
The Value of EphB2 Receptor and Cognate Ephrin Ligands in Prognostic and Predictive Assessments of Human Breast Cancer


Abstract
:1. Introduction
2. Results
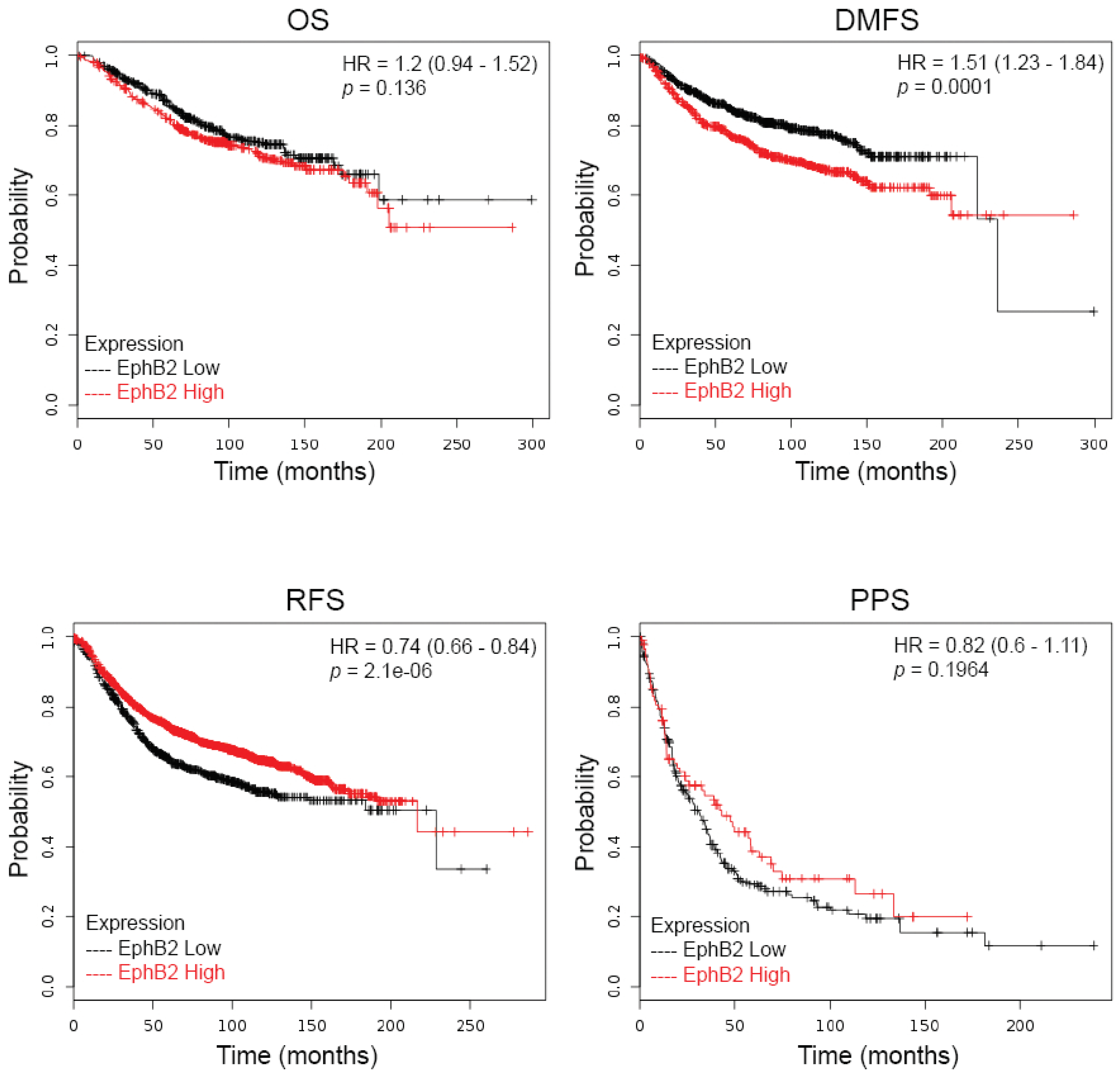
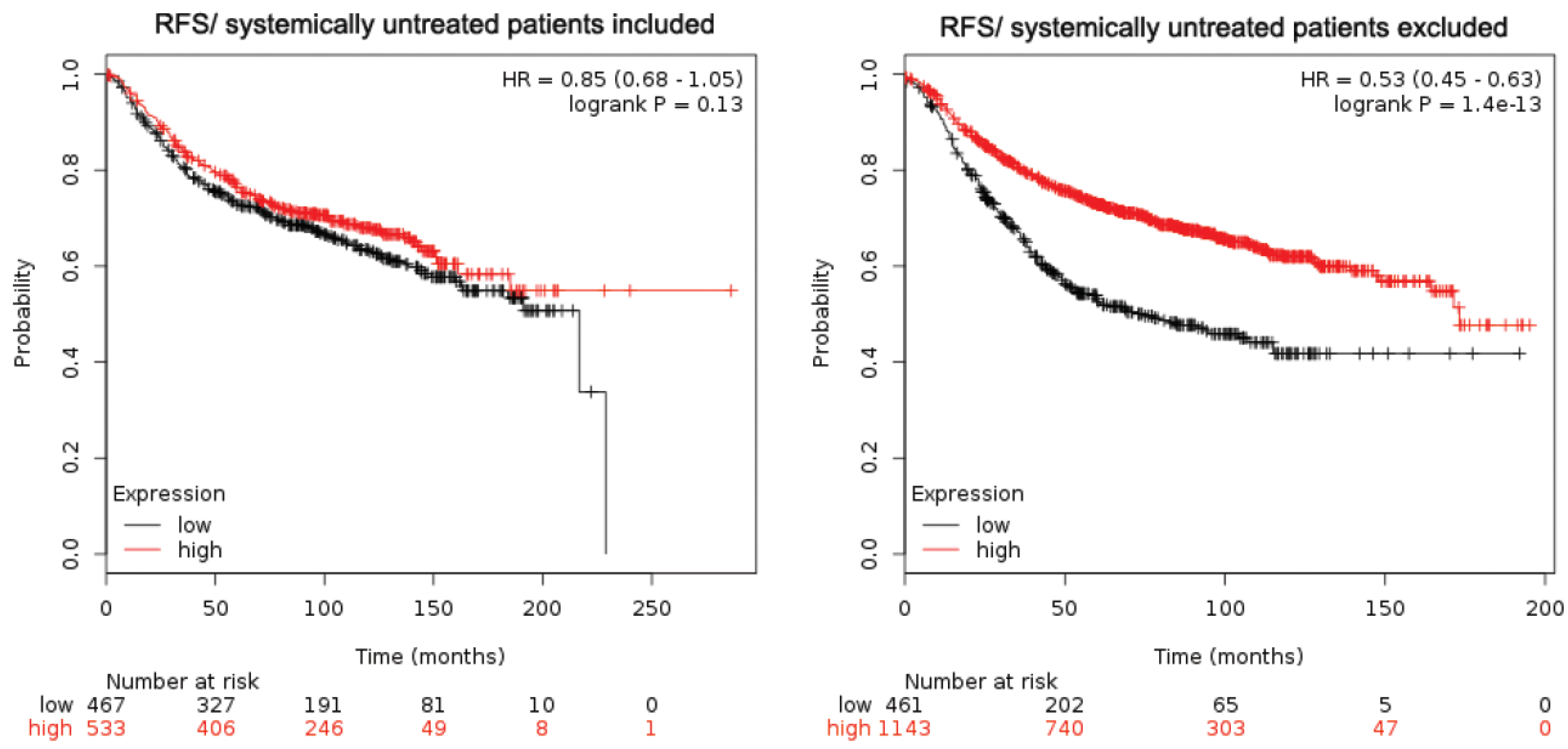
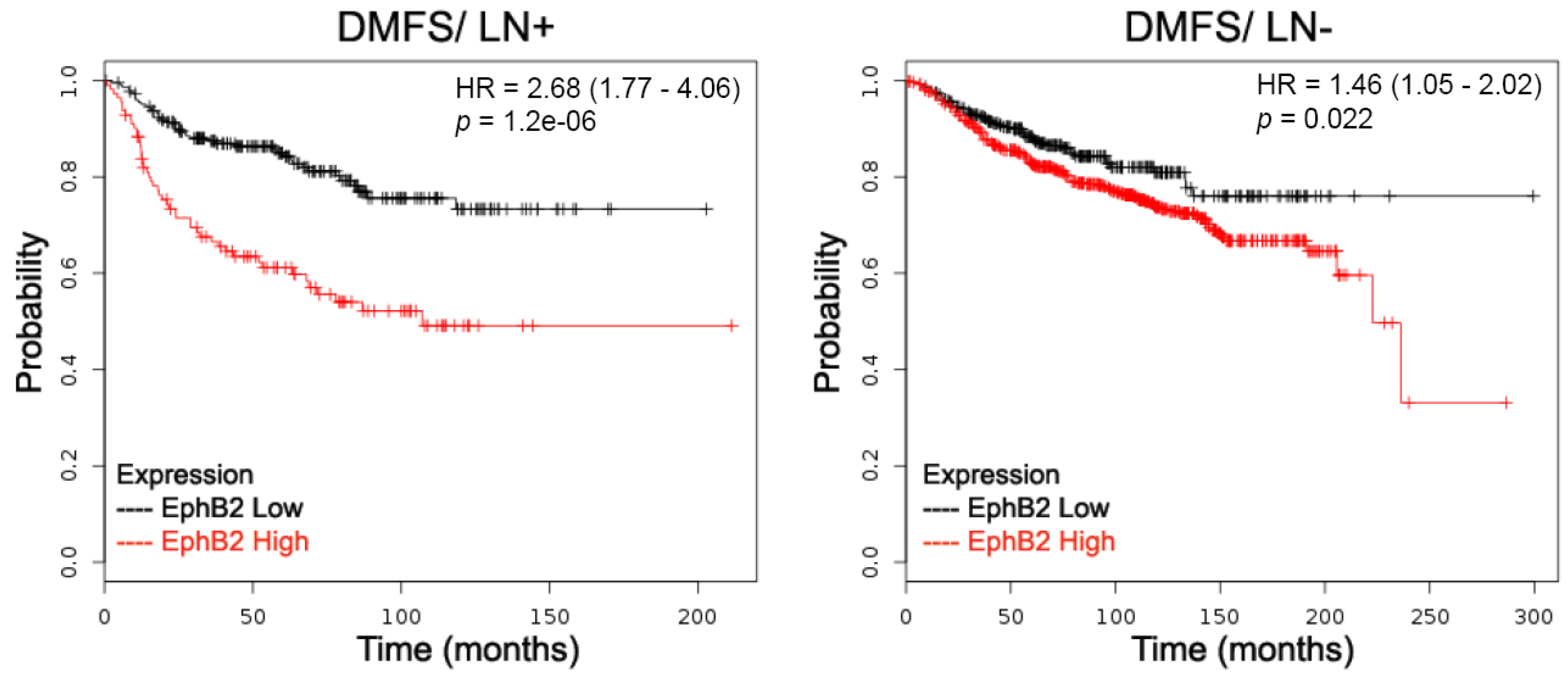
2.1. EphB2 Expression Is an Adverse Prognostic Factor for Distant Metastases and a Positive Predictive Factor for Post-Therapy Relapse
2.2. The Prognostic Value of EphB2 Is Stronger in the Luminal Subtypes of Breast Cancer
2.3. Correlations between Expression Levels of EphB2 Ligands and Patient Survival
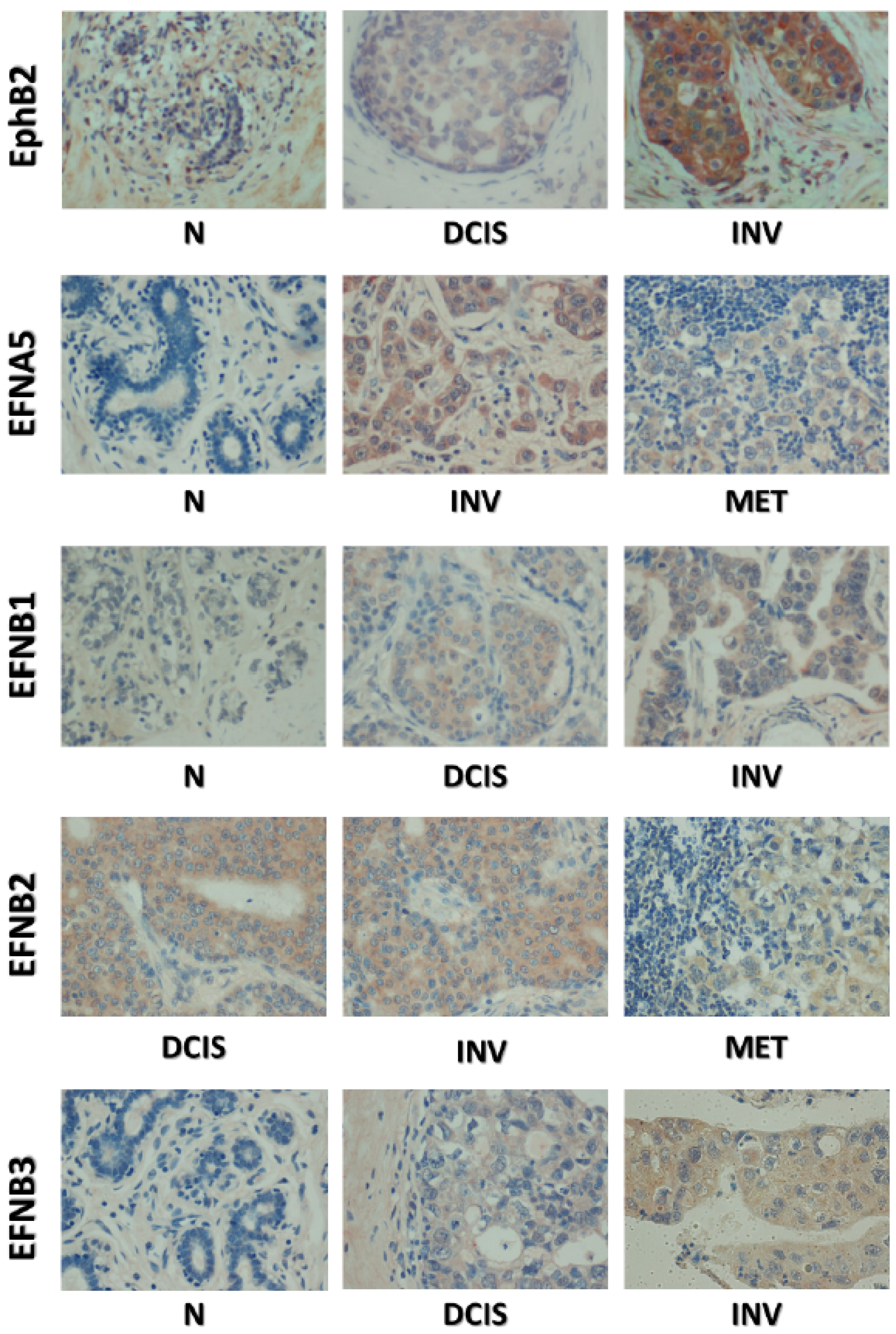
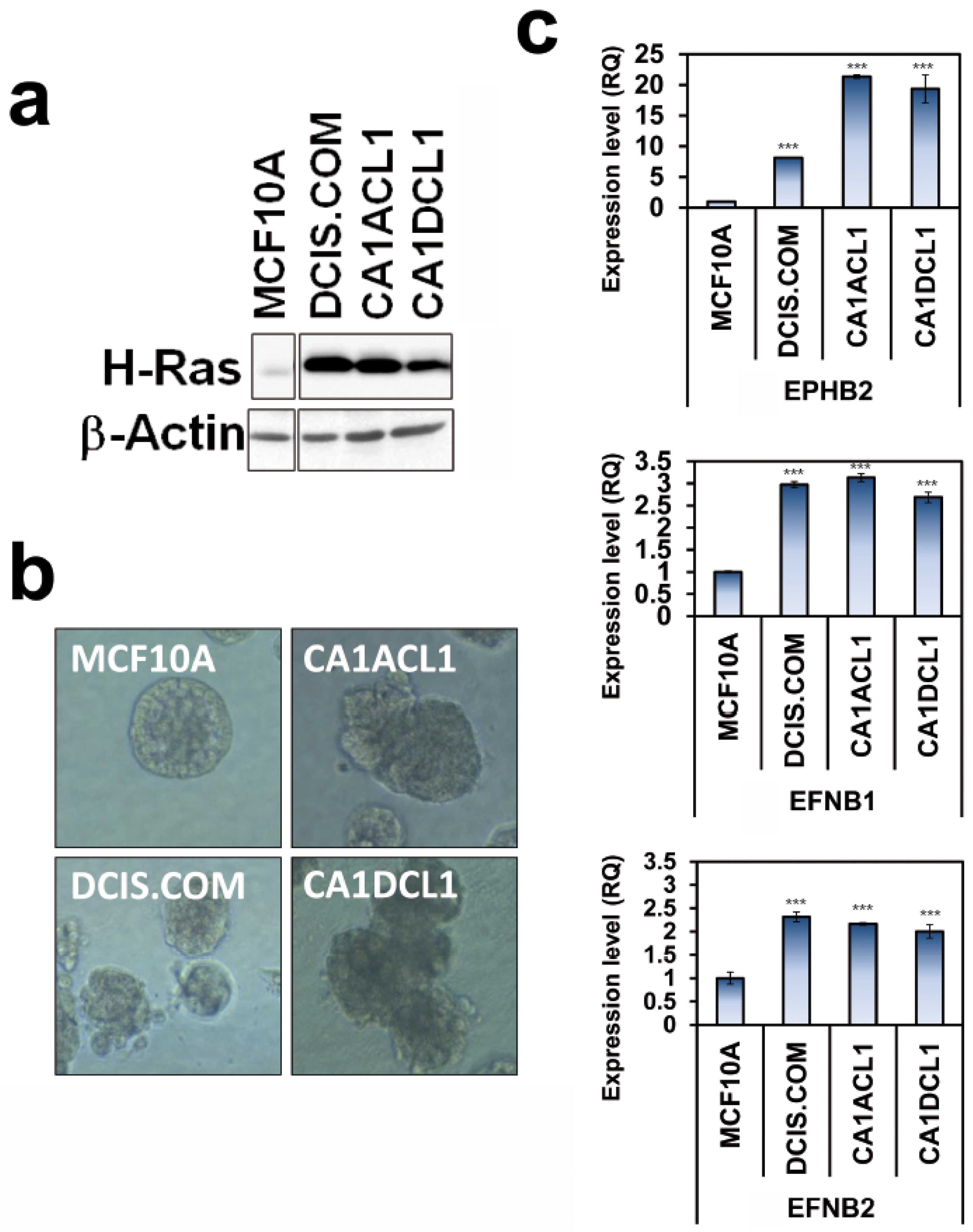
2.4. Co-Expression of EphB2 and Cognate Ligands Is Correlated with Breast Cancer Progression
3. Discussion
4. Materials and Methods
4.1. Kaplan–Meier Plot Analysis
4.2. Tissue Microarray and Immunohistochemistry
4.3. Cell Culture
4.4. Real-Time RT-PCR
4.5. Cell Lysate Preparation and Immunoblotting
4.6. Nomenclature
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| EphB2 | B-type Eph receptor 2 |
| EFN | ephrin |
| RTK | receptor tyrosine kinases |
| TMA | tissue microarray |
References
- Flanagan, J.G.; Vanderhaeghen, P. The ephrins and Eph receptors in neural development. Annu. Rev. Neurosci. 1998, 21, 309–345. [Google Scholar] [CrossRef] [Green Version]
- Frisen, J.; Holmberg, J.; Barbacid, M. Ephrins and their Eph receptors: Multitalented directors of embryonic development. EMBO J. 1999, 18, 5159–5165. [Google Scholar] [CrossRef] [Green Version]
- Holder, N.; Klein, R. Eph receptors and ephrins: Effectors of morphogenesis. Development 1999, 126, 2033–2044. [Google Scholar] [CrossRef]
- O’Leary, D.D.; Wilkinson, D.G. Eph receptors and ephrins in neural development. Curr. Opin. Neurobiol. 1999, 9, 65–73. [Google Scholar] [CrossRef]
- Yancopoulos, G.D.; Klagsbrun, M.; Folkman, J. Vasculogenesis, angiogenesis, and growth factors: Ephrins enter the fray at the border. Cell 1998, 93, 661–664. [Google Scholar] [CrossRef] [Green Version]
- Flenniken, A.M.; Gale, N.W.; Yancopoulos, G.D.; Wilkinson, D.G. Distinct and overlapping expression patterns of ligands for Eph-related receptor tyrosine kinases during mouse embryogenesis. Dev. Biol. 1996, 179, 382–401. [Google Scholar] [CrossRef] [Green Version]
- Friedman, G.C.; O’Leary, D.D. Eph receptor tyrosine kinases and their ligands in neural development. Curr. Opin. Neurobiol. 1996, 6, 127–133. [Google Scholar] [CrossRef]
- Wang, H.U.; Chen, Z.F.; Anderson, D.J. Molecular distinction and angiogenic interaction between embryonic arteries and veins revealed by ephrin-B2 and its receptor Eph-B4. Cell 1998, 93, 741–753. [Google Scholar] [CrossRef] [Green Version]
- Arvanitis, D.; Davy, A. Eph/ephrin signaling: Networks. Genes Dev. 2008, 22, 416–429. [Google Scholar] [CrossRef] [Green Version]
- Klein, R. Bidirectional modulation of synaptic functions by Eph/ephrin signaling. Nat. Neurosci. 2009, 12, 15–20. [Google Scholar] [CrossRef]
- Merlos-Suarez, A.; Batlle, E. Eph-ephrin signalling in adult tissues and cancer. Curr. Opin. Cell Biol. 2008, 20, 194–200. [Google Scholar] [CrossRef]
- Pasquale, E.B. Eph-ephrin bidirectional signaling in physiology and disease. Cell 2008, 133, 38–52. [Google Scholar] [CrossRef] [Green Version]
- Hruska, M.; Dalva, M.B. Ephrin regulation of synapse formation, function and plasticity. Mol. Cell Neurosci. 2012, 50, 35–44. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Fu, A.K.; Ip, N.Y. Eph receptors at synapses: Implications in neurodegenerative diseases. Cell Signal. 2012, 24, 606–611. [Google Scholar] [CrossRef]
- Gale, N.W.; Holland, S.J.; Valenzuela, D.M.; Flenniken, A.; Pan, L.; Ryan, T.E.; Henkemeyer, M.; Strebhardt, K.; Hirai, H.; Wilkinson, D.G.; et al. Eph receptors and ligands comprise two major specificity subclasses and are reciprocally compartmentalized during embryogenesis. Neuron 1996, 17, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Himanen, J.P.; Chumley, M.J.; Lackmann, M.; Li, C.; Barton, W.A.; Jeffrey, P.D.; Vearing, C.; Geleick, D.; Feldheim, D.A.; Boyd, A.W.; et al. Repelling class discrimination: Ephrin-A5 binds to and activates EphB2 receptor signaling. Nat. Neurosci. 2004, 7, 501–509. [Google Scholar] [CrossRef]
- Kandouz, M. The Eph/Ephrin family in cancer metastasis: Communication at the service of invasion. Cancer Metastasis Rev. 2012, 31, 353–373. [Google Scholar] [CrossRef]
- Dodelet, V.C.; Pasquale, E.B. Eph receptors and ephrin ligands: Embryogenesis to tumorigenesis. Oncogene 2000, 19, 5614–5619. [Google Scholar] [CrossRef] [Green Version]
- Lugli, A.; Spichtin, H.; Maurer, R.; Mirlacher, M.; Kiefer, J.; Huusko, P.; Azorsa, D.; Terracciano, L.; Sauter, G.; Kallioniemi, O.P.; et al. EphB2 expression across 138 human tumor types in a tissue microarray: High levels of expression in gastrointestinal cancers. Clin. Cancer Res. 2005, 11, 6450–6458. [Google Scholar] [CrossRef] [Green Version]
- Batlle, E.; Bacani, J.; Begthel, H.; Jonkheer, S.; Gregorieff, A.; van de Born, M.; Malats, N.; Sancho, E.; Boon, E.; Pawson, T.; et al. EphB receptor activity suppresses colorectal cancer progression. Nature 2005, 435, 1126–1130. [Google Scholar] [CrossRef]
- Huusko, P.; Ponciano-Jackson, D.; Wolf, M.; Kiefer, J.A.; Azorsa, D.O.; Tuzmen, S.; Weaver, D.; Robbins, C.; Moses, T.; Allinen, M.; et al. Nonsense-mediated decay microarray analysis identifies mutations of EPHB2 in human prostate cancer. Nat. Genet. 2004, 36, 979–983. [Google Scholar] [CrossRef]
- Kittles, R.A.; Baffoe-Bonnie, A.B.; Moses, T.Y.; Robbins, C.M.; Ahaghotu, C.; Huusko, P.; Pettaway, C.; Vijayakumar, S.; Bennett, J.; Hoke, G.; et al. A common nonsense mutation in EphB2 is associated with prostate cancer risk in African American men with a positive family history. J. Med. Genet. 2006, 43, 507–511. [Google Scholar] [CrossRef]
- Chukkapalli, S.; Amessou, M.; Dilly, A.K.; Dekhil, H.; Zhao, J.; Liu, Q.; Bejna, A.; Thomas, R.D.; Bandyopadhyay, S.; Bismar, T.A.; et al. Role of the EphB2 receptor in autophagy, apoptosis and invasion in human breast cancer cells. Exp. Cell Res. 2014, 320, 233–246. [Google Scholar] [CrossRef]
- Magic, Z.; Sandstrom, J.; Perez-Tenorio, G. EphrinB2 inhibits cell proliferation and motility in vitro and predicts longer metastasisfree survival in breast cancer. Int. J. Oncol. 2019, 55, 1275–1286. [Google Scholar]
- Rutkowski, R.; Mertens-Walker, I.; Lisle, J.E.; Herington, A.C.; Stephenson, S.A. Evidence for a dual function of EphB4 as tumor promoter and suppressor regulated by the absence or presence of the ephrin-B2 ligand. Int. J. Cancer 2012, 131, E614–E624. [Google Scholar] [CrossRef]
- Yin, H.; Lu, C.; Tang, Y.; Wang, H.; Wang, H.; Wang, J. Enhanced expression of EphrinB1 is associated with lymph node metastasis and poor prognosis in breast cancer. Cancer Biomark. 2013, 13, 261–267. [Google Scholar] [CrossRef]
- Dun, M.D.; Chalkley, R.J.; Faulkner, S.; Keene, S.; Avery-Kiejda, K.A.; Scott, R.J.; Falkenby, L.G.; Cairns, M.J.; Larsen, M.R.; Bradshaw, R.A.; et al. Proteotranscriptomic profiling of 231-BR breast cancer cells: Identification of potential biomarkers and therapeutic targets for brain metastasis. Mol. Cell Proteom. 2015, 14, 2316–2330. [Google Scholar] [CrossRef] [Green Version]
- Gyorffy, B.; Lanczky, A.; Eklund, A.C.; Denkert, C.; Budczies, J.; Li, Q.; Szallasi, Z. An online survival analysis tool to rapidly assess the effect of 22,277 genes on breast cancer prognosis using microarray data of 1809 patients. Breast Cancer Res. Treat. 2010, 123, 725–731. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Birkbak, N.J.; Gyorffy, B.; Szallasi, Z.; Eklund, A.C. Jetset: Selecting the optimal microarray probe set to represent a gene. BMC Bioinform. 2011, 12, 474. [Google Scholar] [CrossRef] [Green Version]
- Perou, C.M.; Sorlie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular portraits of human breast tumours. Nature 2000, 406, 747–752. [Google Scholar] [CrossRef]
- Sorlie, T.; Perou, C.M.; Tibshirani, R.; Aas, T.; Geisler, S.; Johnsen, H.; Hastie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; et al. Gene expression patterns of breast carcinomas distinguish tumor subclasses with clinical implications. Proc. Natl. Acad. Sci. USA 2001, 98, 10869–10874. [Google Scholar] [CrossRef] [Green Version]
- Sorlie, T.; Wang, Y.; Xiao, C.; Johnsen, H.; Naume, B.; Samaha, R.R.; Borresen-Dale, A.L. Distinct molecular mechanisms underlying clinically relevant subtypes of breast cancer: Gene expression analyses across three different platforms. BMC Genom. 2006, 7, 127. [Google Scholar] [CrossRef] [Green Version]
- Perou, C.M.; Borresen-Dale, A.L. Systems biology and genomics of breast cancer. Cold Spring Harb. Perspect. Biol. 2011, 3, a003293. [Google Scholar] [CrossRef] [Green Version]
- Pasquale, E.B. The Eph family of receptors. Curr. Opin. Cell Biol. 1997, 9, 608–615. [Google Scholar] [CrossRef]
- Himanen, J.P.; Rajashankar, K.R.; Lackmann, M.; Cowan, C.A.; Henkemeyer, M.; Nikolov, D.B. Crystal structure of an Eph receptor-ephrin complex. Nature 2001, 414, 933–938. [Google Scholar] [CrossRef] [PubMed]
- Gale, N.W.; Flenniken, A.; Compton, D.C.; Jenkins, N.; Copeland, N.G.; Gilbert, D.J.; Davis, S.; Wilkinson, D.G.; Yancopoulos, G.D. Elk-L3, a novel transmembrane ligand for the Eph family of receptor tyrosine kinases, expressed in embryonic floor plate, roof plate and hindbrain segments. Oncogene 1996, 13, 1343–1352. [Google Scholar]
- Brambilla, R.; Schnapp, A.; Casagranda, F.; Labrador, J.P.; Bergemann, A.D.; Flanagan, J.G.; Pasquale, E.B.; Klein, R. Membrane-bound LERK2 ligand can signal through three different Eph-related receptor tyrosine kinases. EMBO J. 1995, 14, 3116–3126. [Google Scholar] [CrossRef]
- Dawson, P.J.; Wolman, S.R.; Tait, L.; Heppner, G.H.; Miller, F.R. MCF10AT: A model for the evolution of cancer from proliferative breast disease. Am. J. Pathol. 1996, 148, 313–319. [Google Scholar]
- Miller, F.R.; Soule, H.D.; Tait, L.; Pauley, R.J.; Wolman, S.R.; Dawson, P.J.; Heppner, G.H. Xenograft model of progressive human proliferative breast disease. J. Natl. Cancer Inst. 1993, 85, 1725–1732. [Google Scholar] [CrossRef] [PubMed]
- Miller, F.R.; Santner, S.J.; Tait, L.; Dawson, P.J. MCF10DCIS.com xenograft model of human comedo ductal carcinoma in situ. J. Natl. Cancer Inst. 2000, 92, 1185–1186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, F.R. Xenograft models of premalignant breast disease. J. Mammary Gland Biol. Neoplasia 2000, 5, 379–391. [Google Scholar] [CrossRef]
- Strickland, L.B.; Dawson, P.J.; Santner, S.J.; Miller, F.R. Progression of premalignant MCF10AT generates heterogeneous malignant variants with characteristic histologic types and immunohistochemical markers. Breast Cancer Res. Treat. 2000, 64, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Karagiannis, G.S.; Condeelis, J.S.; Oktay, M.H. Chemotherapy-induced metastasis in breast cancer. Oncotarget 2017, 8, 110733–110734. [Google Scholar] [CrossRef]
- Karagiannis, G.S.; Condeelis, J.S.; Oktay, M.H. Chemotherapy-induced metastasis: Mechanisms and translational opportunities. Clin. Exp. Metastasis 2018, 35, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Kandouz, M.; Haidara, K.; Zhao, J.; Brisson, M.L.; Batist, G. The EphB2 tumor suppressor induces autophagic cell death via concomitant activation of the ERK1/2 and PI3K pathways. Cell Cycle 2010, 9, 398–407. [Google Scholar] [CrossRef] [Green Version]
- Baehrecke, E.H. Autophagy: Dual roles in life and death? Nat. Rev. Mol. Cell Biol. 2005, 6, 505–510. [Google Scholar] [CrossRef]
- Kroemer, G.; Levine, B. Autophagic cell death: The story of a misnomer. Nat. Rev. Mol. Cell Biol. 2008, 9, 1004–1010. [Google Scholar] [CrossRef]
- Kimmelman, A.C. The dynamic nature of autophagy in cancer. Genes Dev. 2011, 25, 1999–2010. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.K.; Coffelt, S.B.; Cho, C.H.; Wang, X.J.; Lee, C.W.; Chan, F.K.; Yu, J.; Sung, J.J. The autophagic paradox in cancer therapy. Oncogene 2012, 31, 939–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalby, K.N.; Tekedereli, I.; Lopez-Berestein, G.; Ozpolat, B. Targeting the prodeath and prosurvival functions of autophagy as novel therapeutic strategies in cancer. Autophagy 2010, 6, 322–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debnath, J. The multifaceted roles of autophagy in tumors-implications for breast cancer. J. Mammary Gland Biol. Neoplasia 2011, 16, 173–187. [Google Scholar] [CrossRef] [Green Version]
- Fimia, G.M.; Piacentini, M. Regulation of autophagy in mammals and its interplay with apoptosis. Cell Mol. Life Sci. 2010, 67, 1581–1588. [Google Scholar] [CrossRef]
- Scarlatti, F.; Granata, R.; Meijer, A.J.; Codogno, P. Does autophagy have a license to kill mammalian cells? Cell Death Differ. 2009, 16, 12–20. [Google Scholar] [CrossRef] [Green Version]
- Rikiishi, H. Novel insights into the interplay between apoptosis and autophagy. Int. J. Cell Biol. 2012, 2012, 317645. [Google Scholar] [CrossRef] [Green Version]
- Espina, V.; Mariani, B.D.; Gallagher, R.I.; Tran, K.; Banks, S.; Wiedemann, J.; Huryk, H.; Mueller, C.; Adamo, L.; Deng, J.; et al. Malignant precursor cells pre-exist in human breast DCIS and require autophagy for survival. PLoS ONE 2010, 5, e10240. [Google Scholar] [CrossRef]
- Zhang, H.; Cui, Z.; Cheng, D.; Du, Y.; Guo, X.; Gao, R.; Chen, J.; Sun, W.; He, R.; Ma, X.; et al. RNF186 regulates EFNB1 (ephrin B1)-EPHB2-induced autophagy in the colonic epithelial cells for the maintenance of intestinal homeostasis. Autophagy 2020, 1–18. [Google Scholar] [CrossRef]
- Jubb, A.M.; Zhong, F.; Bheddah, S.; Grabsch, H.I.; Frantz, G.D.; Mueller, W.; Kavi, V.; Quirke, P.; Polakis, P.; Koeppen, H. EphB2 is a prognostic factor in colorectal cancer. Clin. Cancer Res. 2005, 11, 5181–5187. [Google Scholar] [CrossRef] [Green Version]
- Guo, D.L.; Zhang, J.; Yuen, S.T.; Tsui, W.Y.; Chan, A.S.; Ho, C.; Ji, J.; Leung, S.Y.; Chen, X. Reduced expression of EphB2 that parallels invasion and metastasis in colorectal tumours. Carcinogenesis 2006, 27, 454–464. [Google Scholar] [CrossRef]
- Nakada, M.; Niska, J.A.; Miyamori, H.; McDonough, W.S.; Wu, J.; Sato, H.; Berens, M.E. The phosphorylation of EphB2 receptor regulates migration and invasion of human glioma cells. Cancer Res. 2004, 64, 3179–3185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakada, M.; Niska, J.A.; Tran, N.L.; McDonough, W.S.; Berens, M.E. EphB2/R-Ras signaling regulates glioma cell adhesion, growth.; invasion. Am. J. Pathol. 2005, 167, 565–576. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.D.; Rath, P.; Lal, B.; Richard, J.P.; Li, Y.; Goodwin, C.R.; Laterra, J.; Xia, S. EphB2 receptor controls proliferation/migration dichotomy of glioblastoma by interacting with focal adhesion kinase. Oncogene 2012, 31, 5132–5143. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Sasaki, K.; Kamata, R.; Sakai, R. The C-terminus of ephrin-B1 regulates metalloproteinase secretion and invasion of cancer cells. J. Cell Sci. 2007, 120, 2179–2189. [Google Scholar] [CrossRef] [Green Version]
- Nakada, M.; Drake, K.L.; Nakada, S.; Niska, J.A.; Berens, M.E. Ephrin-B3 ligand promotes glioma invasion through activation of Rac1. Cancer Res. 2006, 66, 8492–8500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, H.J.; Hwang, Y.S.; Yoon, J.; Lee, M.; Lee, H.G.; Daar, I.O. EphrinB1 promotes cancer cell migration and invasion through the interaction with RhoGDI1. Oncogene 2018, 37, 861–872. [Google Scholar] [CrossRef] [Green Version]
- Sato, S.; Vasaikar, S.; Eskaros, A.; Kim, Y.; Lewis, J.S.; Zhang, B.; Zijlstra, A.; Weaver, A.M. EPHB2 carried on small extracellular vesicles induces tumor angiogenesis via activation of ephrin reverse signaling. JCI Insight 2019, 4, e132447. [Google Scholar] [CrossRef]
- Brantley-Sieders, D.M.; Jiang, A.; Sarma, K.; Badu-Nkansah, A.; Walter, D.L.; Shyr, Y.; Chen, J. Eph/ephrin profiling in human breast cancer reveals significant associations between expression level and clinical outcome. PLoS ONE 2011, 6, e24426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brantley-Sieders, D.M.; Zhuang, G.; Hicks, D.; Fang, W.B.; Hwang, Y.; Cates, J.M.; Coffman, K.; Jackson, D.; Bruckheimer, E.; Muraoka-Cook, R.S.; et al. The receptor tyrosine kinase EphA2 promotes mammary adenocarcinoma tumorigenesis and metastatic progression in mice by amplifying ErbB2 signaling. J. Clin. Investig. 2008, 118, 64–78. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.B.; Brantley-Sieders, D.M.; Parker, M.A.; Reith, A.D.; Chen, J. A kinase-dependent role for EphA2 receptor in promoting tumor growth and metastasis. Oncogene 2005, 24, 7859–7868. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.; Miller, K.D.; Gokmen-Polar, Y.; Jeng, M.H.; Kinch, M.S. EphA2 overexpression decreases estrogen dependence and tamoxifen sensitivity. Cancer Res. 2003, 63, 3425–3429. [Google Scholar]
- Zelinski, D.P.; Zantek, N.D.; Stewart, J.C.; Irizarry, A.R.; Kinch, M.S. EphA2 overexpression causes tumorigenesis of mammary epithelial cells. Cancer Res. 2001, 61, 2301–2306. [Google Scholar]
- Truitt, L.; Freywald, T.; DeCoteau, J.; Sharfe, N.; Freywald, A. The EphB6 receptor cooperates with c-Cbl to regulate the behavior of breast cancer cells. Cancer Res. 2010, 70, 1141–1153. [Google Scholar] [CrossRef] [Green Version]
- Noren, N.K.; Foos, G.; Hauser, C.A.; Pasquale, E.B. The EphB4 receptor suppresses breast cancer cell tumorigenicity through an Abl-Crk pathway. Nat. Cell Biol. 2006, 8, 815–825. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.R.; Singh, J.; Xia, G.; Krasnoperov, V.; Hassanieh, L.; Ley, E.J.; Scehnet, J.; Kumar, N.G.; Hawes, D.; Press, M.F.; et al. Receptor tyrosine kinase EphB4 is a survival factor in breast cancer. Am. J. Pathol. 2006, 169, 279–293. [Google Scholar] [CrossRef] [Green Version]
- Noren, N.K.; Lu, M.; Freeman, A.L.; Koolpe, M.; Pasquale, E.B. Interplay between EphB4 on tumor cells and vascular ephrin-B2 regulates tumor growth. Proc. Natl. Acad. Sci. USA 2004, 101, 5583–5588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berclaz, G.; Flutsch, B.; Altermatt, H.J.; Rohrbach, V.; Djonov, V.; Ziemiecki, A.; Dreher, E.; Andres, A.C. Loss of EphB4 receptor tyrosine kinase protein expression during carcinogenesis of the human breast. Oncol. Rep. 2002, 9, 985–989. [Google Scholar] [CrossRef]
- Harburg, G.C.; Hinck, L. Navigating breast cancer: Axon guidance molecules as breast cancer tumor suppressors and oncogenes. J. Mammary Gland Biol. Neoplasia 2011, 16, 257–270. [Google Scholar] [CrossRef] [Green Version]
- Bhushan, L.; Kandpal, R.P. EphB6 receptor modulates micro RNA profile of breast carcinoma cells. PLoS ONE 2011, 6, e22484. [Google Scholar] [CrossRef] [Green Version]
- Hiramoto-Yamaki, N.; Takeuchi, S.; Ueda, S.; Harada, K.; Fujimoto, S.; Negishi, M.; Katoh, H. Ephexin4 and EphA2 mediate cell migration through a RhoG-dependent mechanism. J. Cell Biol. 2010, 190, 461–477. [Google Scholar] [CrossRef] [Green Version]
- Kaenel, P.; Mosimann, M.; Andres, A.C. The multifaceted roles of Eph-ephrin signaling in breast cancer. Cell Adh. Migr. 2012, 6, 138–147. [Google Scholar] [CrossRef] [Green Version]
- Garber, K. Of Ephs and ephrins: Companies target guidance molecules in cancer. J. Natl. Cancer Inst. 2010, 102, 1692–1694. [Google Scholar] [CrossRef] [PubMed]
- Nievergall, E.; Saunders, T.; Lackmann, M. Targeting of EPH receptor tyrosine kinases for anticancer therapy. Crit. Rev. Oncog. 2012, 17, 211–232. [Google Scholar] [CrossRef]
- Bacus, S.; Flowers, J.L.; Press, M.F.; Bacus, J.W.; McCarty, K.S., Jr. The evaluation of estrogen receptor in primary breast carcinoma by computer-assisted image analysis. Am. J. Clin. Pathol. 1988, 90, 233–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McClelland, R.A.; Berger, U.; Miller, L.S.; Powles, T.J.; Jensen, E.V.; Coombes, R.C. Immunocytochemical assay for estrogen receptor: Relationship to outcome of therapy in patients with advanced breast cancer. Cancer Res. 1986, 46, 4241s–4243s. [Google Scholar] [PubMed]
- Soule, H.D.; Maloney, T.M.; Wolman, S.R.; Peterson, W.D., Jr.; Brenz, R.; McGrath, C.M.; Russo, J.; Pauley, R.J.; Jones, R.F.; Brooks, S.C. Isolation and characterization of a spontaneously immortalized human breast epithelial cell line, MCF-10. Cancer Res. 1990, 50, 6075–6086. [Google Scholar]
- Debnath, J.; Muthuswamy, S.K.; Brugge, J.S. Morphogenesis and oncogenesis of MCF-10A mammary epithelial acini grown in three-dimensional basement membrane cultures. Methods 2003, 30, 256–268. [Google Scholar] [CrossRef]
- Eph Nomenclature Committee. Unified nomenclature for Eph family receptors and their ligands, the ephrins. Cell 1997, 90, 403–404. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Number of Patients | EPHB2 | EFNB1 | EFNB2 | EFNB3 | EFNA5 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HR | p-Value | HR | p-Value | HR | p-Value | HR | p-Value | HR | p-Value | ||||
| RFS | All | All | 3554 | 0.74 | 2.1× 10−6 | 0.77 | 1.5× 10−5 | 1.28 | 3.5× 10−5 | 0.9 | 0.091 | 0.72 | 1.7× 10−8 |
| Lymph Node Status | LN+ | 945 | 1.34 | 0.011 | 0.71 | 0.0059 | 1.29 | 0.024 | 0.83 | 0.15 | 1.09 | 0.43 | |
| LN- | 1813 | 1.16 | 0.1505 | 0.89 | 0.21 | 1.24 | 0.026 | 1.22 | 0.03 | 0.91 | 0.36 | ||
| Intrinsic Subtype | Basal | 580 | 0.69 | 0.0049 | 0.62 | 0.00022 | 1.21 | 0.19 | 1.47 | 0.0071 | 0.68 | 0.0036 | |
| Luminal A | 1764 | 0.7 | 0.00011 | 0.76 | 0.0027 | 1.25 | 0.019 | 1.14 | 0.16 | 0.57 | 3.6× 10−9 | ||
| Luminal B | 1002 | 0.71 | 0.0034 | 0.81 | 0.06 | 1.23 | 0.048 | 0.8 | 0.031 | 0.68 | 0.00022 | ||
| HER2+ | 208 | 0.5 | 0.0018 | 0.69 | 0.083 | 1.75 | 0.0094 | 1.37 | 0.14 | 0.67 | 0.062 | ||
| OS | All | All | 1117 | 1.2 | 0.14 | 0.75 | 0.019 | 1.26 | 0.065 | 0.83 | 0.14 | 0.84 | 0.23 |
| Lymph Node Status | LN+ | 197 | 1.71 | 0.035 | 0.78 | 0.33 | 0.7 | 0.22 | 1.47 | 0.14 | 1.67 | 0.059 | |
| LN- | 425 | 0.65 | 0.044 | 0.52 | 0.0035 | 1.51 | 0.073 | 0.72 | 0.14 | 0.6 | 0.04 | ||
| Intrinsic Subtype | Basal | 204 | 0.47 | 0.031 | 0.63 | 0.1 | 0.75 | 0.31 | 2.05 | 0.01 | 0.69 | 0.2 | |
| Luminal A | 504 | 1.38 | 0.096 | 0.66 | 0.072 | 1.59 | 0.031 | 1.27 | 0.22 | 0.67 | 0.1 | ||
| Luminal B | 320 | 1.48 | 0.082 | 0.62 | 0.026 | 1.41 | 0.19 | 0.7 | 0.097 | 1.49 | 0.074 | ||
| HER2+ | 89 | 0.67 | 0.31 | 1.65 | 0.2 | 1.57 | 0.24 | 1.65 | 0.31 | 2.37 | 0.1 | ||
| DMFS | All | All | 1609 | 1.51 | 5.6× 10−5 | 1.25 | 0.038 | 1.43 | 0.00058 | 1.21 | 0.061 | 0.86 | 0.19 |
| Lymph Node Status | LN+ | 337 | 2.68 | 1.2× 10−6 | 1.4 | 0.11 | 1.44 | 0.091 | 1.53 | 0.046 | 1.5 | 0.054 | |
| LN- | 896 | 1.46 | 0.022 | 1.32 | 0.07 | 1.39 | 0.035 | 1.32 | 0.063 | 0.86 | 0.3 | ||
| Intrinsic Subtype | Basal | 219 | 1.52 | 0.14 | 0.71 | 0.19 | 1.34 | 0.29 | 1.67 | 0.058 | 1.29 | 0.42 | |
| Luminal A | 918 | 1.5 | 0.0068 | 1.36 | 0.058 | 1.55 | 0.0061 | 1.31 | 0.074 | 0.79 | 0.15 | ||
| Luminal B | 361 | 2.15 | 0.00016 | 1.55 | 0.024 | 1.34 | 0.13 | 1.24 | 0.26 | 1.31 | 0.16 | ||
| HER2+ | 111 | 1.1424 | 0.14 | 1.77 | 0.082 | 4.58 | 0.0055 | 2.37 | 0.017 | 2.35 | 0.068 | ||
| EphB2 | EphrinB1 | EphrinB2 | EphrinB3 | EphrinA5 | |
|---|---|---|---|---|---|
| Normal # | 0 (0–0), reference | 0 (0–0), reference | 0 (0–0), reference | 0 (0–0), reference | 0 (0–0), reference |
| DCIS | 80 (20–170), *** | 5 (0–80), *** | 45 (15–58), *** | 10 (0–30), *** | 0 (0–20), *** |
| Ductal Invasive | 35 (0–120), *** | 70 (30–140), *** | 140 (30–180), *** | 10 (0–90), *** | 40 (10–140), *** |
| Metastasis * | 90 (30–190), *** | 65 (25–120), *** | 160 (60–270), *** | 35 (10–150), *** | 50 (0–120), *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ebrahim, A.S.; Hailat, Z.; Bandyopadhyay, S.; Neill, D.; Kandouz, M. The Value of EphB2 Receptor and Cognate Ephrin Ligands in Prognostic and Predictive Assessments of Human Breast Cancer. Int. J. Mol. Sci. 2021, 22, 8098. https://doi.org/10.3390/ijms22158098
Ebrahim AS, Hailat Z, Bandyopadhyay S, Neill D, Kandouz M. The Value of EphB2 Receptor and Cognate Ephrin Ligands in Prognostic and Predictive Assessments of Human Breast Cancer. International Journal of Molecular Sciences. 2021; 22(15):8098. https://doi.org/10.3390/ijms22158098
Chicago/Turabian StyleEbrahim, Abdul Shukkur, Zeyad Hailat, Sudeshna Bandyopadhyay, Daniel Neill, and Mustapha Kandouz. 2021. "The Value of EphB2 Receptor and Cognate Ephrin Ligands in Prognostic and Predictive Assessments of Human Breast Cancer" International Journal of Molecular Sciences 22, no. 15: 8098. https://doi.org/10.3390/ijms22158098
APA StyleEbrahim, A. S., Hailat, Z., Bandyopadhyay, S., Neill, D., & Kandouz, M. (2021). The Value of EphB2 Receptor and Cognate Ephrin Ligands in Prognostic and Predictive Assessments of Human Breast Cancer. International Journal of Molecular Sciences, 22(15), 8098. https://doi.org/10.3390/ijms22158098