
Sex-Based Impact of Creatine Supplementation on Depressive Symptoms, Brain Serotonin and SSRI Efficacy in an Animal Model of Treatment-Resistant Depression

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Impact of Duration at Altitude on Brain Creatine
2.2. Dietary CRMH Supplementation: CRMH Dosage and Impact on Body Weight
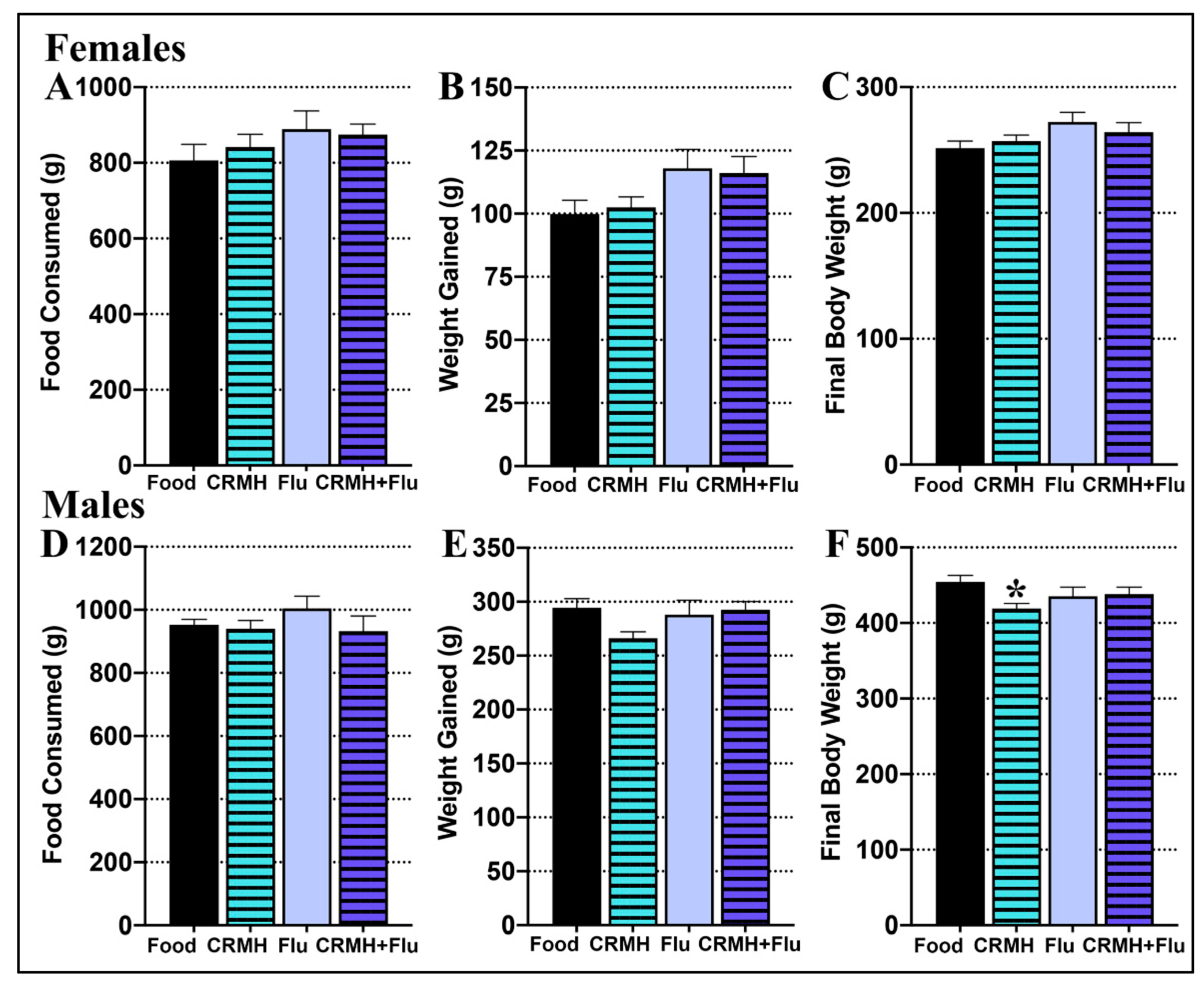
2.2.1. Food Consumed
2.2.2. Body Weight Gain
2.2.3. Final Body Weight
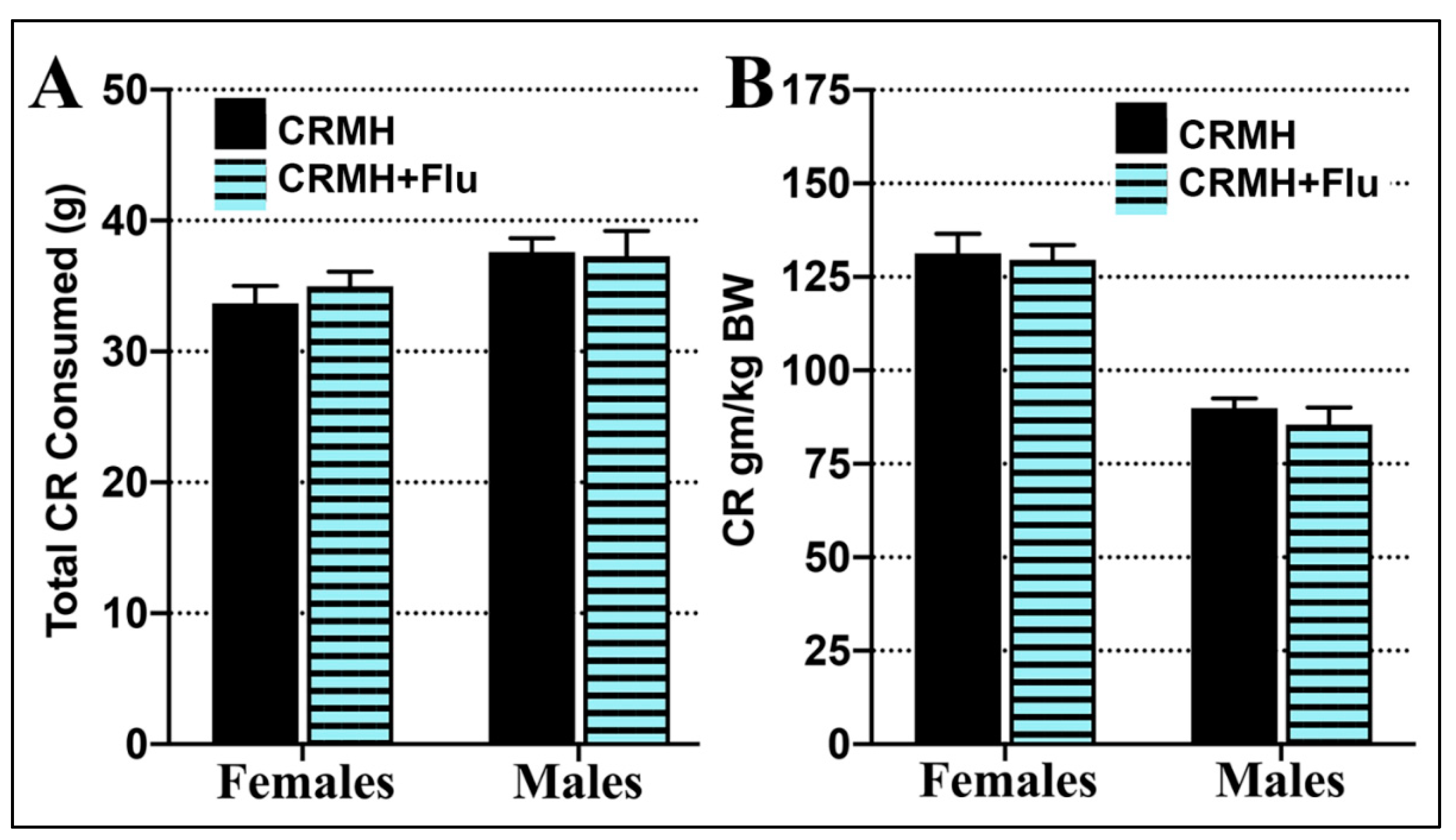
2.2.4. Creatine Consumption
2.3. Creatine Monohydrate Dietary Supplementation (CRDS) Study
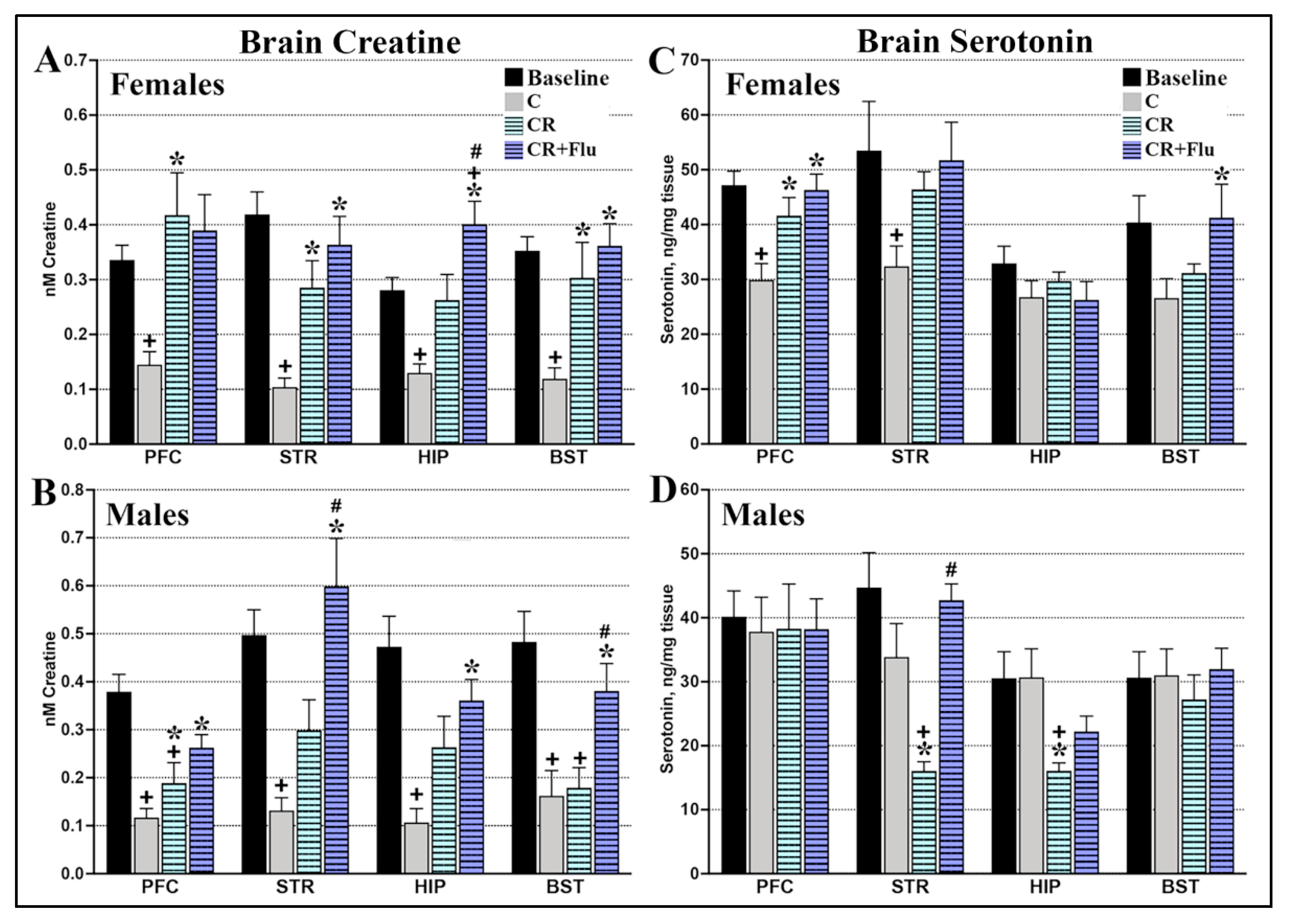
2.3.1. CRDS Impact on Blood and Brain Creatine
2.3.2. CRDS Impact on Brain Serotonin
2.3.3. CRDS Impact on Behavior in the Sucrose Preference Test
2.3.4. CRDS Impact on Behavior in the Open Field Test
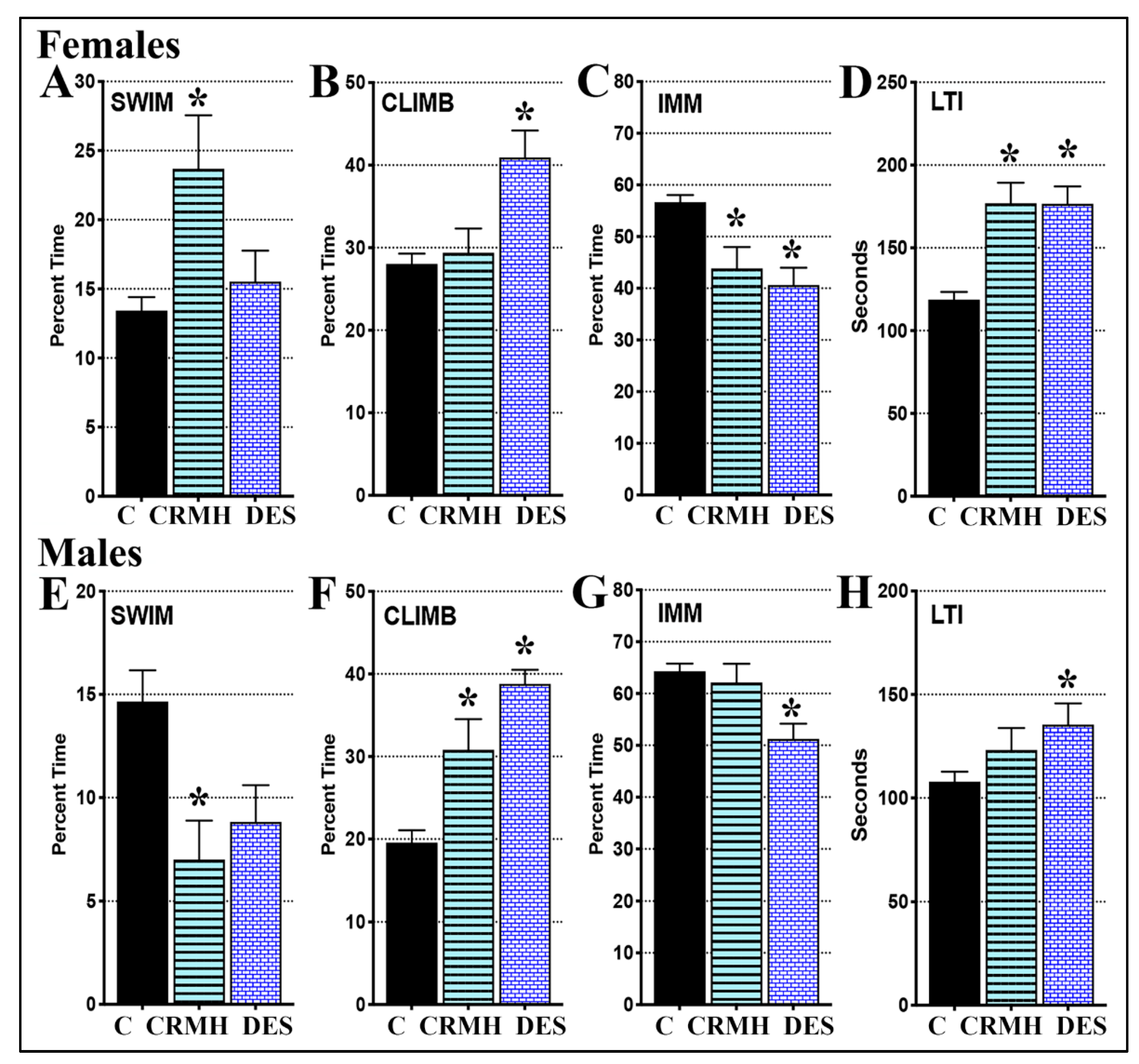
2.3.5. CRDS Impact on Behavior in the Forced Swim Test
2.4. CRDS + SSRI Study
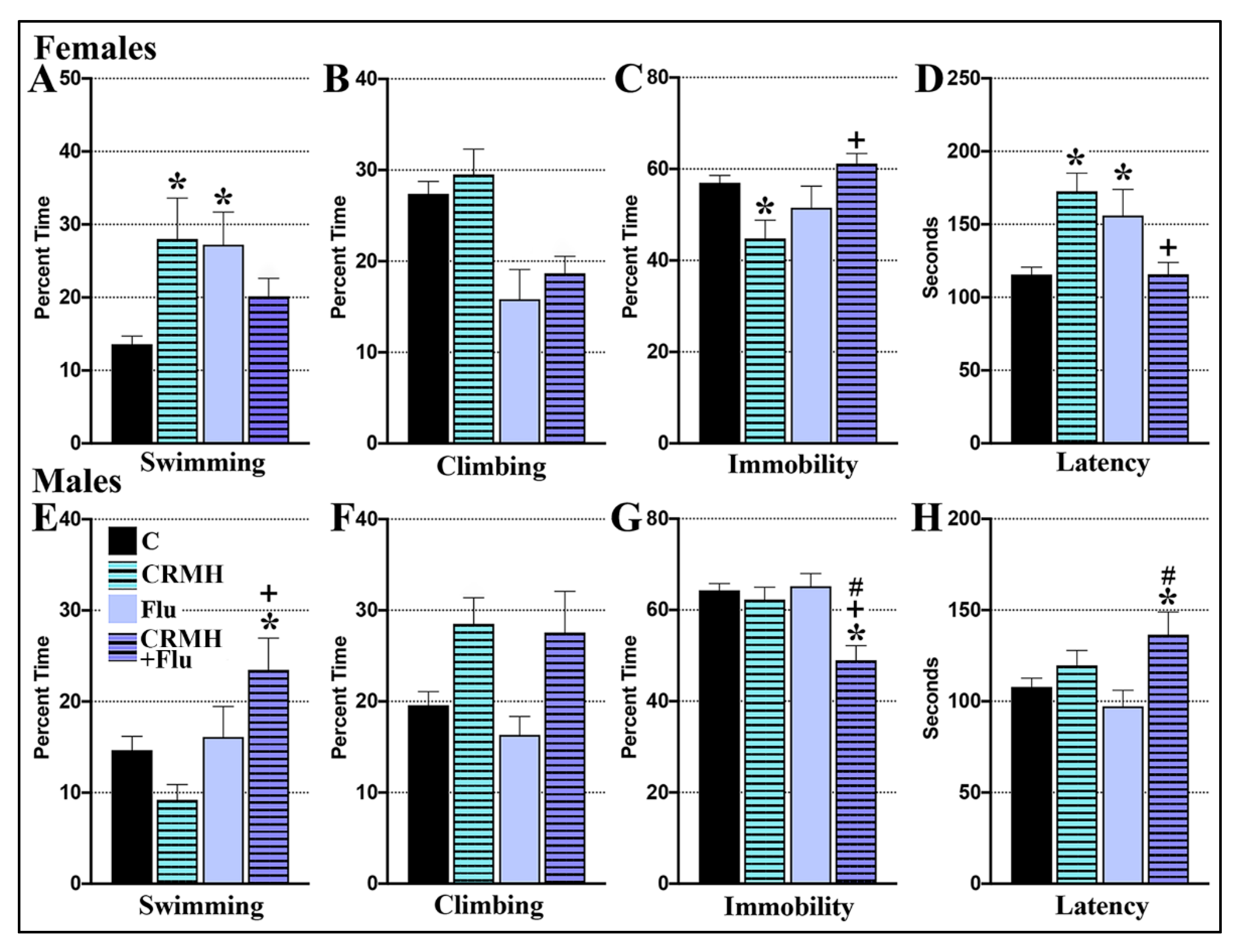
2.4.1. CRDS Impact on SSRI Function in the Forced Swim Test
2.4.2. CRDS + SSRI Impact on Brain Creatine
2.4.3. CRDS + SSRI Impact on Brain Serotonin
3. Discussion
Limitations of Study
4. Materials and Methods
4.1. Animals
4.2. Study Groups
4.3. Drug and Compounds
4.4. Dietary Supplementation
4.5. Forced Swim Test
4.6. Antidepressant Treatment
4.7. Sucrose Preference Test
4.8. Open Field Test
4.9. Dissections
4.10. Tissue Creatine Analysis
4.11. Brain Serotonin Analysis
4.12. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mrazek, D.A.; Hornberger, J.C.; Altar, C.A.; Degtiar, I. A review of the clinical, economic, and societal burden of treatment-resistant depression: 1996–2013. Psychiatr. Serv. 2014, 65, 977–987. [Google Scholar] [CrossRef]
- NIMH. Mental Health Information: Depression. Available online: https://www.nimh.nih.gov/health/topics/depression/index.shtml (accessed on 1 July 2021).
- Al-Harbi, K.S. Treatment-resistant depression: Therapeutic trends, challenges, and future directions. Patient Prefer. Adherence 2012, 6, 369–388. [Google Scholar] [CrossRef] [Green Version]
- DelMastro, K.; Hellem, T.; Kim, N.; Kondo, D.; Sung, Y.H.; Renshaw, P.F. Incidence of major depressive episode correlates with elevation of substate region of residence. J. Affect. Disord. 2011, 129, 376–379. [Google Scholar] [CrossRef] [Green Version]
- Gamboa, J.L.; Caceda, R.; Arregui, A. Is depression the link between suicide and high altitude? High Alt. Med. Biol. 2011, 12, 403–405. [Google Scholar] [CrossRef]
- Zaeh, S.; Miele, C.H.; Putcha, N.; Gilman, R.H.; Miranda, J.J.; Bernabe-Ortiz, A.; Wise, R.A.; Checkley, W. Chronic respiratory disease and high altitude are associated with depressive symptoms in four diverse settings. Int. J. Tuber. Lung Dis. 2016, 20, 1263–1269. [Google Scholar] [CrossRef]
- Risal, A.; Manandhar, K.; Linde, M.; Steiner, T.J.; Holen, A. Anxiety and depression in Nepal: Prevalence, comorbidity and associations. BMC Psychiatry 2016, 16, 102. [Google Scholar] [CrossRef] [Green Version]
- Asiri, S. Suicidal ideation among patients with major depressive disorder living at high altitude. Med. J. Cairo Univ. 2014, 82, 223–228. [Google Scholar]
- Haws, C.A.; Gray, D.D.; Yurgelun-Todd, D.A.; Moskos, M.; Meyer, L.J.; Renshaw, P.F. The possible effect of altitude on regional variation in suicide rates. Med. Hypotheses 2009, 73, 587–590. [Google Scholar] [CrossRef]
- Kim, N.; Mickelson, J.B.; Brenner, B.E.; Haws, C.A.; Yurgelun-Todd, D.A.; Renshaw, P.F. Altitude, gun ownership, rural areas, and suicide. Am. J. Psychiatry 2011, 168, 49–54. [Google Scholar] [CrossRef]
- Brenner, B.; Cheng, D.; Clark, S.; Camargo, C.A., Jr. Positive association between altitude and suicide in 2584 U.S. counties. High Alt. Med. Biol. 2011, 12, 31–35. [Google Scholar] [CrossRef] [Green Version]
- Helbich, M.; Bluml, V.; Leitner, M.; Kapusta, N.D. Does altitude moderate the impact of lithium on suicide? A spatial analysis of Austria. Geospat. Health 2013, 7, 209–218. [Google Scholar] [CrossRef] [Green Version]
- Alameda-Palacios, J.; Ruiz-Ramos, M.; Garcia-Robredo, B. Suicide mortality in Andalusia, Spain: Geographical distribution and relationship with antidepressants, altitude and socioeconomic inequalities. Rev. Esp. Salud Publica 2015, 89, 283–293. [Google Scholar] [CrossRef] [Green Version]
- Bocchetta, A.; Traccis, F. The Sardinian puzzle: Concentration of major psychoses and suicide in the same sub-regions across one century. Clin. Pract. Epidemiol. Ment. Health 2017, 13, 246–254. [Google Scholar] [CrossRef] [Green Version]
- Webb, R.T.; Kontopantelis, E.; Doran, T.; Qin, P.; Creed, F.; Kapur, N. Suicide risk in primary care patients with major physical diseases: A case-control study. Arch. Gen. Psychiatry 2012, 69, 256–264. [Google Scholar] [CrossRef]
- Goodwin, R.D. Asthma and suicide: Current knowledge and future directions. Curr. Psychiatry Rep. 2012, 14, 30–35. [Google Scholar] [CrossRef]
- Goodwin, R.D. Is COPD associated with suicide behavior? J. Psychiatr. Res. 2011, 45, 1269–1271. [Google Scholar] [CrossRef]
- Goodwin, R.D.; Robinson, M.; Sly, P.D.; McKeague, I.W.; Susser, E.S.; Zubrick, S.R.; Stanley, F.J.; Mattes, E. Severity and persistence of asthma and mental health: A birth cohort study. Psychol. Med. 2013, 43, 1313–1322. [Google Scholar] [CrossRef] [Green Version]
- Chung, J.H.; Han, C.H.; Park, S.C.; Kim, C.J. Suicidal ideation and suicide attempts in chronic obstructive pulmonary disease: The Korea National Health and Nutrition Examination Survey (KNHANES IV, V) from 2007–2012. NPJ Prim. Care Respir. Med. 2014, 24, 14094. [Google Scholar] [CrossRef] [Green Version]
- Goodwin, D.R.; Kroenke, K.; Hoven, C.W.; Spitzer, R.L. Major depression, physical illness, and suicidal ideation in primary care. Psychosom. Med. 2003, 65, 501–505. [Google Scholar] [CrossRef]
- Young, S.N. Elevated incidence of suicide in people living at altitude, smokers and patients with chronic obstructive pulmonary disease and asthma: Possible role of hypoxia causing decreased serotonin synthesis. J. Psychiatry Neurosci. 2013, 38, 423–426. [Google Scholar] [CrossRef] [Green Version]
- Katz, I.R. Is there a hypoxic affective syndrome? Psychosomatics 1982, 23, 846, 849–850, 852–853. [Google Scholar] [CrossRef]
- Crapo, R.O.; Jensen, R.L.; Hegewald, M.; Tashkin, D.P. Arterial blood gas reference values for sea level and an altitude of 1400 meters. Am. J. Respir. Crit. Care Med. 1999, 160, 1525–1531. [Google Scholar] [CrossRef]
- Shi, X.F.; Carlson, P.J.; Kim, T.S.; Sung, Y.H.; Hellem, T.L.; Fiedler, K.K.; Kim, S.E.; Glaeser, B.; Wang, K.; Zuo, C.S.; et al. Effect of altitude on brain intracellular pH and inorganic phosphate levels. Psychiatry Res. 2014, 222, 149–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prescot, A.; Huber, R.; Kanekar, S.; Kondo, D.; Prisciandaro, J.; Ongur, D.; Renshaw, P.F. Effect of moderate altitude on human cerebral metabolite levels: A preliminary, multi-site, proton magnetic resonance spectroscopy investigation. Psychiat Res. Neuroimaging 2021, 314, 111314. [Google Scholar] [CrossRef]
- Koundal, S.; Gandhi, S.; Kaur, T.; Khushu, S. Neurometabolic and structural alterations in rat brain due to acute hypobaric hypoxia: In vivo H MRS at 7 T. NMR Biomed. 2014, 27, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Bogdanova, O.V.; Abdullah, O.; Kanekar, S.; Bogdanov, V.B.; Prescot, A.P.; Renshaw, P.F. Neurochemical alterations in frontal cortex of the rat after one week of hypobaric hypoxia. Behav. Brain Res. 2014, 263, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Mayberg, H.S. Positron emission tomography imaging in depression: A neural systems perspective. Neuroimaging Clin. N. Am. 2003, 13, 805–815. [Google Scholar] [CrossRef]
- Rezin, G.T.; Amboni, G.; Zugno, A.I.; Quevedo, J.; Streck, E.L. Mitochondrial dysfunction and psychiatric disorders. Neurochem. Res. 2009, 34, 1021–1029. [Google Scholar] [CrossRef] [PubMed]
- Iosifescu, D.V.; Bolo, N.R.; Nierenberg, A.A.; Jensen, J.E.; Fava, M.; Renshaw, P.F. Brain bioenergetics and response to triiodothyronine augmentation in major depressive disorder. Biol. Psychiatry 2008, 63, 1127–1134. [Google Scholar] [CrossRef] [PubMed]
- Wallimann, T.; Wyss, M.; Brdiczka, D.; Nicolay, K.; Eppenberger, H.M. Intracellular compartmentation, structure and function of creatine kinase isoenzymes in tissues with high and fluctuating energy demands: The “phosphocreatine circuit” for cellular energy homeostasis. Biochem. J. 1992, 281, 21–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakian, A.V.; Huber, R.; Scholl, L.S.; Renshaw, P.F.; Kondo, D. Dietary creatine and depression risk among U.S. adults. Transl. Psychiatry 2020, 10, 52. [Google Scholar] [CrossRef] [Green Version]
- Lyoo, I.K.; Kong, S.W.; Sung, S.M.; Hirashima, F.; Parow, A.; Hennen, J.; Cohen, B.M.; Renshaw, P.F. Multinuclear magnetic resonance spectroscopy of high-energy phosphate metabolites in human brain following oral supplementation of creatine-monohydrate. Psychiatry Res. 2003, 123, 87–100. [Google Scholar] [CrossRef]
- Dechent, P.; Pouwels, P.J.; Wilken, B.; Hanefeld, F.; Frahm, J. Increase of total creatine in human brain after oral supplementation of creatine-monohydrate. Am. J. Physiol. 1999, 277, R698–R704. [Google Scholar] [CrossRef] [PubMed]
- Lyoo, I.K.; Yoon, S.; Kim, T.S.; Hwang, J.; Kim, J.E.; Won, W.; Bae, S.; Renshaw, P.F. A randomized, double-blind placebo-controlled trial of oral creatine monohydrate augmentation for enhanced response to a selective serotonin reuptake inhibitor in women with major depressive disorder. Am. J. Psychiatry 2012, 169, 937–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondo, D.G.; Sung, Y.H.; Hellem, T.L.; Fiedler, K.K.; Shi, X.; Jeong, E.K.; Renshaw, P.F. Open-label adjunctive creatine for female adolescents with SSRI-resistant major depressive disorder: A 31-phosphorus magnetic resonance spectroscopy study. J. Affect. Disord. 2011, 135, 354–361. [Google Scholar] [CrossRef] [Green Version]
- Kondo, D.G.; Forrest, L.N.; Shi, X.; Sung, Y.H.; Hellem, T.L.; Huber, R.S.; Renshaw, P.F. Creatine target engagement with brain bioenergetics: A dose-ranging phosphorus-31 magnetic resonance spectroscopy study of adolescent females with SSRI-resistant depression. Amino Acids 2016, 48, 1941–1954. [Google Scholar] [CrossRef] [Green Version]
- Allen, P.J.; D’Anci, K.E.; Kanarek, R.B.; Renshaw, P.F. Chronic creatine supplementation alters depression-like behavior in rodents in a sex-dependent manner. Neuropsychopharmacology 2010, 35, 534–546. [Google Scholar] [CrossRef] [Green Version]
- Allen, P.J.; D’Anci, K.E.; Kanarek, R.B.; Renshaw, P.F. Sex-specific antidepressant effects of dietary creatine with and without sub-acute fluoxetine in rats. Pharmacol. Biochem. Behav. 2012, 101, 588–601. [Google Scholar] [CrossRef] [Green Version]
- Kanekar, S.; Bogdanova, O.V.; Olson, P.R.; Sung, Y.H.; D’Anci, K.E.; Renshaw, P.F. Hypobaric hypoxia induces depression-like behavior in female Sprague-Dawley rats, but not in males. High Alt. Med. Biol. 2015, 16, 52–60. [Google Scholar] [CrossRef]
- Kanekar, S.; Sheth, C.; Ombach, H.; Brown, J.; Hoffman, M.; Ettaro, R.; Renshaw, P. Sex-based changes in rat brain serotonin and behavior in a model of altitude-related vulnerability to treatment-resistant depression. Psychopharmacology 2021. [Google Scholar] [CrossRef]
- Kanekar, S.; Sheth, C.S.; Ombach, H.J.; Olson, P.R.; Bogdanova, O.V.; Petersen, M.; Renshaw, C.E.; Sung, Y.H.; D’Anci, K.E.; Renshaw, P.F. Hypobaric hypoxia exposure in rats differentially alters antidepressant efficacy of the selective serotonin reuptake inhibitors fluoxetine, paroxetine, escitalopram and sertraline. Pharmacol. Biochem. Behav. 2018, 170, 25–35. [Google Scholar] [CrossRef]
- Vestergaard, M.B.; Lindberg, U.; Aachmann-Andersen, N.J.; Lisbjerg, K.; Christensen, S.J.; Law, I.; Rasmussen, P.; Olsen, N.V.; Larsson, H.B.W. Acute hypoxia increases the cerebral metabolic rate—A magnetic resonance imaging study. J. Cereb. Blood Flow Metab. 2016, 36, 1046–1058. [Google Scholar] [CrossRef] [Green Version]
- Pazini, F.L.; Cunha, M.P.; Rodrigues, A.L.S. The possible beneficial effects of creatine for the management of depression. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2019, 89, 193–206. [Google Scholar] [CrossRef]
- Iosifescu, D.V.; Renshaw, P.E. 31P-magnetic resonance spectroscopy and thyroid hormones in major depressive disorder: Toward a bioenergetic mechanism in depression? Harv. Rev. Psychiatry 2003, 11, 51–63. [Google Scholar]
- Ipsiroglu, O.S.; Stromberger, C.; Ilas, J.; Hoger, H.; Muhl, A.; Stockler-Ipsiroglu, S. Changes of tissue creatine concentrations upon oral supplementation of creatine-monohydrate in various animal species. Life Sci. 2001, 69, 1805–1815. [Google Scholar] [CrossRef]
- Christie, D.L. Functional insights into the creatine transporter. Subcell. Biochem. 2007, 46, 99–118. [Google Scholar]
- Mak, C.S.; Waldvogel, H.J.; Dodd, J.R.; Gilbert, R.T.; Lowe, M.T.; Birch, N.P.; Faull, R.L.; Christie, L.D. Immunohistochemical localisation of the creatine transporter in the rat brain. Neuroscience 2009, 163, 571–585. [Google Scholar] [CrossRef] [PubMed]
- Albert, P.R.; Vahid-Ansari, F.; Luckhart, C. Serotonin-prefrontal cortical circuitry in anxiety and depression phenotypes: Pivotal role of pre- and post-synaptic 5-HT1A receptor expression. Front. Behav. Neurosci. 2014, 8, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nye, J.A.; Purselle, D.; Plisson, C.; Voll, R.J.; Stehouwer, J.S.; Votaw, J.R.; Kilts, C.D.; Goodman, M.M.; Nemeroff, C.B. Decreased brainstem and putamen SERT binding potential in depressed suicide attempters using [11C]-zient PET imaging. Depress. Anxiety 2013, 30, 902–907. [Google Scholar] [CrossRef]
- Garcia-Garcia, A.L.; Meng, Q.; Canetta, S.; Gardier, A.M.; Guiard, B.P.; Kellendonk, C.; Dranovsky, A.; Leonardo, E.D. Serotonin Signaling through prefrontal cortex 5-HT1A receptors during adolescence can determine baseline mood-related behaviors. Cell. Rep. 2017, 18, 1144–1156. [Google Scholar] [CrossRef]
- Dale, E.; Pehrson, A.L.; Jeyarajah, T.; Li, Y.; Leiser, S.C.; Smagin, G.; Olsen, C.K.; Sanchez, C. Effects of serotonin in the hippocampus: How SSRIs and multimodal antidepressants might regulate pyramidal cell function. CNS Spectr. 2016, 21, 143–161. [Google Scholar] [CrossRef]
- Zmudzka, E.; Salaciak, K.; Sapa, J.; Pytka, K. Serotonin receptors in depression and anxiety: Insights from animal studies. Life Sci. 2018, 210, 106–124. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.K. Hypoxia. 3. Hypoxia and neurotransmitter synthesis. Am. J. Physiol. Cell. Physiol. 2011, 300, C743–C751. [Google Scholar] [CrossRef] [Green Version]
- Sheth, C.S.; Ombach, H.; Brown, J.; Renshaw, P.F.; Kanekar, S. Living at moderate altitude may alter rain monoamines to worsen mood and substance use disorders: A sex-based animal model study, in American College of Neuropsychopharmacology. Neuropsychopharmacology 2018, 43, S451. [Google Scholar]
- Damsa, C.; Bumb, A.; Bianchi-Demicheli, F.; Vidailhet, P.; Sterck, R.; Andreoli, A.; Beyenburg, S. “Dopamine-dependent” side effects of selective serotonin reuptake inhibitors: A clinical review. J. Clin. Psychiatry 2004, 65, 1064–1068. [Google Scholar] [CrossRef]
- Morelli, E.; Moore, H.; Rebello, T.J.; Gray, N.; Steele, K.; Esposito, E.; Gingrich, J.A.; Ansorge, M.S. Chronic 5-HT transporter blockade reduces DA signaling to elicit basal ganglia dysfunction. J. Neurosci. 2011, 31, 15742–15750. [Google Scholar] [CrossRef]
- Smeland, O.B.; Meisingset, T.W.; Borges, K.; Sonnewald, U. Chronic acetyl-L-carnitine alters brain energy metabolism and increases noradrenaline and serotonin content in healthy mice. Neurochem. Int. 2012, 61, 100–107. [Google Scholar] [CrossRef]
- Santos, P.M.; Scaini, G.; Rezin, G.T.; Benedet, J.; Rochi, N.; Jeremias, G.C.; Carvalho-Silva, M.; Quevedo, J.; Streck, E.L. Brain creatine kinase activity is increased by chronic administration of paroxetine. Brain Res. Bull. 2009, 80, 327–330. [Google Scholar] [CrossRef]
- Reus, G.Z.; Stringari, R.B.; Rezin, G.T.; Fraga, D.B.; Daufenbach, J.F.; Scaini, G.; Benedet, J.; Rochi, N.; Streck, E.L.; Quevedo, J. Administration of memantine and imipramine alters mitochondrial respiratory chain and creatine kinase activities in rat brain. J. Neural Transm. 2012, 119, 481–491. [Google Scholar] [CrossRef]
- Wyss, M.; Kaddurah-Daouk, R. Creatine and creatinine metabolism. Physiol. Rev. 2000, 80, 1107–1213. [Google Scholar] [CrossRef]
- Wyss, M.; Schulze, A. Health implications of creatine: Can oral creatine supplementation protect against neurological and atherosclerotic disease? Neuroscience 2002, 112, 243–260. [Google Scholar] [CrossRef]
- Cunha, M.P.; Pazini, F.L.; Lieberknecht, V.; Rodrigues, A.L.S. Subchronic administration of creatine produces antidepressant-like effect by modulating hippocampal signaling pathway mediated by FNDC5/BDNF/Akt in mice. J. Psychiatr. Res. 2018, 104, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Pazini, F.L.; Cunha, M.P.; Rosa, J.M.; Colla, A.R.; Lieberknecht, V.; Oliveira, A.; Rodrigues, A.L. Creatine, similar to ketamine, counteracts depressive-like behavior induced by corticosterone via PI3K/Akt/mTOR pathway. Mol. Neurobiol. 2016, 53, 6818–6834. [Google Scholar] [CrossRef]
- Oliveira, M.S.; Furian, A.F.; Fighera, M.R.; Fiorenza, N.G.; Ferreira, J.; Rubin, M.A.; Mello, C.F.; Royes, L.F. The involvement of the polyamines binding sites at the NMDA receptor in creatine-induced spatial learning enhancement. Behav. Brain Res. 2008, 187, 200–204. [Google Scholar] [CrossRef]
- Rapoport, S.I.; Lust, W.D.; Fredericks, W.R. Effects of hypoxia on rat brain metabolism: Unilateral in vivo carotid infusion. Exp. Neurol. 1986, 91, 319–330. [Google Scholar] [CrossRef]
- Engl, E.; Garvert, M.M. A prophylactic role for creatine in hypoxia? J. Neurosci. 2015, 35, 9249–9251. [Google Scholar] [CrossRef] [Green Version]
- Turner, C.E.; Byblow, W.D.; Gant, N. Creatine supplementation enhances corticomotor excitability and cognitive performance during oxygen deprivation. J. Neurosci. 2015, 35, 1773–1780. [Google Scholar] [CrossRef]
- Balestrino, M.; Lensman, M.; Parodi, M.; Perasso, L.; Rebaudo, R.; Melani, R.; Polenov, S.; Cupello, A. Role of creatine and phosphocreatine in neuronal protection from anoxic and ischemic damage. Amino Acids 2002, 23, 221–229. [Google Scholar] [CrossRef]
- Perasso, L.; Lunardi, G.L.; Risso, F.; Pohvozcheva, A.V.; Leko, M.V.; Gandolfo, C.; Florio, T.; Cupello, A.; Burov, S.V.; Balestrino, M. Protective effects of some creatine derivatives in brain tissue anoxia. Neurochem. Res. 2008, 33, 765–775. [Google Scholar] [CrossRef]
- Marques, E.P.; Wyse, A.T.S. Creatine as a neuroprotector: An actor that can play many parts. Neurotox. Res. 2019, 36, 411–423. [Google Scholar] [CrossRef]
- Fiedler, K.K.; Kim, N.; Kondo, D.G.; Renshaw, P.F. Cocaine use in the past year is associated with altitude of residence. J. Addict. Med. 2012, 6, 166–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.S.; Kondo, D.G.; Kim, N.; Renshaw, P.F. Altitude may contribute to regional variation in methamphetamine use in the United States: A population database study. Psychiatry Investig. 2014, 11, 430–436. [Google Scholar] [CrossRef] [Green Version]
- Ombach, H.J.; Scholl, L.S.; Bakian, A.V.; Renshaw, K.T.; Sung, Y.H.; Renshaw, P.F.; Kanekar, S. Association between altitude, prescription opioid misuse, and fatal overdoses. Addict. Behav. Rep. 2019, 9, 100167. [Google Scholar] [CrossRef] [PubMed]
- Hellem, T.L.; Sung, H.; Shi, X.F.; Pett, M.A.; Latendresse, G.; Morgan, J.; Huber, R.S.; Kuykendall, D.; Lundberg, K.J.; Renshaw, P.F. Creatine as a novel treatment for depression in females using methamphetamine: A pilot study. J. Dual Diagn. 2015, 11, 189–202. [Google Scholar] [CrossRef] [Green Version]
- Toler, J.; Gleason, D.; Clare, L.; Buckham, S.; DeWit, R. Disposable plastic rodent feeders for use in an automated filling system. Lab. Anim. 2008, 37, 415–419. [Google Scholar] [CrossRef]
- Detke, M.J.; Lucki, I. Detection of serotonergic and noradrenergic antidepressants in the rat forced swimming test: The effects of water depth. Behav. Brain. Res. 1996, 73, 43–46. [Google Scholar] [CrossRef]
- Bogdanova, O.V.; Kanekar, S.; D’Anci, K.E.; Renshaw, P.F. Factors influencing behavior in the forced swim test. Physiol. Behav. 2013, 118, 227–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slattery, D.A.; Cryan, J.F. Using the rat forced swim test to assess antidepressant-like activity in rodents. Nat. Protoc. 2012, 7, 1009–1014. [Google Scholar] [CrossRef]
- Borsini, F.; Meli, A. Is the forced swimming test a suitable model for revealing antidepressant activity? Psychopharmacology 1988, 94, 147–160. [Google Scholar] [CrossRef] [PubMed]
- Detke, M.J.; Rickels, M.; Lucki, I. Active behaviors in the rat forced swimming test differentially produced by serotonergic and noradrenergic antidepressants. Psychopharmacology 1995, 121, 66–72. [Google Scholar] [CrossRef]
- Overstreet, D.H. Modeling depression in animal models. Methods Mol. Biol. 2012, 829, 125–144. [Google Scholar] [PubMed]
- Sheth, C.; Ombach, H.; Olson, P.; Renshaw, P.F.; Kanekar, S. Increased anxiety and anhedonia in female rats following exposure to altitude. High Alt. Med. Biol. 2018, 19, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Heffner, T.G.; Hartman, J.A.; Seiden, L.S. A rapid method for the regional dissection of the rat brain. Pharmacol. Biochem. Behav. 1980, 13, 453–456. [Google Scholar] [CrossRef]
- Paxinos, G.W.C. The Rat Brain in Stereotaxic Coordinates; Academic Press: Cambridge, MA, USA, 1998. [Google Scholar]
- Mateos, S.S.; Sanchez, C.L.; Paredes, S.D.; Barriga, C.; Rodriguez, A.B. Circadian levels of serotonin in plasma and brain after oral administration of tryptophan in rats. Basic Clin. Pharmacol. Toxicol. 2009, 104, 52–59. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kanekar, S.; Ettaro, R.; Hoffman, M.D.; Ombach, H.J.; Brown, J.; Lynch, C.; Sheth, C.S.; Renshaw, P.F. Sex-Based Impact of Creatine Supplementation on Depressive Symptoms, Brain Serotonin and SSRI Efficacy in an Animal Model of Treatment-Resistant Depression. Int. J. Mol. Sci. 2021, 22, 8195. https://doi.org/10.3390/ijms22158195
Kanekar S, Ettaro R, Hoffman MD, Ombach HJ, Brown J, Lynch C, Sheth CS, Renshaw PF. Sex-Based Impact of Creatine Supplementation on Depressive Symptoms, Brain Serotonin and SSRI Efficacy in an Animal Model of Treatment-Resistant Depression. International Journal of Molecular Sciences. 2021; 22(15):8195. https://doi.org/10.3390/ijms22158195
Chicago/Turabian StyleKanekar, Shami, Robert Ettaro, Michael D. Hoffman, Hendrik J. Ombach, Jadeda Brown, Cayla Lynch, Chandni S. Sheth, and Perry F. Renshaw. 2021. "Sex-Based Impact of Creatine Supplementation on Depressive Symptoms, Brain Serotonin and SSRI Efficacy in an Animal Model of Treatment-Resistant Depression" International Journal of Molecular Sciences 22, no. 15: 8195. https://doi.org/10.3390/ijms22158195
APA StyleKanekar, S., Ettaro, R., Hoffman, M. D., Ombach, H. J., Brown, J., Lynch, C., Sheth, C. S., & Renshaw, P. F. (2021). Sex-Based Impact of Creatine Supplementation on Depressive Symptoms, Brain Serotonin and SSRI Efficacy in an Animal Model of Treatment-Resistant Depression. International Journal of Molecular Sciences, 22(15), 8195. https://doi.org/10.3390/ijms22158195