
Sex Differences in the Triad of Acquired Sensorineural Hearing Loss



Abstract
:1. Introduction
2. Clinical Aspects of Sex Differences in Acquired Sensorineural Hearing Loss
2.1. NIHL
2.2. Ototoxicity
2.3. ARHL
2.4. Other Pathological Diseases Associated to ASNHL
3. Animal Investigations of Sex Differences in Acquired Sensorineural Hearing Loss
3.1. NIHL
3.2. Ototoxicity
3.3. ARHL
4. How Could Hormones Influence Hearing in Molecular Aspects?
4.1. Estrogen in Auditory Function
4.2. Potential Molecular Effects of Estrogen and Receptors on the Auditory System

4.3. The Role of Estrogen on Sex Differences in ASNHL
4.4. Androgen in Auditory Function
5. The Effect of Hormone Therapy on Hearing
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Available online: https://www.who.int/health-topics/hearing-loss#tab=tab_2 (accessed on 17 June 2021).
- Yang, C.H.; Schrepfer, T.; Schacht, J. Age-related hearing impairment and the triad of acquired hearing loss. Front. Cell Neurosci. 2015, 9, 276. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.I.; Nelson, R.Y.; Concha-Barrientos, M.; Fingerhut, M. The global burden of occupational noise-induced hearing loss. Am. J. Ind. Med. 2005, 48, 446–458. [Google Scholar] [CrossRef] [Green Version]
- Dobie, R.A. Hearing conservation in industry. West. J. Med. 1982, 137, 499. [Google Scholar]
- McBride, D.; Williams, S. Audiometric notch as a sign of noise induced hearing loss. Occup. Environ. Med. 2001, 58, 46–51. [Google Scholar] [CrossRef] [Green Version]
- Hong, O.; Kerr, M.J.; Poling, G.L.; Dhar, S. Understanding and preventing noise-induced hearing loss. Dis. Mon. 2013, 59, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Engdahl, B.; Tambs, K.; Borchgrevink, H.M.; Hoffman, H.J. Screened and unscreened hearing threshold levels for the adult population: Results from the Nord-Trøndelag Hearing Loss Study Niveles de umbrales auditivos tamizados y no tamizados en la población adulta. Resultados del estudio Nord-TrøndeIag sobre hipoacusias. Int. J. Audiol. 2005, 44, 213–230. [Google Scholar]
- Engdahl, B.; Tambs, K. Occupation and the risk of hearing impairment—Results from the Nord-Trøndelag study on hearing loss. Scand. J. Work Environ. Health 2010, 36, 250–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Wang, X.; Yang, L.; Han, K.; Huang, Z.; Wu, H. Sex differences in noise-induced hearing loss: A cross-sectional study in China. Biol. Sex Differ. 2021, 12, 24. [Google Scholar] [CrossRef]
- Zhou, J.; Shi, Z.; Zhou, L.; Hu, Y.; Zhang, M. Occupational noise-induced hearing loss in China: A systematic review and meta-analysis. BMJ Open 2020, 10, e039576. [Google Scholar] [CrossRef] [PubMed]
- Chung, D.Y.; Mason, K.; Gannon, R.P.; Willson, G.N. The ear effect as a function of age and hearing loss. J. Acoust. Soc. Am. 1983, 73, 1277–1282. [Google Scholar] [CrossRef]
- Morata, T.C. Young people: Their noise and music exposures and the risk of hearing loss. Int. J. Audiol. 2007, 467, 111–112. [Google Scholar] [CrossRef]
- Warner-Czyz, A.D.; Cain, S. Age and gender differences in children and adolescents’ attitudes toward noise. Int. J. Audiol. 2016, 55, 83–92. [Google Scholar] [CrossRef]
- Portnuff, C.D. Reducing the risk of music-induced hearing loss from overuse of portable listening devices: Understanding the problems and establishing strategies for improving awareness in adolescents. Adolesc. Health Med. Ther. 2016, 7, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmes, A.E.; Kaplan, H.S.; Phillips, R.M.; Kemker, F.J.; Weber, F.T.; Isart, F.A. Screening for hearing loss in adolescents. Lang. Speech Hear. Serv. Sch. 1997, 28, 70–76. [Google Scholar] [CrossRef]
- Park, Y.H.; Shin, S.-H.; Byun, S.W.; Kim, J.Y. Age-and gender-related mean hearing threshold in a highly-screened population: The Korean National Health and Nutrition Examination Survey 2010–2012. PLoS ONE 2016, 11, e0150783. [Google Scholar] [CrossRef] [PubMed]
- Juhn, S.; Rybak, L.; Prado, S. Nature of blood-labyrinth barrier in experimental conditions. Ann. Otol. Rhinol. Laryngol. 1981, 90, 135–141. [Google Scholar] [CrossRef]
- Salt, A.N. Simulation of methods for drug delivery to the cochlear fluids. Adv. Otorhinolaryngol. 2002, 59, 140–148. [Google Scholar]
- Salt, A.N.; Plontke, S.K. Local inner-ear drug delivery and pharmacokinetics. Drug Discov. Today 2005, 10, 1299–1306. [Google Scholar] [CrossRef] [Green Version]
- Forge, A.; Li, L.; Corwin, J.T.; Nevill, G. Ultrastructural evidence for hair cell regeneration in the mammalian inner ear. Science 1993, 259, 1616–1619. [Google Scholar] [CrossRef]
- Franconi, F.; Brunelleschi, S.; Steardo, L.; Cuomo, V. Gender differences in drug responses. Pharmacol. Res. 2007, 55, 81–95. [Google Scholar] [CrossRef]
- Beierle, I.; Meibohm, B.; Derendorf, H. Gender differences in pharmacokinetics and pharmacodynamics. Int. J. Clin. Pharmacol. Ther. 1999, 37, 529–547. [Google Scholar]
- Aznar, M.L.; Marras, T.K.; Elshal, A.S.; Mehrabi, M.; Brode, S.K. Safety and effectiveness of low-dose amikacin in nontuberculous mycobacterial pulmonary disease treated in Toronto, Canada. BMC Pharmacol. Toxicol. 2019, 20, 1–9. [Google Scholar] [CrossRef]
- Ross, C.J.; Katzov-Eckert, H.; Dubé, M.-P.; Brooks, B.; Rassekh, S.R.; Barhdadi, A.; Feroz-Zada, Y.; Visscher, H.; Brown, A.M.; Rieder, M.J. Genetic variants in TPMT and COMT are associated with hearing loss in children receiving cisplatin chemotherapy. Nat. Genet. 2009, 41, 1345–1349. [Google Scholar] [CrossRef] [PubMed]
- Knight, K.R.G.; Kraemer, D.F.; Neuwelt, E.A. Ototoxicity in children receiving platinum chemotherapy: Underestimating a commonly occurring toxicity that may influence academic and social development. J. Clin. Oncol. 2005, 23, 8588–8596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yancey, A.; Harris, M.S.; Egbelakin, A.; Gilbert, J.; Pisoni, D.B.; Renbarger, J. Risk factors for cisplatin-associated ototoxicity in pediatric oncology patients. Pediatric Blood Cancer 2012, 59, 144–148. [Google Scholar] [CrossRef] [Green Version]
- Castelán-Martínez, O.D.; Jiménez-Méndez, R.; Rodríguez-Islas, F.; Fierro-Evans, M.; Vázquez-Gómez, B.E.; Medina-Sansón, A.; Clark, P.; Carleton, B.; Ross, C.; Hildebrand, C. Hearing loss in Mexican children treated with cisplatin. Int. J. Pediatric Otorhinolaryngol. 2014, 78, 1456–1460. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.S.; Kistner, E.O.; Bleibel, W.K.; Shukla, S.J.; Dolan, M.E. Effect of population and gender on chemotherapeutic agent–induced cytotoxicity. Mol. Cancer Ther. 2007, 6, 31–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Womer, R.; Silber, J. Predicting cisplatin ototoxicity in children: The influence of age and the cumulative dose. Eur. J. Cancer 2004, 40, 2445–2451. [Google Scholar] [CrossRef]
- Davis, A.; McMahon, C.M.; Pichora-Fuller, K.M.; Russ, S.; Lin, F.; Olusanya, B.O.; Chadha, S.; Tremblay, K.L. Aging and hearing health: The life-course approach. Gerontologist 2016, 56 (Suppl. S2), S256–S267. [Google Scholar] [CrossRef]
- Nash, S.D.; Cruickshanks, K.J.; Klein, R.; Klein, B.E.; Nieto, F.J.; Huang, G.H.; Pankow, J.S.; Tweed, T.S. The prevalence of hearing impairment and associated risk factors: The Beaver Dam Offspring Study. Arch. Otolaryngol. Head Neck Surg. 2011, 137, 432–439. [Google Scholar] [CrossRef] [Green Version]
- Slade, K.; Plack, C.J.; Nuttall, H.E. The effects of age-related hearing loss on the brain and cognitive function. Trends Neurosci. 2020, 43, 810–821. [Google Scholar] [CrossRef]
- Cardin, V. Effects of aging and adult-onset hearing loss on cortical auditory regions. Front. Neurosci. 2016, 10, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutherford, B.R.; Brewster, K.; Golub, J.S.; Kim, A.H.; Roose, S.P. Sensation and psychiatry: Linking age-related hearing loss to late-life depression and cognitive decline. Am. J. Psychiatry 2018, 175, 215–224. [Google Scholar] [CrossRef] [Green Version]
- Livingston, G.; Huntley, J.; Sommerlad, A.; Ames, D.; Ballard, C.; Banerjee, S.; Brayne, C.; Burns, A.; Cohen-Mansfield, J.; Cooper, C. Dementia prevention, intervention, and care: 2020 report of the Lancet Commission. Lancet 2020, 396, 413–446. [Google Scholar] [CrossRef]
- Pearson, J.D.; Morrell, C.H.; Gordon-Salant, S.; Brant, L.J.; Metter, E.J.; Klein, L.L.; Fozard, J.L. Gender differences in a longitudinal study of age-associated hearing loss. J. Acoust. Soc. Am. 1995, 97, 1196–1205. [Google Scholar] [CrossRef]
- Cruickshanks, K.J.; Wiley, T.L.; Tweed, T.S.; Klein, B.E.; Klein, R.; Mares-Perlman, J.A.; Nondahl, D.M. Prevalence of hearing loss in older adults in Beaver Dam, Wisconsin: The epidemiology of hearing loss study. Am. J. Epidemiol. 1998, 148, 879–886. [Google Scholar] [CrossRef]
- Gates, G.A.; Cooper Jr, J.; Kannel, W.B.; Miller, N.J. Hearing in the elderly: The Framingham cohort, 1983-1985. Part I. Basic audiometric test results. Ear Hear. 1990, 11, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Megighian, D.; Savastano, M.; Salvador, L.; Frigo, A.; Bolzan, M. Audiometric and epidemiological analysis of elderly in the Veneto region. Gerontology 2000, 46, 199–204. [Google Scholar] [CrossRef]
- Jönsson, R.; Rosenhall, U.; Gause-Nilsson, I.; Steen, B. Auditory function in 70-and 75-year-olds of four age cohorts. Scand. Audiol. 1998, 27, 81–93. [Google Scholar] [CrossRef]
- Pedersen, K.E.; Rosenhall, U.; Metier, M.B. Changes in pure-tone thresholds in individuals aged 70-81: Results from a longitudinal study. Audiology 1989, 28, 194–204. [Google Scholar] [CrossRef]
- Bishop, C.E.; Spankovich, C.; Lin, F.R.; Seals, S.R.; Su, D.; Valle, K.; Schweinfurth, J.M. Audiologic profile of the jackson heart study cohort and comparison to other cohorts. Laryngoscope 2019, 129, 2391–2397. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lim, E.J.; Kim, H.S.; Park, J.H.; Jarng, S.S.; Lee, S.H. Sex differences in a cross sectional study of age-related hearing loss in Korean. Clin. Exp. Otorhinolaryngol. 2010, 3, 27. [Google Scholar] [CrossRef] [PubMed]
- Homans, N.C.; Metselaar, R.M.; Dingemanse, J.G.; van der Schroeff, M.P.; Brocaar, M.P.; Wieringa, M.H.; Baatenburg de Jong, R.J.; Hofman, A.; Goedegebure, A. Prevalence of age-related hearing loss, including sex differences, in older adults in a large cohort study. Laryngoscope 2017, 127, 725–730. [Google Scholar] [CrossRef] [PubMed]
- Thomson, R.S.; Auduong, P.; Miller, A.T.; Gurgel, R.K. Hearing loss as a risk factor for dementia: A systematic review. Laryngoscope Investig. Otolaryngol. 2017, 2, 69–79. [Google Scholar] [CrossRef]
- Livingston, G.; Sommerlad, A.; Orgeta, V.; Costafreda, S.G.; Huntley, J.; Ames, D.; Ballard, C.; Banerjee, S.; Burns, A.; Cohen-Mansfield, J. Dementia prevention, intervention, and care. Lancet 2017, 390, 2673–2734. [Google Scholar] [CrossRef] [Green Version]
- Lyu, J.; Kim, H.-Y. Gender-specific associations of sensory impairments with depression and cognitive impairment in later life. Psychiatry Investig. 2018, 15, 926. [Google Scholar] [CrossRef] [Green Version]
- Huang, B.; Cao, G.; Duan, Y.; Yan, S.; Yan, M.; Yin, P.; Jiang, H. Gender Differences in the Association Between Hearing Loss and Cognitive Function. Am. J. Alzheimer’s Dis. Other Dement. 2020, 35, 1533317519871167. [Google Scholar] [CrossRef]
- Denton, M.; Prus, S.; Walters, V. Gender differences in health: A Canadian study of the psychosocial, structural and behavioural determinants of health. Soc. Sci. Med. 2004, 58, 2585–2600. [Google Scholar] [CrossRef] [Green Version]
- Heine, C.; Browning, C.; Cowlishaw, S.; Kendig, H. Trajectories of older adults’ hearing difficulties: Examining the influence of health behaviors and social activity over 10 years. Geriatr. Gerontol. Int. 2013, 13, 911–918. [Google Scholar] [CrossRef] [PubMed]
- Vigil, J.M. Asymmetries in the friendship preferences and social styles of men and women. Hum. Nat. 2007, 18, 143–161. [Google Scholar] [CrossRef]
- Armstrong, N.M.; An, Y.; Beason-Held, L.; Doshi, J.; Erus, G.; Ferrucci, L.; Davatzikos, C.; Resnick, S.M. Sex differences in brain aging and predictors of neurodegeneration in cognitively healthy older adults. Neurobiol. Aging 2019, 81, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Luryi, A.L.; Morse, E.; Michaelides, E. Pathophysiology and Diagnosis of Meniere’s Disease. In Diagnosis and Treatment of Vestibular Disorders; Springer: Berlin/Heidelberg, Germany, 2019; pp. 165–188. [Google Scholar]
- Smith, P.F.; Agrawal, Y.; Darlington, C.L. Sexual dimorphism in vestibular function and dysfunction. J. Neurophysiol. 2019, 121, 2379–2391. [Google Scholar] [CrossRef]
- Simo, H.; Yang, S.; Qu, W.; Preis, M.; Nazzal, M.; Baugh, R. Meniere’s disease: Importance of socioeconomic and environmental factors. Am. J. Otolaryngol. 2015, 36, 393–398. [Google Scholar] [CrossRef]
- Jian, H.; Yu, G.; Chen, G.; Lin, N.; Wang, H. Correlation between auditory-vestibular functions and estrogen levels in postmenopausal patients with Meniere’s disease. J. Clin. Lab. Anal. 2019, 33, e22626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mijovic, T.; Zeitouni, A.; Colmegna, I. Autoimmune sensorineural hearing loss: The otology–rheumatology interface. Rheumatology 2013, 52, 780–789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciorba, A.; Corazzi, V.; Bianchini, C.; Aimoni, C.; Pelucchi, S.; Skarżyński, P.H.; Hatzopoulos, S. Autoimmune inner ear disease (AIED): A diagnostic challenge. Int. J. Immunopathol. Pharmacol. 2018, 32, 2058738418808680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voskuhl, R. Sex differences in autoimmune diseases. Biol. Sex Differ. 2011, 2, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Milon, B.; Mitra, S.; Song, Y.; Margulies, Z.; Casserly, R.; Drake, V.; Mong, J.A.; Depireux, D.A.; Hertzano, R. The impact of biological sex on the response to noise and otoprotective therapies against acoustic injury in mice. Biol. Sex Differ. 2018, 9, 1–14. [Google Scholar] [CrossRef]
- Guimaraes, P.; Zhu, X.; Cannon, T.; Kim, S.; Frisina, R.D. Sex differences in distortion product otoacoustic emissions as a function of age in CBA mice. Hear. Res. 2004, 192, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Henry, K.R. Sex-and age-related elevation of cochlear nerve envelope response (CNER) and auditory brainstem response (ABR) thresholds in C57BL/6 mice. Hear. Res. 2002, 170, 107–115. [Google Scholar] [CrossRef]
- Yager, D.D. Sexual dimorphism of auditory function and structure in praying mantises (Mantodea; Dictyoptera). J. Zool. 1990, 221, 517–537. [Google Scholar] [CrossRef]
- Vassilakis, P.N.; Meenderink, S.W.; Narins, P.M. Distortion product otoacoustic emissions provide clues to hearing mechanisms in the frog ear. J. Acoust. Soc. Am. 2004, 116, 3713–3726. [Google Scholar] [CrossRef]
- Gall, M.D.; Brierley, L.E.; Lucas, J.R. Species and sex effects on auditory processing in brown-headed cowbirds and red-winged blackbirds. Anim. Behav. 2011, 81, 973–982. [Google Scholar] [CrossRef]
- McFadden, D. Masculinization of the mammalian cochlea. Hear. Res. 2009, 252, 37–48. [Google Scholar] [CrossRef] [Green Version]
- McFadden, D.; Pasanen, E.G.; Raper, J.; Lange, H.S.; Wallen, K. Sex differences in otoacoustic emissions measured in rhesus monkeys (Macaca mulatta). Horm. Behav. 2006, 50, 274–284. [Google Scholar] [CrossRef] [PubMed]
- McFadden, D.; Pasanen, E.G.; Valero, M.D.; Roberts, E.K.; Lee, T.M. Effect of prenatal androgens on click-evoked otoacoustic emissions in male and female sheep (Ovis aries). Horm. Behav. 2009, 55, 98–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McFadden, S.L.; Zheng, X.-Y.; Ding, D.-L. Conditioning-induced protection from impulse noise in female and male chinchillas. J. Acoust. Soc. Am. 2000, 107, 2162–2168. [Google Scholar] [CrossRef] [PubMed]
- McFadden, D.; Pasanen, E.G.; Weldele, M.L.; Glickman, S.E.; Place, N.J. Masculinized otoacoustic emissions in female spotted hyenas (Crocuta crocuta). Horm. Behav. 2006, 50, 285–292. [Google Scholar] [CrossRef]
- Escabi, C.D.; Frye, M.D.; Trevino, M.; Lobarinas, E. The rat animal model for noise-induced hearing loss. J. Acoust. Soc. Am. 2019, 146, 3692–3709. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.D. Sex differences in the length of the organ of Corti in humans. J. Acoust. Soc. Am. 2007, 121, EL151–EL155. [Google Scholar] [CrossRef]
- Sato, H.; Sando, I.; Takahashi, H. Sexual dimorphism and development of the human cochlea: Computer 3-D measurement. Acta Oto-Laryngol. 1991, 111, 1037–1040. [Google Scholar] [CrossRef]
- Schrode, K.M.; Muniak, M.A.; Kim, Y.-H.; Lauer, A.M. Central compensation in auditory brainstem after damaging noise exposure. Eneuro 2018, 5. [Google Scholar] [CrossRef] [Green Version]
- McFadden, S.L.; Henselman, L.W.; Zheng, X.-Y. Sex differences in auditory sensitivity of chinchillas before and after exposure to impulse noise. Ear Hear. 1999, 20, 164–174. [Google Scholar] [CrossRef]
- Willott, J.F.; Bross, L. Effects of prolonged exposure to an augmented acoustic environment on the auditory system of middle-aged C57BL/6J mice: Cochlear and central histology and sex differences. J. Comp. Neurol. 2004, 472, 358–370. [Google Scholar] [CrossRef] [PubMed]
- Kirkim, G.; Olgun, Y.; Aktas, S.; Kiray, M.; Kolatan, E.; Altun, Z.; Erçetin, P.; Bagriyanik, A.; Yilmaz, O.; Ellidokuz, H. Is there a gender-related susceptibility for cisplatin ototoxicity? Eur. Arch. Otorhinolaryngol. 2015, 272, 2755–2763. [Google Scholar] [CrossRef]
- DeBacker, J.R.; Harrison, R.T.; Bielefeld, E.C. Cisplatin-induced threshold shift in the CBA/CaJ, C57BL/6J, BALB/cJ mouse models of hearing loss. Hear. Res. 2020, 387, 107878. [Google Scholar] [CrossRef] [PubMed]
- Mills, C.D.; Loos, B.M.; Henley, C.M. Increased susceptibility of male rats to kanamycin-induced cochleotoxicity. Hear. Res. 1999, 128, 75–79. [Google Scholar] [CrossRef]
- Halsey, K.; Skjönsberg, Å.; Ulfendahl, M.; Dolan, D.F. Efferent-mediated adaptation of the DPOAE as a predictor of aminoglycoside toxicity. Hear. Res. 2005, 201, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Henry, K.R.; McGinn, M.D. The mouse as a model for human audition. A review of the literature. Audiology 1992, 31, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Henry, K.R. Males lose hearing earlier in mouse models of late-onset age-related hearing loss; females lose hearing earlier in mouse models of early-onset hearing loss. Hear. Res. 2004, 190, 141–148. [Google Scholar] [CrossRef]
- Johnson, K.R.; Erway, L.C.; Cook, S.A.; Willott, J.F.; Zheng, Q.Y. A major gene affecting age-related hearing loss in C57BL/6J mice. Hear. Res. 1997, 114, 83–92. [Google Scholar] [CrossRef]
- Ohlemiller, K.K.; Dahl, A.R.; Gagnon, P.M. Divergent aging characteristics in CBA/J and CBA/CaJ mouse cochleae. J. Assoc. Res. Otolaryngol. 2010, 11, 605–623. [Google Scholar] [CrossRef] [Green Version]
- Balogová, Z.; Popelář, J.; Chiumenti, F.; Chumak, T.; Burianová, J.S.; Rybalko, N.; Syka, J. Age-related differences in hearing function and cochlear morphology between male and female Fischer 344 rats. Front. Aging Neurosci. 2018, 9, 428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shuster, B.Z.; Depireux, D.A.; Mong, J.A.; Hertzano, R. Sex differences in hearing: Probing the role of estrogen signaling. J. Acoust. Soc. Am. 2019, 145, 3656–3663. [Google Scholar] [CrossRef]
- Fetoni, A.R.; Picciotti, P.M.; Paludetti, G.; Troiani, D. Pathogenesis of presbycusis in animal models: A review. Exp. Gerontol. 2011, 46, 413–425. [Google Scholar] [CrossRef]
- Delhez, A.; Lefebvre, P.; Péqueux, C.; Malgrange, B.; Delacroix, L. Auditory function and dysfunction: Estrogen makes a difference. Cell Mol. Life Sci. 2020, 77, 619–635. [Google Scholar] [CrossRef]
- Dehan, C.P.; Jerger, J. Analysis of gender differences in the auditory brainstem response. Laryngoscope 1990, 100, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Bilger, R.C.; Matthies, M.L.; Hammel, D.R.; Demorest, M.E. Genetic implications of gender differences in the prevalence of spontaneous otoacoustic emissions. J. Speech Lang. Hear. Res. 1990, 33, 418–432. [Google Scholar] [CrossRef]
- Snihur, A.W.; Hampson, E. Sex and ear differences in spontaneous and click-evoked otoacoustic emissions in young adults. Brain Cogn. 2011, 77, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Adriztina, I.; Adnan, A.; Adenin, I.; Haryuna, S.H.; Sarumpaet, S. Influence of hormonal changes on audiologic examination in normal ovarian cycle females: An analytic study. Int. Arch. Otorhinolaryngol. 2016, 20, 294–299. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Kang, B.M.; Chae, H.D.; Kim, C.H. The association between serum estradiol level and hearing sensitivity in postmenopausal women. Obstet. Gynecol. 2002, 99, 726–730. [Google Scholar] [PubMed]
- McFadden, D. Masculinization effects in the auditory system. Arch. Sex. Behav. 2002, 31, 99–111. [Google Scholar] [CrossRef]
- Brann, D.W.; Dhandapani, K.; Wakade, C.; Mahesh, V.B.; Khan, M.M. Neurotrophic and neuroprotective actions of estrogen: Basic mechanisms and clinical implications. Steroids 2007, 72, 381–405. [Google Scholar] [CrossRef] [Green Version]
- Hederstierna, C.; Hultcrantz, M.; Rosenhall, U. Estrogen and hearing from a clinical point of view; characteristics of auditory function in women with Turner syndrome. Hear. Res. 2009, 252, 3–8. [Google Scholar] [CrossRef]
- Hultcrantz, M.; Stenberg, A.E.; Fransson, A.; Canlon, B. Characterization of hearing in an X, 0 ‘Turner mouse’. Hear. Res. 2000, 143, 182–188. [Google Scholar] [CrossRef]
- Heldring, N.; Pike, A.; Andersson, S.; Matthews, J.; Cheng, G.; Hartman, J.; Tujague, M.; Strom, A.; Treuter, E.; Warner, M. Estrogen receptors: How do they signal and what are their targets. Physiol. Rev. 2007, 87, 905–931. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, K.J.; Hewitt, S.C.; Arao, Y.; Korach, K.S. Estrogen Hormone Biology. Curr. Top. Dev. Biol. 2017, 125, 109–146. [Google Scholar]
- Yaşar, P.; Ayaz, G.; User, S.D.; Güpür, G.; Muyan, M. Molecular mechanism of estrogen–estrogen receptor signaling. Reprod. Med. Biol. 2017, 16, 4–20. [Google Scholar] [CrossRef] [PubMed]
- Stenberg, A.; Wang, H.; Fish Iii, J.; Schrott-Fischer, A.; Sahlin, L.; Hultcrantz, M. Estrogen receptors in the normal adult and developing human inner ear and in Turner’s syndrome. Hear. Res. 2001, 157, 87–92. [Google Scholar] [CrossRef]
- Motohashi, R.; Takumida, M.; Shimizu, A.; Konomi, U.; Fujita, K.; Hirakawa, K.; Suzuki, M.; Anniko, M. Effects of age and sex on the expression of estrogen receptor α and β in the mouse inner ear. Acta Oto-Laryngol. 2010, 130, 204–214. [Google Scholar] [CrossRef]
- Charitidi, K.; Canlon, B. Estrogen receptors in the central auditory system of male and female mice. Neuroscience 2010, 165, 923–933. [Google Scholar] [CrossRef]
- Unfer, T.; Figueiredo, C.; Zanchi, M.; Maurer, L.; Kemerich, D.; Duarte, M.; Konopka, C.; Emanuelli, T. Estrogen plus progestin increase superoxide dismutase and total antioxidant capacity in postmenopausal women. Climacteric 2015, 18, 379–388. [Google Scholar] [CrossRef]
- Rao, A.K.; Dietrich, A.K.; Ziegler, Y.S.; Nardulli, A.M. 17β-Estradiol-mediated increase in Cu/Zn superoxide dismutase expression in the brain: A mechanism to protect neurons from ischemia. J. Steroid Biochem. Mol. Biol. 2011, 127, 382–389. [Google Scholar] [CrossRef] [Green Version]
- Yune, T.Y.; Park, H.G.; Lee, J.Y.; Oh, T.H. Estrogen-induced Bcl-2 expression after spinal cord injury is mediated through phosphoinositide-3-kinase/Akt-dependent CREB activation. J. Neurotrauma 2008, 25, 1121–1131. [Google Scholar] [CrossRef]
- Pike, C.J. Estrogen Modulates Neuronal Bcl-xl Expression and β-Amyloid-Induced Apoptosis: Relevance to Alzheimer’s Disease. J. Neurochem. 1999, 72, 1552–1563. [Google Scholar] [CrossRef]
- Nakamagoe, M.; Tabuchi, K.; Uemaetomari, I.; Nishimura, B.; Hara, A. Estradiol protects the cochlea against gentamicin ototoxicity through inhibition of the JNK pathway. Hear. Res. 2010, 261, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Salvi, R.J. Brain derived neurotrophic factor and neurotrophic factor 3 modulate neurotransmitter receptor expressions on developing spiral ganglion neurons. Neuroscience 2009, 164, 1854–1866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meltser, I.; Tahera, Y.; Simpson, E.; Hultcrantz, M.; Charitidi, K.; Gustafsson, J.-Å.; Canlon, B. Estrogen receptor β protects against acoustic trauma in mice. J. Clin. Investig. 2008, 118, 1563–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singer, C.A.; Rogers, K.L.; Strickland, T.M.; Dorsa, D.M. Estrogen protects primary cortical neurons from glutamate toxicity. Neurosci. Lett. 1996, 212, 13–16. [Google Scholar] [CrossRef]
- Tadros, S.F.; D’Souza, M.; Zettel, M.L.; Zhu, X.; Waxmonsky, N.C.; Frisina, R.D. Glutamate-related gene expression changes with age in the mouse auditory midbrain. Brain Res. 2007, 1127, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Liberman, M.C.; Kujawa, S.G. Cochlear synaptopathy in acquired sensorineural hearing loss: Manifestations and mechanisms. Hear. Res. 2017, 349, 138–147. [Google Scholar] [CrossRef]
- Lee, J.H.; Marcus, D.C. Estrogen acutely inhibits ion transport by isolated stria vascularis. Hear. Res. 2001, 158, 123–130. [Google Scholar] [CrossRef]
- Brechtelsbauer, P.; Nuttall, A.; Miller, J. Basal nitric oxide production in regulation of cochlear blood flow. Hear. Res. 1994, 77, 38–42. [Google Scholar] [CrossRef]
- Guo, X.; Razandi, M.; Pedram, A.; Kassab, G.; Levin, E.R. Estrogen induces vascular wall dilation: Mediation through kinase signaling to nitric oxide and estrogen receptors α and β. J. Biol. Chem. 2005, 280, 19704–19710. [Google Scholar] [CrossRef] [Green Version]
- Andrews, J.C.; Honrubia, V. Premenstrual exacerbation of Meniere’s disease revisited. Otolaryngol. Clin. N. Am. 2010, 43, 1029–1040. [Google Scholar] [CrossRef]
- Vernier, M.; Dufour, C.R.; McGuirk, S.; Scholtes, C.; Li, X.; Bourmeau, G.; Kuasne, H.; Park, M.; St-Pierre, J.; Audet-Walsh, E.; et al. Estrogen-related receptors are targetable ROS sensors. Genes Dev. 2020, 34, 544–559. [Google Scholar] [CrossRef] [PubMed]
- Nolan, L.S.; Maier, H.; Hermans-Borgmeyer, I.; Girotto, G.; Ecob, R.; Pirastu, N.; Cadge, B.A.; Hübner, C.; Gasparini, P.; Strachan, D.P.; et al. Estrogen-related receptor gamma and hearing function: Evidence of a role in humans and mice. Neurobiol. Aging 2013, 34, 2077.e1–2077.e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schilit, S.L.; Currall, B.B.; Yao, R.; Hanscom, C.; Collins, R.L.; Pillalamarri, V.; Lee, D.Y.; Kammin, T.; Zepeda-Mendoza, C.J.; Mononen, T.; et al. Estrogen-related receptor gamma implicated in a phenotype including hearing loss and mild developmental delay. Eur. J. Hum. Genet. 2016, 24, 1622–1626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.; Wang, Y.; Lau, C.-C. Effects of Noise Exposure on the Auditory Function of Ovariectomized Rats with Estrogen Deficiency. J. Int. Adv. Otol. 2016, 12, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.-J.; Li, F.-F.; Wang, Y.; Lau, C.-C. Effects of cisplatin on the auditory function of ovariectomized rats with estrogen deficiency. Acta Oto-Laryngol. 2017, 137, 606–610. [Google Scholar] [CrossRef] [PubMed]
- Thakur, M.; Sharma, P. Transcription of estrogen receptor α and β in mouse cerebral cortex: Effect of age, sex, 17β-estradiol and testosterone. Neurochem. Int. 2007, 50, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Wharton, J.A.; Church, G.T. Influence of menopause on the auditory brainstem response. Audiology 1990, 29, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Pessina, M.A.; Hoyt Jr, R.F.; Goldstein, I.; Traish, A.M. BASIC SCIENCE: Differential Regulation of the Expression of Estrogen, Progesterone, and Androgen Receptors by Sex Steroid Hormones in the Vagina: Immunohistochemical Studies. J. Sex. Med. 2006, 3, 804–814. [Google Scholar] [CrossRef]
- Bass, A.H.; McKibben, J.R. Neural mechanisms and behaviors for acoustic communication in teleost fish. Prog. Neurobiol. 2003, 69, 1–26. [Google Scholar] [CrossRef]
- Forlano, P.M.; Marchaterre, M.; Deitcher, D.L.; Bass, A.H. Distribution of androgen receptor mRNA expression in vocal, auditory, and neuroendocrine circuits in a teleost fish. J. Comp. Neurol. 2010, 518, 493–512. [Google Scholar] [CrossRef] [Green Version]
- Miranda, J.A.; Wilczynski, W. Sex differences and androgen influences on midbrain auditory thresholds in the green treefrog, Hyla cinerea. Hear. Res. 2009, 252, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Oghan, F.; Coksuer, H. Does hyperandrogenism have an effect on hearing loss in patients with polycystic ovary syndrome? Auris Nasus Larynx 2012, 39, 365–368. [Google Scholar] [CrossRef] [PubMed]
- Turan, M.; Ucler, R.; Garca, M.F.; Kurdoglu, Z.; Cankaya, H.; Ayral, A.; Bozan, N.; Kıroglu, A.F.; Kurdoglu, M. The relationship between hearing thresholds and hyperandrogenism in polycystic ovary syndrome. Med Sci. Monit. Int. Med. J. Exp. Clin. Res. 2016, 22, 4380. [Google Scholar] [CrossRef] [Green Version]
- Kucur, C.; Kucur, S.K.; Gozukara, I.; Seven, A.; Yuksel, K.B.; Keskin, N.; Oghan, F. Extended high frequency audiometry in polycystic ovary syndrome. Sci. World J. 2013, 2013, 482689. [Google Scholar] [CrossRef] [Green Version]
- Cherian, K.E.; Kapoor, N.; Mathews, S.S.; Paul, T.V. Endocrine glands and hearing: Auditory manifestations of various endocrine and metabolic conditions. Indian J. Endocrinol. Metab. 2017, 21, 464. [Google Scholar]
- Eren, E.; Harman, E.; Arslanoglu, S.; Önal, K.; Katlmis, H. Does hyperandrogenism affect the otoacoustic emissions and medial olivocochlear reflex in female adults? Otol. Neurotol. 2013, 34, 784–789. [Google Scholar] [CrossRef]
- Chun, K.-J.; Lee, C.-H.; Kim, K.-W.; Lee, S.-M.; Kim, S.-Y. Effects of Androgen Receptor Inhibition on Kanamycin-Induced Hearing Loss in Rats. Int. J. Mol. Sci. 2021, 22, 5307. [Google Scholar] [CrossRef]
- He, Z.-Y.; Ren, D.-D. Sex hormones and inner ear. In Sex Hormones in Neurodegenerative Processes and Diseases; IntechOpen: London, UK, 2018; pp. 329–346. [Google Scholar]
- Köşüş, N.; Köşüş, A.; Turhan, N.; Kurtaran, H. Hearing levels in menopausal women and the effect of tibolone on audiological functions. J. Obstet. Gynaecol. 2012, 32, 294–297. [Google Scholar] [CrossRef] [PubMed]
- Kilicdag, E.B.; Yavuz, H.; Bagis, T.; Tarim, E.; Erkan, A.N.; Kazanci, F. Effects of estrogen therapy on hearing in postmenopausal women. Am. J. Obstet. Gynecol. 2004, 190, 77–82. [Google Scholar] [CrossRef]
- Vrtačnik, P.; Ostanek, B.; Mencej-Bedrač, S.; Marc, J. The many faces of estrogen signaling. Biochem. Med. 2014, 24, 329–342. [Google Scholar] [CrossRef] [Green Version]
- Guimaraes, P.; Frisina, S.T.; Mapes, F.; Tadros, S.F.; Frisina, D.R.; Frisina, R.D. Progestin negatively affects hearing in aged women. Proc. Natl. Acad. Sci. USA 2006, 103, 14246–14249. [Google Scholar] [CrossRef] [Green Version]
- Price, K.; Zhu, X.; Guimaraes, P.F.; Vasilyeva, O.N.; Frisina, R.D. Hormone replacement therapy diminishes hearing in peri-menopausal mice. Hear. Res. 2009, 252, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Curhan, S.G.; Eliassen, A.H.; Eavey, R.D.; Wang, M.; Lin, B.M.; Curhan, G.C. Menopause and postmenopausal hormone therapy and risk of hearing loss. Menopause 2017, 24, 1049. [Google Scholar] [CrossRef] [PubMed]
- Williamson, T.T.; Ding, B.; Zhu, X.; Frisina, R.D. Hormone replacement therapy attenuates hearing loss: Mechanisms involving estrogen and the IGF-1 pathway. Aging Cell 2019, 18, e12939. [Google Scholar] [CrossRef] [PubMed]
- Frisina, R.; Bazard, P.; Bauer, M.; Pineros, J.; Zhu, X.; Ding, B. Translational implications of the interactions between hormones and age-related hearing loss. Hear. Res. 2020, 402, 108093. [Google Scholar] [CrossRef] [PubMed]
- Frisina, R.; Budzevich, M.; Zhu, X.; Martinez, G.; Walton, J.; Borkholder, D. Animal model studies yield translational solutions for cochlear drug delivery. Hear. Res. 2018, 368, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Altevogt, B.M.; Wizemann, T.; Pankevich, D.E. Sex Differences and Implications for Translational Neuroscience Research: Workshop Summary; National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Ho, C.H.; Tan, T.Y.; Hwang, C.F.; Lin, W.C.; Wu, C.N.; Yang, C.H. Association of carotid intima-media thickness with the risk of sudden sensorineural hearing loss. PeerJ 2020, 8, e9276. [Google Scholar] [CrossRef] [PubMed]
- Villavisanis, D.F.; Schrode, K.M.; Lauer, A.M. Sex bias in basic and preclinical age-related hearing loss research. Biol. Sex Differ. 2018, 9, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Villavisanis, D.F.; Berson, E.R.; Lauer, A.M.; Cosetti, M.K.; Schrode, K.M. Sex-based differences in hearing loss: Perspectives from non-clinical research to clinical outcomes. Otol. Neurotol. Off Publ. Am. Otol. Soc. Am. Neurotol. Soc. Eur. Acad. Otol. Neurotol. 2020, 41, 290. [Google Scholar] [CrossRef]
- Corazzi, V.; Ciorba, A.; Skarżyński, P.H.; Skarżyńska, M.B.; Bianchini, C.; Stomeo, F.; Bellini, T.; Pelucchi, S.; Hatzopoulos, S. Gender differences in audio-vestibular disorders. Int. J. Immunopathol. Pharmacol. 2020, 34, 2058738420929174. [Google Scholar] [CrossRef]
- Lin, N.; Urata, S.; Cook, R.; Makishima, T. Sex differences in the auditory functions of rodents. Hear. Res. 2021, 108271. [Google Scholar] [CrossRef]
- Nolan, L.S. Age-related hearing loss: Why we need to think about sex as a biological variable. J. Neurosci. Res. 2020, 98, 1705–1720. [Google Scholar] [CrossRef] [PubMed]
- Nourbakhsh, A.; Colbert, B.M.; Nisenbaum, E.; El-Amraoui, A.; Dykxhoorn, D.M.; Koehler, K.R.; Chen, Z.-Y.; Liu, X.Z. Stem Cells and Gene Therapy in Progressive Hearing Loss: The State of the Art. J. Assoc. Res. Otolaryngol. 2021, 22, 95–105. [Google Scholar] [CrossRef]
- Chorath, K.; Willis, M.; Morton-Gonzaba, N.; Moreira, A. Mesenchymal stem cells for sensorineural hearing loss: A systematic review of preclinical studies. Mol. Biol. Rep. 2020, 47, 4723–4736. [Google Scholar] [CrossRef]
- Williams, J.; Mariya, S.; Suparto, I. Gender, age and differences in stem cell expression and efficacy. J. Stem Cell Res. 2017, 3, 00097. [Google Scholar] [CrossRef]
- Zullo, S.; Caenazzo, L. Gene editing and gender-specific medicine: A challenge for dementia research. Palgrave Commun. 2020, 6, 1–6. [Google Scholar] [CrossRef]
- Bean, L.A.; Kumar, A.; Rani, A.; Guidi, M.; Rosario, A.M.; Cruz, P.E.; Golde, T.E.; Foster, T.C. Re-opening the critical window for estrogen therapy. J. Neurosci. 2015, 35, 16077–16093. [Google Scholar] [CrossRef] [PubMed] [Green Version]

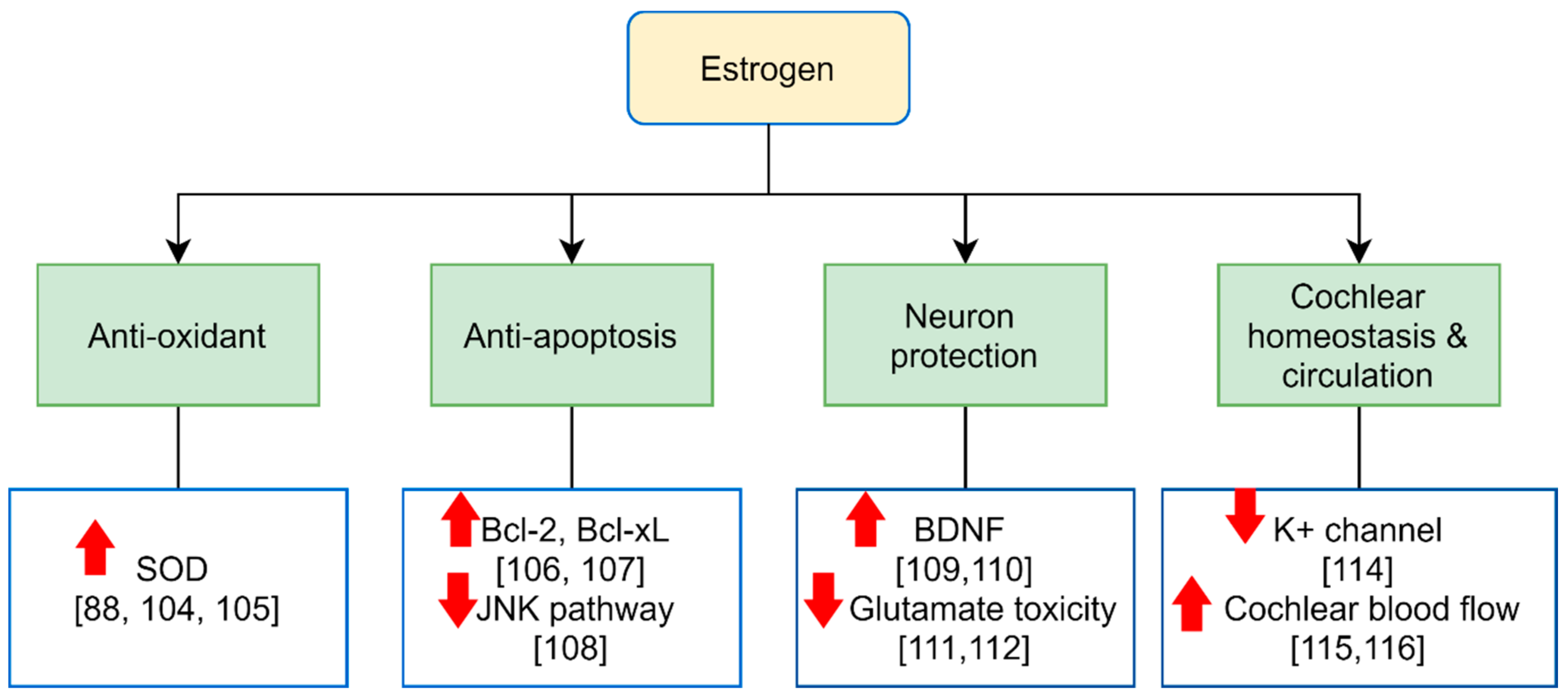{kind=link}
| Noise (NIHL) | Drugs (Ototoxicity) | Age (ARHL) | |
|---|---|---|---|
| Clinical aspects | Females had better hearing [7,8,9,10,11] | Aminoglycosides: females had a higher risk [23] Cisplatin: males had a higher risk [24,25,26,27] or no gender difference [29] | Males had a higher prevalence [36,37,38,39,40,42,43] |
| Animal investigations | B6CBAF1/J mice: females had a reduced permanent threshold shift [60] CBA/CaJ mice: no sex difference [74] Chincillas: females had less hair cell loss [75] C57BL/6J mice: females had more hair cell loss [76] | Cisplatin: Wistar albino rats: females had more apoptotic spiral ganglion neurons [77] CBA/CaJ mice: no sex difference [78] C57BL/6J mice: females had higher threshold shifts [78] BALB/cJ mice: males had higher threshold shifts [78] Aminoglycosides: females had better OAE in the Long–Evans rats [79] and ABR in guinea pigs [80] | CBA mice: females had better hearing during aging [61] CBA/J and CBA/CaJ mice: males had higher high- frequency ABR thresholds in late-onset ARHL [82] |
| Estrogen effect | ERβ agonist reduced the temporary threshold shift after acoustic trauma in mice [110] | Cisplatin: low estrogen increased ABR thresholds [122] Aminoglycosides: estrogen protected against outer hair cell death [108] | Elderly female mice had higher ERβ levels than males and preserved better hearing function [102] Postmenopausal women who had a higher serum estradiol level had better pure tone thresholds [93] and estrogen supplementation helped delay hearing loss [137] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lien, K.-H.; Yang, C.-H. Sex Differences in the Triad of Acquired Sensorineural Hearing Loss. Int. J. Mol. Sci. 2021, 22, 8111. https://doi.org/10.3390/ijms22158111
Lien K-H, Yang C-H. Sex Differences in the Triad of Acquired Sensorineural Hearing Loss. International Journal of Molecular Sciences. 2021; 22(15):8111. https://doi.org/10.3390/ijms22158111
Chicago/Turabian StyleLien, Kuang-Hsu, and Chao-Hui Yang. 2021. "Sex Differences in the Triad of Acquired Sensorineural Hearing Loss" International Journal of Molecular Sciences 22, no. 15: 8111. https://doi.org/10.3390/ijms22158111
APA StyleLien, K. -H., & Yang, C. -H. (2021). Sex Differences in the Triad of Acquired Sensorineural Hearing Loss. International Journal of Molecular Sciences, 22(15), 8111. https://doi.org/10.3390/ijms22158111