
Keratin Profiling by Single-Cell RNA-Sequencing Identifies Human Prostate Stem Cell Lineage Hierarchy and Cancer Stem-Like Cells

 ,
,
Abstract
:1. Introduction
2. Results
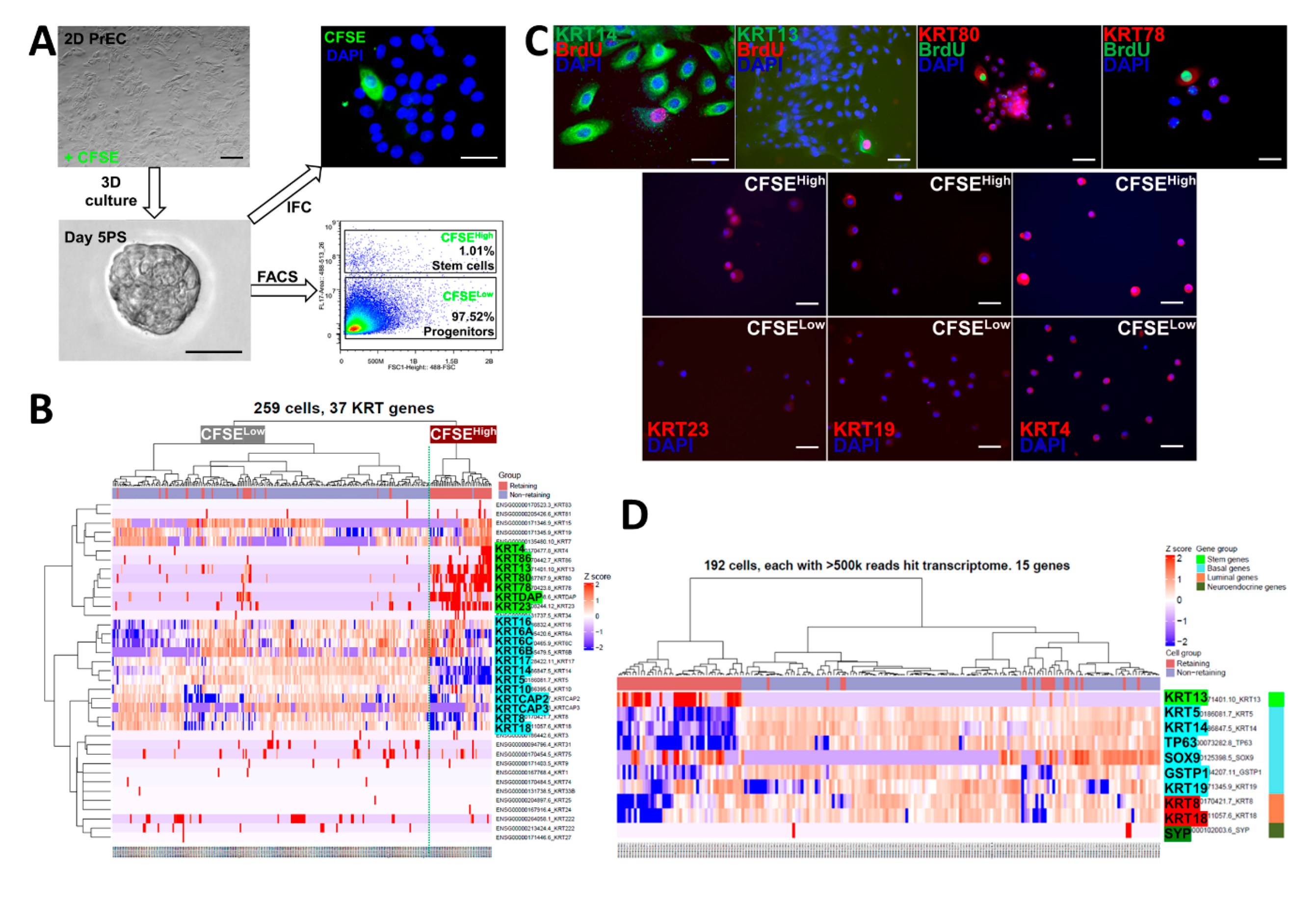
2.1. Differentially Expressed Keratin Genes in Prostate Stem-Like and Progenitor Cells by Fluidigm Single Cell RNA-Seq
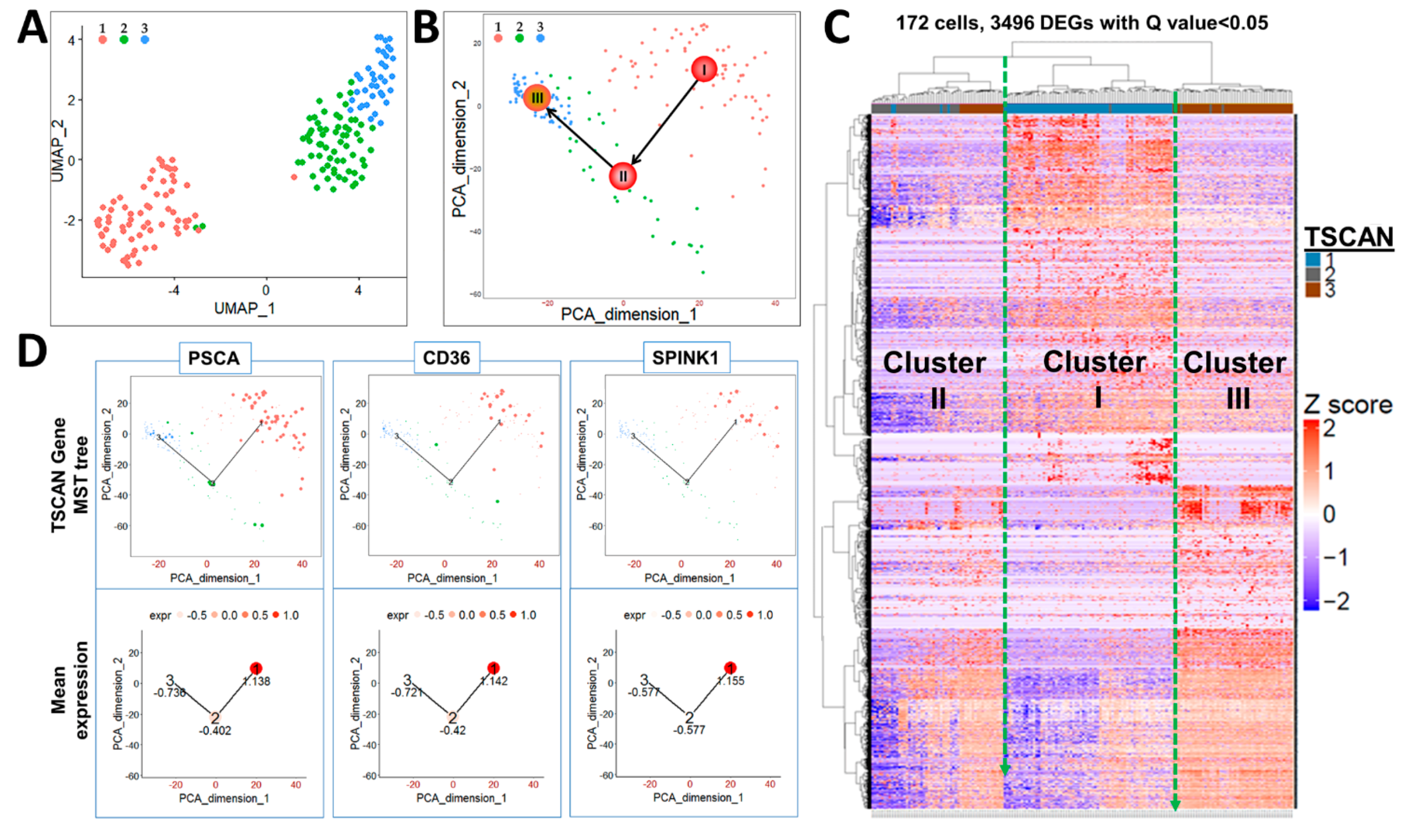
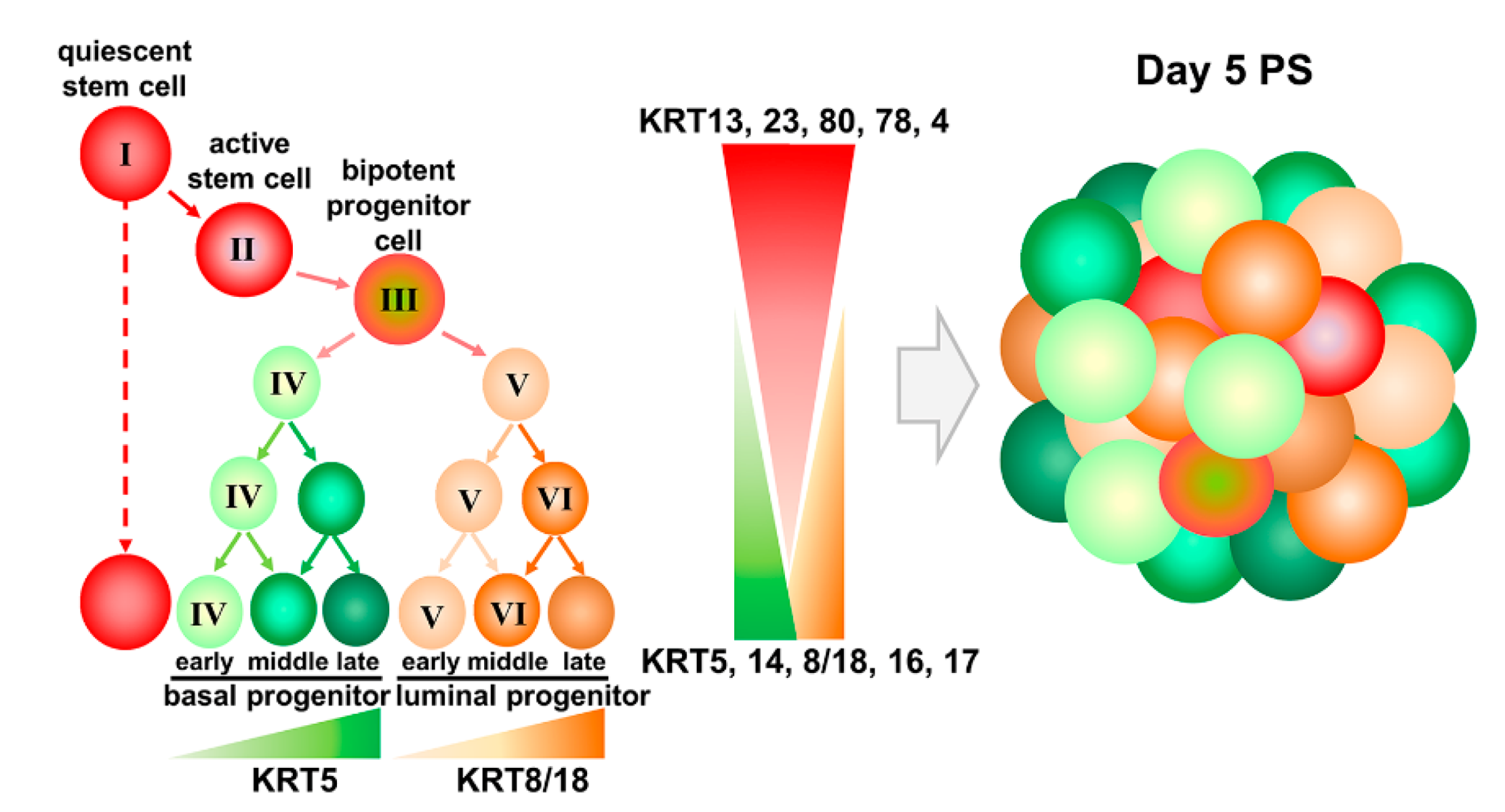
2.2. Identification of Three Subpopulations in Prostasphere CFSEHigh Prostate Stem-Like Cells by Single-Cell Analysis
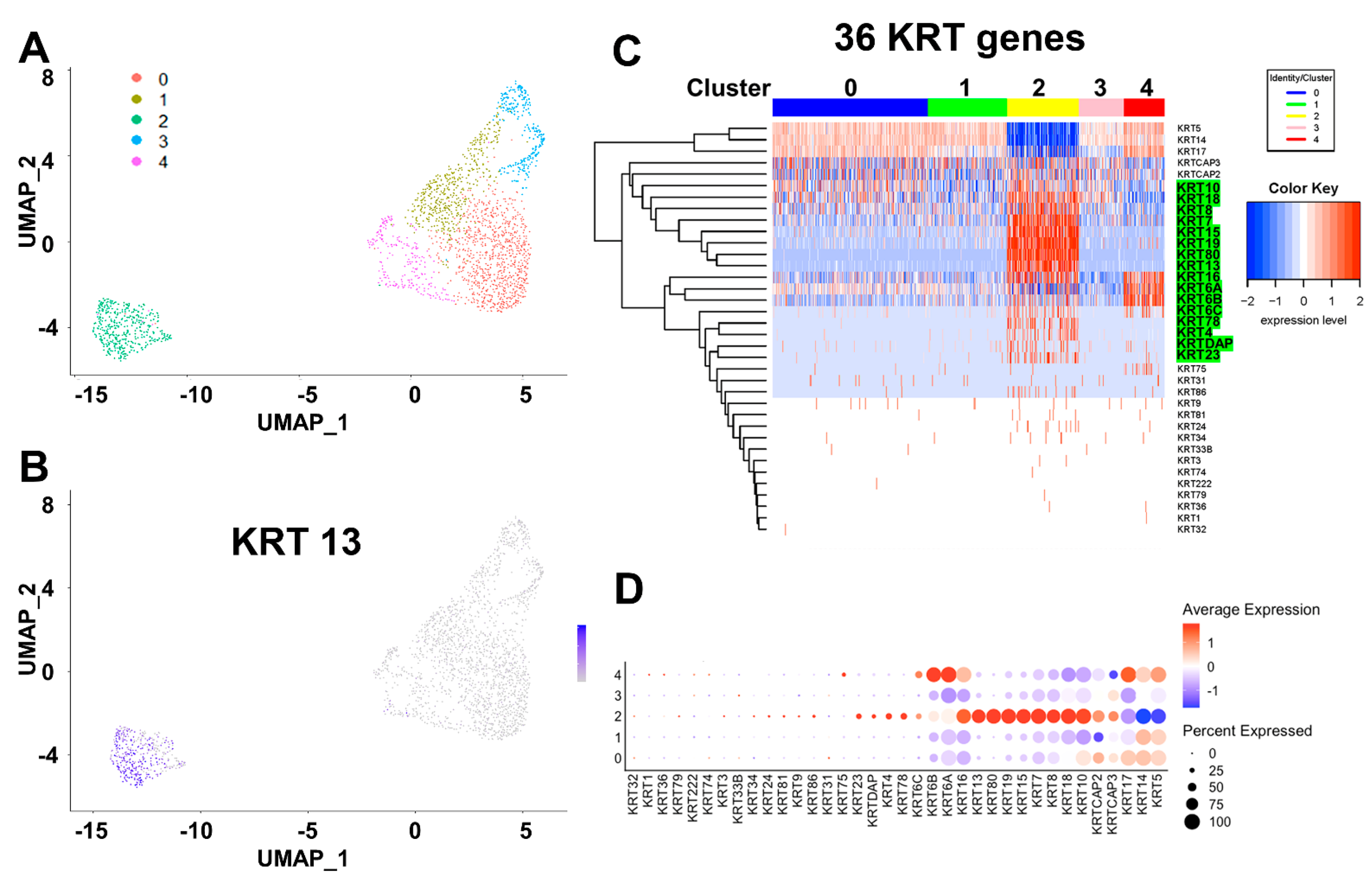
2.3. Keratin Gene Profiling Distinguishes the Prostate Stem and Bipotent Progenitor Cell Clusters
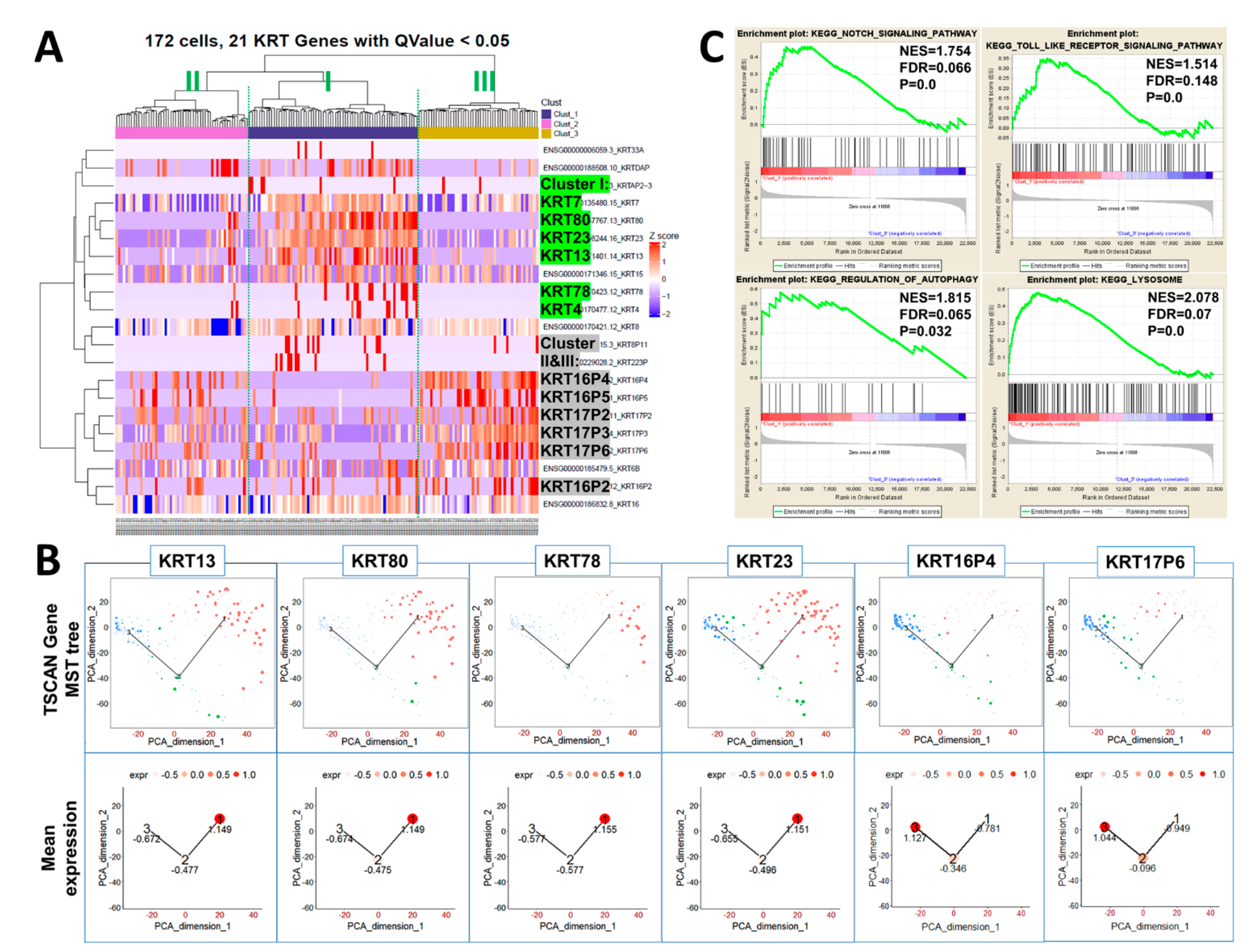
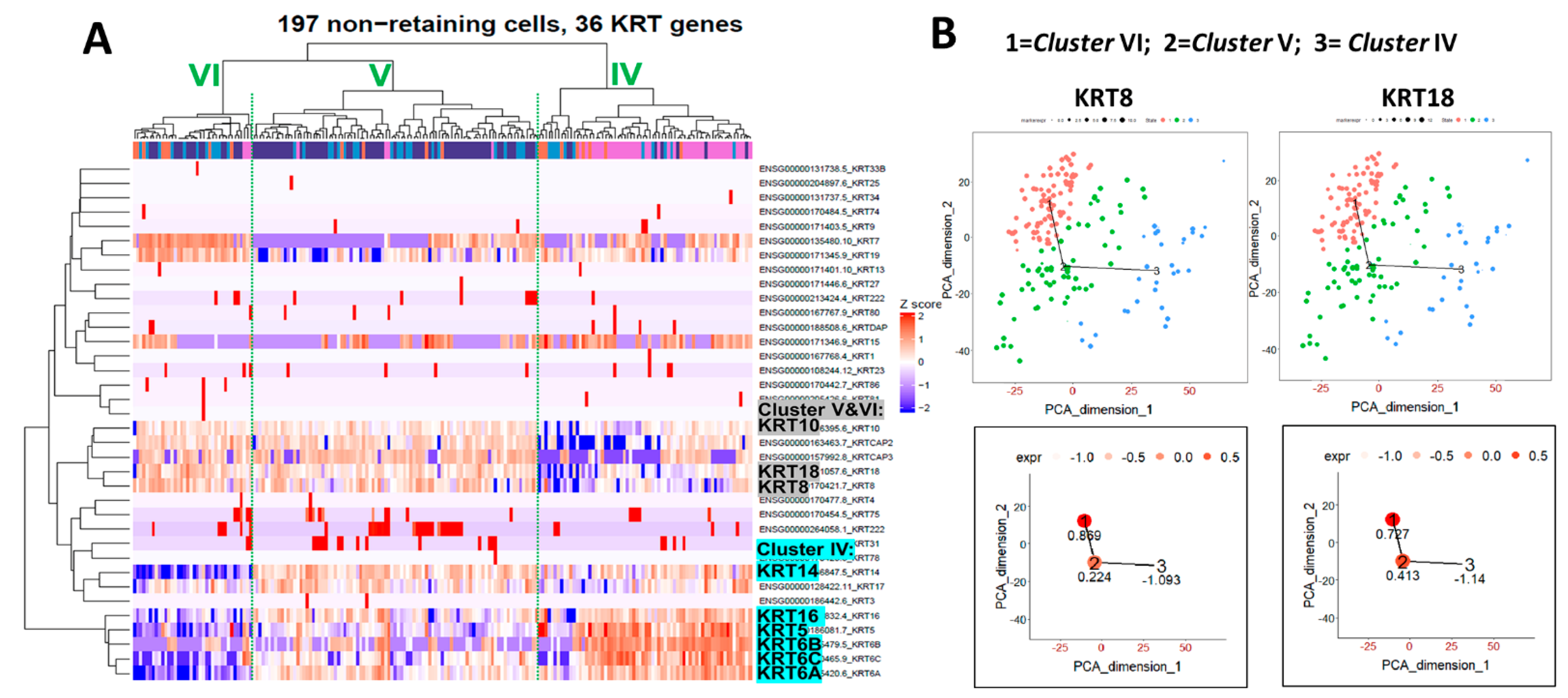
2.4. Heatmap Clustering Analysis Identifies Three Separate Clusters of Prostasphere Progenitor Cells with Differentially Expressed Keratin Genes
2.5. RNA-Seq Reveals Differentially Expressed Genes in Cancer Stem-Like Cells
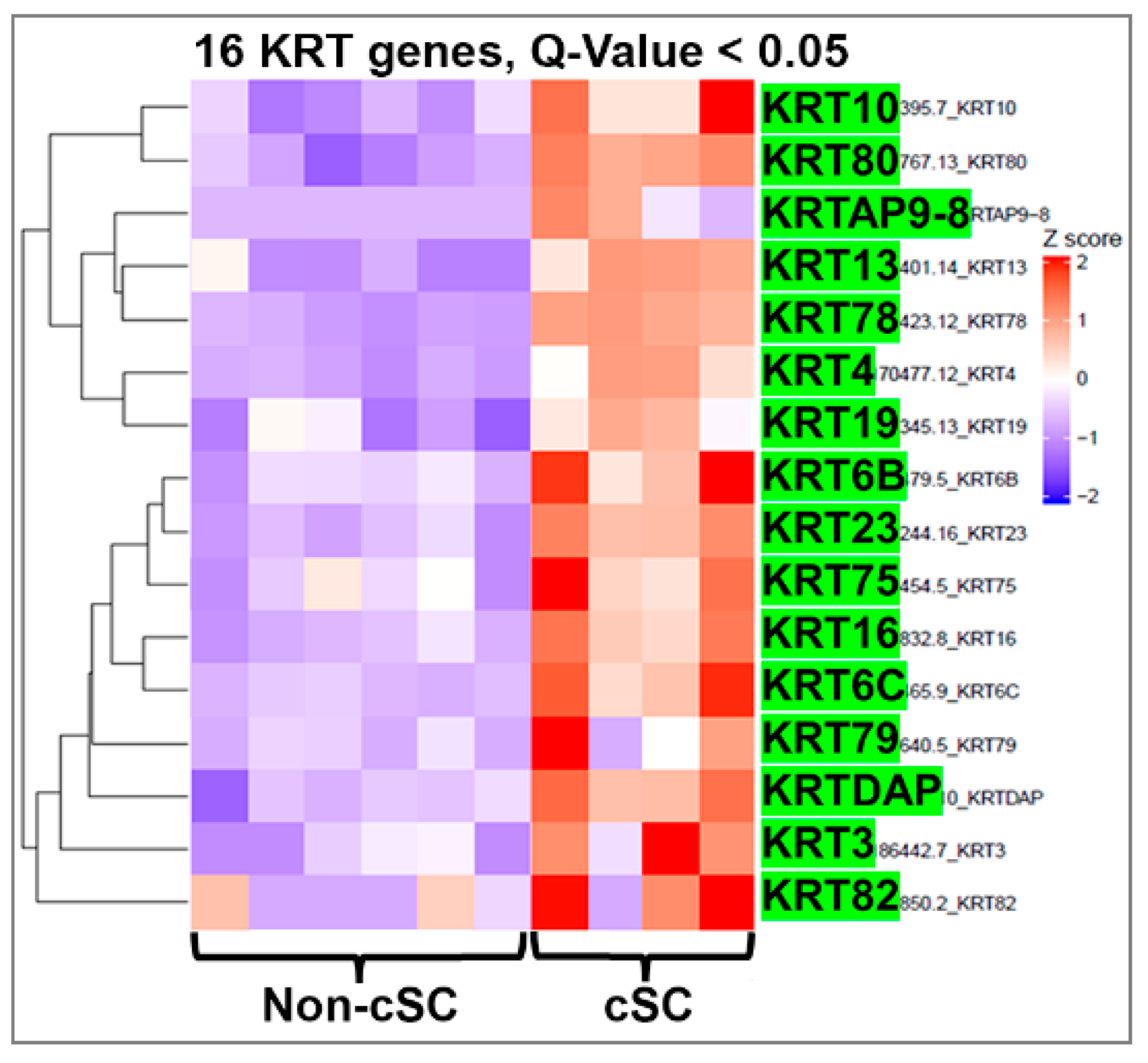
2.6. Prostate Cancer Stem-Like Cells Contain Common and Unique Keratin Genes Compared to Normal Prostate Stem Cells
2.7. Single-Cell RNA-Seq Identifies Cancer Stem-Like Cells with their Differentially Expressed Keratin Genes
3. Discussion
4. Materials and Methods
4.1. Prostate Epithelial Cells and Prostasphere Culture
4.2. Prostasphere-Based Label Retention Assays
4.3. Gene Knockdown by SiRNA
4.4. Next-Generation Bulk RNA-Sequencing Analysis
4.5. Flow Cytometry
4.6. Single-Cell Capture Using the Fluidigm C1-IFC System and Single-Cell RNA-Seq by NextSeq500 Deep Sequencing
4.7. Single-Cell Capture by the 10XGenomics Platform and Single-Cell RNA-Seq Using NovaSeq 6000
4.8. Single-Cell Data Analysis
4.9. Immunocytochemistry (ICC)
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| PrEC | Human prostate epithelial cells |
| PrEGM | ProstaLife epithelial cell growth medium |
| PCa | Prostate cancer |
| KRT | Keratin |
| cSC | Cancer stem-like cells |
| PSCA | Prostate stem cell antigen |
| FACS | Fluorescence-activated cell sorting |
| PCA | Principal component analysis |
| TSCAN | Tool of single cell analysis |
| ICC | Immunocytochemistry |
| CFSE | 5(6)-Carboxyfluorescein N-hydroxysuccinimidyl ester |
| BrdU | Bromodeoxyuridine/5-bromo-2′-deoxyuridine |
| GSEA | Gene set enrichment analysis |
| PSA | Prostate-specific antigen |
References
- Presnell, S.C.; Petersen, B.; Heidaran, M. Stem cells in adult tissues. Semin. Cell Dev. Biol. 2002, 13, 369–376. [Google Scholar] [CrossRef]
- Smith, S.; Neaves, W.; Teitelbaum, S. Adult Versus Embryonic Stem Cells: Treatments. Science 2007, 316, 1422–1423. [Google Scholar] [CrossRef] [PubMed]
- De Marzo, A.M.; Nelson, W.G.; Meeker, A.K.; Coffey, D.S. Stem cell features of benign and malignant prostate epithelial cells. J. Urol. 1998, 160, 2381–2392. [Google Scholar] [CrossRef]
- Goldstein, A.S.; Lawson, D.A.; Cheng, D.; Sun, W.; Garraway, I.P.; Witte, O.N. Trop2 identifies a subpopulation of murine and human prostate basal cells with stem cell characteristics. Porc. Natl. Acad. Sci. USA 2008, 105, 20882–20887. [Google Scholar] [CrossRef] [Green Version]
- Prins, G.S.; Lindgren, M. Accessory sex glands in the male. In Knobil and Neill’s Physiology of Reproduction, 4th ed.; Plant, T.M., Zeleznik, A.J., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 773–804. [Google Scholar]
- Xin, L.; Lukacs, R.U.; Lawson, D.A.; Cheng, D.; Witte, O.N. Self-renewal and multilineage differentiation in vitro from murine prostate stem cells. Stem Cells 2007, 25, 2760–2769. [Google Scholar] [CrossRef]
- Leong, K.G.; Wang, B.E.; Johnson, L.; Gao, W.Q. Generation of a prostate from a single cell. Nature 2008, 456, 804–808. [Google Scholar] [CrossRef]
- Prins, G.S.; Birch, L. The developmental pattern of androgen receptor expression in rat prostate lobes is altered after neonatal exposure to estrogen. Endocrinology 1995, 136, 1303–1314. [Google Scholar] [CrossRef]
- Wang, X.; Xu, H.; Cheng, C.; Ji, Z.; Zhao, H.; Sheng, Y.; Li, X.; Wang, J.; Shu, Y.; He, Y.; et al. Identification of a Zeb1 expressing basal stem cell subpopulation in the prostate. Nat. Commun. 2020, 11, 706. [Google Scholar] [CrossRef] [Green Version]
- Moad, M.; Hannezo, E.; Buczacki, S.J.; Wilson, L.; El-Sherif, A.; Sims, D.; Pickard, R.; Wright, N.A.; Williamson, S.C.; Turnbull, D.M.; et al. Multipotent Basal Stem Cells, Maintained in Localized Proximal Niches, Support Directed Long-Ranging Epithelial Flows in Human Prostates. Cell Rep. 2017, 20, 1609–1622. [Google Scholar] [CrossRef] [Green Version]
- Blackwood, J.K.; Williamson, S.C.; Greaves, L.C.; Wilson, L.; Rigas, A.C.; Sandher, R.; Pickard, R.S.; Robson, C.N.; Turnbull, D.M.; Taylor, R.W.; et al. In situ lineage tracking of human prostatic epithelial stem cell fate reveals a common clonal origin for basal and luminal cells. J. Pathol. 2011, 225, 181–188. [Google Scholar]
- Wang, J.; Zhu, H.H.; Chu, M.; Liu, Y.; Zhang, C.; Liu, G.; Yang, X.; Yang, R.; Gao, W.Q. Symmetrical and asymmetrical division analysis provides evidence for a hierarchy of prostate epithelial cell lineages. Nat. Commun. 2014, 5, 4758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horton, C.; Liu, Y.; Yu, C.; Xie, Q.; Wang, Z.A. Luminal-contact-inhibition of epithelial basal stem cell multipotency in prostate organogenesis and homeostasis. Biol. Open 2019, 8, bio045724. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.A.; Mitrofanova, A.; Bergren, S.K.; Abate-Shen, C.; Cardiff, R.D.; Califano, A.; Shen, M.M. Lineage analysis of basal epithelial cells reveals their unexpected plasticity and supports a cell-of-origin model for prostate cancer heterogeneity. Nat. Cell Biol. 2013, 15, 274–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crowley, L.; Cambuli, F.; Aparicio, L.; Shibata, M.; Robinson, B.D.; Xuan, S.; Li, W.; Hibshoosh, H.; Loda, M.; Rabadan, R.; et al. A single-cell atlas of the mouse and human prostate reveals heterogeneity and conservation of epithelial progenitors. eLife 2020, 9, e59465. [Google Scholar] [CrossRef]
- Toivanen, R.; Mohan, A.; Shen, M.M. Basal Progenitors Contribute to Repair of the Prostate Epithelium Following Induced Luminal Anoikis. Stem Cell Rep. 2016, 6, 660–667. [Google Scholar] [CrossRef] [Green Version]
- Karthaus, W.R.; Iaquinta, P.J.; Drost, J.; Gracanin, A.; van Boxtel, R.; Wongvipat, J.; Dowling, C.M.; Gao, D.; Begthel, H.; Sachs, N.; et al. Identification of multipotent luminal progenitor cells in human prostate organoid cultures. Cell 2014, 159, 163–175. [Google Scholar] [CrossRef] [Green Version]
- Karthaus, W.R.; Hofree, M.; Choi, D.; Linton, E.L.; Turkekul, M.; Bejnood, A.; Carver, B.; Gopalan, A.; Abida, W.; Laudone, V.; et al. Regenerative potential of prostate luminal cells revealed by single-cell analysis. Science 2020, 368, 497–505. [Google Scholar] [CrossRef]
- Hu, W.Y.; Shi, G.B.; Lam, H.M.; Hu, D.P.; Ho, S.M.; Madueke, I.C.; Kajdacsy-Balla, A.; Prins, G.S. Estrogen-initiated transformation of prostate epithelium derived from normal human prostate stem-progenitor cells. Endocrinology 2011, 152, 2150–2163. [Google Scholar] [CrossRef] [Green Version]
- Garraway, I.P.; Sun, W.; Tran, C.P.; Perner, S.; Zhang, B.; Goldstein, A.S.; Hahm, S.A.; Haider, M.; Head, C.S.; Reiter, R.E.; et al. Human prostate sphere-forming cells represent a subset of basal epithelial cells capable of glandular regeneration in vivo. Prostate 2010, 70, 491–501. [Google Scholar] [CrossRef]
- Hu, W.Y.; Shi, G.B.; Hu, D.P.; Nelles, J.L.; Prins, G.S. Actions of endocrine disrupting chemicals on human prostate stem/progenitor cells and prostate cancer risk. Mol. Cell. Endocrinol. 2012, 354, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Prins, G.S.; Hu, W.Y.; Shi, G.B.; Hu, D.P.; Majumdar, S.; Li, G.; Huang, K.; Nelles, J.; Ho, S.M.; Walker, C.L.; et al. Bisphenol A Promotes Human Prostate Stem-Progenitor Cell Self-Renewal and Increases In Vivo Carcinogenesis in Human Prostate Epithelium. Endocrinology 2014, 155, 805–817. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.Y.; Hu, D.P.; Xie, L.; Li, Y.; Majumdar, S.; Nonn, L.; Shioda, T.; Prins, G.S. Isolation and functional interrogation of adult human prostate epithelial stem cells at single cell resolution. Stem Cell Res. 2017, 23, 1–12. [Google Scholar] [CrossRef]
- Hu, W.Y.; Hu, D.P.; Xie, L.; Birch, L.A.; Prins, G.S. Isolation of Stem-like Cells from 3-Dimensional Spheroid Cultures. J. Vis. Exp. 2019, 154, e60357. [Google Scholar] [CrossRef]
- Moll, R.; Divo, M.; Langbein, L. The human keratins: Biology and pathology. Histochem. Cell Biol. 2008, 129, 705–733. [Google Scholar] [CrossRef] [Green Version]
- Bragulla, H.H.; Homberger, D.G. Structure and functions of keratin proteins in simple, stratified, keratinized and cornified epithelia. J. Anat. 2009, 214, 516–559. [Google Scholar] [CrossRef]
- Windoffer, R.; Beil, M.; Magin, T.M.; Leube, R.E. Cytoskeleton in motion: The dynamics of keratin intermediate filaments in epithelia. J. Cell Biol. 2011, 194, 669–678. [Google Scholar] [CrossRef] [Green Version]
- Karantza, V. Keratins in health and cancer: More than mere epithelial cell markers. Oncogene 2011, 30, 127–138. [Google Scholar] [CrossRef] [Green Version]
- Trompetter, M.; Smedts, F.; van der Wijk, J.; Schoots, C.; de Jong, H.J.; Hopman, A.; de la Rosette, J. Keratin profiling in the developing human prostate. A different approach to understanding epithelial lineage. Anticancer. Res. 2008, 28, 237–243. [Google Scholar] [PubMed]
- Okada, H.; Tsubura, A.; Okamura, A.; Senzaki, H.; Naka, Y.; Komatz, Y.; Morii, S. Keratin profiles in normal/hyperplastic prostates and prostate carcinoma. Virchows Arch. A Pathol. Anat. Histopathol. 1992, 421, 157–161. [Google Scholar] [CrossRef]
- Malecha, M.J.; Miettinen, M. Expression of keratin 13 in human epithelial neoplasms. Virchows Arch. A Pathol. Anat. Histopathol. 1991, 418, 249–254. [Google Scholar] [CrossRef]
- Li, Q.; Yin, L.; Jones, L.W.; Chu, G.C.; Wu, J.B.; Huang, J.M.; Li, Q.; You, S.; Kim, J.; Lu, Y.T.; et al. Keratin 13 expression reprograms bone and brain metastases of human prostate cancer cells. Oncotarget 2016, 7, 84645–84657. [Google Scholar] [CrossRef]
- Liu, S.; Cadaneanu, R.M.; Zhang, B.; Huo, L.; Lai, K.; Li, X.; Galet, C.; Grogan, T.R.; Elashoff, D.; Freedland, S.J.; et al. Keratin 13 Is Enriched in Prostate Tubule-Initiating Cells and May Identify Primary Prostate Tumors that Metastasize to the Bone. PLoS ONE 2016, 11, e0163232. [Google Scholar]
- Trapnell, C.; Cacchiarelli, D.; Grimsby, J.; Pokharel, P.; Li, S.; Morse, M.; Lennon, N.J.; Livak, K.J.; Mikkelsen, T.S.; Rinn, J.L. The dynamics and regulators of cell fate decisions are revealed by pseudotemporal ordering of single cells. Nat. Biotechnol. 2014, 32, 381–386. [Google Scholar] [CrossRef] [Green Version]
- Ji, Z.; Ji, H. TSCAN: Pseudo-time reconstruction and evaluation in single-cell RNA-seq analysis. Nucleic Acids Res. 2016, 44, e117. [Google Scholar] [CrossRef] [Green Version]
- Lang, S.H.; Anderson, E.; Fordham, R.; Collins, A.T. Modeling the prostate stem cell niche: An evaluation of stem cell survival and expansion in vitro. Stem Cells Dev. 2010, 19, 537–546. [Google Scholar] [CrossRef] [Green Version]
- Blum, R.; Gupta, R.; Burger, P.E.; Ontiveros, C.S.; Salm, S.N.; Xiong, X.; Kamb, A.; Wesche, H.; Marshall, L.; Cutler, G.; et al. Molecular signatures of the primitive prostate stem cell niche reveal novel mesenchymal-epithelial signaling pathways. PLoS ONE 2010, 5, e13024. [Google Scholar] [CrossRef] [Green Version]
- Richards, Z.; McCray, T.; Marsili, J.; Zenner, M.L.; Manlucu, J.T.; Garcia, J.; Kajdacsy-Balla, A.; Murray, M.; Voisine, C.; Murphy, A.B.; et al. Prostate Stroma Increases the Viability and Maintains the Branching Phenotype of Human Prostate Organoids. iScience 2019, 12, 304–317. [Google Scholar] [CrossRef] [Green Version]
- Gu, Z.; Thomas, G.; Yamashiro, J.; Shintaku, I.P.; Dorey, F.; Raitano, A.; Witte, O.N.; Said, J.W.; Loda, M.; Reiter, R.E. Prostate stem cell antigen (PSCA) expression increases with high gleason score, advanced stage and bone metastasis in prostate cancer. Oncogene 2000, 19, 1288–1296. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, R.; Manzar, N.; Bhatia, V.; Yadav, A.; Nengroo, M.A.; Datta, D.; Carskadon, S.; Gupta, N.; Sigouros, M.; Khani, F.; et al. Androgen deprivation upregulates SPINK1 expression and potentiates cellular plasticity in prostate cancer. Nat. Commun. 2020, 11, 384. [Google Scholar] [CrossRef] [Green Version]
- Hale, J.S.; Otvos, B.; Sinyuk, M.; Alvarado, A.G.; Hitomi, M.; Stoltz, K.; Wu, Q.; Flavahan, W.; Levison, B.; Johansen, M.L.; et al. Cancer stem cell-specific scavenger receptor CD36 drives glioblastoma progression. Stem Cells 2014, 32, 1746–1758. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Fleming, A.; Ricketts, T.; Pavel, M.; Virgin, H.; Menzies, F.M.; Rubinsztein, D.C. Autophagy regulates Notch degradation and modulates stem cell development and neurogenesis. Nat. Commun. 2016, 7, 10533. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Shi, B.; Liu, X.; Jiang, M.; Yuan, C.; Jiang, B.; Song, Y.; Zeng, Y.; Wang, G. The activation of Toll-like receptor 4 reverses tumor differentiation in human glioma U251 cells via Notch pathway. Int. Immunopharmacol. 2018, 64, 33–41. [Google Scholar] [CrossRef]
- Kobayashi, T.; Piao, W.; Takamura, T.; Kori, H.; Miyachi, H.; Kitano, S.; Iwamoto, Y.; Yamada, M.; Imayoshi, I.; Shioda, S.; et al. Enhanced lysosomal degradation maintains the quiescent state of neural stem cells. Nat. Commun. 2019, 10, 5446. [Google Scholar] [CrossRef] [PubMed]
- Gaisa, N.T.; Graham, T.A.; McDonald, S.A.; Poulsom, R.; Heidenreich, A.; Jakse, G.; Knuechel, R.; Wright, N.A. Clonal architecture of human prostatic epithelium in benign and malignant conditions. J. Pathol. 2011, 225, 172–180. [Google Scholar] [CrossRef]
- Manzo, G. Similarities Between Embryo Development and Cancer Process Suggest New Strategies for Research and Therapy of Tumors: A New Point of View. Front. Cell Dev. Biol. 2019, 7, 20. [Google Scholar] [CrossRef]
- Naxerova, K.; Bult, C.J.; Peaston, A.; Fancher, K.; Knowles, B.B.; Kasif, S.; Kohane, I.S. Analysis of gene expression in a developmental context emphasizes distinct biological leitmotifs in human cancers. Genome Biol. 2008, 9, R108. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, D.T.; Zheng, Y.; Wittner, B.S.; Lee, R.J.; Zhu, H.; Broderick, K.T.; Desai, R.; Fox, D.B.; Brannigan, B.W.; Trautwein, J.; et al. RNA-Seq of single prostate CTCs implicates noncanonical Wnt signaling in antiandrogen resistance. Science 2015, 349, 1351–1356. [Google Scholar] [CrossRef] [Green Version]
- Xie, L.; Hu, W.Y.; Hu, D.P.; Shi, G.B.; Li, Y.; Yang, J.F.; Prins, G.S. Effects of Inorganic Arsenic on Human Prostate Stem-Progenitor Cell Transformation, Autophagic Flux Blockade, and NRF2 Pathway Activation. Environ. Health Perspect. 2020, 128, 67008. [Google Scholar] [CrossRef] [PubMed]
- Chua, C.W.; Shibata, M.; Lei, M.; Toivanen, R.; Barlow, L.J.; Bergren, S.K.; Badani, K.K.; McKiernan, J.M.; Benson, M.C.; Hibshoosh, H.; et al. Single luminal epithelial progenitors can generate prostate organoids in culture. Nat. Cell Biol. 2014, 16, 951–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasper, S. Exploring the origins of the normal prostate and prostate cancer stem cell. Stem Cell Rev. 2008, 4, 193–201. [Google Scholar] [CrossRef]
- Yang, Y.; Hao, J.; Liu, X.; Dalkin, B.; Nagle, R.B. Differential expression of cytokeratin mRNA and protein in normal prostate, prostatic intraepithelial neoplasia, and invasive carcinoma. Am. J. Pathol. 1997, 150, 693–704. [Google Scholar] [PubMed]
- Xue, Y.; Verhofstad, A.; Lange, W.; Smedts, F.; Debruyne, F.; de la Rosette, J.; Schalken, J. Prostatic neuroendocrine cells have a unique keratin expression pattern and do not express Bcl-2: Cell kinetic features of neuroendocrine cells in the human prostate. Am. J. Pathol. 1997, 151, 1759–1765. [Google Scholar] [PubMed]
- van Leenders, G.J.; Aalders, T.W.; Hulsbergen-van de Kaa, C.A.; Ruiter, D.J.; Schalken, J.A. Expression of basal cell keratins in human prostate cancer metastases and cell lines. J. Pathol. 2001, 195, 563–570. [Google Scholar] [CrossRef]
- Ferronika, P.; Triningsih, F.X.; Ghozali, A.; Moeljono, A.; Rahmayanti, S.; Shadrina, A.N.; Naim, A.E.; Wudexi, I.; Arnurisa, A.M.; Nanwani, S.T.; et al. p63 cytoplasmic aberrance is associated with high prostate cancer stem cell expression. Asian Pac. J. Cancer Prev. 2012, 13, 1943–1948. [Google Scholar] [CrossRef]
- Dhillon, P.K.; Barry, M.; Stampfer, M.J.; Perner, S.; Fiorentino, M.; Fornari, A.; Ma, J.; Fleet, J.; Kurth, T.; Rubin, M.A.; et al. Aberrant cytoplasmic expression of p63 and prostate cancer mortality. Cancer Epidemiol. Biomark. Prev. 2009, 18, 595–600. [Google Scholar] [CrossRef] [Green Version]
- Maitland, N.J.; Frame, F.M.; Polson, E.S.; Lewis, J.L.; Collins, A.T. Prostate Cancer Stem Cells: Do They Have a Basal or Luminal Phenotype? Horm. Cancer 2011, 2, 47–61. [Google Scholar] [CrossRef] [Green Version]
- Gu, G.; Yuan, J.; Wills, M.; Kasper, S. Prostate cancer cells with stem cell characteristics reconstitute the original human tumor in vivo. Cancer Res. 2007, 67, 4807–4815. [Google Scholar] [CrossRef] [Green Version]
- Collins, A.T.; Berry, P.A.; Hyde, C.; Stower, M.J.; Maitland, N.J. Prospective identification of tumorigenic prostate cancer stem cells. Cancer Res. 2005, 65, 10946–10951. [Google Scholar] [CrossRef] [Green Version]
- Skvortsov, S.; Skvortsova, I.I.; Tang, D.G.; Dubrovska, A. Concise Review: Prostate Cancer Stem Cells: Current Understanding. Stem Cells. 2018, 36, 1457–1474. [Google Scholar] [CrossRef] [Green Version]
- Hudson, D.L. Epithelial stem cells in human prostate growth and disease. Prostate Cancer Prostatic Dis. 2004, 7, 188–194. [Google Scholar] [CrossRef]
- Vander Griend, D.J.; Karthaus, W.L.; Dalrymple, S.; Meeker, A.; DeMarzo, A.M.; Isaacs, J.T. The role of CD133 in normal human prostate stem cells and malignant cancer-initiating cells. Cancer Res. 2008, 68, 9703–9711. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Li, Q.; Liu, X.; Liu, C.; Liu, R.; Rycaj, K.; Zhang, D.; Liu, B.; Jeter, C.; Calhoun-Davis, T.; et al. Defining a Population of Stem-like Human Prostate Cancer Cells That Can Generate and Propagate Castration-Resistant Prostate Cancer. Clin. Cancer Res. 2016, 22, 4505–4516. [Google Scholar] [CrossRef] [Green Version]
- Smith, B.A.; Sokolov, A.; Uzunangelov, V.; Baertsch, R.; Newton, Y.; Graim, K.; Mathis, C.; Cheng, D.; Stuart, J.M.; Witte, O.N. A basal stem cell signature identifies aggressive prostate cancer phenotypes. Porc. Natl. Acad. Sci. USA 2015, 112, E6544–E6552. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Porc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Macosko, E.Z.; Basu, A.; Satija, R.; Nemesh, J.; Shekhar, K.; Goldman, M.; Tirosh, I.; Bialas, A.R.; Kamitaki, N.; Martersteck, E.M.; et al. Highly Parallel Genome-wide Expression Profiling of Individual Cells Using Nanoliter Droplets. Cell 2015, 161, 1202–1214. [Google Scholar] [CrossRef] [Green Version]
- Stuart, T.; Butler, A.; Hoffman, P.; Hafemeister, C.; Papalexi, E.; Mauck WM 3rd Hao, Y.; Stoeckius, M.; Smibert, P.; Satija, R. Comprehensive Integration of Single-Cell Data. Cell 2019, 177, 1888–1902. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | cSC/non-cSC | |
|---|---|---|
| Symbol | Gene Name | (Fold Change) |
| SCGB2A1 | Secretoglobulin family 2A1 | 67,370 |
| SPINK1 | Serine peptidase inhibitor kazak-type 1 | 2270 |
| PSCA | Prostate stem cell antigen | 1621 |
| CD36 | Scavenger receptor class B member 3 | 990 |
| TMPRSS11E | Transmembrane serine protease 11E | 593 |
| HMOX1 | Heme oxygenase 1 | 128 |
| IGF2 | Insulin growth factor 2 | 117 |
| (A) Pathways enriched in cSC cells (Prostate_Cancer_916_genes_Qvalue_0.05_genelist) | |||||
| Maps | Total | p-Value | FDR | In Data | Network Objects from Active Data |
| Cytoskeleton remodeling: Keratin filaments | 36 | 2.7 × 10−8 | 1.8 × 10−5 | 10 | Keratin 16, 14-3-3 gamma, Keratin 4, PPL (periplakin), Keratin 6C, Keratin 6A, Keratin 19, Keratin 13, Keratin 4/13, Plakophilin 1 |
| Ligand-independent activation of androgen receptor in PCa | 67 | 1.3 × 10−5 | 3.1 × 10−3 | 10 | GAB1, Bcl-XL, PP2A regulatory, c-Myc, ERK1 (MAPK3), S5AR2, PP2A catalytic, FGFR1, Kallikrein 3 (PSA), ErbB3 |
| Androgen receptor activation and downstream signaling in PCa | 110 | 8.5 × 10−4 | 6.2 × 10−2 | 10 | GAB1, Bcl-XL, TMPRSS2, c-Myc, ERK1 (MAPK3), S5AR2, Kallikrein 2, FGFR1, PSCA, Kallikrein 3 (PSA) |
| Transcription: HIF-1 targets | 95 | 5.4 × 10−5 | 0.0071 | 11 | ROR-alpha, TGM2, Mxi1, CITED2, Heme oxygenase 1, HXK2, c-Myc, Cyclin G2, LOXL2, AK3, CTGF |
| Development: IGF-1 receptor signaling | 51 | 0.00044 | 0.0416 | 7 | Bcl-XL, MNK2(GPRK7), c-Myc, IGF-2, IBP, FOXO3A, ERK1/2 |
| Autophagy | 35 | 0.00246 | 0.0993 | 5 | GATE-16, Bcl-XL, WIPI2, MAP1LC3A, ULK1 |
| (B) Pathways enriched in non-cSC cells (Prostate_Cancer_394_genes_Qvalue_0.05_genelist) | |||||
| Maps | Total | p-Value | FDR | In Data | Network Objects from Active Data |
| Cell cycle: Chromosome condensation in prometaphase | 21 | 4.4 × 10−14 | 2 × 10−11 | 11 | Aurora-B, BRRN1, CAP-H/H2, CAP-G, CAP-E, Cyclin B, CAP-C, TOP2, CAP-G/G2, Histone H1, CDK1 (p34) |
| Cell cycle: The metaphase checkpoint | 36 | 2.3 × 10−12 | 6 × 10−10 | 12 | Nek2A, Aurora-B, HEC, CDCA1, CDC20, HZwint-1, Rod, CENP-F, DSN1, SPBC24, CENP-E, PLK1 |
| Cell cycle: Transition and termination of DNA replication | 27 | 1.4 × 10−9 | 2 × 10−7 | 9 | TOP2 alpha, POLD reg (p68), Brca1/Bard1, Brca1, MCM2, TOP2, FEN1, Bard1, CDK1 (p34) |
| Cell cycle: Start of DNA replication in early S phase | 32 | 2.5 × 10−6 | 0.0001 | 7 | CDC18L (CDC6), MCM10, Geminin, MCM4, MCM2, Histone H1, CDC45L |
| Cell cycle: Initiation of mitosis | 26 | 2.5 × 10−8 | 3 × 10−6 | 8 | Lamin B, Cyclin B1, Cyclin B2, FOXM1, Kinase MYT1, Histone H1, CDK1 (p34), PLK1 |
| Signal transduction: NF-kB activation pathways | 51 | 5 × 10−8 | 4 × 10−6 | 10 | NF-kB2 (p100), IL-1 beta, NF-kB2 (p52), I-kB, NF-kB, c-IAP1, c-IAP2, NF-kB1 (p105), BAFF(TNFSF13B), NF-kB1 (p50) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, W.-Y.; Hu, D.-P.; Xie, L.; Nonn, L.; Lu, R.; Abern, M.; Shioda, T.; Prins, G.S. Keratin Profiling by Single-Cell RNA-Sequencing Identifies Human Prostate Stem Cell Lineage Hierarchy and Cancer Stem-Like Cells. Int. J. Mol. Sci. 2021, 22, 8109. https://doi.org/10.3390/ijms22158109
Hu W-Y, Hu D-P, Xie L, Nonn L, Lu R, Abern M, Shioda T, Prins GS. Keratin Profiling by Single-Cell RNA-Sequencing Identifies Human Prostate Stem Cell Lineage Hierarchy and Cancer Stem-Like Cells. International Journal of Molecular Sciences. 2021; 22(15):8109. https://doi.org/10.3390/ijms22158109
Chicago/Turabian StyleHu, Wen-Yang, Dan-Ping Hu, Lishi Xie, Larisa Nonn, Ranli Lu, Michael Abern, Toshihiro Shioda, and Gail S. Prins. 2021. "Keratin Profiling by Single-Cell RNA-Sequencing Identifies Human Prostate Stem Cell Lineage Hierarchy and Cancer Stem-Like Cells" International Journal of Molecular Sciences 22, no. 15: 8109. https://doi.org/10.3390/ijms22158109
APA StyleHu, W. -Y., Hu, D. -P., Xie, L., Nonn, L., Lu, R., Abern, M., Shioda, T., & Prins, G. S. (2021). Keratin Profiling by Single-Cell RNA-Sequencing Identifies Human Prostate Stem Cell Lineage Hierarchy and Cancer Stem-Like Cells. International Journal of Molecular Sciences, 22(15), 8109. https://doi.org/10.3390/ijms22158109