
Phytochemicals in Malignant Pleural Mesothelioma Treatment—Review on the Current Trends of Therapies


Abstract
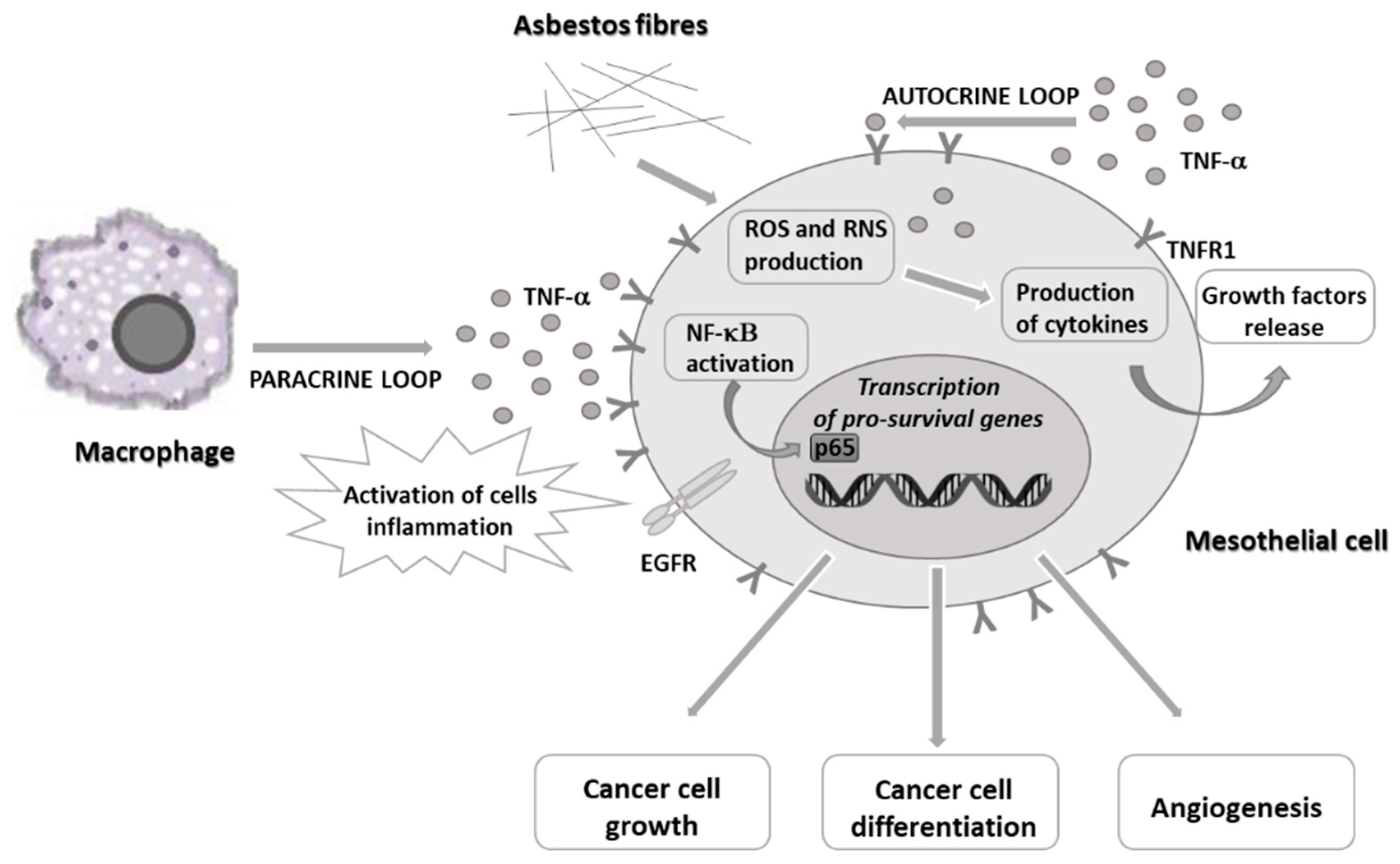
:1. Introduction
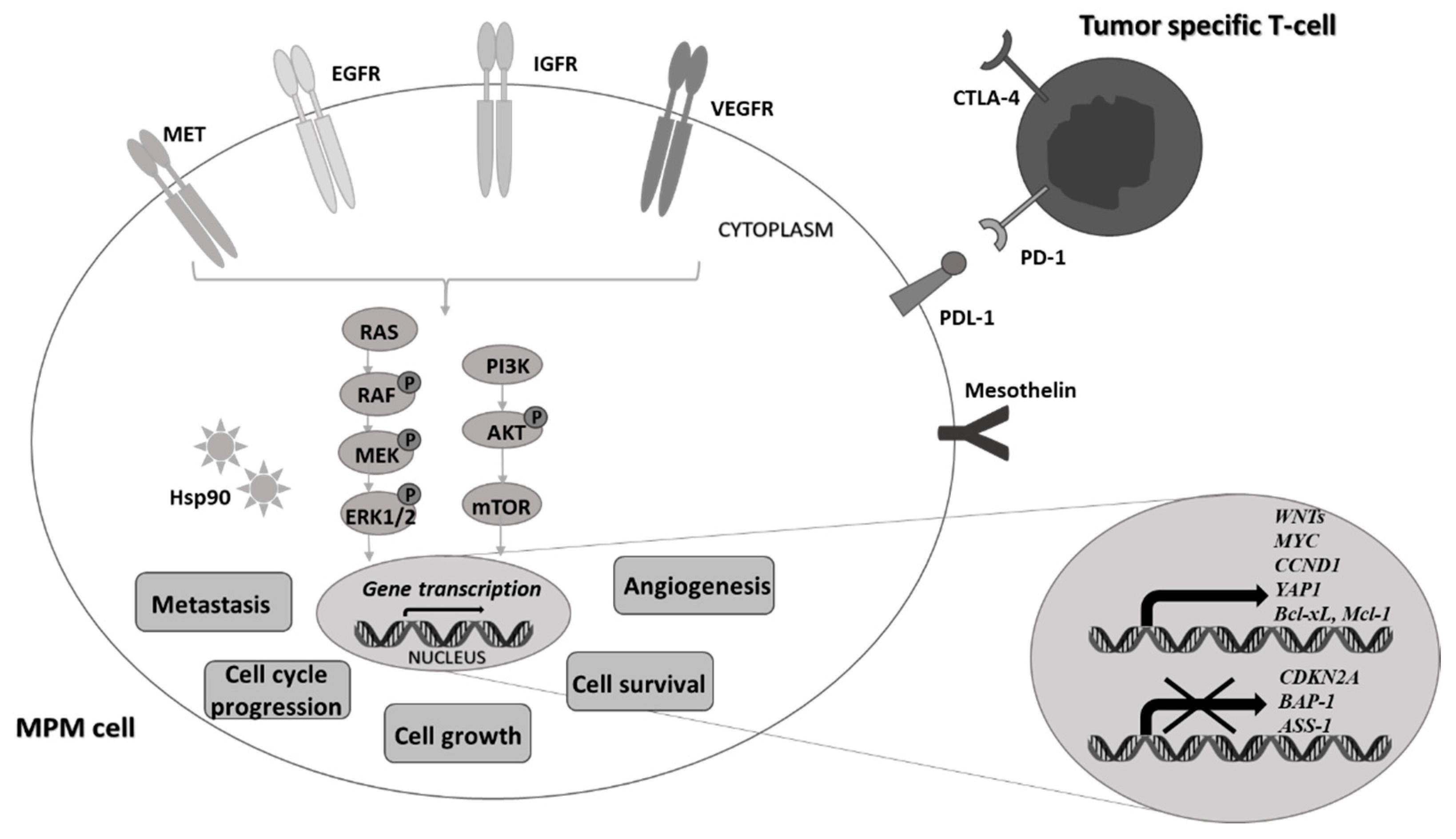
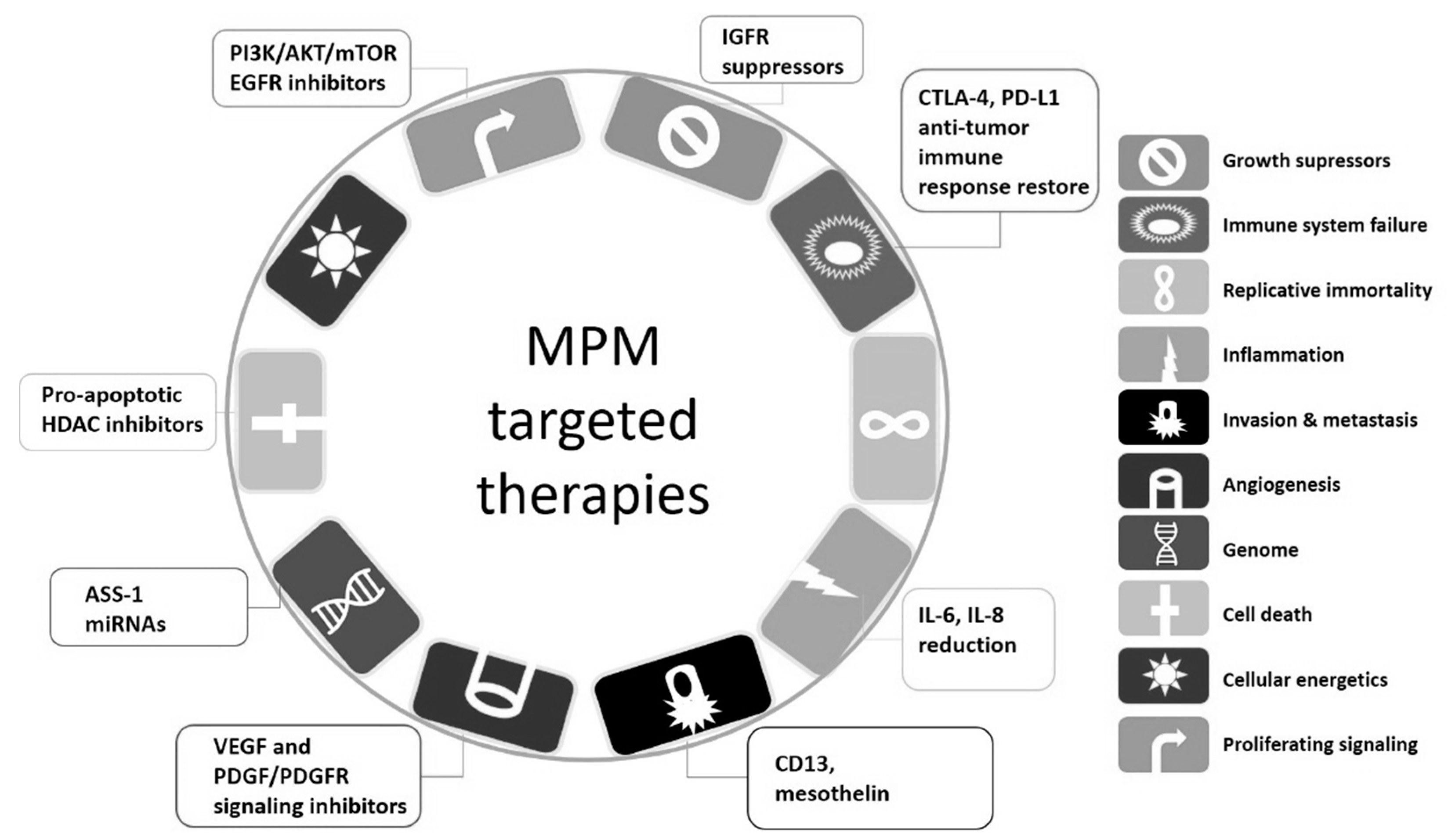
2. Novel Targeted Therapies against Malignant Pleural Mesothelioma Cells
3. Polyphenols and Natural Compounds in Cancer Treatment
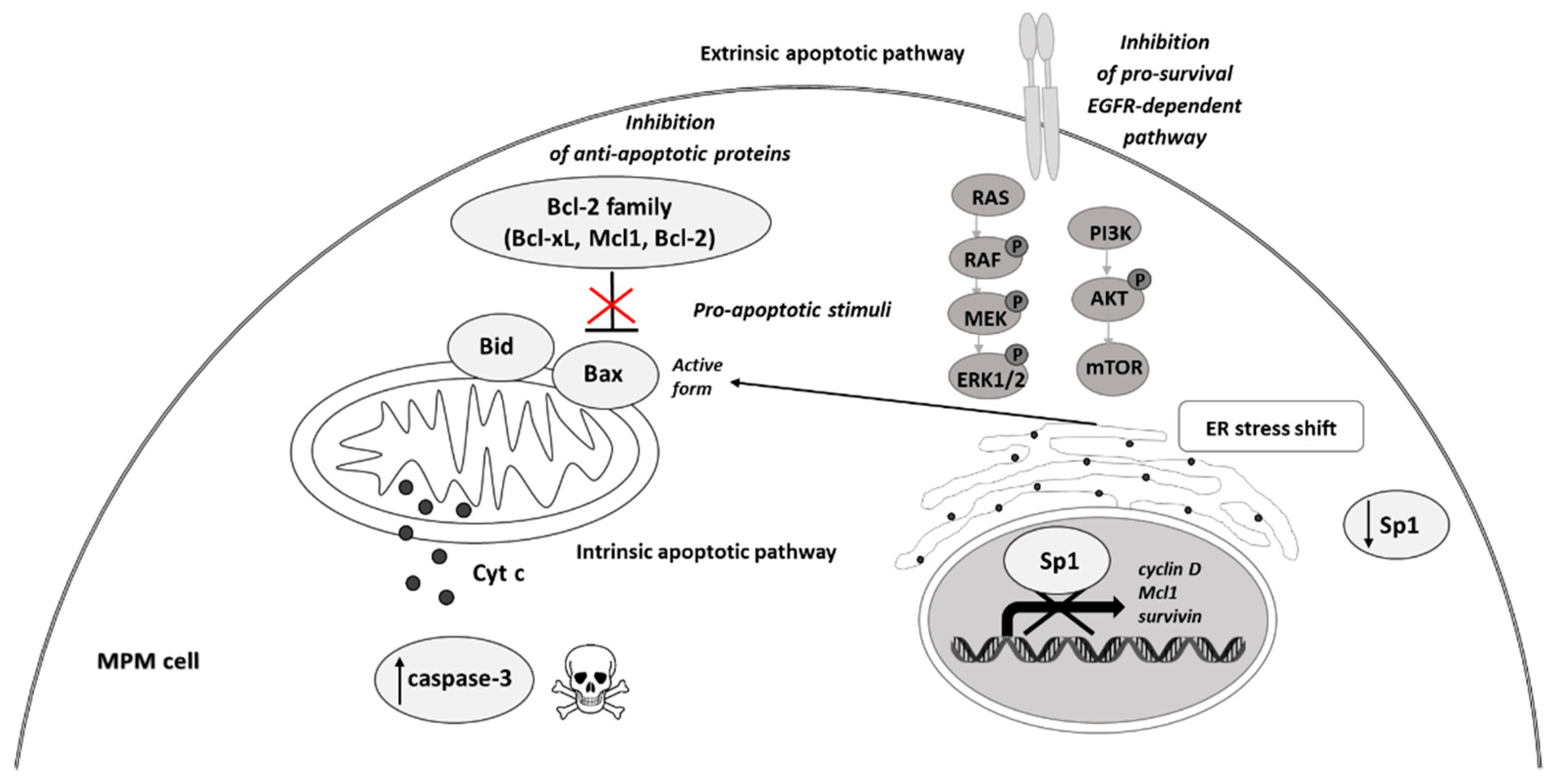
4. Polyphenols in Malignant Pleural Mesothelioma Therapy
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Stahel, R.A.; Weder, W.; Lievens, Y.; Felip, E. Malignant pleural mesothelioma: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. On behalf of the ESMO Guidelines Working Group. Ann. Oncol. 2010, 21 (Suppl. 5), v126–v128. [Google Scholar] [CrossRef] [PubMed]
- Parkin, D.M.; Bray, F.; Ferlay, J.; Pisani, P. Global cancer statistics, 2002. CA Cancer J. Clin. 2005, 55, 74–108. [Google Scholar] [CrossRef] [PubMed]
- Kameda, T.; Takahashi, K.; Kim, R.; Jiang, Y.; Movahed, M.; Park, E.K.; Rantanen, J. Asbestos: Use, bans and disease burden in Europe. Bull. World Health Organ 2014, 92, 790–797. [Google Scholar] [CrossRef] [PubMed]
- Acencio, M.M.; Soares, B.; Marchi, E.; Silva, C.S.; Teixeira, L.R.; Broaddus, V.C. Inflammatory cytokines contribute to asbestos-induced injury of mesothelial cells. Lung 2015, 193, 831–837. [Google Scholar] [CrossRef] [PubMed]
- Carbone, M.; Yang, H. Molecular pathways: Targeting mechanisms of asbestos and erionite carcinogenesis in mesothelioma. Clin. Cancer Res. 2012, 18, 598–604. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Rivera, Z.; Jube, S.; Nasu, M.; Bertino, P.; Goparaju, C.; Franzoso, G.; Lotze, M.T.; Krausz, T.; Pass, H.I.; et al. Programmed necrosis induced by asbestos in human mesothelial cells causes high-mobility group box 1 protein release and resultant inflammation. Proc. Nat. Acad. Sci. USA 2010, 107, 12611–12616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zucali, P.A.; Ceresoli, G.L.; de Vincenzo, F.; Simonelli, M.; Lorenzi, E.; Gianoncelli, L.; Santoro, A. Advances in the biology of malignant pleural mesotheliomamesothalioma. Cancer Treat. Rev. 2011, 37, 543–558. [Google Scholar] [CrossRef]
- Yang, H.; Bocchetta, M.; Kroczynska, B.; Elmishad, A.G.; Chen, Y.; Liu, Z.; Bubici, C.; Mossman, B.T.; Pass, H.I.; Testa, J.R.; et al. TNF-alpha inhibits asbestos-induced cytotoxicity via a NF-kappa-B-dependent pathway, a possible mechanism for asbestos-induced oncogenesis. Proc. Natl. Acad. Sci. USA 2006, 103, 10397–10402. [Google Scholar] [CrossRef] [Green Version]
- Jean, D.; Daubriac, J.; Le Pimpec-Barthes, F.; Galateau-Salle, F.; Jaurand, M.C. Molecular changes in mesothelioma with an impact on prognosis and treatment. Arch. Pathol. Lab. Med. 2012, 136, 277–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Kadariya, Y.; Cheung, M.; Pei, J.; Talarchek, J.; Sementino, E.; Tan, Y.; Menges, C.W.; Cai, K.Q.; Litwin, S.; et al. Germline mutation of BAP1 accelerates development of asbestos-induced malignant mesothelioma. Cancer Res. 2014, 74, 4388–4397. [Google Scholar] [CrossRef] [Green Version]
- Szlosarek, P.W.; Klabatsa, A.; Pallaska, A.; Sheaff, M.; Smith, P.; Crook, T.; Grimshaw, M.J.; Steele, J.P.; Rudd, R.M.; Balkwill, F.R.; et al. In Vivo loss of expression of argininosuccinate synthetase in malignant pleural mesothelioma is a biomarker for susceptibility to arginine depletion. Clin. Cancer Res. 2006, 12, 7126–7131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chew, S.H.; Toyokuni, S. Malignant mesothelioma as an oxidative stress-induced cancer: An update. Free Radic. Biol. Med. 2015, 86, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Scherpereel, A.; Wallyn, F.; Albelda, S.M.; Munck, C. Novel therapies for malignant pleural mesothelioma. Lancet Oncol. 2018, 19, e161–e172. [Google Scholar] [CrossRef]
- Bonelli, M.A.; Fumarola, C.; La Monica, S.; Alfieri, R. New therapeutic strategies in malignant pleural mesothelioma. Biochem. Pharmacol. 2017, 123, 8–18. [Google Scholar] [CrossRef]
- Delage, B.; Fennell, D.A.; Nicholson, L.; McNeish, L.; Lemoine, N.R.; Crook, T.; Szlosarek, P.W. Arginine deprivation and argininosuccinate synthetase expression in the treatment of cancer. Int. J. Cancer 2010, 126, 2762–2772. [Google Scholar] [CrossRef]
- Ensor, C.M.; Holtsberg, F.W.; Bomalaski, J.S.; Clark, M.A. Pegylated arginine deiminase (ADI-SS PEG20,000mw) inhibits human melanomas and hepatocellular carcinomas in vitro and in vivo. Cancer Res. 2002, 62, 5443–5450. [Google Scholar]
- Allen, M.; Luong, P.; Hudson, C.; Leyton, J.; Delage, B.; Ghazaly, E.; Cutts, R.; Yuan, M.; Syed, N.; Lo Nigro, C.; et al. Prognostic and therapeutic impact of argininosuccinate synthase-1 control in bladder cancer as monitored longituf=dinally by PET imaging. Cancer Res. 2014, 74, 896–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szlosarek, P.W.; Steele, J.P.; Nolan, L.; Gilligan, D.; Taylor, P.; Spicer, J.; Lind, M.; Mitra, S.; Shamash, J.; Phillips, M.M.; et al. Arginine deprivation with pegylated arginine deiminase in patients with argininosuccinate synthetase 1–deficient malignant pleural mesothelioma: A randomized clinical trial. JAMA Oncol. 2017, 3, 58–66. [Google Scholar] [CrossRef]
- Phillips, M.M.; Szyszko, T.; Hall, P.; Cook, G.J.R.; Khadeir, R.; Steele, J.P.C.; Spicer, J.F.; Feng, X.; Hategan, M.; Rashid, S.; et al. Expansion study of ADI-PEG 20, pemetrexed and cisplatin in patients with ASS1-deficient malignant pleural mesothelioma (TRAP). Proc. Am. Soc. Clin. Oncol. 2017, 35 (Suppl. 15), 8553. [Google Scholar] [CrossRef]
- Topalian, S.L.; Drake, C.G.; Pardoll, D.M. Immune checkpoint blockade: A common denominator approach to cancer therapy. Cancer Cell 2015, 27, 450–461. [Google Scholar] [CrossRef] [Green Version]
- Hotta, K.; Fujimoto, N. Current evidence and future perspectives of immune-checkpoint inhibitors in unresectable malignant pleural mesothelioma. J. Immunother Cancer 2020, 8, e000461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, R.; Kindler, H.L.; Jahan, T.; Bazhenova, L.; Reck, M.; Thomas, A.; Pastan, I.; Parno, J.; O’Shannessy, D.J.; Fatato, P.; et al. Phase II clinical trial of amatuximab, a chimeric anti-mesothelin antibody with pemetrexed and cisplatincisplatin in advanced unresectable pleural mesothelioma. Clin. Cancer Res. 2014, 20, 5927–5936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, R.; Alley, E.; Kindler, H.; Antonia, S.; Jahan, T.; Honarmand, S.; Nair, N.; Whiting, C.C.; Enstrom, A.; Lemmens, E.; et al. Clinical Response of Live-Attenuated, Listeria monocytogenes Expressing Mesothelin (CRS-207) with Chemotherapy in Patients with Malignant Pleural Mesothelioma. Clin. Cancer Res. 2019, 25, 5787–5798. [Google Scholar] [CrossRef]
- Reid, G.; Johnson, T.G.; van Zandwijk, N. Manipulating microRNAs for the Treatment of Malignant Pleural Mesothelioma: Past, Present and Future. Front. Oncol. 2020, 10, 105. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Bukowiecka-Matusiak, M.; Turek, I.A.; Woźniak, L.A. Natural Phenolic Antioxidants and Their Synthetic Derivatives. In Systems Biology of FreeRadicals and Antioxidants; Laher, I., Ed.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 4047–4061. [Google Scholar]
- Fantini, M.; Benvenuto, M.; Masuelli, L.; Frajese, G.V.; Tresoldi, I.; Modesti, A.; Bei, R. In Vitro and in vivo antitumoral effects of combinations of polyphenols, or polyphenols and anticancer drugs: Perspectives on cancer treatment. Int. J. Mol. Sci. 2015, 16, 9236–9282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piasek, A.; Bartoszek, A.; Namieśnik, J. Phytochemicals that counteract the cardiotoxic side effects of cancer chemotherapy. Postepy Hig. Med. Dosw. 2009, 63, 142–158. [Google Scholar]
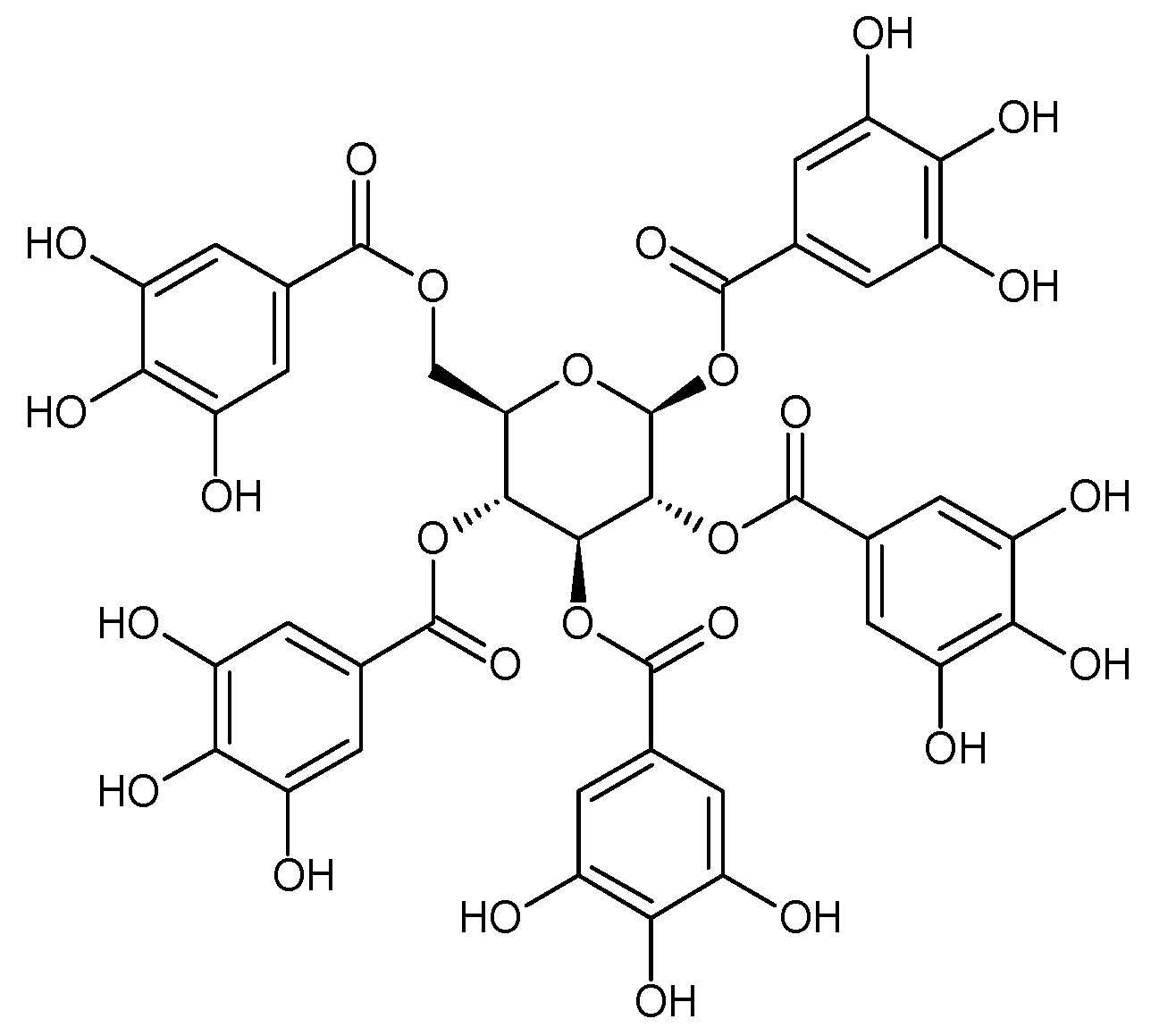
- Zhang, J.; Li, L.; Kim, S.H.; Hagerman, A.E.; Lü, J. Anti-cancer, anti-diabetic and other pharmacologic and biological activities of penta-galloyl-glucose. Pharm. Res. 2009, 26, 2066–2080. [Google Scholar] [CrossRef] [Green Version]
- Oh, G.S.; Pae, H.O.; Oh, H.; Hong, S.G.; Kim, I.K.; Chai, K.Y.; Yun, Y.G.; Kwon, T.O.; Chung, H.T. In Vitro antiproliferative effect of 1,2,3,4,6-penta-O-galloyl-beta-D-glucose on the human hepatocellular carcinoma cell line, SK-HEP-1 cells. Cancer Lett. 2001, 174, 17–24. [Google Scholar] [CrossRef]
- Kitagawa, S.; Nabekura, T.; Nakamura, Y.; Takahashi, T.; Kashiwada, Y. Inhibition of P-glycoprotein function by tannic acid and pentagalloylglucose. J. Pharm. Pharmacol. 2007, 59, 965–969. [Google Scholar] [CrossRef]
- Ryu, H.G.; Jeong, S.J.; Kwon, H.Y.; Lee, H.J.; Lee, E.O.; Lee, M.H.; Choi, S.H.; Ahn, K.S.; Kim, S.H. Penta-O-galloyl-β-D-glucose attenuates cisplatin-induced nephrotoxicity via reactive oxygen species reduction in renal epithelial cells and enhances antitumor activity in Caki-2 renal cancer cells. Toxicol. In Vitro 2012, 26, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Ceci, C.; Lacal, P.M.; Tentori, L.; de Martino, M.G.; Miano, R.; Graziani, G. Experimental Evidence of the Antitumor, Antimetastatic and Antiangiogenic Activity of Ellagic Acid. Nutrients 2018, 10, 1756. [Google Scholar] [CrossRef] [Green Version]
- Zhou, D.H.; Wang, X.; Yang, M.; Shi, X.; Huang, W.; Feng, Q. Combination of low concentration of (-)-epigallocatechin gallate (EGCG) and curcumin strongly suppresses the growth of non-small cell lung cancer in vitro and in vivo through causing cell cycle arrest. Int. J. Mol. Sci. 2013, 14, 12023–12036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somers-Edgar, T.J.; Scandlyn, M.J.; Stuart, E.C.; Le Nedelec, M.J.; Valentine, S.P.; Rosengren, R.J. The combination of epigallocatechin gallate and curcumin suppresses ER alpha-breast cancer cell growth in vitro and in vivo. Int. J. Cancer 2008, 122, 1966–1971. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wang, B.; Chung, S.; Wu, Y.; Henning, S.M.; Vadgama, J.V. Increased chemopreventive effect by combining arctigenin, green tea polyphenol and curcumin in prostate and breast cancer cells. RSC Adv. 2014, 4, 35242–35250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mertens-Talcott, S.U.; Percival, S.S. Ellagic acid and quercetin interact synergistically with resveratrol in the induction of apoptosis and cause transient cell cycle arrest in human leukemia cells. Cancer Lett. 2005, 218, 141–151. [Google Scholar] [CrossRef] [PubMed]
- León-Gonzáles, A.J.; Auger, C.; Schini-Kerth, V.B. Pro-oxidant activity of polyphenols and its implication on cancer chemoprevention and chemotherapy. Biochem. Pharmacol. 2015, 98, 371–380. [Google Scholar] [CrossRef]
- Wojcik, M.; Krawczyk, M.; Wojcik, P.; Cypryk, K.; Wozniak, L.A. Molecular Mechanisms Underlying Curcumin-Mediated Therapeutic Effects in Type 2 Diabetes and Cancer. Oxidative Med. Cell. Longev. 2018, 2018, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Baldi, A.; De Luca, A.; Maiorano, P.; D’Angelo, C.; Giordano, A. Curcumin as an anticancer agent in malignant mesothelioma: A review. Int. J. Mol. Sci. 2020, 21, 1839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Rishi, A.K.; Wu, W.; Polin, L.; Sharma, S.; Levi, E.; Albelda, S.; Pass, H.I.; Wali, A. Curcumin suppresses the growth of mesothelioma cells in vitro and in vivo, in part, by stimulating apoptosis. Mol. Cell Biochem. 2011, 357, 83–94. [Google Scholar] [CrossRef] [Green Version]
- Ghiringhelli, F.; Rebe, C.; Hichami, A.; Delmas, D. Immunomodulation and anti-inflammatory roles of polyphenols as anticancer agents. Anticancer Agents Med. Chem. 2012, 12, 852–873. [Google Scholar] [CrossRef]
- Miller, J.M.; Thompson, J.K.; MacPherson, M.B.; Beuschel, S.L.; Westbom, C.M.; Sayan, M.; Shukla, A. Curcumin: A double hit on malignant mesothelioma. Cancer Prev. Res. 2014, 7, 330–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuelli, L.; Benvenuto, M.; di Stefano, E.; Mattera, R.; Fantini, M.; de Feudis, G.; de Smaele, E.; Tresoldi, I.; Giganti, M.G.; Modesti, A.; et al. Curcumin blocks autophagy and activates apoptosis of malignant mesothelioma cell lines and increases the survival of mice intraperitoneally transplanted with a malignant mesothelioma cell line. Oncotarget 2017, 8, 34405–34422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Meo, F.; Filosa, S.; Madonna, M.; Giello, G.; Di Pardo, A.; Maglione, V.; Baldi, A.; Crispi, S. Curcumin C3 complex/Bioperine has antineoplastic activity in mesothelioma: An in vitro and in vivo analysis. J. Exp. Clin. Cancer Res. 2019, 38, 360–371. [Google Scholar] [CrossRef] [PubMed]
- Pouliquen, D.L.; Boissard, A.; Henry, C.; Blandin, S.; Richomme, P.; Coqueret, O.; Guette, C. Curcumin Treatment Identifies Therapeutic Targets within Biomarkers of Liver Colonization by Highly Invasive Mesothelioma Cells-Potential Links with Sarcomas. Cancers 2020, 12, 3384. [Google Scholar] [CrossRef]
- Pouliquen, D.L.; Nawrocki-Raby, B.; Nader, J.; Blandin, S.; Robard, M.; Birembaut, P.; Grégoire, M. Evaluation of intracavitary administration of curcumin for the treatment of sarcomatoid mesothelioma. Oncotarget 2017, 8, 57552–57573. [Google Scholar] [CrossRef] [Green Version]
- Hocking, A.; Tommasi, S.; Sordillo, P.; Klebe, S. The safety and exploration of the pharmacokinetics of intrapleural liposomal curcumin. Int. J. Nanomed. 2020, 15, 943–952. [Google Scholar] [CrossRef]
- Valenti, D.; de Bari, L.; Manente, G.A.; Rossi, L.; Mutti, L.; Moro, L.; Vacca, R.A. Negative modulation of mitochondrial oxidative phosphorylation by epigallocatechin-3 gallate leads to growth arrest and apoptosis in human malignant pleural mesothelioma cells. Biochim. Biophys. Acta 2013, 1832, 2085–2096. [Google Scholar] [CrossRef]
- Ranzato, E.; Martinotti, S.; Magnelli, V.; Murer, B.; Biffo, S.; Mutti, L.; Burlando, B. Epigallocatechin-3-gallate induces mesothelioma cell death via H2O2 -dependent T-type Ca2+ channel opening. J. Cell Mol. Med. 2012, 16, 2667–2678. [Google Scholar] [CrossRef]
- Satoh, M.; Takemura, Y.; Hamada, H.; Sekido, Y.; Kubota, S. EGCG induces human mesothelioma cell death by inducing reactive oxygen species and autophagy. Cancer Cell Int. 2013, 13, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Martinotti, S.; Ranzato, E.; Burlando, B. In Vitro screening of synergistic ascorbate-drug combinations for the treatment of malignant mesothelioma. Toxicol. In Vitro 2011, 25, 1568–1574. [Google Scholar] [CrossRef]
- Martinotti, S.; Ranzato, E.; Parodi, M.; Vitale, M.; Burlando, B. Combination of ascorbate/epigallocatechin-3-gallate/gemcitabine synergistically induces cell cycle deregulation and apoptosis in mesothelioma cells. Toxicol. Appl. Pharmacol. 2014, 274, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Martinotti, S.; Ranzato, E.; Burlando, B. (−)- Epigallocatechin-3-gallate induces GRP78 accumulation in the ER and shifts mesothelioma constitutive UPR into pro-apoptotic ER stress. J. Cell. Physiol. 2018, 233, 7082–7090. [Google Scholar] [CrossRef]
- Lee, Y.J.; Im, J.H.; Lee, D.M.; Park, J.S.; Won, S.Y.; Cho, M.K.; Nam, H.S.; Lee, Y.J.; Lee, S.H. Synergistic inhibition of mesothelioma cell growth by the combination of clofarabine and resveratrol involves Nrf2 downregulation. BMB Rep. 2012, 45, 647–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.J.; Lee, Y.J.; Im, J.H.; Won, S.Y.; Kim, Y.B.; Cho, M.K.; Nam, H.S.; Choi, Y.J.; Lee, S.H. Synergistic anticancer effects of resveratrol and chemotherapeutic agent clofarabine against human malignant mesothelioma MSTO-211H cells. Food Chem. Toxicol. 2013, 52, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Lee, G.J.; Yi, S.S.; Heo, S.H.; Park, C.R.; Nam, H.S.; Cho, M.K.; Lee, S.H. Cisplatin and resveratrol induce apoptosis and autophagy following oxidative stress in malignant mesothelioma cells. Food Chem. Toxicol. 2016, 97, 96–107. [Google Scholar] [CrossRef]
- Motohashi, H.; Yamamoto, M. Nrf2-Keap1 defines a physiologically important stress response mechanism. Trends Mol. Med. 2004, 10, 549–557. [Google Scholar] [CrossRef]
- Beishline, K.; Azizkhan-Clifford, J. Sp1 and the ‘hallmarks of cancer’. FEBS J. 2015, 282, 224–258. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.A.; Lee, Y.J.; Ban, J.O.; Lee, Y.J.; Lee, S.H.; Cho, M.K.; Nam, H.S.; Hong, J.T.; Shim, J.H. The flavonoid resveratrol suppresses growth of human malignant pleural mesothelioma cells through direct inhibition of specificity protein 1. Int. J. Mol. Med. 2012, 30, 21–27. [Google Scholar]
- Lee, Y.J.; Lee, Y.J.; Lee, S.H. Resveratrol and clofarabine induces a preferential apoptosis-activating effect on malignant mesothelioma cells by Mcl-1 down-regulation and caspase-3 activation. BMB Rep. 2015, 48, 166–171. [Google Scholar] [CrossRef]
- Chae, J.I.; Cho, J.H.; Lee, K.A.; Choi, N.J.; Seo, K.S.; Kim, S.B.; Lee, S.H.; Shim, J.H. Role of transcription factor Sp1 in the quercetin-mediated inhibitory effect on human malignant pleural mesothelioma. Int. J. Mol. Med. 2012, 30, 835–841. [Google Scholar] [CrossRef] [Green Version]
- Demiroglu-Zergeroglu, A.; Basara-Cigerim, B.; Kilic, E.; Yanikkaya-Demirel, G. The investigation of effects of quercetin and its combination with cisplatinCisplatin on malignant mesothelioma cells in vitro. J. Biomed. Biotechnol. 2010, 2010, 851589. [Google Scholar] [CrossRef]
- Demiroglu-Zergeroglu, A.; Ergene, E.; Ayvali, N.; Kuete, V.; Sivas, H. Quercetin and Cisplatin combined treatment altered cell cycle and mitogen-activated protein kinase expressions in malignant mesothelioma cells. BMC Complement Altern. Med. 2016, 16, 281. [Google Scholar] [CrossRef] [Green Version]
- Demiroglu-Zergeroglu, A.; Candemir, G.; Turhanlar, E.; Sagir, F.; Ayvali, N. EGFR-dependent signalling reduced and p38 dependent apoptosis required by Gallic acid in Malignant Mesothelioma cells. Biomed. Pharmacother. 2016, 84, 2000–2007. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.A.; Chae, J.I.; Shim, J.H. Natural diterpenes from coffee, cafestol and kahweol induce apoptosis through regulation of specificity protein 1 expression in human malignant pleural mesothelioma. J. Biomed. Sci. 2012, 19, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huber, W.W.; Rossmanith, W.; Grusch, M.; Haslinger, E.; Prustomersky, S.; Peter-Vörösmarty, B.; Parzefall, W.; Scharf, G.; Schulte-Hermann, R. Effects of coffee and its chemopreventive components kahweol and cafestol on cytochrome P450 and sulfotransferase in rat liver. Food Chem. Toxicol. 2008, 46, 1230–1238. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.A.; Lee, S.H.; Lee, Y.J.; Baeg, S.M.; Shim, J.H. Hesperidin induces apoptosis by inhibiting Sp1 and its regulatory protein in MSTO-211H cells. Biomol. Ther. 2012, 20, 273–279. [Google Scholar] [CrossRef] [Green Version]
- Chae, J.I.; Jeon, Y.J.; Shim, J.H. Downregulation of Sp1 is involved in honokiol-induced cell cycle arrest and apoptosis in human malignant pleural mesothelioma cells. Oncol. Rep. 2013, 29, 2318–2324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolbe, L.; Immeyer, J.; Batzer, J.; Wensorra, U.; tom Dieck, K.; Mundt, C.; Wolber, R.; Stäb, F.; Schönrock, U.; Ceilley, R.I.; et al. Anti-inflammatory efficacy of Licochalcone A: Correlation of clinical potency and in vitro effects. Arch. Dermatol. Res. 2006, 298, 23–30. [Google Scholar] [CrossRef]
- Kim, K.H.; Yoon, G.; Cho, J.J.; Cho, J.H.; Cho, Y.S.; Chae, J.I.; Shim, J.H. Licochalcone A induces apoptosis in malignant pleural mesothelioma through downregulation of Sp1 and subsequent activation of mitochondria-related apoptotic pathway. Int. J. Oncol. 2015, 46, 1385–1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chirumamilla, C.; Pérez-Novo, C.; Van Ostade, X.; Vanden Berghe, W. Molecular insights into cancer therapeutic effects of the dietary medicinal phytochemical withaferin A. Proc. Nutr. Soc. 2017, 76, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Wang, Y.; Cheryan, V.T.; Wu, W.; Cui, C.Q.; Polin, L.A.; Pass, H.I.; Dou, Q.P.; Rishi, A.K.; Wali, A. Withaferin A inhibits the proteasome activity in mesothelioma in vitro and in vivo. PLoS ONE 2012, 7, e41214. [Google Scholar] [CrossRef] [PubMed]
- Sayeed, M.A.; Bracci, M.; Lucarini, G.; Lazzarini, R.; Di Primio, R.; Santarelli, L. Regulation of microRNA using promising dietary phytochemicals: Possible preventive and treatment option of malignant mesothelioma. Biomed. Pharmacother. 2017, 94, 1197–1224. [Google Scholar] [CrossRef] [PubMed]
- Sohn, E.J.; Won, G.; Lee, J.; Yoon, S.S.; Lee, I.; Kim, H.J.; Kim, S.H. Blockage of epithelial to mesenchymal transition and upregulation of let 7b are critically involved in ursolic acid induced apoptosis in malignant mesothelioma cell. Int. J. Biol. Sci. 2016, 12, 1279–1288. [Google Scholar] [CrossRef] [Green Version]
- Roomi, M.W.; Ivanov, V.; Kalinovsky, T.; Niedzwiecki, A.; Rath, M. Inhibition of malignant mesothelioma cell matrix metalloproteinase production and invasion by a novel nutrient mixture. Exp. Lung Res. 2006, 32, 69–79. [Google Scholar] [CrossRef]
- Roomi, M.W.; Ivanov, V.; Netke, S.P.; Niedzwiecki, A.; Rath, M. Serum markers of the liver, heart, and kidney and lipid profile and histopathology in ODS rats treated with nutrient synergy. J. Am. Coll. Nutr. 2003, 22, 477. [Google Scholar]
- Pulito, C.; Mori, F.; Sacconi, A.; Casadei, L.; Ferraiuolo, M.; Valerio, M.C.; Santoro, R.; Geoman, F.; Maidecchi, A.; Mattoli, L.; et al. Cynara scolymus affects malignant pleural mesothelioma by promoting apoptosis and restraining invasion. Oncotarget 2015, 6, 18134–18150. [Google Scholar] [CrossRef] [Green Version]
- Chmielewska-Kassassir, M.; Sobierajska, K.; Ciszewski, W.M.; Bukowiecka-Matusiak, M.; Szczesna, D.; Burzynska-Pedziwiatr, I.; Wiczkowski, W.; Wagner, W.; Wozniak, L.A. Polyphenol Extract from Evening Primrose (Oenothera paradoxa) Inhibits Invasion Properties of Human Malignant Pleural Mesothelioma Cells. Biomolecules 2020, 10, 1574. [Google Scholar] [CrossRef]
- Kiss, A.K.; Derwińska, M.; Dawidowska, A.; Naruszewicz, M. Novel biological properties of Oenothera paradoxa defatted seed extracts: Effect on metallopeptidase activity. J. Agric. Food Chem. 2008, 56, 7845–7852. [Google Scholar] [CrossRef]
- Birch, A.E.; Fenner, G.P.; Watkins, R.; Boyd, L.C. Antioxidant properties of evening primrose seed extracts. J. Agric. Food Chem. 2001, 49, 4502–4507. [Google Scholar] [CrossRef]
- Wettasinghe, M.; Shahidi, F. Evening primrose meal: A source of natural antioxidants and scavenger of hydrogen peroxide and oxygen-derived free radicals. J. Agric. Food Chem. 1999, 47, 1801–1812. [Google Scholar] [CrossRef] [PubMed]
- Wettasinghe, M.; Shahidi, F. Iron (II) chelation activity of extracts of borage and evening primrose meals. Food Res. Int. 2002, 35, 65–71. [Google Scholar] [CrossRef]
- Kiss, A.; Filipek, A.; Czerwińska, M.; Naruszewicz, M. Oenothera paradoxa defatted seeds extract, and its bioactive component penta-O-gallolyl-β-D-glucose decreased production of reactive oxygen species and inhibited release of leukotriene B4, interleukin-8, elastase and myeloperoxidase, in human neutrophils. J. Agric. Food Chem. 2010, 58, 9960–9966. [Google Scholar]
- Burzynska-Pedziwiatr, I.; Bukowiecka-Matusiak, M.; Wojcik, M.; Machala, W.; Bienkiewicz, M.; Spolnik, G.; Danikiewicz, W.; Wozniak, L.A. Dual stimulus-dependent effect of Oenothera paradoxa extract on the respiratory burst in human leukocytes: Suppressing for Escherichia coli and phorbol myristate acetate and stimulating for formyl-methionyl-leucyl-phenylalanine. Oxid. Med. Cell Longev. 2014, 2014, 764367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellegrina, C.D.; Padovani, G.; Mainente, F.; Zoccatelli, G.; Bissoli, G.; Mosconi, S.; Veneri, G.; Peruffo, A.; Andrighetto, G.; Rizzi, C.; et al. Anti-tumor potential of a gallic acid-containing phenolic fraction from Oenothera biennis. Cancer Lett. 2005, 226, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Kiss, A.K.; Kaplon-Cieslicka, A.; Filipiak, K.J.; Opolski, G.; Naruszewicz, M. Ex Vivo effects of an Oenothera paradoxa extract on the reactive oxygen species generation and neutral endopeptidase activity in neutrophils from patients after acute myocardial infarction. Phytother. Res. 2011, 26, 482–487. [Google Scholar] [CrossRef] [PubMed]
- Granica, S.; Czerwińska, M.E.; Piwowarski, J.P.; Ziaja, M.; Kiss, A.K. Chemical composition, antioxidative and anti-inflammatory activity of extracts prepared from aerial parts of Oenothera biennis L. and Oenothera paradoxa hudziok obtained after seeds cultivation. J. Agric. Food Chem. 2013, 61, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Sałaga, M.; Lewandowska, U.; Sosnowska, D.; Zakrzewski, P.K.; Cygankiewicz, A.I.; Piechota-Polańczyk, A.; Sobczak, M.; Mosinska, P.; Chen, C.; Krajewska, W.M.; et al. Polyphenol extract from evening primrose pomace alleviates experimental colitis after intracolonic and oral administration in mice. Naunyn Schmiedeberg’s Arch. Pharmacol. 2014, 387, 1069–1078. [Google Scholar] [CrossRef] [Green Version]
- Jaszewska, E.; Kośmider, A.; Kiss, A.K.; Naruszewicz, M. Pro-oxidative and pro-apoptotic action of defatted seeds of Oenothera paradoxa on human skin melanoma cells. J. Agric. Food Chem. 2009, 57, 8282–8289. [Google Scholar] [CrossRef]
- Gorlach, S.; Wagner, W.; Podsedek, A.; Sosnowska, D.; Dastych, J.; Koziołkiewicz, M. Polyphenols from evening primrose (Oenothera paradoxa) defatted seeds induce apoptosis in human colon cancer Caco-2 cells. J. Agric. Food Chem. 2011, 59, 6985–6997. [Google Scholar] [CrossRef] [PubMed]
- Arimura, T.; Kojima-Yuasa, A.; Tatsumi, Y.; Kennedy, D.O.; Matsui-Yuasa, I. Involvement of polyamines in evening primrose extract-induced apoptosis in Ehrlich ascites tumor cells. Amino Acids 2005, 28, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Lewandowska, U.; Owczarek, K.; Szewczyk, K.; Podsędek, A.; Koziołkiewicz, M.; Hrabec, M. Influence of polyphenol extract from evening primrose (Oenothera paradoxa) seeds on human prostate and breast cancer cell lines. Postepy Hig. Med. Dosw. 2014, 68, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Jaszewska, E.; Kosmider, A.; Kiss, A.K.; Naruszewicz, M. Oenothera paradoxa defatted seeds extract containing pentagalloylglucose and procyanidins potentiates the cytotoxicity of vincristine. J. Physiol. Pharm. 2010, 61, 637–643. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Flavonoids | Flavonols | Flavones | Flavanols | ||
quercetin | luteolin | catechin epigallocatechin-3-gallate (EGCG) | |||
| Anthocyanidins | Isoflavones | Flavanones | |||
cyanidin | genistein | hesperidin | |||
| Phenolic acids | gallic acid | caffeic acid | protocatechuic acid | ||
| Stilbenes | resveratrol | Lignans | pinoresinol | Other polyphenols | curcumin |
| Compound | Molecular Target Effect | Effect | Reference |
|---|---|---|---|
| Curcumin | ↑ Bax proteins and caspase-9 expression; Induction of XAF1, CARP1/CCAR1, SULF1 | Stimulation of pro-apoptotic factors; induction of novel apoptosis signaling transducers; | [41] [44] |
| reduction of tumorigenic properties of MPM cells in an animal model | [43] | ||
| Epigallocatechin-3-gallate (+ascorbate and gemcitabine) | ↑ ROS generation | Impairment of mitochondrial membrane potential; | [51] |
| ↓ NF-κB activation (↓ p65 subunit) | restoring programmed cell death; | [53] | |
| pro-apoptotic ER stress (UPR shift and GRP78 accumulation in ER) | induction of pro-apoptotic signals | [54] | |
| Resveratrol (+cisplatin or +clofarabine) | ↑ROS generation in mitochondria ↓ Sp1 protein level ↓ expression of Sp1 regulatory proteins: p21, p27, cyclin D1, Mcl1, and survivin ↑ caspase-3 activation↓ anti-apoptotic Bcl-xL ↑ pro-apoptotic Bax | Activation of the apoptotic pathway; MSTO-H211 cells sensitization to the antitumor effect of clofarabine and cisplatin | [55,56,57] [60,61] |
Quercetin (+ cisplatin) | ↓ Sp1 protein and mRNA level ↑ caspase-3 signaling ↓ anti-apoptotic Bcl-xL ↑ pro-apoptotic Bax | S-phase arrest; activation of the apoptotic pathway; sensitization to the antitumor effect of cisplatin | [62,63,64] |
Gallic acid | ↓ expression of cyclin D and Bcl-2 genes; altered expression of p21 | G1 phase arrest; mitochondria dysfunction; apoptosis | [65] |
Cafestol  Kahweol  | ↓ Sp1 protein level ↓ expression of Sp1 regulatory proteins: cyclin D1, Mcl1, and survivin ↑ caspase-3 signaling ↓ anti-apoptotic Bcl-xL ↑ pro-apoptotic Bax, Bid | Growth arrest of MSTO-H211 cellsat Sub-G1 phase; activation of apoptotic signals | [66] |
Hesperidin  | ↓ Sp1 mRNA and protein level ↓ expression of Sp1 regulatory proteins: cyclin D1, Mcl1, and survivin ↑ caspase-3 signaling ↓ anti-apoptotic Bcl-xL ↑ pro-apoptotic Bax, Bid | Growth arrest of MSTO-H211 cellsat Sub-G1 phase; activation of apoptotic signals | [68] |
Honokiol  | ↓ Sp1 mRNA and protein level ↓ expression of Sp1 regulatory proteins: cyclin D1, Mcl1, and survivin ↑ caspase-3 signaling ↓ anti-apoptotic Bcl-xL ↑ pro-apoptotic Bax, Bid | Proliferation inhibition; growth arrest of MSTO-H211 cellsat Sub-G1 phase; activation of apoptotic signals | [69] |
Licochalcone A  | ↓ Sp1 mRNA and protein level ↓ expression of Sp1 regulatory proteins: cyclin D1, Mcl1, and survivin ↑ caspase-3 signaling ↓ anti-apoptotic Bcl-xL ↑ pro-apoptotic Bax, Bid | Growth arrest of MSTO-H211 and NCI-H28 cells at Sub-G1 phase; depolarization of mitochondria membrane; activation of apoptotic signals | [71] |
Withaferin A | ↑ p38 stress-activated caspase-3, ↑ Bax protein, PARP cleavage, induction of CARP1/CCAR1 | Increase of pro-apoptotic proteins; blockage in the chymotryptic activity of proteasome | [73] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chmielewska-Kassassir, M.; Wozniak, L.A. Phytochemicals in Malignant Pleural Mesothelioma Treatment—Review on the Current Trends of Therapies. Int. J. Mol. Sci. 2021, 22, 8279. https://doi.org/10.3390/ijms22158279
Chmielewska-Kassassir M, Wozniak LA. Phytochemicals in Malignant Pleural Mesothelioma Treatment—Review on the Current Trends of Therapies. International Journal of Molecular Sciences. 2021; 22(15):8279. https://doi.org/10.3390/ijms22158279
Chicago/Turabian StyleChmielewska-Kassassir, Malgorzata, and Lucyna A. Wozniak. 2021. "Phytochemicals in Malignant Pleural Mesothelioma Treatment—Review on the Current Trends of Therapies" International Journal of Molecular Sciences 22, no. 15: 8279. https://doi.org/10.3390/ijms22158279
APA StyleChmielewska-Kassassir, M., & Wozniak, L. A. (2021). Phytochemicals in Malignant Pleural Mesothelioma Treatment—Review on the Current Trends of Therapies. International Journal of Molecular Sciences, 22(15), 8279. https://doi.org/10.3390/ijms22158279