
The Rice Cation/H+ Exchanger Family Involved in Cd Tolerance and Transport



Abstract
:1. Introduction
2. Results
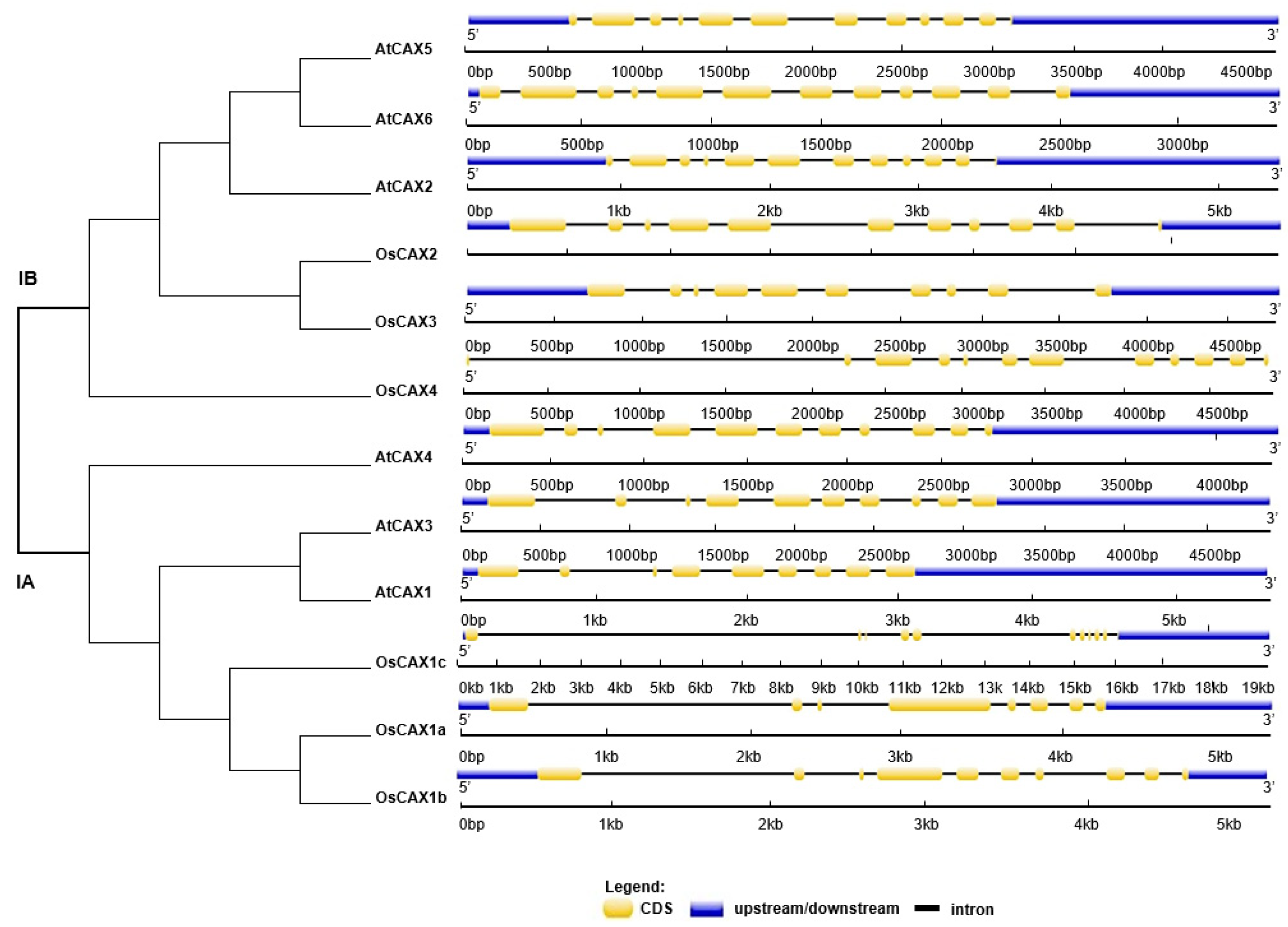
2.1. Bioinformatics Analyses of CAX Family Genes in Oryza sativa and Arabidopsis thaliana
2.2. Response of Rice CAX Family Genes to Cd Stress
2.3. Functional Analysis of Rice CAX Genes in Yeast
2.4. Expression Pattern and Subcellular Localization of the Rice CAX Genes
3. Discussion
3.1. The CAX Family Genes in Rice Have High Identity to Their Homologues in Arabidopsis Thaliana
3.2. N-Terminal Auto-Inhibition Was Not Found for the Rice CAX Genes Tested Using Cd Treatment
3.3. Rice CAX Genes, Particularly OsCAX1a, OsCAX1c and OsCAX4, Might Be Important in Controlling Cd Uptake and Translocation in Rice
4. Materials and Method
4.1. Identification and Bioinformatics Analyses of the Cation/H+ Antiporters from Oryza sativa and Arabidopsis thaliana Species
4.2. Plant Materials and Growth Conditions
4.3. RNA Extraction and Real-Time PCR
4.4. Functional Analysis of Rice CAX Genes in Yeast
4.5. Tissue Expression Assay
4.6. Subcellular-Localization Assay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ABC | ATP-binding cassette |
| CAX | Cation/H+ exchanger |
| CaCA | Ca2+/cation antiporter |
| CCX | Cation/Ca2+ exchanger |
| CDT | Cation diffusion facilitator |
| HMA | Heavy metal ATPase |
| ICP-MS | Inductively coupled plasma mass spectrometry |
| MAPK | Mitogen-activated protein kinase |
| MHX | Mg2+/H+ exchanger |
| MTP | Metal tolerance protein |
| NRR | N-terminal regulatory region |
| TF | Transcription factors |
| TMDs | Transmembrane domains |
References
- Parmar, P.; Kumari, N.; Sharma, V. Structural and functional alterations in photosynthetic apparatus of plants under cadmium stress. Bot. Stud. 2013, 54, 55. [Google Scholar] [CrossRef] [Green Version]
- Pereira, A.S.; Dorneles, A.O.S.; Bernardy, K.; Sasso, V.M.; Bernardy, D.; Possebom, G.; Rossato, L.V.; Dressler, V.L.; Tabaldi, L.A. Selenium and silicon reduce cadmium uptake and mitigate cadmium toxicity in Pfaffia glomerata (Spreng.) Pedersen plants by activation antioxidant enzyme system. Environ. Sci. Pollut. Res. Int. 2018, 25, 18548–18558. [Google Scholar] [CrossRef]
- Shi, Z.; Yang, S.; Han, D.; Zhou, Z.; Li, X.; Liu, Y.; Zhang, B. Silicon alleviates cadmium toxicity in wheat seedlings (Triticum aestivum L.) by reducing cadmium ion uptake and enhancing antioxidative capacity. Environ. Sci. Pollut. Res. Int. 2018, 25, 7638–7646. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, M.; Liu, Z.; Shi, Y.; Han, T.; Ye, Y.; Gong, N.; Sun, J.; Zhu, C. Different responses of low grain-Cd-accumulating and high grain-Cd-accumulating rice cultivars to Cd stress. Plant Physiol. Biochem. 2015, 96, 261–269. [Google Scholar] [CrossRef]
- Qayyum, M.F.; Rehman, M.Z.U.; Ali, S.; Rizwan, M.; Naeem, A.; Maqsood, M.A.; Khalid, H.; Rinklebe, J.; Ok, Y.S. Residual effects of monoammonium phosphate, gypsum and elemental sulfur on cadmium phytoavailability and translocation from soil to wheat in an effluent irrigated field. Chemosphere 2017, 174, 515–523. [Google Scholar] [CrossRef]
- Hussain, A.; Ali, S.; Rizwan, M.; Zia, U.R.M.; Javed, M.R.; Imran, M.; Chatha, S.A.S.; Nazir, R. Zinc oxide nanoparticles alter the wheat physiological response and reduce the cadmium uptake by plants. Environ. Pollut. 2018, 242, 1518–1526. [Google Scholar] [CrossRef] [PubMed]
- Rehman, M.Z.U.; Rizwan, M.; Khalid, H.; Ali, S.; Naeem, A.; Yousaf, B.; Liu, G.; Sabir, M.; Farooq, M. Farmyard manure alone and combined with immobilizing amendments reduced cadmium accumulation in wheat and rice grains grown in field irrigated with raw effluents. Chemosphere 2018, 199, 468–476. [Google Scholar] [CrossRef]
- Hu, Y.; Cheng, H.; Tao, S. The Challenges and Solutions for Cadmium-contaminated Rice in China: A Critical Review. Environ. Int. 2016, 92–93, 515–532. [Google Scholar] [CrossRef]
- Yang, Q.; Li, Z.; Lu, X.; Duan, Q.; Huang, L.; Bi, J. A review of soil heavy metal pollution from industrial and agricultural regions in China: Pollution and risk assessment. Sci. Total Environ. 2018, 642, 690–700. [Google Scholar] [CrossRef]
- Karahan, F.; Ozyigit, I.I.; Saracoglu, I.A.; Yalcin, I.E.; Ozyigit, A.H.; Ilcim, A. Heavy Metal Levels and Mineral Nutrient Status in Different Parts of Various Medicinal Plants Collected from Eastern Mediterranean Region of Turkey. Biol. Trace Elem. Res. 2020, 197, 316–329. [Google Scholar] [CrossRef]
- Qin, G.; Niu, Z.; Yu, J.; Li, Z.; Ma, J.; Xiang, P. Soil heavy metal pollution and food safety in China: Effects, sources and removing technology. Chemosphere 2021, 267, 129205. [Google Scholar] [CrossRef]
- Kussaga, J.B.; Jacxsens, L.; Tiisekwa, B.P.; Luning, P.A. Food safety management systems performance in African food processing companies: A review of deficiencies and possible improvement strategies. J. Sci. Food Agric. 2014, 94, 2154–2169. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Wang, J.; Fang, W.; Yuan, J.; Yang, Z. Cadmium accumulation in different rice cultivars and screening for pollution-safe cultivars of rice. Sci. Total Environ. 2006, 370, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Deng, S.; Tan, D.; Long, J.; Lei, M. Heavy metal distribution, translocation, and human health risk assessment in the soil-rice system around Dongting Lake area, China. Environ. Sci. Pollut. Res. Int. 2019, 26, 17655–17665. [Google Scholar] [CrossRef]
- Chen, Q.; Wu, F.B. Breeding for low cadmium accumulation cereals. J. Zhejiang Univ. Sci. B 2020, 21, 442–459. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.G.; Liang, J.S.; Li, K.Q.; Zhang, Z.J.; Yu, B.Y.; Lu, X.L.; Yang, J.C.; Zhu, Q.S. Correlations between cadmium and mineral nutrients in absorption and accumulation in various genotypes of rice under cadmium stress. Chemosphere 2003, 52, 1467–1473. [Google Scholar] [CrossRef]
- Tezuka, K.; Miyadate, H.; Katou, K.; Kodama, I.; Matsumoto, S.; Kawamoto, T.; Masaki, S.; Satoh, H.; Yamaguchi, M.; Sakurai, K. A single recessive gene controls cadmium translocation in the cadmium hyperaccumulating rice cultivar Cho-Ko-Koku. Theor. Appl. Genet. 2010, 120, 1175–1182. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zou, W.; Meng, L.; Fan, X.; Xu, G.; Ye, G. Advances in the Uptake and Transport Mechanisms and QTLs Mapping of Cadmium in Rice. Int. J. Mol. Sci. 2019, 20, 3417. [Google Scholar] [CrossRef] [Green Version]
- Uraguchi, S.; Fujiwara, T. Rice breaks ground for cadmium-free cereals. Curr. Opin. Plant Biol. 2013, 16, 328–334. [Google Scholar] [CrossRef]
- Agrawal, G.K.; Rakwal, R.; Iwahashi, H. Isolation of novel rice (Oryza sativa L.) multiple stress responsive MAP kinase gene, OsMSRMK2, whose mRNA accumulates rapidly in response to environmental cues. Biochem. Biophys. Res. Commun. 2002, 294, 1009–1016. [Google Scholar] [CrossRef]
- Agrawal, G.K.; Agrawal, S.K.; Shibato, J.; Iwahashi, H.; Rakwal, R. Novel rice MAP kinases OsMSRMK3 and OsWJUMK1 involved in encountering diverse environmental stresses and developmental regulation. Biochem. Biophys. Res. Commun. 2003, 300, 775–783. [Google Scholar] [CrossRef]
- Yeh, C.M.; Hsiao, L.J.; Huang, H.J. Cadmium activates a mitogen-activated protein kinase gene and MBP kinases in rice. Plant Cell Physiol. 2004, 45, 1306–1312. [Google Scholar] [CrossRef]
- Kuramata, M.; Masuya, S.; Takahashi, Y.; Kitagawa, E.; Inoue, C.; Ishikawa, S.; Youssefian, S.; Kusano, T. Novel cysteine-rich peptides from Digitaria ciliaris and Oryza sativa enhance tolerance to cadmium by limiting its cellular accumulation. Plant Cell Physiol. 2009, 50, 106–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oda, K.; Otani, M.; Uraguchi, S.; Akihiro, T.; Fujiwara, T. Rice ABCG43 is Cd inducible and confers Cd tolerance on yeast. Biosci. Biotechnol. Biochem. 2011, 75, 1211–1213. [Google Scholar] [CrossRef] [PubMed]
- Moons, A. Ospdr9, which encodes a PDR-type ABC transporter, is induced by heavy metals, hypoxic stress and redox perturbations in rice roots. FEBS Lett. 2003, 553, 370–376. [Google Scholar] [CrossRef] [Green Version]
- Fu, S.; Lu, Y.; Zhang, X.; Yang, G.; Chao, D.; Wang, Z.; Shi, M.; Chen, J.; Chao, D.Y.; Li, R. The ABC transporter ABCG36 is required for cadmium tolerance in rice. J. Exp. Bot. 2019, 70, 5909–5918. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, S.A.; Shin, R.; Eide, D.J.; Schachtman, D.P. Differential metal selectivity and gene expression of two zinc transporters from rice. Plant Physiol. 2003, 133, 126–134. [Google Scholar] [CrossRef] [Green Version]
- Chou, T.S.; Chao, Y.Y.; Huang, W.; Hong, C.Y.; Kao, C.H. Effect of magnesium deficiency on antioxidant status and cadmium toxicity in rice seedlings. J. Plant Physiol. 2011, 168, 1021–1030. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; An, G. Over-expression of OsIRT1 leads to increased iron and zinc accumulations in rice. Plant Cell Environ. 2009, 32, 408–416. [Google Scholar] [CrossRef]
- Tan, L.; Zhu, Y.; Fan, T.; Peng, C.; Wang, J.; Sun, L.; Chen, C. OsZIP7 functions in xylem loading in roots and inter-vascular transfer in nodes to deliver Zn/Cd to grain in rice. Biochem. Biophys. Res. Commun. 2019, 512, 112–118. [Google Scholar] [CrossRef]
- Sasaki, A.; Yamaji, N.; Ma, J.F. Overexpression of OsHMA3 enhances Cd tolerance and expression of Zn transporter genes in rice. J. Exp. Bot. 2014, 65, 6013–6021. [Google Scholar] [CrossRef] [Green Version]
- Ueno, D.; Milner, M.J.; Yamaji, N.; Yokosho, K.; Koyama, E.; Clemencia, Z.M.; Kaskie, M.; Ebbs, S.; Kochian, L.V.; Ma, J.F. Elevated expression of TcHMA3 plays a key role in the extreme Cd tolerance in a Cd-hyperaccumulating ecotype of Thlaspi caerulescens. Plant J. 2011, 66, 852–862. [Google Scholar] [CrossRef]
- Lee, S.; Kim, Y.Y.; Lee, Y.; An, G. Rice P1B-type heavy-metal ATPase, OsHMA9, is a metal efflux protein. Plant Physiol. 2007, 145, 831–842. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, R.; Ishimaru, Y.; Shimo, H.; Ogo, Y.; Senoura, T.; Nishizawa, N.K.; Nakanishi, H. The OsHMA2 transporter is involved in root-to-shoot translocation of Zn and Cd in rice. Plant Cell Environ. 2012, 35, 1948–1957. [Google Scholar] [CrossRef]
- Yamaji, N.; Xia, J.; Mitani-Ueno, N.; Yokosho, K.; Ma, J.F. Preferential delivery of zinc to developing tissues in rice is mediated by P-type heavy metal ATPase OsHMA2. Plant Physiol. 2013, 162, 927–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueno, D.; Yamaji, N.; Kono, I.; Huang, C.F.; Ando, T.; Yano, M.; Ma, J.F. Gene limiting cadmium accumulation in rice. Proc. Natl. Acad. Sci. USA 2010, 107, 16500–16505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, L.; Yang, S.; Liu, B.; Zhang, M.; Wu, K. Molecular characterization of a rice metal tolerance protein, OsMTP1. Plant Cell Rep. 2012, 31, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Liu, B. Identification of a rice metal tolerance protein OsMTP11 as a manganese transporter. PLoS ONE 2017, 12, e0174987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Yang, W.; Zhang, S.; Yang, T.; Liu, Q.; Dong, J.; Fu, H.; Mao, X.; Liu, B. Genome-wide association study and candidate gene analysis of rice cadmium accumulation in grain in a diverse rice collection. Rice 2018, 11, 61. [Google Scholar] [CrossRef] [Green Version]
- Ishimaru, Y.; Takahashi, R.; Bashir, K.; Shimo, H.; Senoura, T.; Sugimoto, K.; Ono, K.; Yano, M.; Ishikawa, S.; Arao, T. Characterizing the role of rice NRAMP5 in Manganese, Iron and Cadmium Transport. Sci. Rep. 2012, 2, 286. [Google Scholar] [CrossRef]
- Sasaki, A.; Yamaji, N.; Yokosho, K.; Ma, J.F. Nramp5 is a major transporter responsible for manganese and cadmium uptake in rice. Plant Cell 2012, 24, 2155–2167. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Zhang, Y.; Zhang, L.; Hu, J.; Zhang, X.; Lu, K.; Dong, H.; Wang, D.; Zhao, F.J.; Huang, C.F.; et al. OsNRAMP5 contributes to manganese translocation and distribution in rice shoots. J. Exp. Bot. 2014, 65, 4849–4861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, R.; Ishimaru, Y.; Shimo, H.; Bashir, K.; Senoura, T.; Sugimoto, K.; Ono, K.; Suzui, N.; Kawachi, N.; Ishii, S. From laboratory to field: OsNRAMP5-knockdown rice is a promising candidate for Cd phytoremediation in paddy fields. PLoS ONE 2014, 9, e98816. [Google Scholar]
- Tang, L.; Mao, B.; Li, Y.; Lv, Q.; Zhang, L.; Chen, C.; He, H.; Wang, W.; Zeng, X.; Shao, Y. Knockout of OsNramp5 using the CRISPR/Cas9 system produces low Cd-accumulating indica rice without compromising yield. Sci. Rep. 2017, 7, 14438. [Google Scholar] [CrossRef] [Green Version]
- Taneja, M.; Tyagi, S.; Sharma, S.; Upadhyay, S.K. Ca2+/Cation Antiporters (CaCA): Identification, Characterization and Expression Profiling in Bread Wheat (Triticum aestivum L.). Front. Plant Sci. 2016, 7, 1775. [Google Scholar] [CrossRef] [Green Version]
- Hirschi, K. Expression of Arabidopsis CAX1 in tobacco: Altered calcium homeostasis and increased stress sensitivity. Plant Cell 1999, 11, 2113–2122. [Google Scholar] [CrossRef] [Green Version]
- Choe, M.; Choe, W.; Cha, S.; Lee, I. Changes of cationic transport in AtCAX5 transformant yeast by electromagnetic field environments. J. Biol. Phys. 2018, 44, 433–448. [Google Scholar] [CrossRef] [PubMed]
- Baliardini, C.; Meyer, C.L.; Salis, P.; Saumitou-Laprade, P.; Verbruggen, N. CATION EXCHANGER1 Cosegregates with Cadmium Tolerance in the Metal Hyperaccumulator Arabidopsis halleri and Plays a Role in Limiting Oxidative Stress in Arabidopsis spp. Plant Physiol. 2015, 169, 549–559. [Google Scholar] [CrossRef] [Green Version]
- Ahmadi, H.; Corso, M.; Weber, M.; Verbruggen, N.; Clemens, S. CAX1 suppresses Cd-induced generation of reactive oxygen species in Arabidopsis halleri. Plant Cell Environ. 2018, 41, 2435–2448. [Google Scholar] [CrossRef] [PubMed]
- Korenkov, V.; Hirschi, K.; Crutchfield, J.D.; Wagner, G.J. Enhancing tonoplast Cd/H antiport activity increases Cd, Zn, and Mn tolerance, and impacts root/shoot Cd partitioning in Nicotiana tabacum L. Planta 2007, 226, 1379–1387. [Google Scholar] [CrossRef]
- Koren’kov, V.; Park, S.; Cheng, N.H.; Sreevidya, C.; Lachmansingh, J.; Morris, J.; Hirschi, K.; Wagner, G.J. Enhanced Cd2+-selective root-tonoplast-transport in tobaccos expressing Arabidopsis cation exchangers. Planta 2007, 225, 403–411. [Google Scholar] [CrossRef]
- Korenkov, V.; King, B.; Hirschi, K.; Wagner, G.J. Root-selective expression of AtCAX4 and AtCAX2 results in reduced lamina cadmium in field-grown Nicotiana tabacum L. Plant Biotechnol. J. 2009, 7, 219–226. [Google Scholar] [CrossRef]
- Qiao, K.; Wang, F.; Liang, S.; Hu, Z.; Chai, T. Heterologous expression of TuCAX1a and TuCAX1b enhances Ca2+ and Zn2+ translocation in Arabidopsis. Plant Cell Rep. 2019, 38, 597–607. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hao, J.; Bao, M.; Hasi, A.; Niu, Y. Cloning and characterization of a Ca2+/H+ exchanger from the halophyte Salicornia europaea L. Plant Physiol. Biochem. 2015, 96, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Catala, R.; Santos, E.; Alonso, J.M.; Ecker, J.R.; Martinez-Zapater, J.M.; Salinas, J. Mutations in the Ca2+/H+ transporter CAX1 increase CBF/DREB1 expression and the cold-acclimation response in Arabidopsis. Plant Cell 2003, 15, 2940–2951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirschi, K.D.; Zhen, R.G.; Cunningham, K.W.; Rea, P.A.; Fink, G.R. CAX1, an H+/Ca2+ antiporter from Arabidopsis. Proc. Natl. Acad. Sci. USA 1996, 93, 8782–8786. [Google Scholar] [CrossRef] [Green Version]
- Shigaki, T.; Cheng, N.H.; Pittman, J.K.; Hirschi, K. Structural determinants of Ca2+ transport in the Arabidopsis H+/Ca2+ antiporter CAX1. J. Biol. Chem. 2001, 276, 43152–43159. [Google Scholar] [CrossRef] [Green Version]
- Edmond, C.; Shigaki, T.; Ewert, S.; Nelson, M.D.; Connorton, J.M.; Chalova, V.; Noordally, Z.; Pittman, J.K. Comparative analysis of CAX2-like cation transporters indicates functional and regulatory diversity. Biochem. J. 2009, 418, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.K.; Han, J.S.; Lee, H.S.; Oh, J.Y.; Shigaki, T.; Park, S.H.; Hirschi, K. Expression of an Arabidopsis CAX2 variant in potato tubers increases calcium levels with no accumulation of manganese. Plant Cell Rep. 2006, 25, 1226–1232. [Google Scholar] [CrossRef]
- Cheng, N.H.; Pittman, J.K.; Barkla, B.J.; Shigaki, T.; Hirschi, K.D. The Arabidopsis cax1 mutant exhibits impaired ion homeostasis, development, and hormonal responses and reveals interplay among vacuolar transporters. Plant Cell 2003, 15, 347–364. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Shigaki, T.; Mei, H.; Guo, Y.Q.; Cheng, N.H.; Hirschi, K.D. Interaction between Arabidopsis Ca2+/H+ exchangers CAX1 and CAX3. J. Biol. Chem. 2009, 2844, 605–4615. [Google Scholar]
- Bu, Y.; Fu, W.; Chen, J.; Takano, T.; Liu, S. Description of AtCAX4 in Response to Abiotic Stress in Arabidopsis. Int. J. Mol. Sci. 2021, 22, 856. [Google Scholar] [CrossRef]
- Kamiya, T.; Akahori, T.; Maeshima, M. Expression profile of the genes for rice cation/H+ exchanger family and functional analysis in yeast. Plant Cell Physiol. 2005, 46, 1735–1740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, N.; Theerawitaya, C.; Cha-um, S.; Kirdmanee, C.; Takabe, T. Expression and functional analysis of putative vacuolar Ca2+-transporters (CAXs and ACAs) in roots of salt tolerant and sensitive rice cultivars. Protoplasma 2014, 251, 1067–1075. [Google Scholar] [CrossRef] [PubMed]
- Liao, Q.; Jian, S.F.; Song, H.X.; Guan, C.Y.; Lepo, J.; Ismail, A.M.; Zhang, Z.H. Balance between nitrogen use efficiency and cadmium tolerance in Brassica napus and Arabidopsis thaliana. Plant Sci. 2019, 284, 57–66. [Google Scholar] [CrossRef]
- Cheng, N.H.; Pittman, J.K.; Shigaki, T.; Lachmansingh, J.; LeClere, S.; Lahner, B.; Salt, D.E.; Hirschi, K.D. Functional association of Arabidopsis CAX1 and CAX3 is required for normal growth and ion homeostasis. Plant Physiol. 2005, 138, 2048–2060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Zhang, X.; Takano, T.; Liu, S. Characterization of a PutCAX1 gene from Puccinellia tenuiflora that confers Ca2+ and Ba2+ tolerance in yeast. Biochem. Biophys. Res. Commun. 2009, 383, 392–396. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Zahid, K.R.; He, L.; Zhang, W.; He, X.; Zhang, X.; Yang, X.; Zhu, L. GhCAX3 gene, a novel Ca2+/H+ exchanger from cotton, confers regulation of cold response and ABA induced signal transduction. PLoS ONE 2013, 8, e66303. [Google Scholar] [CrossRef]
- Martins, V.; Carneiro, F.; Conde, C.; Sottomay, M.; Geros, H. The grapevine VvCAX3 is a cation/H(+) exchanger involved in vacuolar Ca(2+) homeostasis. Planta 2017, 246, 1083–1096. [Google Scholar] [CrossRef]
- Pittman, J.K.; Hirschi, K.D. Phylogenetic analysis and protein structure modelling identifies distinct Ca2+/Cation antiporters and conservation of gene family structure within Arabidopsis and rice species. Rice 2016, 9, 3. [Google Scholar] [CrossRef] [Green Version]
- Pittman, J.K.; Hirschi, K.D. Regulation of CAX1, an Arabidopsis Ca2+/H+ antiporter. Identification of an N-terminal autoinhibitory domain. Plant Physiol. 2001, 127, 1020–1029. [Google Scholar] [CrossRef]
- Shigaki, T.; Mei, H.; Marshall, J.; Li, X.; Manohar, M.; Hirschi, K.D. The expression of the open reading frame of Arabidopsis CAX1, but not its cDNA, confers metal tolerance in yeast. Plant Biol. 2010, 12, 935–939. [Google Scholar] [CrossRef] [PubMed]
- Hirschi, K.D.; Korenkov, V.D.; Wilganowski, N.L.; Wagner, G.J. Expression of Arabidopsis CAX2 in tobacco. Altered metal accumulation and increased manganese tolerance. Plant Physiol. 2000, 124, 125–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, A.K.; Hallett, M.B.; Daw, R.A.; Luzio, J.P.; Siddle, K. The importance of measuring intracellular free Ca2+. Biochem. Soc. Trans. 1979, 7, 865–869. [Google Scholar] [CrossRef] [PubMed]
- Hoang, X.L.T.; Nhi, D.N.H.; Thu, N.B.A.; Thao, N.P.; Tran, L.P. Transcription Factors and Their Roles in Signal Transduction in Plants under Abiotic Stresses. Curr. Genom. 2017, 18, 483–497. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.Q.; Jiang, Z.H.; Yi, Y.Y.; Yang, Y.; Ke, L.P.; Pei, Z.M.; Zhu, S. Biotic and Abiotic Stresses Activate Different Ca2+ Permeable Channels in Arabidopsis. Front. Plant Sci. 2017, 8, 83. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Ouyang, Y.; Fan, Y.; Qiu, B.; Zhang, G.; Zeng, F. The pathway of transmembrane cadmium influx via calcium-permeable channels and its spatial characteristics along rice root. J. Exp. Bot. 2018, 69, 5279–5291. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2002, 25, 402–408. [Google Scholar] [CrossRef]
- Cheng, N.H.; Pittman, J.K.; Zhu, J.K.; Hirschi, K.D. The protein kinase SOS2 activates the Arabidopsis H+/Ca2+ antiporter CAX1 to integrate calcium transport and salt tolerance. J. Biol. Chem. 2004, 279, 2922–2926. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Connorton, J.M.; Guo, Y.; Li, X.; Shigaki, T.; Hirschi, K.D.; Pittman, J.K. Functional studies of split Arabidopsis Ca2+/H+ exchangers. J. Biol. Chem. 2009, 284, 34075–34083. [Google Scholar] [CrossRef] [Green Version]
- Punshon, T.; Hirschi, K.; Yang, J.; Lanzirotti, A.; Lai, B.; Guerinot, M.L. The role of CAX1 and CAX3 in elemental distribution and abundance in Arabidopsis seed. Plant Physiol. 2012, 158, 352–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Fan, X.; Qian, K.; Zhang, Y.; Song, M.; Liu, Y.; Xu, G.; Fan, X. pOsNAR2.1:OsNAR2.1 expression enhances nitrogen uptake efficiency and grain yield in transgenic rice plants. Plant Biotechnol. J. 2017, 15, 1273–1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naz, M.; Luo, B.; Guo, X.; Li, B.; Chen, J.; Fan, X. Overexpression of Nitrate Transporter OsNRT2.1 Enhances Nitrate-Dependent Root Elongation. Genes 2019, 10, 290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bart, R.; Chern, M.; Park, C.J.; Bartley, L.; Ronald, P.C. A novel system for gene silencing using siRNAs in rice leaf and stem-derived protoplasts. Plant Methods 2006, 2, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Locus | Length of CDS (bp) | No. of Amino Acids (aa) | Chromosome | MW (KDa) | pI | Type | No. of Transmembrane Domains |
|---|---|---|---|---|---|---|---|---|
| OsCAX2 | LOC_Os03g27960 | 1317 | 439 | 3 | 47.48 | 4.57 | IB | 10 |
| OsCAX3 | LOC_Os04g55940 | 1254 | 418 | 4 | 45.45 | 4.76 | IB | 11 |
| OsCAX4 | LOC_Os02g04630 | 1089 | 363 | 2 | 39.07 | 7.00 | IB | 8 |
| OsCAX1a | LOC_Os01g37690 | 1356 | 452 | 1 | 47.71 | 6.78 | IA | 10 |
| OsCAX1b | LOC_Os05g51610 | 1362 | 454 | 5 | 49.11 | 6.34 | IA | 10 |
| OsCAX1c | LOC_Os02g21009 | 1353 | 451 | 2 | 48.16 | 5.62 | IA | 8 |
| AtCAX1 | AT2G38170 | 1428 | 475 | 2 | 51.64 | 6.25 | IA | 10 |
| AtCAX2 | AT3G13320 | 1326 | 441 | 3 | 48.21 | 4.45 | IB | 10 |
| AtCAX3 | AT3G51860 | 1380 | 459 | 3 | 49.85 | 5.39 | IA | 11 |
| AtCAX4 | AT5G01490 | 1365 | 454 | 5 | 49.61 | 6.51 | IA | 10 |
| AtCAX5 | AT1G55730 | 1326 | 441 | 1 | 48.10 | 4.61 | IB | 10 |
| AtCAX6 | AT1G55720 | 1404 | 467 | 1 | 51.84 | 5.40 | IB | 10 |
| % Identity | OsCAX1a | OsCAX1b | OsCAX1c | OsCAX2 | OsCAX3 | OsCAX4 | AtCAX1 | AtCAX2 | AtCAX3 | AtCAX4 | AtCAX5 | AtCAX6 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| % | % | % | % | % | % | % | % | % | % | % | % | |
| OsCAX1a | 100 | |||||||||||
| OsCAX1b | 69 | 100 | ||||||||||
| OsCAX1c | 59 | 58 | 100 | |||||||||
| OsCAX2 | 50 | 52 | 43 | 100 | ||||||||
| OsCAX3 | 49 | 51 | 40 | 74 | 100 | |||||||
| OsCAX4 | 36 | 38 | 31 | 55 | 55 | 100 | ||||||
| AtCAX1 | 64 | 62 | 57 | 48 | 49 | 38 | 100 | |||||
| AtCAX2 | 46 | 49 | 44 | 72 | 72 | 54 | 46 | 100 | ||||
| AtCAX3 | 67 | 66 | 56 | 50 | 48 | 39 | 79 | 48 | 100 | |||
| AtCAX4 | 56 | 59 | 49 | 46 | 47 | 34 | 57 | 47 | 59 | 100 | ||
| AtCAX5 | 48 | 49 | 44 | 72 | 73 | 52 | 48 | 87 | 49 | 47 | 100 | |
| AtCAX6 | 47 | 45 | 38 | 66 | 65 | 53 | 46 | 82 | 47 | 45 | 88 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, W.; Chen, J.; Meng, L.; Chen, D.; He, H.; Ye, G. The Rice Cation/H+ Exchanger Family Involved in Cd Tolerance and Transport. Int. J. Mol. Sci. 2021, 22, 8186. https://doi.org/10.3390/ijms22158186
Zou W, Chen J, Meng L, Chen D, He H, Ye G. The Rice Cation/H+ Exchanger Family Involved in Cd Tolerance and Transport. International Journal of Molecular Sciences. 2021; 22(15):8186. https://doi.org/10.3390/ijms22158186
Chicago/Turabian StyleZou, Wenli, Jingguang Chen, Lijun Meng, Dandan Chen, Haohua He, and Guoyou Ye. 2021. "The Rice Cation/H+ Exchanger Family Involved in Cd Tolerance and Transport" International Journal of Molecular Sciences 22, no. 15: 8186. https://doi.org/10.3390/ijms22158186
APA StyleZou, W., Chen, J., Meng, L., Chen, D., He, H., & Ye, G. (2021). The Rice Cation/H+ Exchanger Family Involved in Cd Tolerance and Transport. International Journal of Molecular Sciences, 22(15), 8186. https://doi.org/10.3390/ijms22158186