
Cell Types and Synapses Expressing the SNARE Complex Regulating Proteins Complexin 1 and Complexin 2 in Mammalian Retina

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Results
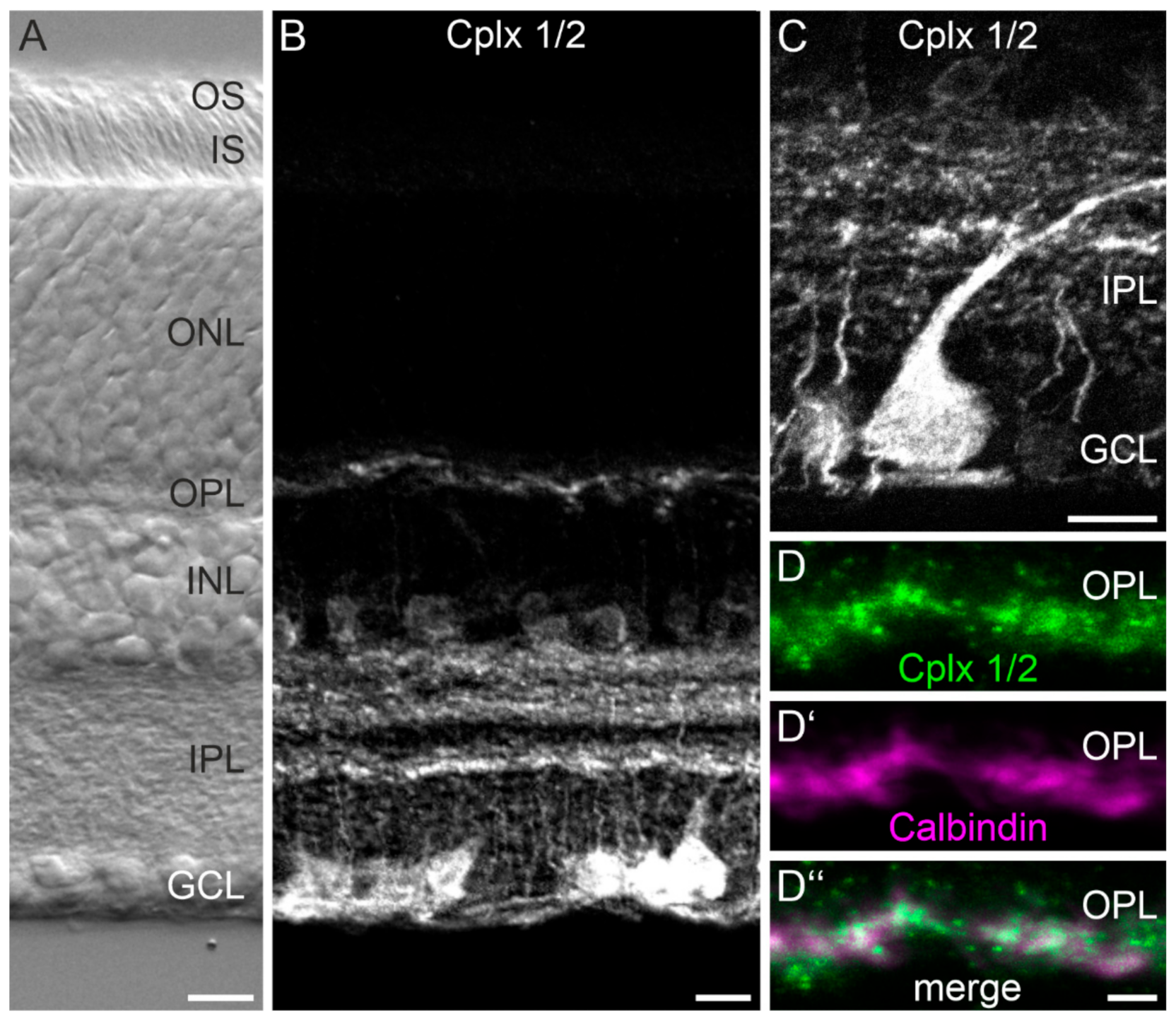
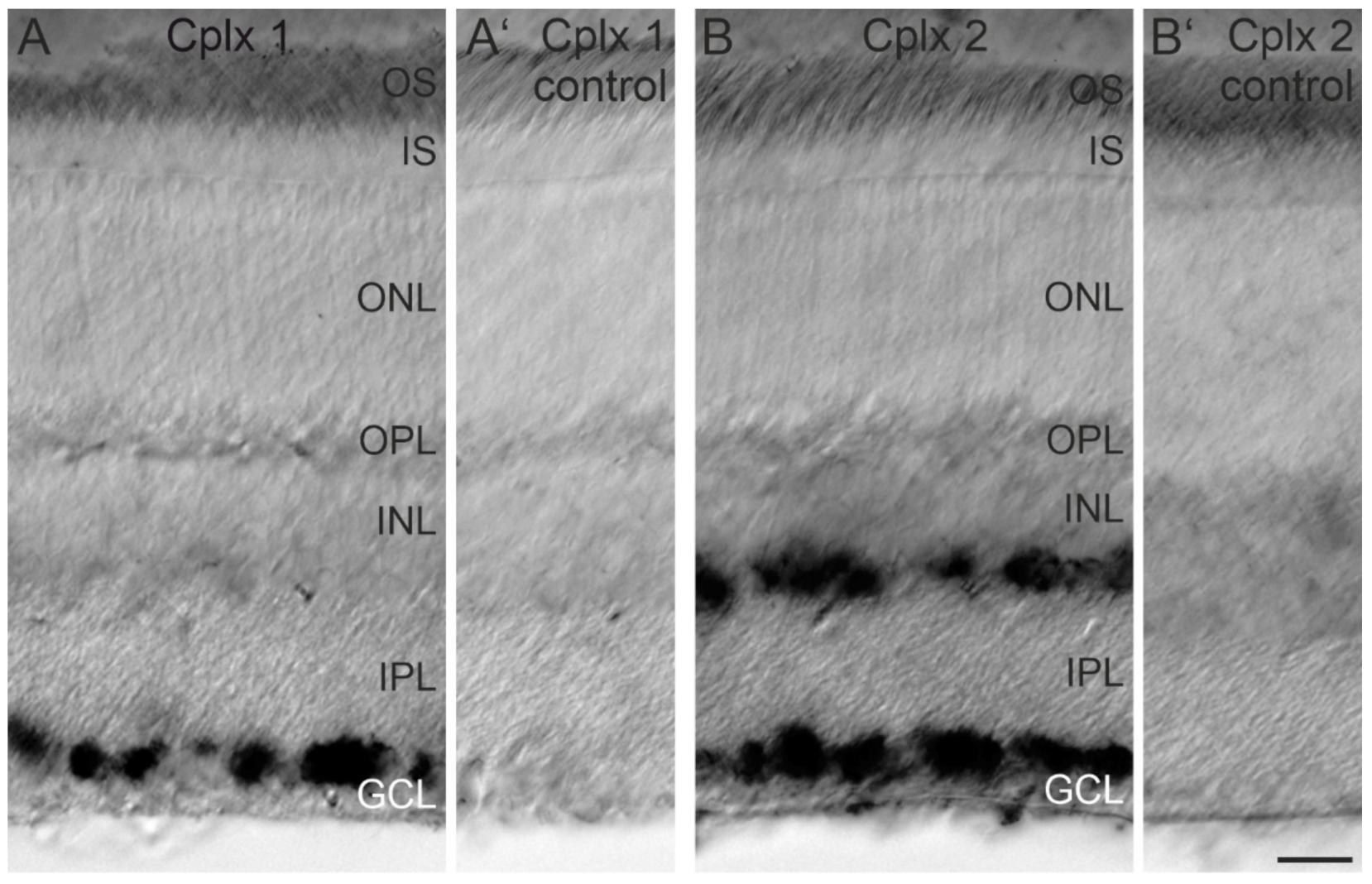
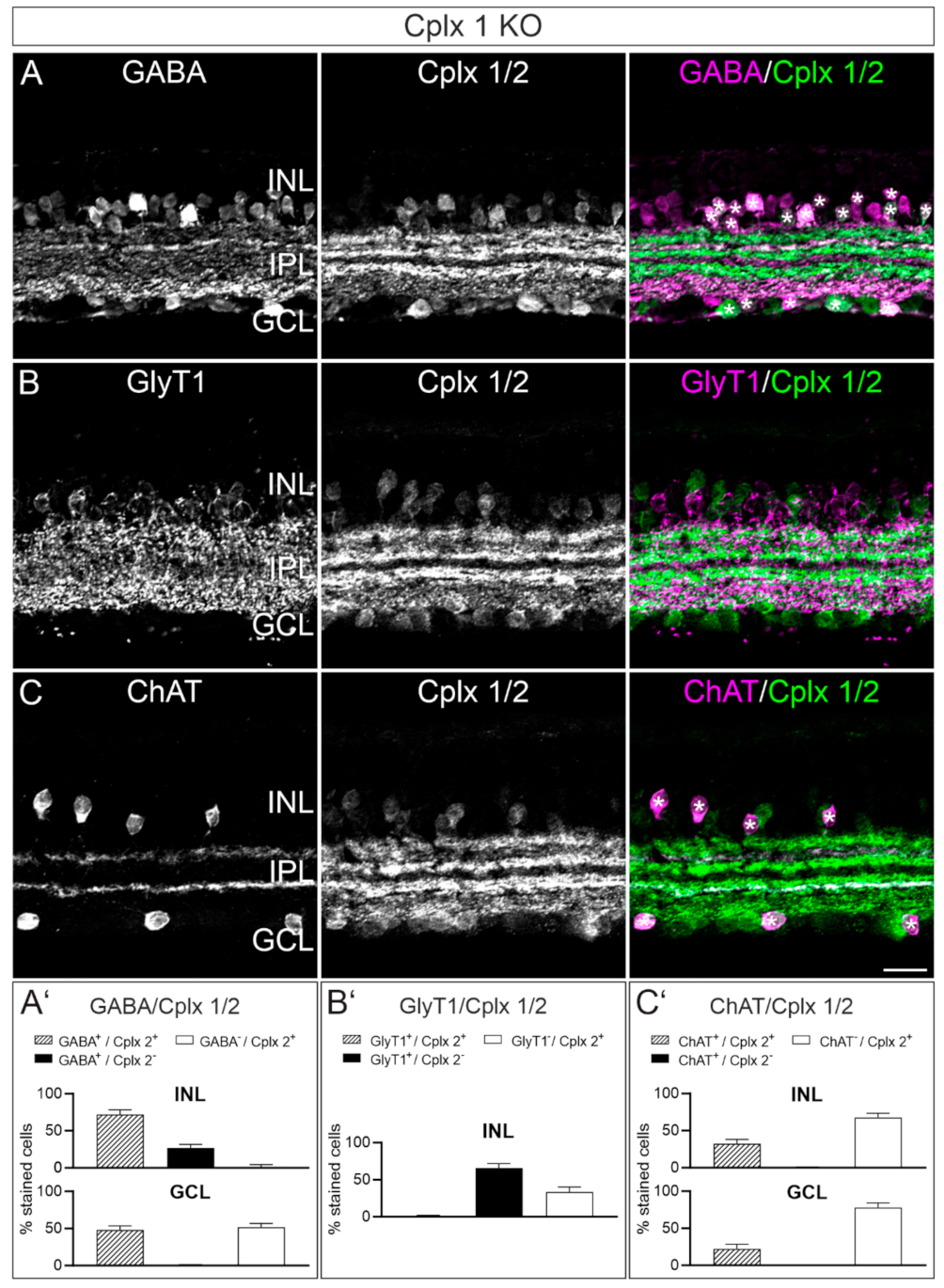
2.1. Cplx 2 but Not Cplx 1 Is Expressed by Cells in the Inner Nuclear Layer in Mouse Retina
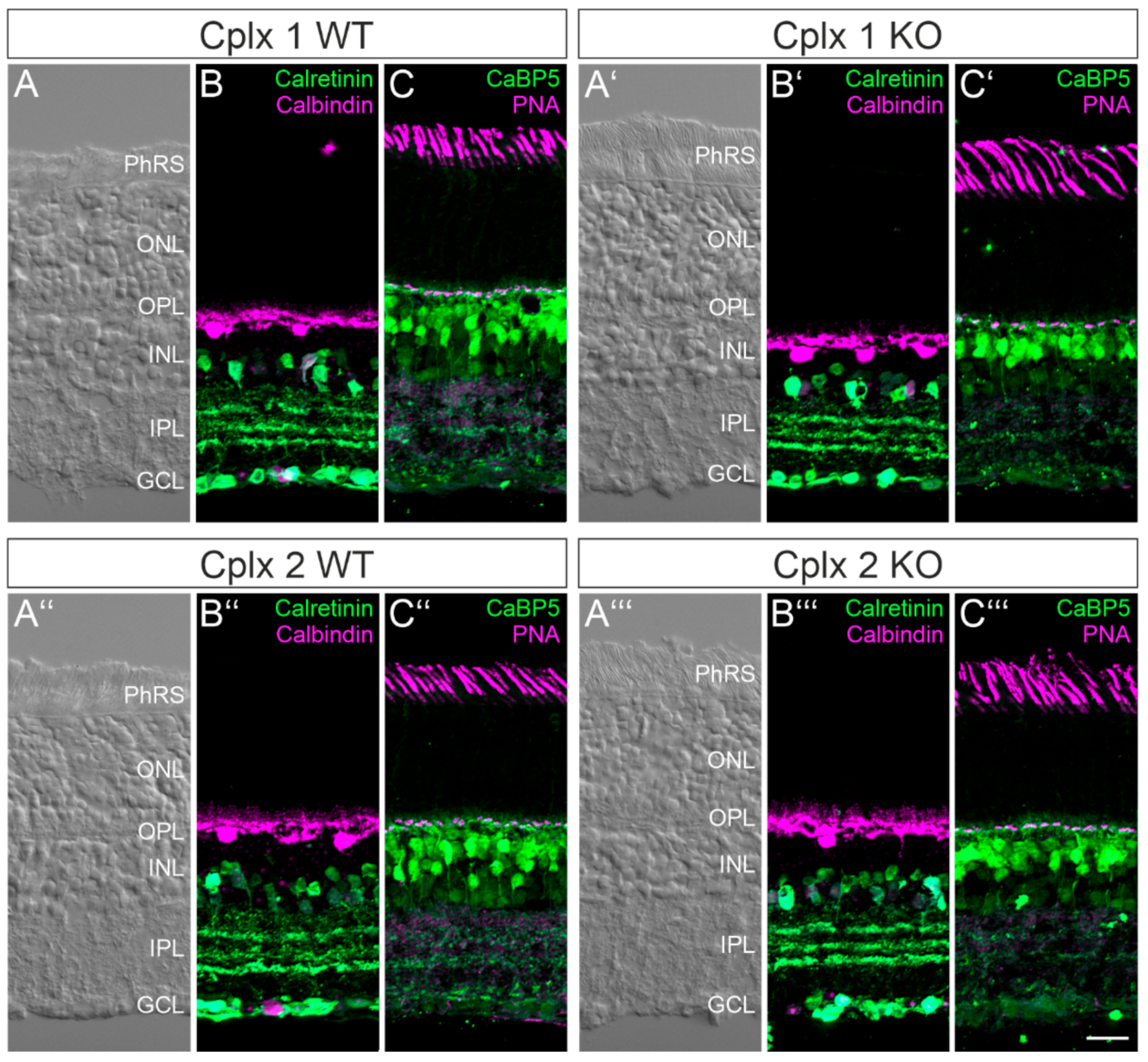
2.2. Retinal Structure Is Unaltered in Cplx 1 and Cplx 2 KO Mice
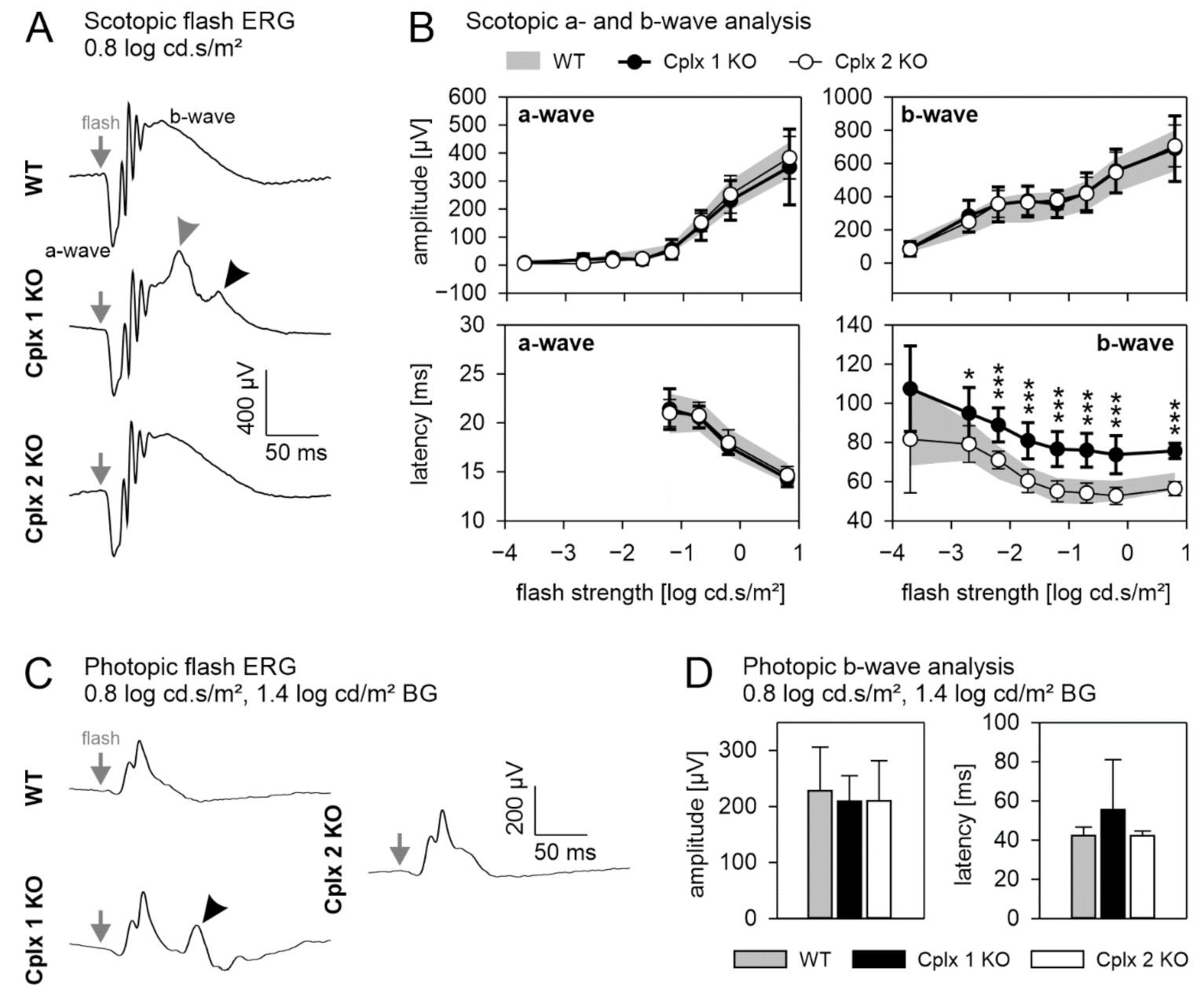
2.3. Retinal Function Is Altered in Cplx 1 but Not in Cplx 2 KO Mice
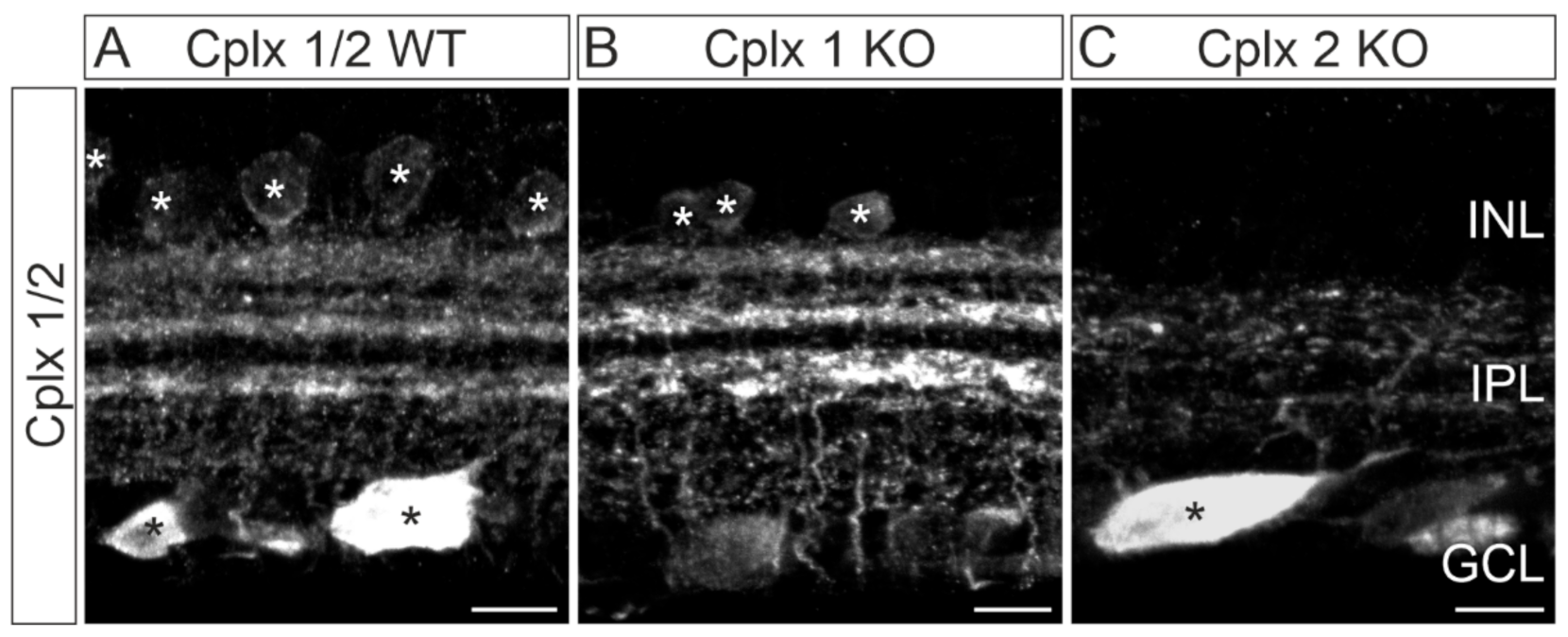
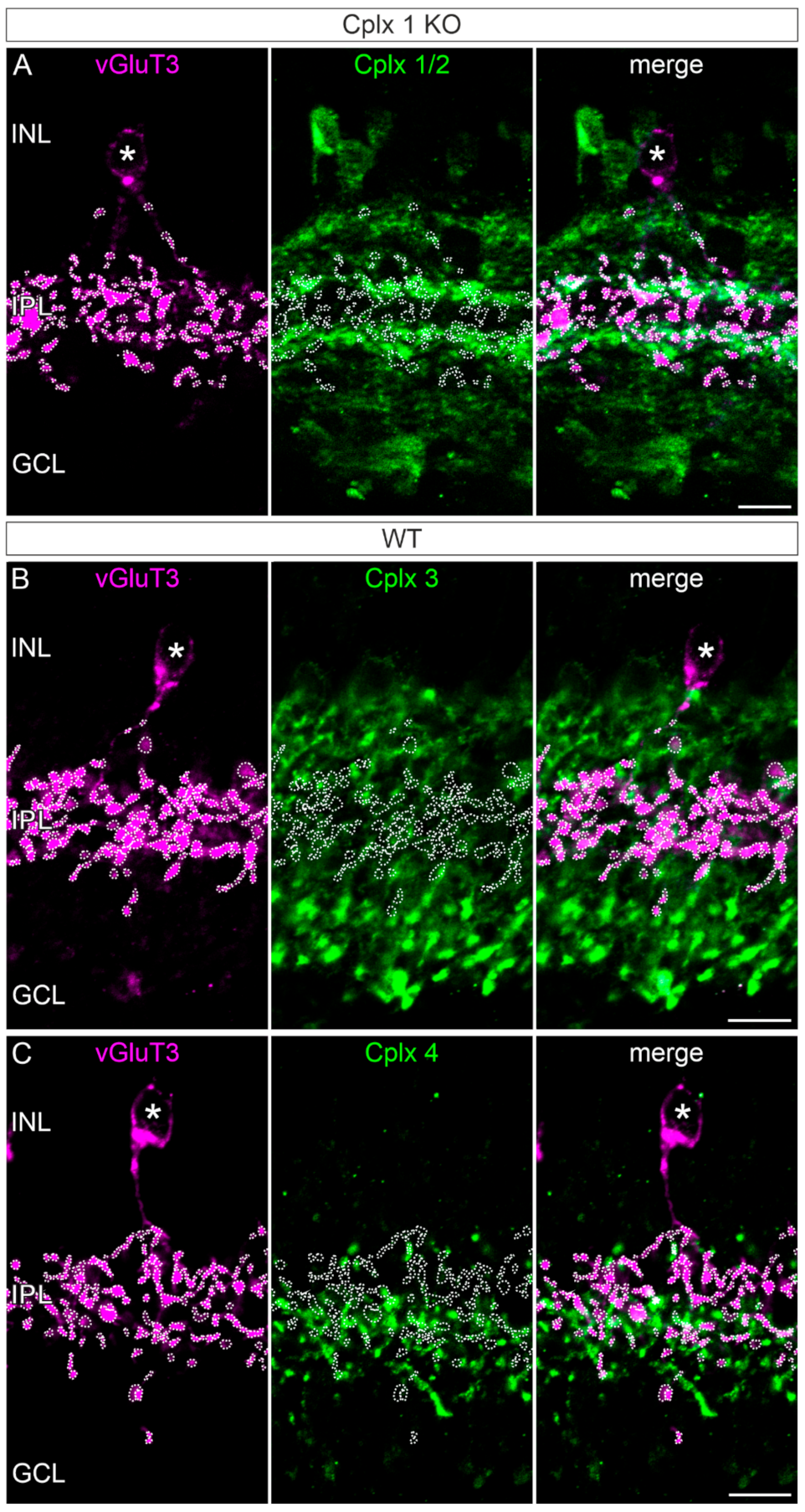
2.4. Cplx 2 Expressing Amacrine Cell Types in Mouse Retina
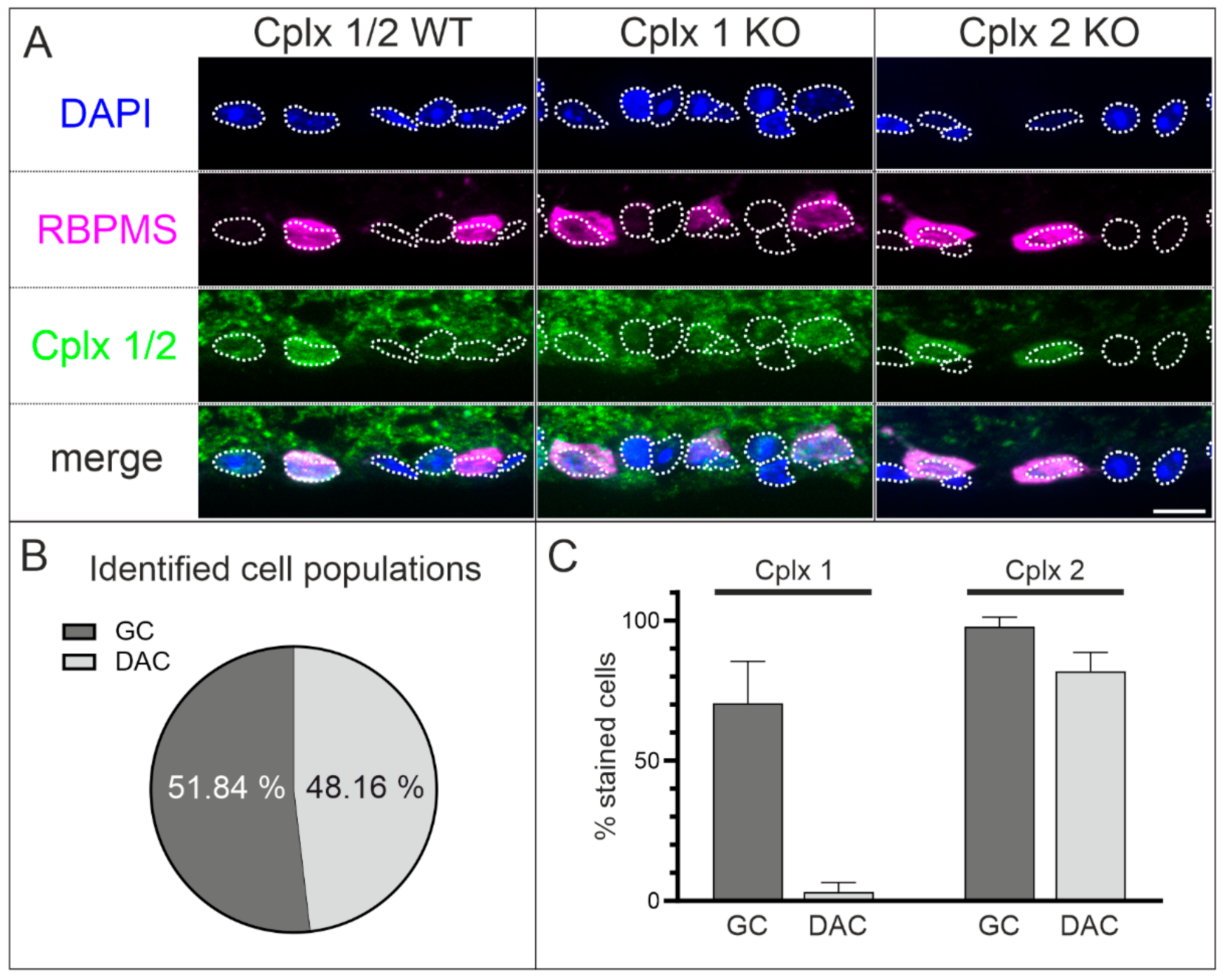
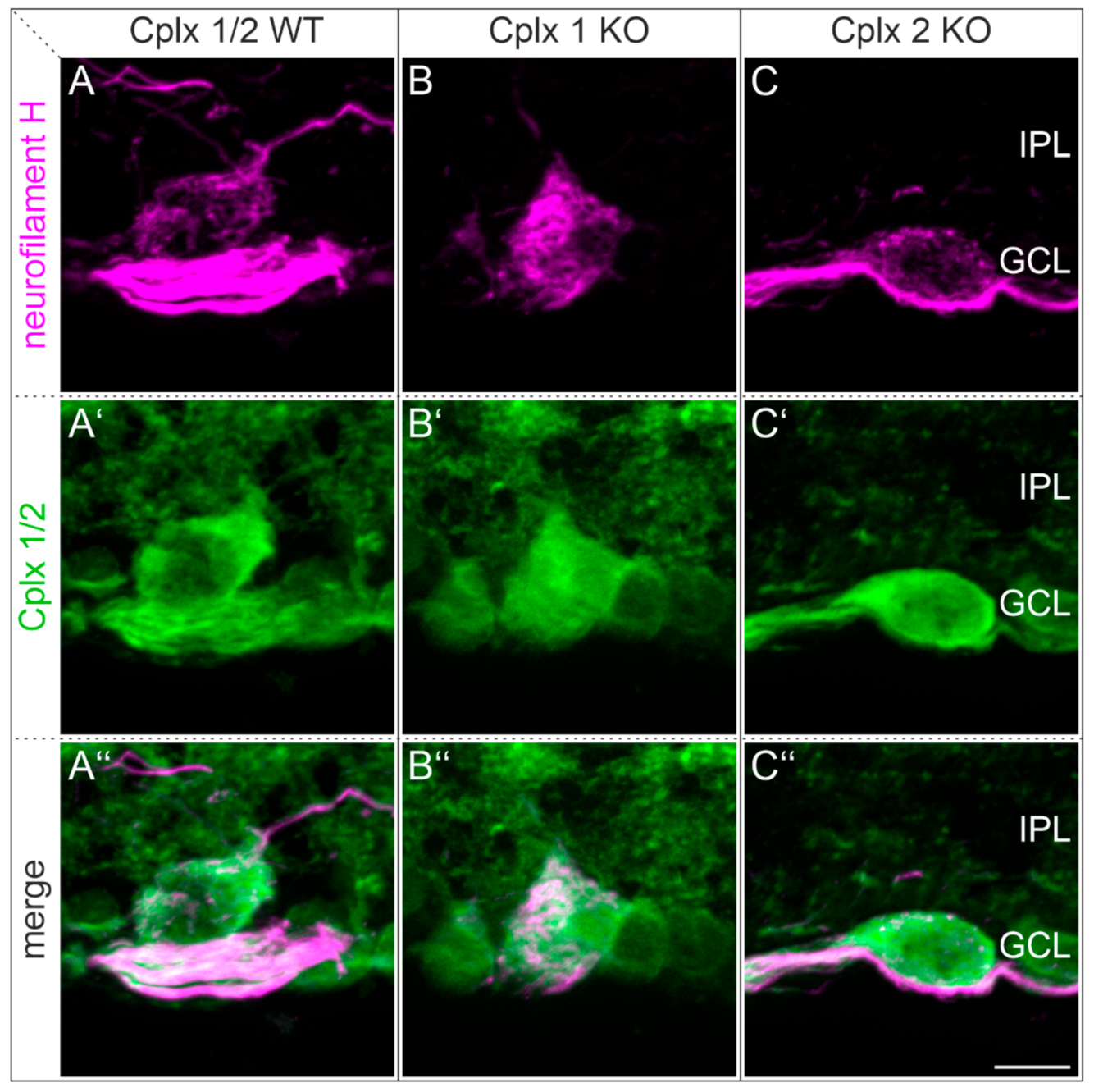
2.5. Cplx 1 and Cplx 2 Expressing Cell Types in the Ganglion Cell Layer in Mouse Retinae
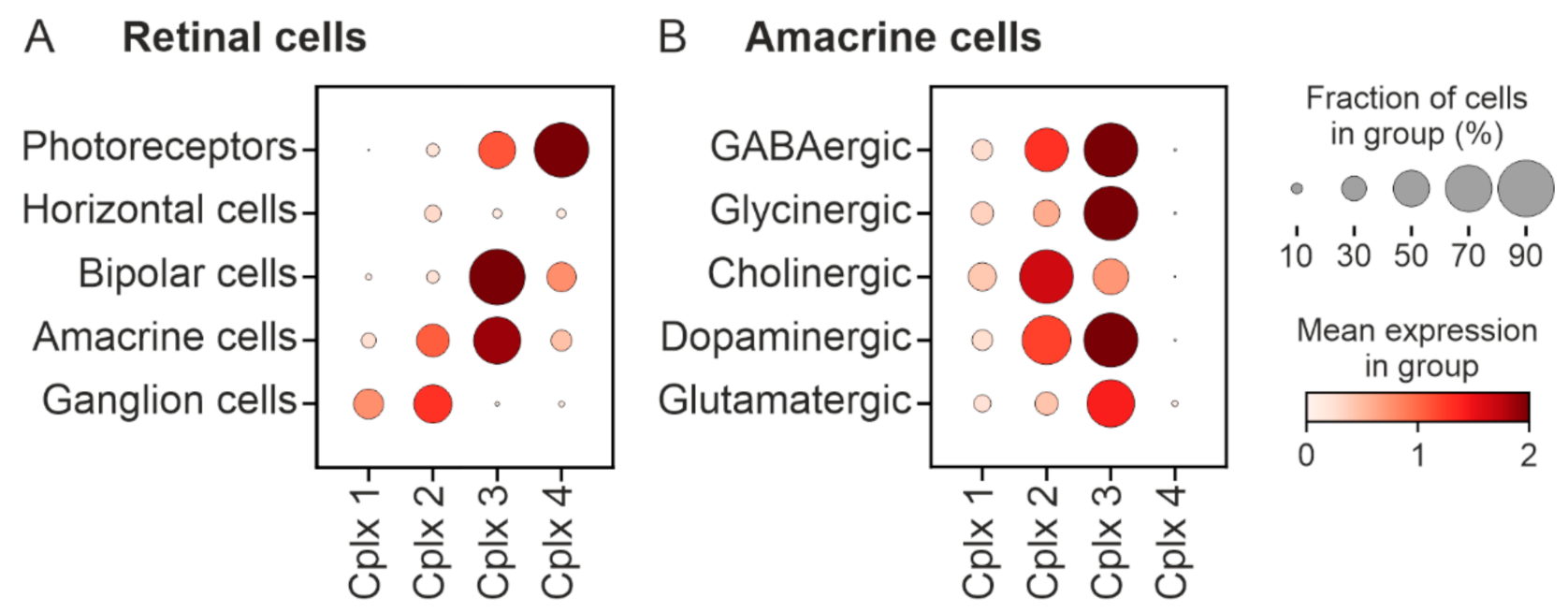
2.6. Bioinformatics Analysis of the Cell Types Expressing the Genes Encoding Cplx 1 and Cplx 2 in Mouse Retina
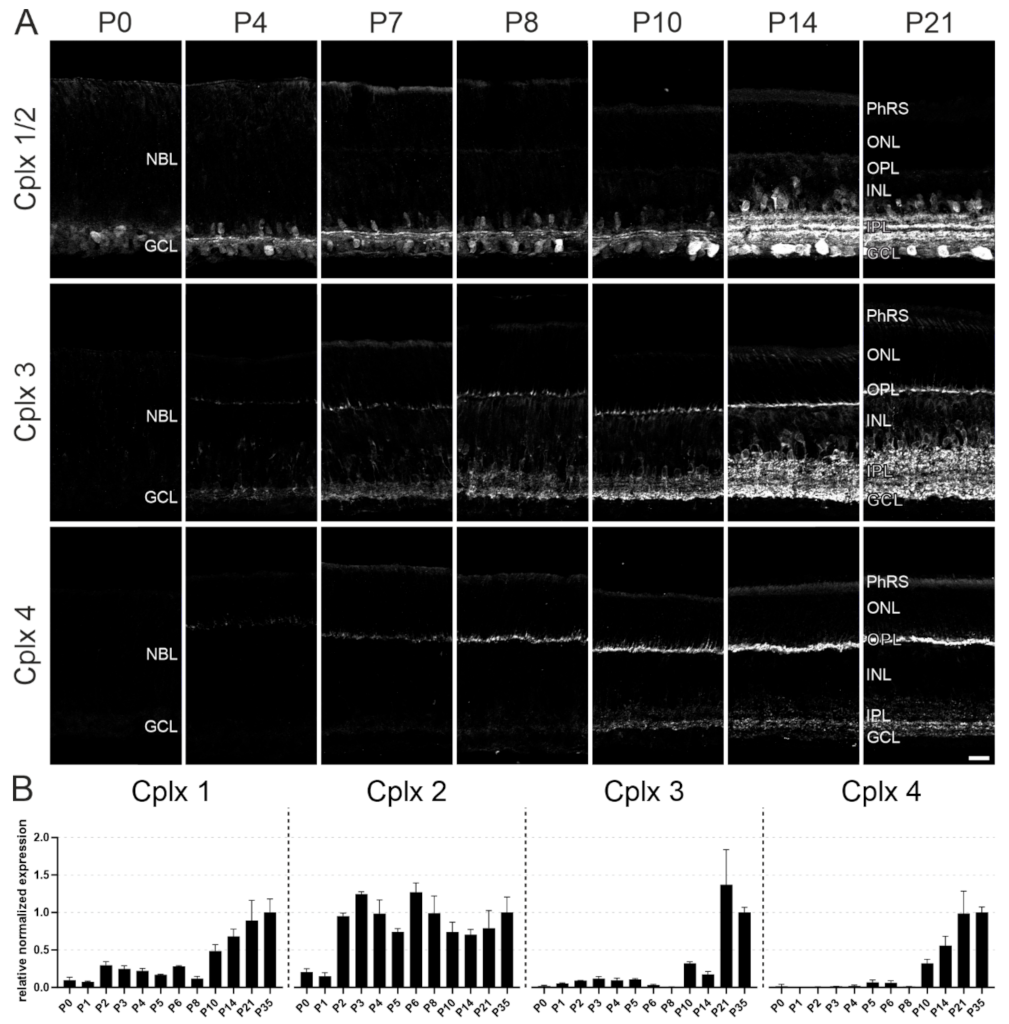
2.7. Postnatal Developmental Expression of the Cplxs 1 to 4 in Mouse Retina
3. Discussion
3.1. Presence of Cplx 2 but Not Cplx 1 at Conventional Chemical Synapses in Mouse Retinae
3.2. Presence of Cplx 1 and Cplx 2 in Ganglion Cells in Mouse Retina
4. Materials and Methods
4.1. Animals
4.2. Antibodies
4.3. Immunocytochemistry
4.4. Light Microscopy and Analysis of Immunocytochemical Data
4.5. Electroretinographic (ERG) Recordings and Data Analysis
4.6. In Situ Hybridization
4.7. RT-qPCR
4.8. Bioinformatics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AC | amacrine cell |
| DAC | displaced amacrine cell |
| GC | ganglion cell |
| GCL | ganglion cell layer |
| INL | inner nuclear layer |
| IPL | inner plexiform layer |
| KO | knock-out |
| ONL | outer nuclear layer |
| OP | oscillatory potentials |
| OPL | outer plexiform layer |
| PhRS | photoreceptor segments |
References
- Jahn, R.; Scheller, R.H. SNAREs—Engines for Membrane Fusion. Nat. Rev. Mol. Cell Biol. 2006, 7, 631–643. [Google Scholar] [CrossRef]
- Reim, K. Complexins. In Reference Module in Neuroscience and Biobehavioral Psychology; Elsevier: Amsterdam, The Netherlands, 2017; pp. 1–17. ISBN 978-0-12-809324-5. [Google Scholar]
- McMahon, H.T.; Missler, M.; Li, C.; Südhof, T.C. Complexins: Cytosolic Proteins That Regulate SNAP Receptor Function. Cell 1995, 83, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Reim, K.; Wegmeyer, H.; Brandstätter, J.H.; Xue, M.; Rosenmund, C.; Dresbach, T.; Hofmann, K.; Brose, N. Structurally and Functionally Unique Complexins at Retinal Ribbon Synapses. J. Cell Biol. 2005, 169, 669–680. [Google Scholar] [CrossRef] [Green Version]
- tom Dieck, S.; Brandstätter, J.H. Ribbon Synapses of the Retina. Cell Tissue Res. 2006, 326, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Thoreson, W.B.; Rabl, K.; Townes-Anderson, E.; Heidelberger, R. A Highly Ca2+-Sensitive Pool of Vesicles Contributes to Linearity at the Rod Photoreceptor Ribbon Synapse. Neuron 2004, 42, 595–605. [Google Scholar] [CrossRef] [Green Version]
- tom Dieck, S.; Altrock, W.D.; Kessels, M.M.; Qualmann, B.; Regus, H.; Brauner, D.; Fejtová, A.; Bracko, O.; Gundelfinger, E.D.; Brandstätter, J.H. Molecular Dissection of the Photoreceptor Ribbon Synapse: Physical Interaction of Bassoon and RIBEYE Is Essential for the Assembly of the Ribbon Complex. J. Cell Biol. 2005, 168, 825–836. [Google Scholar] [CrossRef] [Green Version]
- Sterling, P.; Matthews, G. Structure and Function of Ribbon Synapses. Trends Neurosci. 2005, 28, 20–29. [Google Scholar] [CrossRef]
- Reim, K.; Regus-Leidig, H.; Ammermuller, J.; El-Kordi, A.; Radyushkin, K.; Ehrenreich, H.; Brandstatter, J.H.; Brose, N. Aberrant Function and Structure of Retinal Ribbon Synapses in the Absence of Complexin 3 and Complexin 4. J. Cell Sci. 2009, 122, 1352–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babai, N.; Sendelbeck, A.; Regus-Leidig, H.; Fuchs, M.; Mertins, J.; Reim, K.; Brose, N.; Feigenspan, A.; Brandstätter, J.H. Functional Roles of Complexin 3 and Complexin 4 at Mouse Photoreceptor Ribbon Synapses. J. Neurosci. 2016, 36, 6651–6667. [Google Scholar] [CrossRef] [Green Version]
- Mortensen, L.S.; Park, S.J.H.; Ke, J.; Cooper, B.H.; Zhang, L.; Imig, C.; Löwel, S.; Reim, K.; Brose, N.; Demb, J.B.; et al. Complexin 3 Increases the Fidelity of Signaling in a Retinal Circuit by Regulating Exocytosis at Ribbon Synapses. Cell Rep. 2016, 15, 2239–2250. [Google Scholar] [CrossRef] [Green Version]
- Vaithianathan, T.; Zanazzi, G.; Henry, D.; Akmentin, W.; Matthews, G. Stabilization of Spontaneous Neurotransmitter Release at Ribbon Synapses by Ribbon-Specific Subtypes of Complexin. J. Neurosci. 2013, 33, 8216–8226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaithianathan, T.; Henry, D.; Akmentin, W.; Matthews, G. Functional Roles of Complexin in Neurotransmitter Release at Ribbon Synapses of Mouse Retinal Bipolar Neurons. J. Neurosci. 2015, 35, 4065–4070. [Google Scholar] [CrossRef] [PubMed]
- Reim, K.; Mansour, M.; Varoqueaux, F.; McMahon, H.T.; Südhof, T.C.; Brose, N.; Rosenmund, C. Complexins Regulate a Late Step in Ca2+-Dependent Neurotransmitter Release. Cell 2001, 104, 71–81. [Google Scholar] [CrossRef] [Green Version]
- Rogers, J.H. Calretinin: A Gene for a Novel Calcium-Binding Protein Expressed Principally in Neurons. J. Cell Biol. 1987, 105, 1343–1353. [Google Scholar] [CrossRef] [Green Version]
- Haverkamp, S.; Wässle, H. Immunocytochemical Analysis of the Mouse Retina. J. Comp. Neurol. 2000, 424, 1–23. [Google Scholar] [CrossRef]
- Haeseleer, F.; Sokal, I.; Verlinde, C.L.M.J.; Erdjument-Bromage, H.; Tempst, P.; Pronin, A.N.; Benovic, J.L.; Fariss, R.N.; Palczewski, K. Five Members of a Novel Ca2+-Binding Protein (CABP) Subfamily with Similarity to Calmodulin. J. Biol. Chem. 2000, 275, 1247–1260. [Google Scholar] [CrossRef] [Green Version]
- Blanks, J.C.; Johnson, L.V. Selective Lectin Binding of the Developing Mouse Retina. J. Comp. Neurol. 1983, 221, 31–41. [Google Scholar] [CrossRef]
- MacNeil, M.A.; Masland, R.H. Extreme Diversity among Amacrine Cells: Implications for Function. Neuron 1998, 20, 971–982. [Google Scholar] [CrossRef] [Green Version]
- Yan, W.; Laboulaye, M.A.; Tran, N.M.; Whitney, I.E.; Benhar, I.; Sanes, J.R. Mouse Retinal Cell Atlas: Molecular Identification of over Sixty Amacrine Cell Types. J. Neurosci. 2020, 40, 5177–5195. [Google Scholar] [CrossRef]
- Vaney, D.I. The Mosaic of Amacrine Cells in the Mammalian Retina. Prog. Ret. Res. 1990, 9, 49–100. [Google Scholar] [CrossRef]
- Wässle, H.; Boycott, B.B. Functional Architecture of the Mammalian Retina. Physiol. Rev. 1991, 71, 447–480. [Google Scholar] [CrossRef]
- Eulenburg, V.; Knop, G.; Sedmak, T.; Schuster, S.; Hauf, K.; Schneider, J.; Feigenspan, A.; Joachimsthaler, A.; Brandstätter, J.H. GlyT1 Determines the Glycinergic Phenotype of Amacrine Cells in the Mouse Retina. Brain Struct. Funct. 2018, 223, 3251–3266. [Google Scholar] [CrossRef] [PubMed]
- Vaney, D.I.; Nelson, J.C.; Pow, D.V. Neurotransmitter Coupling through Gap Junctions in the Retina. J. Neurosci. 1998, 18, 10594–10602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brecha, N.; Johnson, D.; Peichl, L.; Wässle, H. Cholinergic Amacrine Cells of the Rabbit Retina Contain Glutamate Decarboxylase and Gamma-Aminobutyrate Immunoreactivity. Proc. Natl. Acad. Sci. USA 1988, 85, 6187–6191. [Google Scholar] [CrossRef] [Green Version]
- Kosaka, T.; Tauchi, M.; Dahl, J.L. Cholinergic Neurons Containing GABA-like and/or Glutamic Acid Decarboxylase-like Immunoreactivities in Various Brain Regions of the Rat. Exp. Brain Res. 1988, 70, 605–617. [Google Scholar] [CrossRef]
- O’Malley, D.M.; Masland, R.H. Co-Release of Acetylcholine and y-Aminobutyric Acid by a Retinal Neuron. Proc. Natl. Acad. Sci. USA 1989, 86, 3414–3418. [Google Scholar] [CrossRef] [Green Version]
- Vaney, D.I.; Young, H.M. GABA-like Immunoreactivity in Cholinergic Amacrine Cells of the Rabbit Retina. Brain Res. 1988, 438, 369–373. [Google Scholar] [CrossRef]
- Jeon, C.-J.; Strettoi, E.; Masland, R.H. The Major Cell Populations of the Mouse Retina. J. Neurosci. 1998, 18, 8936–8946. [Google Scholar] [CrossRef] [Green Version]
- Haverkamp, S.; Wässle, H. Characterization of an Amacrine Cell Type of the Mammalian Retina Immunoreactive for Vesicular Glutamate Transporter 3. J. Comp. Neurol. 2004, 468, 251–263. [Google Scholar] [CrossRef]
- Johnson, J.; Sherry, D.M.; Liu, X.; Fremeau, R.T., Jr.; Seal, R.P.; Edwards, R.H.; Copenhagen, D.R. Vesicular Glutamate Transporter 3 Expression Identifies Glutamatergic Amacrine Cells in the Rodent Retina. J. Comp. Neurol. 2004, 477, 386–398. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Chen, L.; Chen, M.; Ye, M.; Seal, R.P.; Zhou, Z.J. An Unconventional Glutamatergic Circuit in the Retina Formed by VGluT3 Amacrine Cells. Neuron 2014, 84, 708–715. [Google Scholar] [CrossRef] [Green Version]
- Tien, N.-W.; Kim, T.; Kerschensteiner, D. Target-Specific Glycinergic Transmission from VGluT3-Expressing Amacrine Cells Shapes Suppressive Contrast Responses in the Retina. Cell Rep. 2016, 15, 1369–1375. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, A.R.; de Sevilla Müller, L.P.; Brecha, N.C. The RNA Binding Protein RBPMS Is a Selective Marker of Ganglion Cells in the Mammalian Retina. J. Comp. Neurol. 2014, 522, 1411–1443. [Google Scholar] [CrossRef] [Green Version]
- Krieger, B.; Qiao, M.; Rousso, D.L.; Sanes, J.R.; Meister, M. Four Alpha Ganglion Cell Types in Mouse Retina: Function, Structure, and Molecular Signatures. PLoS ONE 2017, 12, e0180091. [Google Scholar] [CrossRef] [Green Version]
- Peichl, L.; Ott, H.; Boycott, B.B. Alpha Ganglion Cells in Mammalian Retinae. Proc. R. Soc. Lond. B 1987, 231, 169–197. [Google Scholar] [CrossRef]
- Jiang, D.; Burger, C.A.; Casasent, A.K.; Albrecht, N.E.; Li, F.; Samuel, M.A. Spatiotemporal Gene Expression Patterns Reveal Molecular Relatedness between Retinal Laminae. J. Comp. Neurol. 2020, 528, 729–755. [Google Scholar] [CrossRef] [PubMed]
- Macosko, E.Z.; Basu, A.; Satija, R.; Nemesh, J.; Shekhar, K.; Goldman, M.; Tirosh, I.; Bialas, A.R.; Kamitaki, N.; Martersteck, E.M.; et al. Highly Parallel Genome-Wide Expression Profiling of Individual Cells Using Nanoliter Droplets. Cell 2015, 161, 1202–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, L.J. Development of Synaptic Arrays in the Inner Plexiform Layer of Neonatal Mouse Retina. J. Comp. Neurol. 1979, 187, 359–372. [Google Scholar] [CrossRef] [PubMed]
- Sassoè-Pognetto, M.; Wässle, H. Synaptogenesis in the Rat Retina: Subcellular Localization of Glycine Receptors, GABA(A) Receptors, and the Anchoring Protein Gephyrin. J. Comp. Neurol. 1997, 381, 158–174. [Google Scholar] [CrossRef]
- Blanks, J.C.; Adinolfi, A.M.; Lolley, R.N. Synaptogenesis in the Photoreceptor Terminal of the Mouse Retina. J. Comp. Neurol. 1974, 156, 81–93. [Google Scholar] [CrossRef]
- Regus-Leidig, H.; tom Dieck, S.; Specht, D.; Meyer, L.; Brandstätter, J.H. Early Steps in the Assembly of Photoreceptor Ribbon Synapses in the Mouse Retina: The Involvement of Precursor Spheres. J. Comp. Neurol. 2009, 512, 814–824. [Google Scholar] [CrossRef]
- Brose, N. For Better or for Worse: Complexins Regulate SNARE Function and Vesicle Fusion. Traffic 2008, 9, 1403–1413. [Google Scholar] [CrossRef]
- Trimbuch, T.; Rosenmund, C. Should I Stop or Should I Go? The Role of Complexin in Neurotransmitter Release. Nat. Rev. Neurosci. 2016, 17, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Rosolen, S.G.; Rigaudière, F.; LeGargasson, J.-F.; Chalier, C.; Rufiange, M.; Racine, J.; Joly, S.; Lachapelle, P. Comparing the Photopic ERG I-Wave in Different Species. Vet. Ophthalmol. 2004, 7, 189–192. [Google Scholar] [CrossRef]
- Glynn, D.; Drew, C.J.; Reim, K.; Brose, N.; Morton, A.J. Profound Ataxia in Complexin I Knockout Mice Masks a Complex Phenotype That Includes Exploratory and Habituation Deficits. Hum. Mol. Genet. 2005, 14, 2369–2385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Zhang, Y.; Chen, M.; Zhou, Z.J. Segregated Glycine-Glutamate Co-Transmission from VGluT3 Amacrine Cells to Contrast-Suppressed and Contrast-Enhanced Retinal Circuits. Neuron 2016, 90, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Landgraf, I.; Mühlhans, J.; Dedek, K.; Reim, K.; Brandstätter, J.H.; Ammermüller, J. The Absence of Complexin 3 and Complexin 4 Differentially Impacts the ON and OFF Pathways in Mouse Retina. Eur. J. Neurosci. 2012, 36, 2470–2481. [Google Scholar] [CrossRef]
- Nouvian, R.; Neef, J.; Bulankina, A.V.; Reisinger, E.; Pangršič, T.; Frank, T.; Sikorra, S.; Brose, N.; Binz, T.; Moser, T. Exocytosis at the Hair Cell Ribbon Synapse Apparently Operates without Neuronal SNARE Proteins. Nat. Neurosci. 2011, 14, 411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strenzke, N.; Chanda, S.; Kopp-Scheinpflug, C.; Khimich, D.; Reim, K.; Bulankina, A.V.; Neef, A.; Wolf, F.; Brose, N.; Xu-Friedman, M.A.; et al. Complexin-I Is Required for High-Fidelity Transmission at the Endbulb of Held Auditory Synapse. J. Neurosci. 2009, 29, 7991–8004. [Google Scholar] [CrossRef] [Green Version]
- Beurg, M.; Michalski, N.; Safieddine, S.; Bouleau, Y.; Schneggenburger, R.; Chapman, E.R.; Petit, C.; Dulon, D. Control of Exocytosis by Synaptotagmins and Otoferlin in Auditory Hair Cells. J. Neurosci. 2010, 30, 13281–13290. [Google Scholar] [CrossRef] [Green Version]
- Reisinger, E.; Bresee, C.; Neef, J.; Nair, R.; Reuter, K.; Bulankina, A.; Nouvian, R.; Koch, M.; Bückers, J.; Kastrup, L.; et al. Probing the Functional Equivalence of Otoferlin and Synaptotagmin 1 in Exocytosis. J. Neurosci. 2011, 31, 4886–4895. [Google Scholar] [CrossRef]
- Ishizuka, T.; Saisu, H.; Odani, S.; Kumanishi, T.; Abe, T. Distinct Regional Distribution in the Brain of Messenger RNAs for the Two Isoforms of Synaphin Associated with the Docking/Fusion Complex. Neuroscience 1999, 88, 295–306. [Google Scholar] [CrossRef]
- Yamada, M.; Saisu, H.; Ishizuka, T.; Takahashi, H.; Abe, T. Immunohistochemical Distribution of the Two Isoforms of Synaphin/Complexin Involved in Neurotransmitter Release: Localization at the Distinct Central Nervous System Regions and Synaptic Types. Neuroscience 1999, 93, 7–18. [Google Scholar] [CrossRef]
- Freeman, W.; Morton, A.J. Differential Messenger RNA Expression of Complexins in Mouse Brain. Brain Res. Bull. 2004, 63, 33–44. [Google Scholar] [CrossRef]
- Gierke, K.; von Wittgenstein, J.; Hemmerlein, M.; Atorf, J.; Joachimsthaler, A.; Kremers, J.; Cooper, B.H.; Varoqueaux, F.; Regus-Leidig, H.; Brandstätter, J.H. Heterogeneous Presynaptic Distribution of Munc13 Isoforms at Retinal Synapses and Identification of an Unconventional Bipolar Cell Type with Dual Expression of Munc13 Isoforms: A Study Using Munc13-EXFP Knock-in Mice. Int. J. Mol. Sci. 2020, 21, 7848. [Google Scholar] [CrossRef]
- Thoreson, W.B.; Mangel, S.C. Lateral Interactions in the Outer Retina. Prog. Retin. Eye Res. 2012, 31, 407–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirano, A.A.; Vuong, H.E.; Kornmann, H.L.; Schietroma, C.; Stella, S.L.; Barnes, S.; Brecha, N.C. Vesicular Release of GABA by Mammalian Horizontal Cells Mediates Inhibitory Output to Photoreceptors. Front. Cell. Neurosci. 2020, 14, 600777. [Google Scholar] [CrossRef]
- Hirano, A.A.; Brandstätter, J.H.; Brecha, N.C. Cellular Distribution and Subcellular Localization of Molecular Components of Vesicular Transmitter Release in Horizontal Cells of Rabbit Retina. J. Comp. Neurol. 2005, 488, 70–81. [Google Scholar] [CrossRef] [Green Version]
- Boycott, B.; Wässle, H. Parallel Processing in the Mammalian Retina: The Proctor Lecture. Investig. Ophthalmol. Vis. Sci. 1999, 40, 1313–1327. [Google Scholar]
- Baden, T.; Berens, P.; Franke, K.; Román Rosón, M.; Bethge, M.; Euler, T. The Functional Diversity of Retinal Ganglion Cells in the Mouse. Nature 2016, 529, 345–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, J.A.; Mu, S.; Kim, J.S.; Turner, N.L.; Tartavull, I.; Kemnitz, N.; Jordan, C.S.; Norton, A.D.; Silversmith, W.M.; Prentki, R.; et al. Digital Museum of Retinal Ganglion Cells with Dense Anatomy and Physiology. Cell 2018, 173, 1293–1306.e19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rheaume, B.A.; Jereen, A.; Bolisetty, M.; Sajid, M.S.; Yang, Y.; Renna, K.; Sun, L.; Robson, P.; Trakhtenberg, E.F. Single Cell Transcriptome Profiling of Retinal Ganglion Cells Identifies Cellular Subtypes. Nat. Commun. 2018, 9, 2759. [Google Scholar] [CrossRef]
- Tran, N.M.; Shekhar, K.; Whitney, I.E.; Jacobi, A.; Benhar, I.; Hong, G.; Yan, W.; Adiconis, X.; Arnold, M.E.; Lee, J.M.; et al. Single-Cell Profiles of Retinal Ganglion Cells Differing in Resilience to Injury Reveal Neuroprotective Genes. Neuron 2019, 104, 1039–1055.e12. [Google Scholar] [CrossRef] [PubMed]
- Hidaka, S.; Akahori, Y.; Kurosawa, Y. Dendrodendritic Electrical Synapses between Mammalian Retinal Ganglion Cells. J. Neurosci. 2004, 24, 10553–10567. [Google Scholar] [CrossRef]
- Puller, C.; Duda, S.; Lotfi, E.; Arzhangnia, Y.; Block, C.T.; Ahlers, M.T.; Greschner, M. Electrical Coupling of Heterotypic Ganglion Cells in the Mammalian Retina. J. Neurosci. 2020, 40, 1302–1310. [Google Scholar] [CrossRef]
- Chang, S.; Reim, K.; Pedersen, M.; Neher, E.; Brose, N.; Taschenberger, H. Complexin Stabilizes Newly Primed Synaptic Vesicles and Prevents Their Premature Fusion at the Mouse Calyx of Held Synapse. J. Neurosci. 2015, 35, 8272–8290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, M.; Polepalli, J.S.; Goswami, D.; Yang, X.; Kaeser-Woo, Y.J.; Südhof, T.C.; Malenka, R.C. Postsynaptic Complexin Controls AMPA Receptor Exocytosis during LTP. Neuron 2012, 73, 260–267. [Google Scholar] [CrossRef] [Green Version]
- Harazny, J.; Scholz, M.; Buder, T.; Lausen, B.; Kremers, J. Electrophysiological Deficits in the Retina of the DBA/2J Mouse. Doc. Ophthalmol. 2009, 119, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, P.; Gommers, R.; Oliphant, T.E.; Haberland, M.; Reddy, T.; Cournapeau, D.; Burovski, E.; Peterson, P.; Weckesser, W.; Bright, J.; et al. SciPy 1.0: Fundamental Algorithms for Scientific Computing in Python. Nat. Methods 2020, 17, 261–272. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forward | Reverse | |
|---|---|---|
| Cplx 1 | GATCGTCCACCTTGGAAG | GATGGCTTGGTTCCTCAG |
| Cplx 2 | AGTGGCTTAGACGGTTG | TGCAGGCTTTGGTTAATG |
| Cplx 3 | GAAGAGTACGAGGAGTATC | CTTCCTCTGTGTGAACTG |
| Cplx 4 | GGCTAAAGGGATGACTAG | CTCTCTCCATCTTCTCTTC |
| Actin | TTCCTCCCTGGAGAAGAG | CACTGTGTTGGCATAGAG |
| GAPDH | CAACTTTGTCAAGCTCATT | TCTGGGATGGAAATTGTG |
| PBGD | TACCCTGGCATACAGTTT | CGTTTTCTAGCTCCTTGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lux, U.T.; Ehrenberg, J.; Joachimsthaler, A.; Atorf, J.; Pircher, B.; Reim, K.; Kremers, J.; Gießl, A.; Brandstätter, J.H. Cell Types and Synapses Expressing the SNARE Complex Regulating Proteins Complexin 1 and Complexin 2 in Mammalian Retina. Int. J. Mol. Sci. 2021, 22, 8131. https://doi.org/10.3390/ijms22158131
Lux UT, Ehrenberg J, Joachimsthaler A, Atorf J, Pircher B, Reim K, Kremers J, Gießl A, Brandstätter JH. Cell Types and Synapses Expressing the SNARE Complex Regulating Proteins Complexin 1 and Complexin 2 in Mammalian Retina. International Journal of Molecular Sciences. 2021; 22(15):8131. https://doi.org/10.3390/ijms22158131
Chicago/Turabian StyleLux, Uwe Thorsten, Johanna Ehrenberg, Anneka Joachimsthaler, Jenny Atorf, Bianca Pircher, Kerstin Reim, Jan Kremers, Andreas Gießl, and Johann Helmut Brandstätter. 2021. "Cell Types and Synapses Expressing the SNARE Complex Regulating Proteins Complexin 1 and Complexin 2 in Mammalian Retina" International Journal of Molecular Sciences 22, no. 15: 8131. https://doi.org/10.3390/ijms22158131
APA StyleLux, U. T., Ehrenberg, J., Joachimsthaler, A., Atorf, J., Pircher, B., Reim, K., Kremers, J., Gießl, A., & Brandstätter, J. H. (2021). Cell Types and Synapses Expressing the SNARE Complex Regulating Proteins Complexin 1 and Complexin 2 in Mammalian Retina. International Journal of Molecular Sciences, 22(15), 8131. https://doi.org/10.3390/ijms22158131