, T.M.; Hamzavi, I.H.; Qureshi, A.A. Vitiligo: Targeted Therapies Add Color to Disease Pathophysiology. J. Investig. Dermatol. 2020, 140, 1498–1500. [Google Scholar] [CrossRef]
- Marçon, C.R.; Maia, M. Albinism: Epidemiology, genetics, cutaneous characterization, psychosocial factors. An. Bras. Dermatol. 2019, 94, 503–520. [Google Scholar] [CrossRef]
- Saleem, M.D.; Oussedik, E.; Picardo, M.; Schoch, J.J. Acquired disorders with hypopigmentation: A clinical approach to diagnosis and treatment. J. Am. Acad. Dermatol. 2019, 80, 1233–1250. [Google Scholar] [CrossRef]
- Dina, Y.; McKesey, J.; Pandya, A.G. Disorders of Hypopigmentation. J. Drugs Dermatol. 2019, 18, s115–s116. [Google Scholar]
- Slominski, A.; Tobin, D.J.; Shibahara, S.; Wortsman, J. Melanin pigmentation in mammalian skin and its hormonal regulation. Physiol. Rev. 2004, 84, 1155–1228. [Google Scholar] [CrossRef]
- Ohbayashi, N.; Fukuda, M. Recent advances in understanding the molecular basis of melanogenesis in melanocytes. F1000Research 2020, 9, 608. [Google Scholar] [CrossRef]
- Lee, H.J.; Park, M.K.; Kim, S.Y.; Park Choo, H.Y.; Lee, A.Y.; Lee, C.H. Serotonin induces melanogenesis via serotonin receptor 2A. Br. J. Dermatol. 2011, 165, 1344–1348. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Kosmadaki, M.; Yaar, M.; Gilchrest, B. Cellular mechanisms regulating human melanogenesis. Cell. Mol. Life Sci. 2009, 66, 1493–1506. [Google Scholar] [CrossRef] [PubMed]
- Rzepka, Z.; Buszman, E.; Beberok, A.; Wrześniok, D. From tyrosine to melanin: Signaling pathways and factors regulating melanogenesis. Postepy Hig. Med. Dosw. (Online) 2016, 70, 695–708. [Google Scholar] [CrossRef] [PubMed]
- Kondo, T.; Hearing, V.J. Update on the regulation of mammalian melanocyte function and skin pigmentation. Expert Rev. Dermatol. 2011, 6, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Lambert, M.W.; Maddukuri, S.; Karanfilian, K.M.; Elias, M.L.; Lambert, W.C. The physiology of melanin deposition in health and disease. Clin. Dermatol. 2019, 37, 402–417. [Google Scholar] [CrossRef]
- Land, E.; Ito, S.; Wakamatsu, K.; Riley, P. Rate constants for the first two chemical steps of eumelanogenesis. Pigment cell Res. 2003, 16, 487–493. [Google Scholar] [CrossRef]
- Ito, S.; Wakamatsu, K. Diversity of human hair pigmentation as studied by chemical analysis of eumelanin and pheomelanin. J. Eur. Acad. Dermatol. Venereol. 2011, 25, 1369–1380. [Google Scholar] [CrossRef]
- O’Sullivan, J.D.; Nicu, C.; Picard, M.; Chéret, J.; Bedogni, B.; Tobin, D.J.; Paus, R. The biology of human hair greying. Biol. Rev. 2021, 96, 107–128. [Google Scholar] [CrossRef]
- Palumbo, A.; Solano, F.; Misuraca, G.; Aroca, P.; Borron, J.C.G.; Lozano, J.A.; Prota, G. Comparative action of dopachrome tautomerase and metal ions on the rearrangement of dopachrome. Biochim. Biophys. Acta (BBA)-Gen. Subj. 1991, 1115, 1–5. [Google Scholar] [CrossRef]
- Wakamatsu, K.; Ohtara, K.; Ito, S. Chemical analysis of late stages of pheomelanogenesis: Conversion of dihydrobenzothiazine to a benzothiazole structure. Pigment Cell Melanoma Res. 2009, 22, 474–486. [Google Scholar] [CrossRef]
- Olivares, C.; Solano, F. New insights into the active site structure and catalytic mechanism of tyrosinase and its related proteins. Pigment Cell Melanoma Res. 2009, 22, 750–760. [Google Scholar] [CrossRef]
- Tanaka, H.; Yamashita, Y.; Umezawa, K.; Hirobe, T.; Ito, S.; Wakamatsu, K. The pro-oxidant activity of pheomelanin is significantly enhanced by UVA irradiation: Benzothiazole moieties are more reactive than benzothiazine moieties. Int. J. Mol. Sci. 2018, 19, 2889. [Google Scholar] [CrossRef] [Green Version]
- García-Borrón, J.C.; Solano, F. Molecular anatomy of tyrosinase and its related proteins: Beyond the histidine-bound metal catalytic center. Pigment Cell Res. 2002, 15, 162–173. [Google Scholar] [CrossRef]
- Zucca, F.A.; Basso, E.; Cupaioli, F.A.; Ferrari, E.; Sulzer, D.; Casella, L.; Zecca, L. Neuromelanin of the human substantia nigra: An update. Neurotox. Res. 2014, 25, 13–23. [Google Scholar] [CrossRef]
- Ghanem, G.; Fabrice, J. Tyrosinase related protein 1 (TYRP1/gp75) in human cutaneous melanoma. Mol. Oncol. 2011, 5, 150–155. [Google Scholar] [CrossRef] [Green Version]
- Vila, M. Neuromelanin, aging, and neuronal vulnerability in Parkinson’s disease. Mov. Disord. 2019, 34, 1440–1451. [Google Scholar] [CrossRef]
- Hearing, V.J. Determination of melanin synthetic pathways. J. Investig. Dermatol. 2011, 131, E8–E11. [Google Scholar] [CrossRef] [Green Version]
- Agrup, G.; Hansson, C.; Rorsman, H.; Rosengren, A.; Rosengren, E. Trichochromes in red human hair. Acta Derm-Venereol. 1978, 58, 357–358. [Google Scholar]
- Yamaguchi, Y.; Morita, A.; Maeda, A.; Hearing, V.J. Regulation of skin pigmentation and thickness by Dickkopf 1 (DKK1). J. Investig. Dermatol. Symp. Proc. 2009, 14, 73–75. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Knapp, S.K.; Iden, S. Mechanisms of melanocyte polarity and differentiation: What can we learn from other neuroectoderm-derived lineages? Curr. Opin. Cell Biol. 2020, 67, 99–108. [Google Scholar] [CrossRef]
- Lee, H.J.; Park, M.K.; Lee, E.J.; Kim, Y.L.; Kim, H.J.; Kang, J.H.; Kim, H.M.; Lee, A.Y.; Lee, C.H. Histamine receptor 2-mediated growth-differentiation factor-15 expression is involved in histamine-induced melanogenesis. Int. J. Biochem. Cell Biol. 2012, 44, 2124–2128. [Google Scholar] [CrossRef] [PubMed]
- Vandamme, N.; Berx, G. From neural crest cells to melanocytes: Cellular plasticity during development and beyond. Cell. Mol. Life Sci. 2019, 76, 1919–1934. [Google Scholar] [CrossRef] [PubMed]
- Cichorek, M.; Wachulska, M.; Stasiewicz, A.; Tymińska, A. Skin melanocytes: Biology and development. Adv. Dermatol. Allergol. /Postȩpy Dermatol. I Alergol. 2013, 30, 30. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Kurfurst, R.; Nizard, C.; Schnebert, S.; Perrier, E.; Tobin, D.J. Melanin transfer in human skin cells is mediated by filopodia—A model for homotypic and heterotypic lysosome-related organelle transfer. FASEB J. 2010, 24, 3756–3769. [Google Scholar] [CrossRef]
- Vijayasaradhi, S.; Bouchard, B.; Houghton, A. The melanoma antigen gp75 is the human homologue of the mouse b (brown) locus gene product. J. Exp. Med. 1990, 171, 1375–1380. [Google Scholar] [CrossRef]
- Liu, T.F.; Kandala, G.; Setaluri, V. PDZ domain protein GIPC interacts with the cytoplasmic tail of melanosomal membrane protein gp75 (tyrosinase-related protein-1). J. Biol. Chem. 2001, 276, 35768–35777. [Google Scholar] [CrossRef] [Green Version]
- Jackson, I.J. A cDNA encoding tyrosinase-related protein maps to the brown locus in mouse. Proc. Natl. Acad. Sci. USA 1988, 85, 4392–4396. [Google Scholar] [CrossRef] [Green Version]
- Halaban, R.; Moellmann, G. Murine and human b locus pigmentation genes encode a glycoprotein (gp75) with catalase activity. Proc. Natl. Acad. Sci. USA 1990, 87, 4809–4813. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, T.; Hearing, V.J. Direct interaction of tyrosinase with Tyrp1 to form heterodimeric complexes in vivo. J. Cell Sci. 2007, 120, 4261–4268. [Google Scholar] [CrossRef] [Green Version]
- Mockus, S.M.; Vrana, K.E. Advances in the molecular characterization of tryptophan hydroxylase. J. Mol. Neurosci. 1998, 10, 163–179. [Google Scholar] [CrossRef]
- Hoashi, T.; Sato, S.; Yamaguchi, Y.; Passeron, T.; Tamaki, K.; Hearing, V.J. Glycoprotein nonmetastatic melanoma protein b, a melanocytic cell marker, is a melanosome-specific and proteolytically released protein. FASEB J. 2010, 24, 1616–1629. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Beaulieu, J.M.; Sotnikova, T.D.; Gainetdinov, R.R.; Caron, M.G. Tryptophan hydroxylase-2 controls brain serotonin synthesis. Science 2004, 305, 217. [Google Scholar] [CrossRef]
- Theos, A.C.; Tenza, D.; Martina, J.A.; Hurbain, I.; Peden, A.A.; Sviderskaya, E.V.; Stewart, A.; Robinson, M.S.; Bennett, D.C.; Cutler, D.F. Functions of adaptor protein (AP)-3 and AP-1 in tyrosinase sorting from endosomes to melanosomes. Mol. Biol. Cell 2005, 16, 5356–5372. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Hearing, V.J. Physiological factors that regulate skin pigmentation. Biofactors 2009, 35, 193–199. [Google Scholar] [CrossRef] [Green Version]
- Brenner, M.; Hearing, V.J. The protective role of melanin against UV damage in human skin. Photochem. Photobiol. 2008, 84, 539–549. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Li, Y.; Hu, Z.; Yue, X.; Proetto, M.T.; Jones, Y.; Gianneschi, N.C. Mimicking melanosomes: Polydopamine nanoparticles as artificial microparasols. ACS Cent. Sci. 2017, 3, 564–569. [Google Scholar] [CrossRef] [Green Version]
- Land, E.J.; Ramsden, C.A.; Riley, P.A. Quinone chemistry and melanogenesis. Methods Enzym. 2004, 378, 88–109. [Google Scholar] [CrossRef]
- Schallreuter, K.U.; Kothari, S.; Chavan, B.; Spencer, J.D. Regulation of melanogenesis—Controversies and new concepts. Exp. Dermatol. 2008, 17, 395–404. [Google Scholar] [CrossRef]
- Kobayashi, T.; Urabe, K.; Winder, A.; Jiménez-Cervantes, C.; Imokawa, G.; Brewington, T.; Solano, F.; García-Borrón, J.; Hearing, V. Tyrosinase related protein 1 (TRP1) functions as a DHICA oxidase in melanin biosynthesis. EMBO J. 1994, 13, 5818–5825. [Google Scholar] [CrossRef]
- Plonka, P.M.; Handjiski, B.; Michalczyk, D.; Popik, M.; Paus, R. Oral zinc sulphate causes murine hair hypopigmentation and is a potent inhibitor of eumelanogenesis in vivo. Br. J. Derm. 2006, 155, 39–49. [Google Scholar] [CrossRef]
- Vachtenheim, J.; Borovanský, J. “Transcription physiology” of pigment formation in melanocytes: Central role of MITF. Exp. Dermatol. 2010, 19, 617–627. [Google Scholar] [CrossRef]
- Ongenae, K.; Van Geel, N.; Naeyaert, J.M. Evidence for an autoimmune pathogenesis of vitiligo. Pigment Cell Res. 2003, 16, 90–100. [Google Scholar] [CrossRef]
- Fan, K.-C.; Yang, T.-H.; Huang, Y.-C. Vitiligo and thyroid disease: A systematic review and meta-analysis. Eur. J. Dermatol. 2018, 28, 750–763. [Google Scholar]
- Alkhateeb, A.; Fain, P.R.; Thody, A.; Bennett, D.C.; Spritz, R.A. Epidemiology of vitiligo and associated autoimmune diseases in Caucasian probands and their families. Pigment. Cell Res. 2003, 16, 208–214. [Google Scholar] [CrossRef]
- Goldgeier, M.H.; Klein, L.E.; Klein-Angerer, S.; Moellmann, G.; Nordlund, J.J. The distribution of melanocytes in the leptomeninges of the human brain. J. Investig. Dermatol. 1984, 82, 235–238. [Google Scholar] [CrossRef] [Green Version]
- Garcia Hernandez, F.J.; Ocana Medina, C.; Castillo Palma, M.J.; Sanchez Roman, J.; Rueda Rueda, T.; Pulido Aguilera, M.C.; Lopez Checa, F. [Vogt-Koyanagi-Harada disease. Characteristics of a series of Andalusian patients]. Rev. Clin. Esp. 2006, 206, 388–391. [Google Scholar] [CrossRef]
- Okamura, K.; Suzuki, T. Current landscape of Oculocutaneous Albinism in Japan. Pigment Cell Melanoma Res. 2021, 34, 190–203. [Google Scholar] [CrossRef]
- Somashekar, P.H.; Girisha, K.M.; Nampoothiri, S.; Gowrishankar, K.; Devi, R.R.; Gupta, N.; Narayanan, D.L.; Kaur, A.; Bajaj, S.; Jagadeesh, S.; et al. Locus and allelic heterogeneity and phenotypic variability in Waardenburg syndrome. Clin. Genet. 2019, 95, 398–402. [Google Scholar] [CrossRef]
- Ansons, A.M.; Davis, H. Diagnosis and Management of Ocular Motility Disorders, 3rd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Summers, C.G. Albinism: Classification, clinical characteristics, and recent findings. Optom. Vis. Sci. 2009, 86, 659–662. [Google Scholar] [CrossRef]
- Fistarol, S.K.; Itin, P.H. Disorders of pigmentation. JDDG J. Dtsch. Dermatol. Ges. 2010, 8, 187–202. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Lee, P.D. A new mechanism for freckle initiation based on microstructural level simulation. Acta Mater. 2012, 60, 4917–4926. [Google Scholar] [CrossRef]
- Ebanks, J.P.; Wickett, R.R.; Boissy, R.E. Mechanisms regulating skin pigmentation: The rise and fall of complexion coloration. Int. J. Mol. Sci. 2009, 10, 4066–4087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ak, M. A comprehensive review of acne vulgaris. J. Clin. Pharm. 2019, 1, 17–45. [Google Scholar]
- Jallouli, M.; Francès, C.; Piette, J.-C.; Moguelet, P.; Factor, C.; Zahr, N.; Miyara, M.; Saadoun, D.; Mathian, A.; Haroche, J. Hydroxychloroquine-induced pigmentation in patients with systemic lupus erythematosus: A case-control study. JAMA Dermatol. 2013, 149, 935–940. [Google Scholar] [CrossRef]
- Dai, N.-T.; Chang, H.-I.; Wang, Y.-W.; Fu, K.-Y.; Huang, T.-C.; Huang, N.-C.; Li, J.-K.; Hsieh, P.-S.; Dai, L.-G.; Hsu, C.-K. Restoration of skin pigmentation after deep partial or full-thickness burn injury. Adv. Drug Deliv. Rev. 2018, 123, 155–164. [Google Scholar] [CrossRef]
- Adhikari, M.; Ali, A.; Kaushik, N.K.; Choi, E.H. Perspective in pigmentation disorders. In Comprehensive Clinical Plasma Medicine; Springer: Cham, Switzerland, 2018; pp. 363–400. [Google Scholar]
- Stulberg, D.L.; Clark, N.; Tovey, D. Common hyperpigmentation disorders in adults: Part II. Melanoma, seborrheic keratoses, acanthosis nigricans, melasma, diabetic dermopathy, tinea versicolor, and postinflammatory hyperpigmentation. Am. Fam. Physician 2003, 68, 1963–1968. [Google Scholar]
- Sinha, S.; Schwartz, R.A. Juvenile acanthosis nigricans. J. Am. Acad. Dermatol. 2007, 57, 502–508. [Google Scholar] [CrossRef]
- Ono, T.; Egawa, K.; Kayashima, K.I.; Kitoh, M. Late onset dermal melanocytosis: An upper back variant. J. Dermatol. 1991, 18, 97–103. [Google Scholar] [CrossRef]
- Nakamura, M.; Morita, A.; Seité, S.; Haarmann-Stemmann, T.; Grether-Beck, S.; Krutmann, J. Environment-induced lentigines: Formation of solar lentigines beyond ultraviolet radiation. Exp. Dermatol. 2015, 24, 407–411. [Google Scholar] [CrossRef]
- Remington, B.K.; Remington, T.K. Treatment of facial lentigines in Peutz–Jeghers syndrome with an intense pulsed light source. Dermatol. Surg. 2002, 28, 1079–1081. [Google Scholar] [CrossRef]
- Martínez-Quintana, E.; Rodríguez-González, F. LEOPARD syndrome: Clinical features and gene mutations. Mol. Syndromol. 2012, 3, 145–157. [Google Scholar] [CrossRef] [Green Version]
- Engwa, G.A.; Ferdinand, P.U.; Nwalo, F.N.; Unachukwu, M.N. Mechanism and health effects of heavy metal toxicity in humans. In Poisoning in the Modern World—New Tricks for an Old Dog? Intechopen: London, UK, 2019; Volume 10, pp. 1–23. [Google Scholar]
- Sotaniemi, E.; Kreus, K.; Kaipainen, W. Pigmentation and oral contraceptives. Br. Med. J. 1968, 2, 120. [Google Scholar] [CrossRef] [Green Version]
- Barthel, A.; Benker, G.; Berens, K.; Diederich, S.; Manfras, B.; Gruber, M.; Kanczkowski, W.; Kline, G.; Kamvissi-Lorenz, V.; Hahner, S. An update on Addison’s disease. Exp. Clin. Endocrinol. Diabetes 2019, 127, 165–175. [Google Scholar] [CrossRef] [Green Version]
- Mills, P.R.; Skerrow, C.J.; MacKie, R.M. Melanin pigmentation of the skin in primary biliary cirrhosis. J. Cutan. Pathol. 1981, 8, 404–410. [Google Scholar] [CrossRef]
- García, R.M.G.; Molina, S.C. Drug-induced hyperpigmentation: Review and case series. J. Am. Board Fam. Med. 2019, 32, 628–638. [Google Scholar] [CrossRef] [Green Version]
- Das, S.; Kourosh, A.S. Pigment changes and drug reactions. In Cutaneous Drug Eruptions; Springer: London, UK, 2015; pp. 87–106. [Google Scholar]
- Granstein, R.D.; Sober, A.J. Drug-and heavy metal-induced hyperpigmentation. J. Am. Acad. Dermatol. 1981, 5, 1–18. [Google Scholar] [CrossRef]
- Dang, N.; Meng, X.; Song, H. Nicotinic acetylcholine receptors and cancer. Biomed. Rep. 2016, 4, 515–518. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Cheuk, I.W.Y.; Shin, V.Y.; Kwong, A. Acetylcholine receptors: Key players in cancer development. Surg. Oncol. 2019, 31, 46–53. [Google Scholar] [CrossRef]
- Ishii, M.; Kurachi, Y. Muscarinic acetylcholine receptors. Curr. Pharm. Des. 2006, 12, 3573–3581. [Google Scholar] [CrossRef]
- Kawashima, K.; Fujii, T. Basic and clinical aspects of non-neuronal acetylcholine: Overview of non-neuronal cholinergic systems and their biological significance. J. Pharmacol. Sci. 2008, 106, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Elwary, S.M.; Chavan, B.; Schallreuter, K.U. The vesicular acetylcholine transporter is present in melanocytes and keratinocytes in the human epidermis. J. Investig. Dermatol. 2006, 126, 1879–1884. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Xia, Y.; Dai, K.; Bai, P.; Kwan, K.K.L.; Guo, M.S.S.; Dong, T.T.X.; Tsim, K.W.K. Solar light induces the release of acetylcholine from skin keratinocytes affecting melanogenesis. FASEB J. 2020, 34, 8941–8958. [Google Scholar] [CrossRef]
- Wu, Q.; Fung, A.H.Y.; Xu, M.L.; Poon, K.; Liu, E.Y.L.; Kong, X.P.; Yao, P.; Xiong, Q.P.; Dong, T.T.X.; Tsim, K.W.K. Microphthalmia-associated transcription factor up-regulates acetylcholinesterase expression during melanogenesis of murine melanoma cells. J. Biol. Chem. 2018, 293, 14417–14428. [Google Scholar] [CrossRef] [Green Version]
- Lamacz, M.; Tonon, M.C.; Louiset, E.; Cazin, L.; Strosberg, D.; Vaudry, H. Acetylcholine stimulates alpha-melanocyte-stimulating hormone release from frog pituitary melanotrophs through activation of muscarinic and nicotinic receptors. Endocrinology 1989, 125, 707–714. [Google Scholar] [CrossRef]
- Fujii, R.; Miyashita, Y.; Fujii, Y. Muscarinic cholinoceptors mediate neurally evoked pigment aggregation in glass catfish melanophores. J. Neural Transm. 1982, 54, 29–39. [Google Scholar] [CrossRef]
- Buchli, R.; Ndoye, A.; Arredondo, J.; Webber, R.J.; Grando, S.A. Identification and characterization of muscarinic acetylcholine receptor subtypes expressed in human skin melanocytes. Mol. Cell. Biochem. 2001, 228, 57–72. [Google Scholar] [CrossRef]
- Hasse, S.; Chernyavsky, A.I.; Grando, S.A.; Paus, R. The M4 muscarinic acetylcholine receptor plays a key role in the control of murine hair follicle cycling and pigmentation. Life Sci. 2007, 80, 2248–2252. [Google Scholar] [CrossRef] [Green Version]
- Van Strien, F.J.; Roubos, E.W.; Vaudry, H.; Jenks, B.G. Acetylcholine autoexcites the release of proopiomelanocortin-derived peptides from melanotrope cells of Xenopus laevis via an M1 muscarinic receptor. Endocrinology 1996, 137, 4298–4307. [Google Scholar] [CrossRef] [Green Version]
- Khanam, S.; Siddique, Y.H. Dopamine: Agonists and neurodegenerative disorders. Curr. Drug Targets 2018, 19, 1599–1611. [Google Scholar] [CrossRef]
- Vallone, D.; Picetti, R.; Borrelli, E. Structure and function of dopamine receptors. Neurosci. Biobehav. Rev. 2000, 24, 125–132. [Google Scholar] [CrossRef]
- Tursunov, D. Assessment of L-Dopa induced dyskinesia. Parkinsonism Relat. Disord. 2018, 46, e45. [Google Scholar] [CrossRef]
- Amenta, F.; Ricci, A.; Rossodivita, I.; Avola, R.; Tayebati, S. The dopaminergic system in hypertension. Clin. Exp. Hypertens. 2001, 23, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Beaulieu, J.-M.; Gainetdinov, R.R. The physiology, signaling, and pharmacology of dopamine receptors. Pharmacol. Rev. 2011, 63, 182–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tammaro, A.; Cavallotti, C.; Gaspari, A.; Narcisi, A.; Parisella, F. Dopaminergic receptors in the human skin. J. Biol. Regul. Homeost. Agents 2012, 26, 789–795. [Google Scholar] [PubMed]
- Lezcano, N.E.; Salvatierra, N.A.; Celis, M.E. α-Melanotropin hormone inhibits the binding of [3H] SCH 23390 to the dopamine D1 receptor in vitro. Eur. J. Pharmacol. 1998, 363, 211–215. [Google Scholar] [CrossRef]
- Chu, C.Y.; Liu, Y.L.; Chiu, H.C.; Jee, S.H. Dopamine-induced apoptosis in human melanocytes involves generation of reactive oxygen species. Br. J. Dermatol. 2006, 154, 1071–1079. [Google Scholar] [CrossRef] [PubMed]
- Hedges, D.M.; Yorgason, J.T.; Perez, A.W.; Schilaty, N.D.; Williams, B.M.; Watt, R.K.; Steffensen, S.C. Spontaneous Formation of Melanin from Dopamine in the Presence of Iron. Antioxidants 2020, 9, 1285. [Google Scholar] [CrossRef]
- Kim, B.; Hwang, J.S.; Kim, H.S. N-Nicotinoyl dopamine inhibits skin pigmentation by suppressing of melanosome transfer. Eur. J. Pharmacol. 2015, 769, 250–256. [Google Scholar] [CrossRef]
- Kim, Y.H.; Park, J.I.; hwan Myung, C.; eun Lee, J.; Bang, S.; Chang, S.E.; Hwang, J.S. 1-Phenyl-3-(2-thiazolyl)-2-thiourea inhibits melanogenesis via a dual-action mechanism. Arch. Dermatol. Res. 2016, 308, 473–479. [Google Scholar] [CrossRef]
- Leoty-Okombi, S.; Bonnet, S.; Rival, D.; Degrave, V.; Lin, X.; Vogelgesang, B.; Andre-Frei, V. In vitro melanogenesis inhibitory effects of N-feruloyldopamine. J. Cosmet. Sci. 2013, 64, 133–144. [Google Scholar]
- Burchill, S.A.; Thody, A.J. Dopaminergic inhibition of tyrosinase activity in hair follicular melanocytes of the mouse. J. Endocrinol. 1986, 111, 233–237. [Google Scholar] [CrossRef]
- Jung, J.M.; Kim, S.Y.; Lee, W.J.; Hwang, J.S.; Chang, S.E. Dopamine D4 receptor antagonist inhibits melanogenesis through transcriptional downregulation of MITF via ERK signalling. Exp. Dermatol. 2016, 25, 325–328. [Google Scholar] [CrossRef] [Green Version]
- Le Poole, I.; Van den Wijngaard, R.; Smit, N.; Oosting, J.; Westerhof, W.; Pavel, S. Catechol-O-methyltransferase in vitiligo. Arch. Dermatol. Res. 1994, 286, 81–86. [Google Scholar] [CrossRef]
- Myburgh, J. Norepinephrine: More of a neurohormone than a vasopressor. Crit. Care 2010, 14, 196. [Google Scholar] [CrossRef] [Green Version]
- Mittal, R.; Debs, L.H.; Patel, A.P.; Nguyen, D.; Patel, K.; O’Connor, G.; Grati, M.; Mittal, J.; Yan, D.; Eshraghi, A.A.; et al. Neurotransmitters: The Critical Modulators Regulating Gut-Brain Axis. J. Cell. Physiol. 2017, 232, 2359–2372. [Google Scholar] [CrossRef] [Green Version]
- Axelrod, J.; Reisine, T.D. Stress hormones: Their interaction and regulation. Science 1984, 224, 452–459. [Google Scholar] [CrossRef] [Green Version]
- Tank, A.W.; Lee Wong, D. Peripheral and central effects of circulating catecholamines. Compr. Physiol. 2015, 5, 1–15. [Google Scholar] [CrossRef]
- Pierce, K.L.; Premont, R.T.; Lefkowitz, R.J. Seven-transmembrane receptors. Nat. Rev. Mol. Cell Biol. 2002, 3, 639–650. [Google Scholar] [CrossRef]
- Bylund, D.B.; Eikenberg, D.C.; Hieble, J.P.; Langer, S.Z.; Lefkowitz, R.J.; Minneman, K.P.; Molinoff, P.B.; Ruffolo, R.R., Jr.; Trendelenburg, U. International Union of Pharmacology nomenclature of adrenoceptors. Pharmacol. Rev. 1994, 46, 121–136. [Google Scholar]
- Schallreuter, K.U.; Korner, C.; Pittelkow, M.R.; Swanson, N.N.; Gardner, M.L. The induction of the alpha-1-adrenoceptor signal transduction system on human melanocytes. Exp. Dermatol. 1996, 5, 20–23. [Google Scholar] [CrossRef]
- Scarparo, A.C.; Visconti, M.A.; de Oliveira, A.R.; Castrucci, A.M. Adrenoceptors in normal and malignant human melanocytes. Arch. Dermatol. Res. 2000, 292, 265–267. [Google Scholar] [CrossRef]
- Gillbro, J.M.; Marles, L.K.; Hibberts, N.A.; Schallreuter, K.U. Autocrine catecholamine biosynthesis and the beta-adrenoceptor signal promote pigmentation in human epidermal melanocytes. J. Investig. Dermatol. 2004, 123, 346–353. [Google Scholar] [CrossRef] [Green Version]
- Moretti, S.; Massi, D.; Farini, V.; Baroni, G.; Parri, M.; Innocenti, S.; Cecchi, R.; Chiarugi, P. beta-adrenoceptors are upregulated in human melanoma and their activation releases pro-tumorigenic cytokines and metalloproteases in melanoma cell lines. Lab. Investig. 2013, 93, 279–290. [Google Scholar] [CrossRef] [Green Version]
- Kindermann, C.; Narayan, E.J.; Hero, J.M. The neuro-hormonal control of rapid dynamic skin colour change in an amphibian during amplexus. PLoS ONE 2014, 9, e114120. [Google Scholar] [CrossRef] [Green Version]
- Kjær, A.; Knigge, U.; Matzen, S.; Warberg, J. β-Adrenergic receptors are involved in histamine-induced secretion of proopiomelanocortin-derived peptides and prolactin in rats. Eur. J. Endocrinol. 1995, 132, 223–228. [Google Scholar] [CrossRef]
- Lan, W.-J.; Wang, H.-Y.; Lan, W.; Wang, K.-Y.; Wang, R.-M. Evidence that geniposide abrogates norepinephrine-induced hypopigmentation by the activation of GLP-1R-dependent c-kit receptor signaling in melanocyte. J. Ethnopharmacol. 2008, 118, 154–158. [Google Scholar] [CrossRef]
- Orecchia, G. Alternative therapies for vitiligo. In Vitiligo-a Monograph on the Basic and Clinical Science; Hann, S.K., Nordlund, J.J., Eds.; Blackwell Science: Oxford, UK, 2000. [Google Scholar]
- Mendelsohn, A.R.; Larrick, J.W. The Danger of Being Too Sympathetic: Norepinephrine in Alzheimer’s Disease and Graying of Hair. Rejuvenation Res. 2020, 23, 68–72. [Google Scholar] [CrossRef] [Green Version]
- Schallreuter, K.U.; Wood, J.M.; Lemke, R.; LePoole, C.; Das, P.; Westerhof, W.; Pittelkow, M.R.; Thody, A.J. Production of catecholamines in the human epidermis. Biochem. Biophys. Res. Commun. 1992, 189, 72–78. [Google Scholar] [CrossRef]
- Watanabe, M.; Maemura, K.; Kanbara, K.; Tamayama, T.; Hayasaki, H. GABA and GABA receptors in the central nervous system and other organs. Int. Rev. Cytol. 2002, 213, 1–47. [Google Scholar] [CrossRef]
- Denda, M.; Inoue, K.; Inomata, S.; Denda, S. γ-aminobutyric acid (A) receptor agonists accelerate cutaneous barrier recovery and prevent epidermal hyperplasia induced by barrier disruption. J. Investig. Dermatol. 2002, 119, 1041–1047. [Google Scholar] [CrossRef] [Green Version]
- Stoebner, P.; Carayon, P.; Penarier, G.; Frechin, N.; Barneon, G.; Casellas, P.; Cano, J.; Meynadier, J.; Meunier, L. The expression of peripheral benzodiazepine receptors in human skin: The relationship with epidermal cell differentiation. Br. J. Dermatol. 1999, 140, 1010–1016. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.R.; Skeath, J.B.; Johnson, S.L. Maintenance of Melanocyte Stem Cell Quiescence by GABA-A Signaling in Larval Zebrafish. Genetics 2019, 213, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Pomeranz Krummel, D.A.; Nasti, T.H.; Kaluzova, M.; Kallay, L.; Bhattacharya, D.; Melms, J.C.; Izar, B.; Xu, M.; Burnham, A.; Ahmed, T.; et al. Melanoma Cell Intrinsic GABAA Receptor Enhancement Potentiates Radiation and Immune Checkpoint Inhibitor Response by Promoting Direct and T Cell-Mediated Antitumor Activity. Int. J. Radiat. Oncol. Biol. Phys. 2021, 109, 1040–1053. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.A.; Jeon, H.Y.; Kim, M.; Lee, Y.J.; Han, E.T.; Park, W.S.; Hong, S.H.; Kim, Y.M.; Ha, K.S. Anti-metastatic effect of midazolam on melanoma B16F10 cells in the lungs of diabetic mice. Biochem. Pharmacol. 2020, 178, 114052. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Fu, Y.; Gao, R.; Li, J.; Kang, M.; Song, G.; Yun, C. Diazepam enhances melanogenesis, melanocyte dendricity and melanosome transport via the PBR/cAMP/PKA pathway. Int. J. Biochem. Cell Biol. 2019, 116, 105620. [Google Scholar] [CrossRef] [PubMed]
- Matthew, E.; Laskin, J.D.; Zimmerman, E.A.; Weinstein, I.B.; Hsu, K.C.; Engelhardt, D.L. Benzodiazepines have high-affinity binding sites and induce melanogenesis in B16/C3 melanoma cells. Proc. Natl. Acad. Sci. USA 1981, 78, 3935–3939. [Google Scholar] [CrossRef] [Green Version]
- Fuziwara, S.; Inoue, K.; Denda, M. NMDA-type glutamate receptor is associated with cutaneous barrier homeostasis. J. Investig. Dermatol. 2003, 120, 1023–1029. [Google Scholar] [CrossRef] [Green Version]
- Teh, J.L.; Shah, R.; Shin, S.S.; Wen, Y.; Mehnert, J.M.; Goydos, J.; Chen, S. Metabotropic glutamate receptor 1 mediates melanocyte transformation via transactivation of insulin-like growth factor 1 receptor. Pigment Cell Melanoma Res. 2014, 27, 621–629. [Google Scholar] [CrossRef] [Green Version]
- Hoogduijn, M.; Hitchcock, I.; Smit, N.; Gillbro, J.; Schallreuter, K.; Genever, P. Glutamate receptors on human melanocytes regulate the expression of MiTF. Pigment Cell Res. 2006, 19, 58–67. [Google Scholar] [CrossRef]
- Choi, K.Y.; Chang, K.; Pickel, J.M.; Badger, J.D., 2nd; Roche, K.W. Expression of the metabotropic glutamate receptor 5 (mGluR5) induces melanoma in transgenic mice. Proc. Natl. Acad. Sci. USA 2011, 108, 15219–15224. [Google Scholar] [CrossRef] [Green Version]
- Devi, S.; Markandeya, Y.; Maddodi, N.; Dhingra, A.; Vardi, N.; Balijepalli, R.C.; Setaluri, V. Metabotropic glutamate receptor 6 signaling enhances TRPM1 calcium channel function and increases melanin content in human melanocytes. Pigment Cell Melanoma Res. 2013, 26, 348–356. [Google Scholar] [CrossRef] [Green Version]
- Ni, J.; Wang, N.; Gao, L.; Li, L.; Zheng, S.; Liu, Y.; Ozukum, M.; Nikiforova, A.; Zhao, G.; Song, Z. The effect of the NMDA receptor-dependent signaling pathway on cell morphology and melanosome transfer in melanocytes. J. Dermatol. Sci. 2016, 84, 296–304. [Google Scholar] [CrossRef]
- Hajdu, T.; Juhasz, T.; Szucs-Somogyi, C.; Racz, K.; Zakany, R. NR1 and NR3B Composed Intranuclear N-methyl-d-aspartate Receptor Complexes in Human Melanoma Cells. Int. J. Mol. Sci. 2018, 19, 1929. [Google Scholar] [CrossRef] [Green Version]
- Yang, N.; Mu, L.; Zhao, B.; Wang, M.; Hu, S.; Zhao, B.; Chen, Y.; Wu, X. RNAi-mediated SLC7A11 knockdown inhibits melanogenesis-related genes expression in rabbit skin fibroblasts. J. Genet. 2018, 97, 463–468. [Google Scholar] [CrossRef]
- Marieb, E.; Hoehn, K. The Respiratory System. In Human Anatomy and Physiology; Benjamin Cummings: San Francisco, CA, USA, 2001. [Google Scholar]
- Nieto-Alamilla, G.; Márquez-Gómez, R.; García-Gálvez, A.-M.; Morales-Figueroa, G.-E.; Arias-Montaño, J.-A. The histamine H3 receptor: Structure, pharmacology, and function. Mol. Pharmacol. 2016, 90, 649–673. [Google Scholar] [CrossRef] [Green Version]
- Lieberman, P. The basics of histamine biology. Ann. Allergy Asthma Immunol. 2011, 106, S2–S5. [Google Scholar] [CrossRef]
- Albrecht, M.; Dittrich, A. Expression and function of histamine and its receptors in atopic dermatitis. Mol. Cell. Pediatrics 2015, 2, 1–8. [Google Scholar] [CrossRef]
- Reynolds, J.; Akhter, J.; Morris, D. In vitro effect of histamine and histamine H1 and H2 receptor antagonists on cellular proliferation of human malignant melanoma cell lines. Melanoma Res. 1996, 6, 95–99. [Google Scholar] [CrossRef]
- McEwan, M.T.; Parsons, P.G. Regulation of tyrosinase expression and activity in human melanoma cells via histamine receptors. J. Investig. Dermatol. 1991, 97, 868–873. [Google Scholar] [CrossRef] [Green Version]
- Chang, T.-S.; Chen, C.-T. Inhibitory effect of homochlorcyclizine on melanogenesis in α-melanocyte stimulating hormone-stimulated mouse B16 melanoma cells. Arch. Pharmacal Res. 2012, 35, 119–127. [Google Scholar] [CrossRef]
- Jangi, S.-M.; Díaz-Pérez, J.L.; Ochoa-Lizarralde, B.; Martín-Ruiz, I.; Asumendi, A.; Pérez-Yarza, G.; Gardeazabal, J.; Díaz-Ramón, J.L.; Boyano, M.D. H1 histamine receptor antagonists induce genotoxic and caspase-2-dependent apoptosis in human melanoma cells. Carcinogenesis 2006, 27, 1787–1796. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.R.; Jo, S.Y.; Kim, H.T.; Lee, W.J.; Won, C.H.; Lee, M.W.; Choi, J.H.; Chang, S.E. Loratadine, an H1 Antihistamine, Inhibits Melanogenesis in Human Melanocytes. BioMed Res. Int. 2019, 2019, 5971546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, M.; Takahashi, Y.; Inoue, S. Histamine induces melanogenesis and morphologic changes by protein kinase A activation via H2 receptors in human normal melanocytes. J. Investig. Dermatol 2000, 114, 334–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, N.H.; Lee, A.Y. Histamine effect on melanocyte proliferation and vitiliginous keratinocyte survival. Exp. Dermatol. 2010, 19, 1073–1079. [Google Scholar] [CrossRef] [PubMed]
- Le Gros, G.; Zhang, X.; Parsons, P. Alteration of tyrosinase activity in human melanocytes and melanoma cells by histamine H2 and H3 ligands. Melanoma Res. 1994, 4, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Massari, N.A.; Nicoud, M.B.; Sambuco, L.; Cricco, G.P.; Lamas, D.J.M.; Ducloux, M.V.H.; Blanco, H.; Rivera, E.S.; Medina, V.A. Histamine therapeutic efficacy in metastatic melanoma: Role of histamine H4 receptor agonists and opportunity for combination with radiation. Oncotarget 2017, 8, 26471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pytliak, M.; Vargova, V.; Mechirova, V.; Felsoci, M. Serotonin receptors—From molecular biology to clinical applications. Physiol. Res. 2011, 60, 15–25. [Google Scholar] [CrossRef]
- Johansson, O.; Liu, P.-Y.; Liang, Y.; Gangi, S.; Bondesson, L.; Nordlind, K.; Olsson, M.J.; Löntz, W.; Verhofstad, A. A serotonin-like immunoreactivity is present in human cutaneous melanocytes. J. Investig. Dermatol. 1998, 111, 1010–1014. [Google Scholar] [CrossRef]
- English, K.B.; Wang, Z.Z.; Stayner, N.; Stensaas, L.J.; Martin, H.; Tuckett, R.P. Serotonin-like immunoreactivity in Merkel cells and their afferent neurons in touch domes from the hairy skin of rats. Anat. Rec. 1992, 232, 112–120. [Google Scholar] [CrossRef]
- Slominski, A.; Pisarchik, A.; Semak, I.; Sweatman, T.; Wortsman, J.; Szczesniewski, A.; Slugocki, G.; McNulty, J.; Kauser, S.; Tobin, D.J. Serotoninergic and melatoninergic systems are fully expressed in human skin. FASEB J. 2002, 16, 896–898. [Google Scholar] [CrossRef] [Green Version]
- Lefebvre, H.; Compagnon, P.; Contesse, V.; Delarue, C.; Thuillez, C.; Vaudry, H.; Kuhn, J.-M. Production and metabolism of serotonin (5-HT) by the human adrenal cortex: Paracrine stimulation of aldosterone secretion by 5-HT. J. Clin. Endocrinol. Metab. 2001, 86, 5001–5007. [Google Scholar] [CrossRef]
- Fink, K.B.; Göthert, M. 5-HT receptor regulation of neurotransmitter release. Pharmacol. Rev. 2007, 59, 360–417. [Google Scholar] [CrossRef]
- Talley, N.J. Serotoninergic neuroenteric modulators. Lancet 2001, 358, 2061–2068. [Google Scholar] [CrossRef]
- Slominski, A.; Pisarchik, A.; Zbytek, B.; Tobin, D.J.; Kauser, S.; Wortsman, J. Functional activity of serotoninergic and melatoninergic systems expressed in the skin. J. Cell. Physiol. 2003, 196, 144–153. [Google Scholar] [CrossRef]
- Bockaert, J.; Claeysen, S.; Bécamel, C.; Dumuis, A.; Marin, P. Neuronal 5-HT metabotropic receptors: Fine-tuning of their structure, signaling, and roles in synaptic modulation. Cell Tissue Res. 2006, 326, 553–572. [Google Scholar] [CrossRef]
- Slominski, A.; Wortsman, J.; Tobin, D.J. The cutaneous serotoninergic/melatoninergic system: Securing a place under the sun. FASEB J. 2005, 19, 176–194. [Google Scholar] [CrossRef]
- Lundeberg, L.; El-Nour, H.; Mohabbati, S.; Morales, M.; Azmitia, E.; Nordlind, K. Expression of serotonin receptors in allergic contact eczematous human skin. Arch. Dermatol. Res. 2002, 294, 393–398. [Google Scholar] [CrossRef]
- Nordlind, K.; Azmitia, E.C.; Slominski, A. The skin as a mirror of the soul: Exploring the possible roles of serotonin. Exp. Dermatol. 2008, 17, 301–311. [Google Scholar] [CrossRef]
- Slominski, A.; Pisarchik, A.; Johansson, O.; Jing, C.; Semak, I.; Slugocki, G.; Wortsman, J. Tryptophan hydroxylase expression in human skin cells. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2003, 1639, 80–86. [Google Scholar] [CrossRef] [Green Version]
- Oh, E.J.; Park, J.I.; Lee, J.E.; Myung, C.H.; Kim, S.Y.; Chang, S.E.; Hwang, J.S. A Novel Role of Serotonin Receptor 2B Agonist as an Anti-Melanogenesis Agent. Int. J. Mol. Sci. 2016, 17, 546. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Cai, M.; Ren, Y.; Wu, H.; Liu, M.; Chen, H.; Shang, J. The different roles of 5-HT1A/2A receptors in fluoxetine ameliorated pigmentation of C57BL/6 mouse skin in response to stress. J. Dermatol. Sci. 2018, 92, 222–229. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Fu, M.; Pei, S.; Zhou, L.; Shang, J. R-Fluoxetine Increases Melanin Synthesis Through a 5-HT1A/2A Receptor and p38 MAPK Signaling Pathways. Int. J. Mol. Sci. 2018, 20, 80. [Google Scholar] [CrossRef] [Green Version]
- Perdomo, J.; Quintana, C.; Gonzalez, I.; Hernandez, I.; Rubio, S.; Loro, J.F.; Reiter, R.J.; Estevez, F.; Quintana, J. Melatonin Induces Melanogenesis in Human SK-MEL-1 Melanoma Cells Involving Glycogen Synthase Kinase-3 and Reactive Oxygen Species. Int. J. Mol. Sci. 2020, 21, 4970. [Google Scholar] [CrossRef]
- Kim, T.K.; Lin, Z.; Tidwell, W.J.; Li, W.; Slominski, A.T. Melatonin and its metabolites accumulate in the human epidermis in vivo and inhibit proliferation and tyrosinase activity in epidermal melanocytes in vitro. Mol. Cell. Endocrinol. 2015, 404, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Cabrera, J.; Negrin, G.; Estevez, F.; Loro, J.; Reiter, R.J.; Quintana, J. Melatonin decreases cell proliferation and induces melanogenesis in human melanoma SK-MEL-1 cells. J. Pineal. Res. 2010, 49, 45–54. [Google Scholar] [CrossRef]
- Slominski, A.; Pruski, D. Melatonin inhibits proliferation and melanogenesis in rodent melanoma cells. Exp. Cell Res. 1993, 206, 189–194. [Google Scholar] [CrossRef]
- BinSaif, G.A.; Al Samary, A.; Al Mohizea, S. Failure of botulinum toxin treatment for localized vitiligo. J. Drugs Dermatol. 2010, 9, 1092–1094. [Google Scholar]
- Iyengar, B. Modulation of melanocytic activity by acetylcholine. Acta Anat. (Basel) 1989, 136, 139–141. [Google Scholar] [CrossRef] [PubMed]
- Kurzen, H.; Schallreuter, K.U. Novel aspects in cutaneous biology of acetylcholine synthesis and acetylcholine receptors. Exp. Dermatol. 2004, 13, 27–30. [Google Scholar] [CrossRef]
- Said, E.R.; Nagui, N.; Rashed, L.A.; Mostafa, W.Z. Oxidative stress and the cholinergic system in non-segmental vitiligo: Effect of narrow band ultraviolet b. Photodermatol. Photoimmunol. Photomed. 2021, 37, 306–312. [Google Scholar] [CrossRef]
- Schallreuter, K.U.; Elwary, S.M.; Gibbons, N.C.; Rokos, H.; Wood, J.M. Activation/deactivation of acetylcholinesterase by H2O2: More evidence for oxidative stress in vitiligo. Biochem. Biophys. Res. Commun. 2004, 315, 502–508. [Google Scholar] [CrossRef] [PubMed]
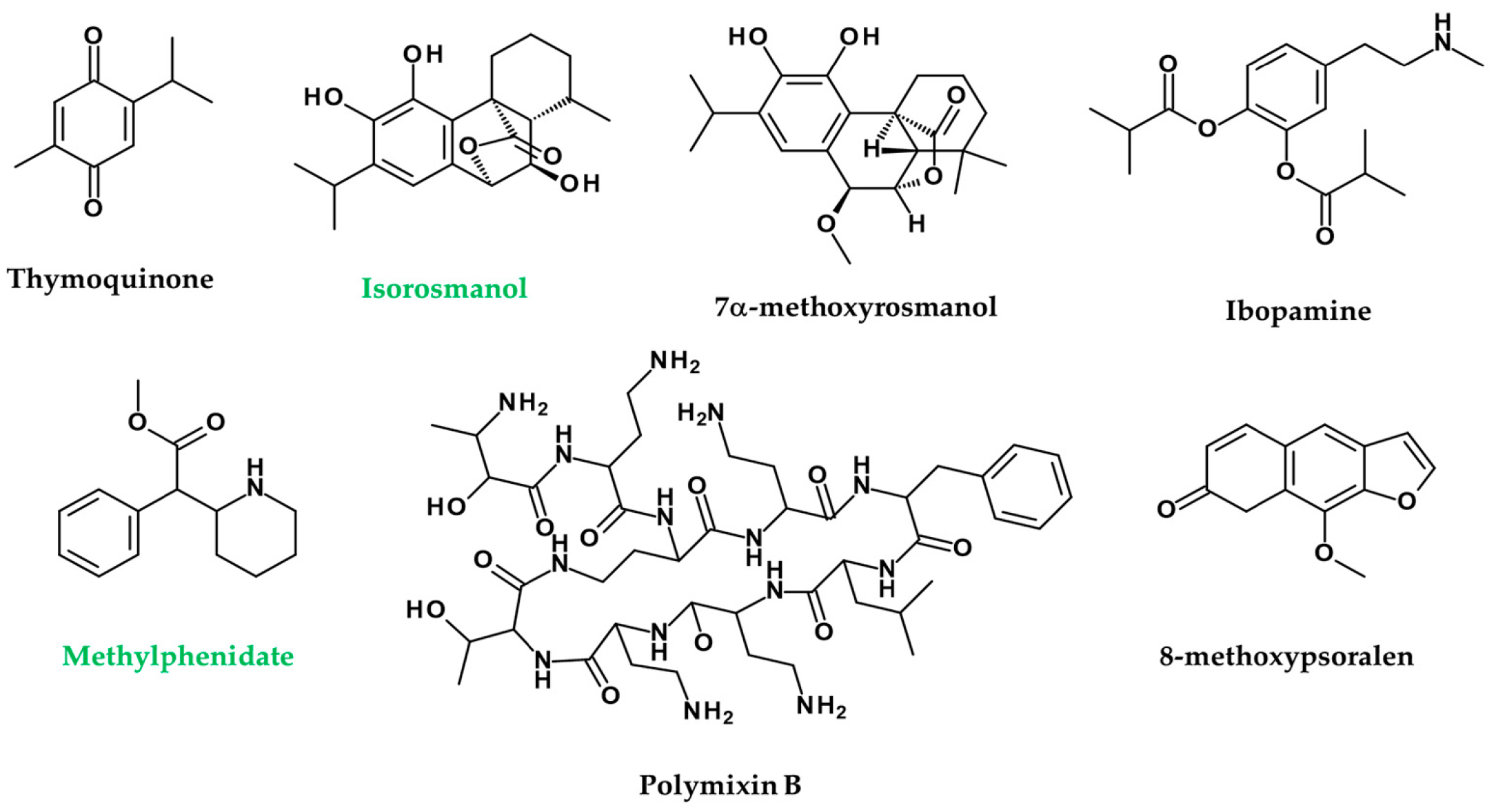
- Ali, S.A.; Meitei, K.V. Nigella sativa seed extract and its bioactive compound thymoquinone: The new melanogens causing hyperpigmentation in the wall lizard melanophores. J. Pharm. Pharmacol. 2011, 63, 741–746. [Google Scholar] [CrossRef] [PubMed]
- Sallam, A.; Mira, A.; Ashour, A.; Shimizu, K. Acetylcholine esterase inhibitors and melanin synthesis inhibitors from Salvia officinalis. Phytomedicine 2016, 23, 1005–1011. [Google Scholar] [CrossRef]
- Kotb El-Sayed, M.-I.; El-Ghany, A.; Ahmed, A.; Mohamed, R.R. Neural and endocrinal pathobiochemistry of vitiligo: Comparative study for a hypothesized mechanism. Front. Endocrinol. 2018, 9, 197. [Google Scholar] [CrossRef]
- Reimann, E.; Kingo, K.; Karelson, M.; Reemann, P.; Loite, U.; Keermann, M.; Abram, K.; Vasar, E.; Silm, H.; Koks, S. Expression profile of genes associated with the dopamine pathway in vitiligo skin biopsies and blood sera. Dermatology 2012, 224, 168–176. [Google Scholar] [CrossRef]
- Kingo, K.; Aunin, E.; Karelson, M.; Philips, M.-A.; Rätsep, R.; Silm, H.; Vasar, E.; Soomets, U.; Kõks, S. Gene expression analysis of melanocortin system in vitiligo. J. Dermatol. Sci. 2007, 48, 113–122. [Google Scholar] [CrossRef]
- Kanoff, J.M.; Colby, K. Pigmented deposits on a Boston keratoprosthesis from topical ibopamine. Cornea 2010, 29, 1069–1071. [Google Scholar] [CrossRef]
- Guler Aksu, G.; Akdere, P.; Toros, F. Methylphenidate associated vitiligo in a child: A case report. Asia-Pac. Psychiatry 2019, 11, e12351. [Google Scholar] [CrossRef]
- Giordano, F.; Bonetti, C.; Surace, E.M.; Marigo, V.; Raposo, G. The ocular albinism type 1 (OA1) G-protein-coupled receptor functions with MART-1 at early stages of melanogenesis to control melanosome identity and composition. Hum. Mol. Genet. 2009, 18, 4530–4545. [Google Scholar] [CrossRef] [Green Version]
- Vetrini, F.; Auricchio, A.; Du, J.; Angeletti, B.; Fisher, D.E.; Ballabio, A.; Marigo, V. The microphthalmia transcription factor (Mitf) controls expression of the ocular albinism type 1 gene: Link between melanin synthesis and melanosome biogenesis. Mol. Cell. Biol. 2004, 24, 6550–6559. [Google Scholar] [CrossRef] [Green Version]
- Lopez, V.M.; Decatur, C.L.; Stamer, W.D.; Lynch, R.M.; McKay, B.S. L-DOPA is an endogenous ligand for OA1. PLoS Biol. 2008, 6, e236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basnet, B.; Bhushan, A.; Khan, R.; Kumar, G.; Sharma, V.K.; Sharma, A.; Gupta, S. Plasma & urinary catecholamines & urinary vanillylmandelic acid levels in patients with generalized vitiligo. Indian J. Med. Res. 2018, 147, 384–390. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.S.; Yu, H.S.; Chang, H.R.; Yu, C.L.; Yu, C.L.; Wu, B.N. Cutaneous blood flow and adrenoceptor response increase in segmental-type vitiligo lesions. J. Dermatol. Sci. 2000, 23, 53–62. [Google Scholar] [CrossRef]
- Schallreuter, K.U.; Wood, J.M.; Pittelkow, M.R.; Buttner, G.; Swanson, N.; Korner, C.; Ehrke, C. Increased monoamine oxidase A activity in the epidermis of patients with vitiligo. Arch. Dermatol. Res. 1996, 288, 14–18. [Google Scholar] [CrossRef]
- Cucchi, M.L.; Frattini, P.; Santagostino, G.; Preda, S.; Orecchia, G. Catecholamines increase in the urine of non-segmental vitiligo especially during its active phase. Pigment Cell Res. 2003, 16, 111–116. [Google Scholar] [CrossRef]
- Schallreuter, K.U. Epidermal adrenergic signal transduction as part of the neuronal network in the human epidermis. J. Investig. Dermatol. Symp. Proc. 1997, 2, 37–40. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Zhou, W.; Liu, Y.; Wang, Y.; Li, Z.; Song, Z. Critical role of metabotropic glutamate receptor 4 in bone marrow-derived dendritic cells in the Th17 cell differentiation and the melanogenesis of B16 cells. Braz. J. Med. Biol. Res. 2020, 53, e9282. [Google Scholar] [CrossRef]
- Lin, X.; Meng, X.; Song, Z.; Lin, J. Nuclear factor erythroid 2-related factor 2 (Nrf2) as a potential therapeutic target for vitiligo. Arch. Biochem. Biophys. 2020, 696, 108670. [Google Scholar] [CrossRef]
- Natarajan, V.T.; Singh, A.; Kumar, A.A.; Sharma, P.; Kar, H.K.; Marrot, L.; Meunier, J.R.; Natarajan, K.; Rani, R.; Gokhale, R.S. Transcriptional upregulation of Nrf2-dependent phase II detoxification genes in the involved epidermis of vitiligo vulgaris. J. Investig. Dermatol. 2010, 130, 2781–2789. [Google Scholar] [CrossRef] [Green Version]
- Flores-Cantu, H.; Camara-Lemarroy, C.R.; Calderon-Hernandez, H.J.; Zapata-Rivera, M.A.; Villareal-Perez, J.Z.; Villareal-Velazquez, H.J. Anti-GAD antibody, seizures, cerebellar ataxias and vitiligo: A diagnostic challenge. Cerebellum 2015, 14, 375–377. [Google Scholar] [CrossRef]
- Jog, M.S.; Lambert, C.D.; Lang, A.E. Stiff-person syndrome. Can. J. Neurol. Sci. 1992, 19, 383–388. [Google Scholar] [CrossRef] [Green Version]
- Caballero, P.E.J. Stiff person syndrome: Presentation of a case with repetitive complex discharges in electromiograms. Neurologist 2009, 15, 227–229. [Google Scholar] [CrossRef]
- Burbridge, T.; Choudhury, B.; Collard, K. The uptake of gamma-aminobutyric acid and glutamate by synaptosomes from the visual cortex of albino and pigmented rabbits. J. Neural Transm. 1996, 103, 299–305. [Google Scholar] [CrossRef]
- Yoshida, M.; Hirotsu, S.; Nakahara, M.; Uchiwa, H.; Tomita, Y. Histamine is involved in ultraviolet B-induced pigmentation of guinea pig skin. J. Investig Dermatol. 2002, 118, 255–260. [Google Scholar] [CrossRef] [Green Version]
- Tomita, Y.; Maeda, K.; Tagami, H. Histamine stimulates normal human melanocytes in vitro: One of the possible inducers of hyperpigmentation in urticaria pigmentosa. J. Dermatol. Sci. 1993, 6, 146–154. [Google Scholar] [CrossRef]
- Liu, J.; Xu, Y.; Lin, T.-K.; Lv, C.; Elias, P.M.; Man, M.-Q. Topical histamine stimulates repigmentation of nonsegmental vitiligo by a receptor-dependent mechanism. Ski. Pharmacol. Physiol. 2017, 30, 139–145. [Google Scholar] [CrossRef] [Green Version]
- Bushby, S.; Green, A. The release of histamine by polymyxin B and polymyxin E. Br. J. Pharmacol. Chemother. 1955, 10, 215. [Google Scholar] [CrossRef]
- Mattos, K.; Cintra, M.; Gouvêa, I.; Ferreira, L.; Velho, P.; Moriel, P. Skin hyperpigmentation following intravenous polymyxin B treatment associated with melanocyte activation and inflammatory process. J. Clin. Pharm. Ther. 2017, 42, 573–578. [Google Scholar] [CrossRef]
- Panja, S.K.; Bhattacharya, B.; Lahiri, S.C. Role of histamine as a toxic mediator in the pathogenesis of vitiligo. Indian J. Dermatol. 2013, 58, 421. [Google Scholar] [CrossRef]
- Foldès, C.; Wallach, D.; Launay, J.-M.; Chirio, R. Congenital dyschromia with erythrocyte, platelet, and tryptophan metabolism abnormalities. J. Am. Acad. Dermatol. 1988, 19, 642–655. [Google Scholar] [CrossRef]
- Lekman, A.; Witt-Engerström, I.; Gottfries, J.; Hagberg, B.A.; Percy, A.K.; Svennerholm, L. Rett syndrome: Biogenic amines and metabolites in postmortem brain. Pediatric Neurol. 1989, 5, 357–362. [Google Scholar] [CrossRef]
- Gerritsen, S.; Akkerman, J.; Nijmeijer, B.; Sixma, J.; Witkop, C.; White, J. The Hermansky-Pudlak Syndrome: Evidence for a Lowered 5-Hydroxytryptamine Content in Platelets of Heterozygotes. Scand. J. Haematol. 1977, 18, 249–256. [Google Scholar] [CrossRef]
- Weiss, H.J.; Tschopp, T.B.; Rogers, J.; Brand, H. Studies of platelet 5-hydroxytryptamine (serotonin) in storage pool disease and albinism. J. Clin. Investig. 1974, 54, 421–432. [Google Scholar] [CrossRef]
- Kurbanov, K.; Beresov, T.T. Metabolism of tryptophan in vitiligo. Vopr. Meditsinskoj Khimii 1976, 22, 683. [Google Scholar]
- Cucchi, M.L.; Frattini, P.; Santagostino, G.; Orecchia, G. Higher plasma catecholamine and metabolite levels in the early phase of nonsegmental vitiligo. Pigment. Cell Res. 2000, 13, 28–32. [Google Scholar] [CrossRef]
- Lindsay, H. Leukoderma—its treatment. Calif. West. Med. 1932, 37, 36. [Google Scholar]
- Menon, A. Ultra-violet therapy in cases of leucoderma. Indian Med. Gaz. 1945, 80, 612. [Google Scholar]
- El Mofty, A.M. A preliminary clinical report on the treatment of leucodermia with Ammi majus Linn. J. Egypt. Med. Assoc. 1948, 31, 651–665. [Google Scholar]
- Iyengar, B. Photomodulation of the Melanocyte Cell cycle by Indoleamines. Neurosignals 1998, 7, 345–350. [Google Scholar] [CrossRef]
- Iyengar, B. Indoleamines and the UV-light-sensitive photoperiodic responses of the melanocyte network: A biological calendar? Experientia 1994, 50, 733–736. [Google Scholar] [CrossRef]
- Van Gool, A.R.; Fekkes, D.; Kruit, W.H.; Mulder, P.G.; Ten Hagen, T.L.; Bannink, M.; Maes, M.; Eggermont, A.M. Serum amino acids, biopterin and neopterin during long-term immunotherapy with interferon-alpha in high-risk melanoma patients. Psychiatry Res. 2003, 119, 125–132. [Google Scholar] [CrossRef]
- Hu, M.-f.; Li, Y.-l.; Zhuang, L. A case report of concomitant vitiligo in a patient treated with interferon alfa-1b for chronic hepatitis B infection. Chin. J. Hepatol. 2010, 18, 872. [Google Scholar]
- Hamadah, I.; Binamer, Y.; Sanai, F.M.; Abdo, A.A.; Alajlan, A. Interferon-induced vitiligo in hepatitis C patients: A case series. Int. J. Dermatol. 2010, 49, 829–833. [Google Scholar] [CrossRef] [PubMed]
- Kocer, B.; Nazliel, B.; Oztas, M.; Batur, H. Vitiligo and multiple sclerosis in a patient treated with interferon beta-1a: A case report. Eur. J. Neurol. 2009, 16, e78. [Google Scholar] [CrossRef]
- Namian, A.-M.; Shahbaz, S.; Salmanpoor, R.; Namazi, M.-R.; Dehghani, F.; Kamali-Sarvestani, E. Association of interferon-gamma and tumor necrosis factor alpha polymorphisms with susceptibility to vitiligo in Iranian patients. Arch. Dermatol. Res. 2009, 301, 21–25. [Google Scholar] [CrossRef]
- Anbar, T.; Abdel-Rahman, A.; Ahmad, H. Vitiligo occurring at site of interferon-a 2b injection in a patient with chronic viral hepatitis C: A case report. Clin. Exp. Dermatol. 2008, 33, 503. [Google Scholar] [CrossRef]
- Taffaro, M.; Pyrsopoulos, N.; Cedron, H.; Cacayorin, E.; Weppler, D.; Moon, J.; Nishida, S.; Levi, D.; Kato, T.; Selvaggi, G. Vitiligo improvement in a hepatitis C patient after treatment with PEG-interferon α-2A and ribavirin: A case report. Dig. Dis. Sci. 2007, 52, 3435. [Google Scholar] [CrossRef]
- Tinio, P.; Hadi, S.; Al-Ghaithi, K.; Al-Qari, H.; Rudikoff, D. Segmental Vitiligo and Hair Curling After Interferon α and Ribavirin Treatment for Hepatitis C. Skinmed 2006, 5, 50–51. [Google Scholar] [CrossRef]
- Seçkin, D.; Durusoy, Ç.; Şahin, S. Concomitant vitiligo and psoriasis in a patient treated with interferon alfa-2a for chronic hepatitis B infection. Pediatric Dermatol. 2004, 21, 577–579. [Google Scholar] [CrossRef]
- Primo, J.; Merino, C.; Belda, A.G. Vitiligo and alopecia in patients with chronic hepatitis C treated with alpha interferon associated or not with ribavirin. Gastroenterol. Hepatol. 2000, 23, 362–363. [Google Scholar]
- Nouri, K.; Busso, M.; Machler, B.C. Vitiligo associated with alpha-interferon in a patient with chronic active hepatitis C. Cutis 1997, 60, 289–290. [Google Scholar]
- Simsek, H.; Savas, C.; Akkiz, H.; Telatar, H. Interferon-lnduced Vitiligo in a Patient with Chronic Viral Hepatitis C Infection. Dermatology 1996, 193, 65–66. [Google Scholar] [CrossRef]
- Harris, J.; Bines, S.; Das Gupta, T. Therapy of disseminated malignant melanoma with recombinant α 2b-interferon and piroxicam: Clinical results with a report of an unusual response-associated feature (Vitiligo) and unusual toxicity (diffuse pulmonary interstitial fibrosis). Med. Pediatric Oncol. 1994, 22, 103–106. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Enkhtaivan, E.; Lee, C.H. Role of Amine Neurotransmitters and Their Receptors in Skin Pigmentation: Therapeutic Implication. Int. J. Mol. Sci. 2021, 22, 8071. https://doi.org/10.3390/ijms22158071
Enkhtaivan E, Lee CH. Role of Amine Neurotransmitters and Their Receptors in Skin Pigmentation: Therapeutic Implication. International Journal of Molecular Sciences. 2021; 22(15):8071. https://doi.org/10.3390/ijms22158071
Chicago/Turabian StyleEnkhtaivan, Enkhmend, and Chang Hoon Lee. 2021. "Role of Amine Neurotransmitters and Their Receptors in Skin Pigmentation: Therapeutic Implication" International Journal of Molecular Sciences 22, no. 15: 8071. https://doi.org/10.3390/ijms22158071
APA StyleEnkhtaivan, E., & Lee, C. H. (2021). Role of Amine Neurotransmitters and Their Receptors in Skin Pigmentation: Therapeutic Implication. International Journal of Molecular Sciences, 22(15), 8071. https://doi.org/10.3390/ijms22158071