
Neural and Hormonal Basis of Opposite-Sex Preference by Chemosensory Signals

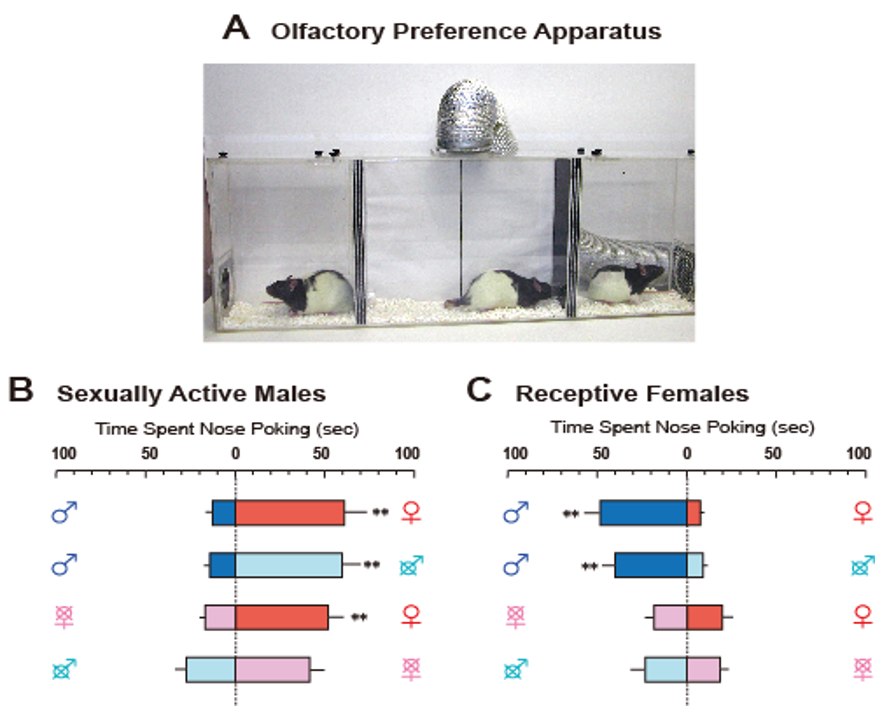{kind=link}
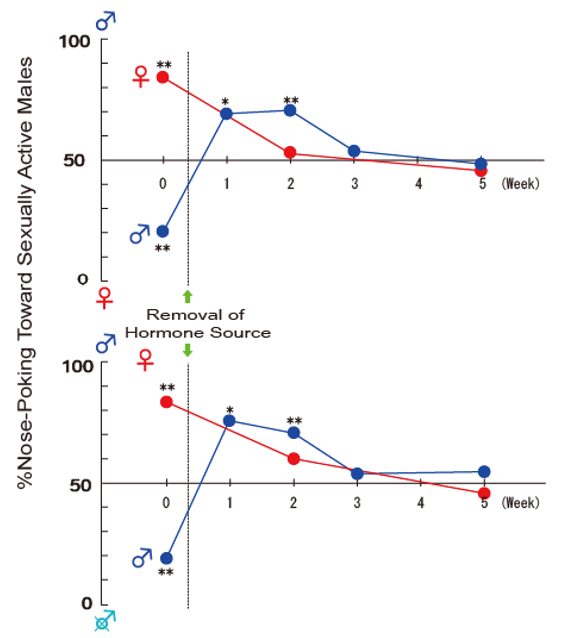{kind=link}
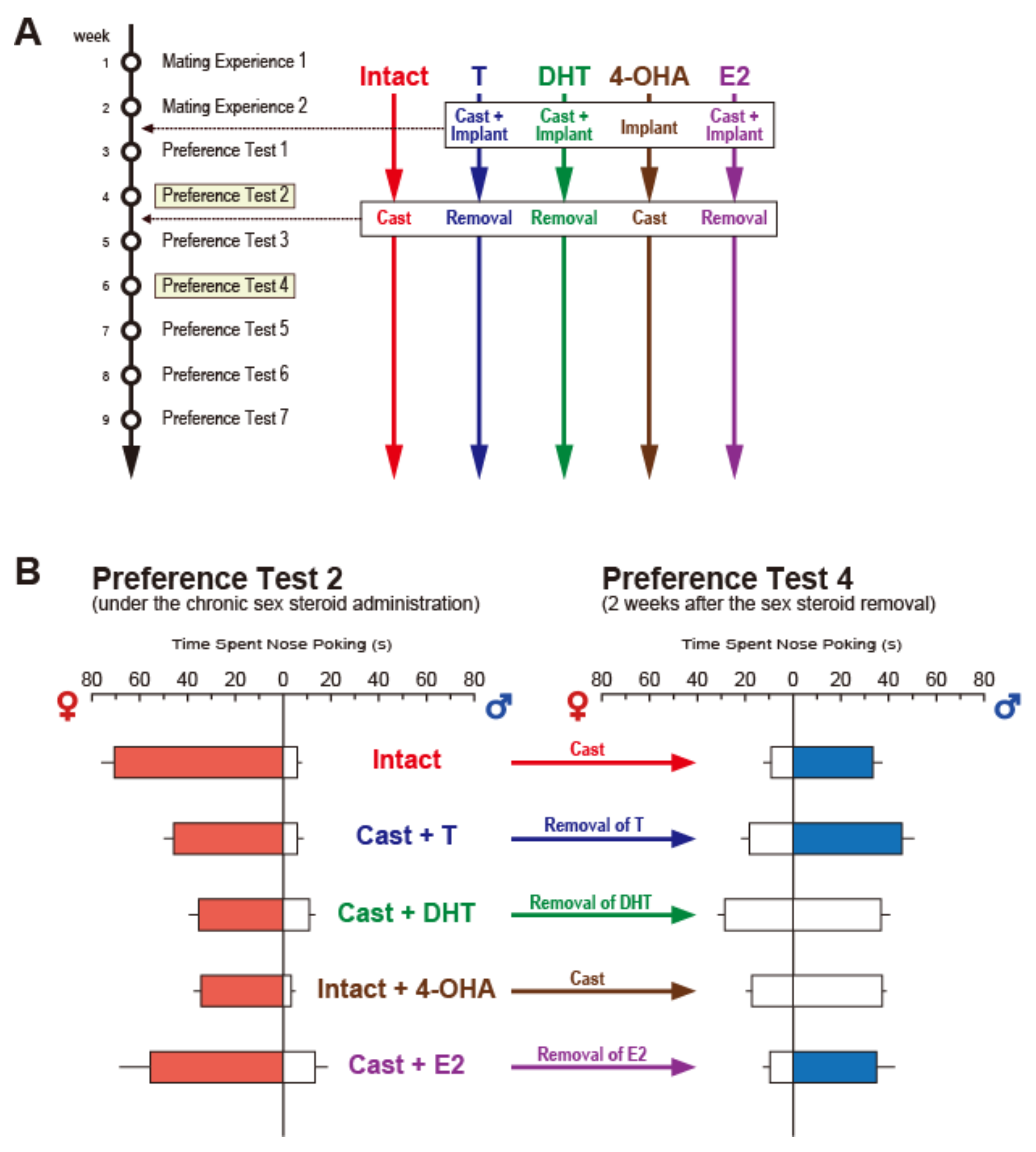{kind=link}
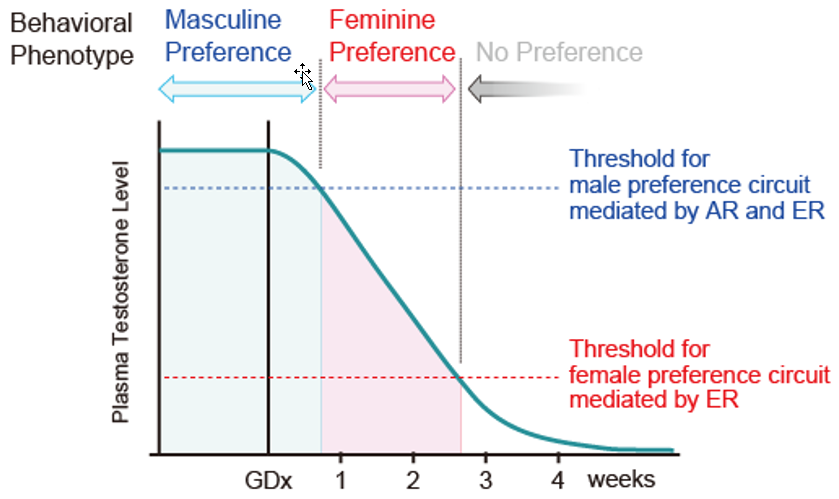{kind=link}
{kind=link}
Abstract
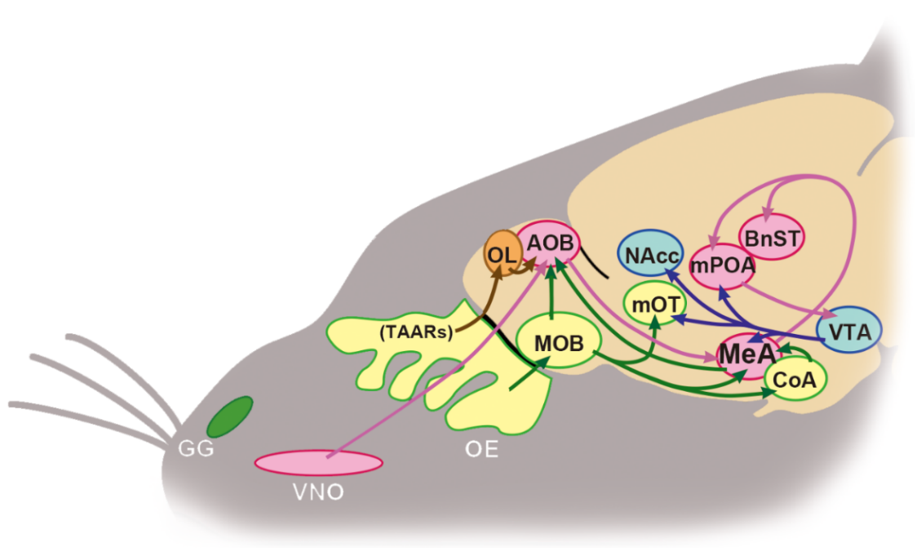
:1. Introduction
2. Detection of Sexual Chemicals
3. Sexual Stimuli Attributes
4. Sexually Dimorphic Preference and Organizational Effects of Sex Steroids
5. Hormonal Control for Opposite-Sex Preference
6. Regulation of Opposite-Sex Preference by Integrating the Main and Accessory Olfactory Systems
7. Experience and Sexual Preference
8. Neuropeptidergic Regulation of Opposite-Sex Preference
9. Future Outlook
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Slob, A.; DeKlerk, L.; Brand, T. Homosexual and heterosexual partner preference in ovariectomized female rats: Effects of testosterone, estradiol and mating experience. Physiol. Behav. 1987, 41, 571–576. [Google Scholar] [CrossRef]
- Broekman, M.; De Bruin, M.; Smeenk, J.; Slob, A.K.; Van Der Schoot, P. Partner preference behavior of estrous female rats affected by castration of tethered male incentives. Horm. Behav. 1988, 22, 324–337. [Google Scholar] [CrossRef]
- Merkx, J.; Slob, A.; Bosch, J.V.D.W.T. Preference for an estrous female over a non-estrous female evinced by female rats requires dihydrotestosterone plus estradiol. Horm. Behav. 1989, 23, 466–472. [Google Scholar] [CrossRef]
- Brand, T.; Kroonen, J.; Mos, J.; Slob, A. Adult partner preference and sexual behavior of male rats affected by perinatal endocrine manipulations. Horm. Behav. 1991, 25, 323–341. [Google Scholar] [CrossRef] [Green Version]
- Bakker, J.; Van Ophemert, J.; Eijskoot, F.; Slob, A. A semiautomated test apparatus for studying partner preference behavior in the rat. Physiol. Behav. 1994, 56, 597–601. [Google Scholar] [CrossRef] [Green Version]
- Xiao, K.; Kondo, Y.; Sakuma, Y. Sex-specific effects of gonadal steroids on conspecific odor preference in the rat. Horm. Behav. 2004, 46, 356–361. [Google Scholar] [CrossRef]
- Brechbühl, J.; Klaey, M.; Broillet, M.-C. Grueneberg Ganglion Cells Mediate Alarm Pheromone Detection in Mice. Science 2008, 321, 1092–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brechbühl, J.; Moine, F.; Klaey, M.; Nenniger-Tosato, M.; Hurni, N.; Sporkert, F.; Giroud, C.; Broillet, M.-C. Mouse alarm pheromone shares structural similarity with predator scents. Proc. Natl. Acad. Sci. USA 2013, 110, 4762–4767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brechbuhl, J.; Klaey, M.; Moine, F.; Bovay, E.; Hurni, N.; Nenniger-Tosato, M.; Broillet, M.C. Morphological and physiological species-dependent characteristics of the rodent Grueneberg ganglion. Front. Neuroanat. 2014, 8, 87. [Google Scholar]
- Stowers, L.; Kuo, T.-H. Mammalian pheromones: Emerging properties and mechanisms of detection. Curr. Opin. Neurobiol. 2015, 34, 103–109. [Google Scholar] [CrossRef] [Green Version]
- Halem, H.A.; Baum, M.J.; Cherry, J.A. Sex Difference and Steroid Modulation of Pheromone-Induced Immediate Early Genes in the Two Zones of the Mouse Accessory Olfactory System. J. Neurosci. 2001, 21, 2474–2480. [Google Scholar] [CrossRef] [Green Version]
- Haga-Yamanaka, S.; Ma, L.; He, J.; Qiu, Q.; Lavis, L.D.; Looger, L.L.; Yu, C.R. Integrated action of pheromone signals in promoting courtship behavior in male mice. eLife 2014, 3, e03025. [Google Scholar] [CrossRef] [PubMed]
- Kaur, A.W.; Ackels, T.; Kuo, T.-H.; Cichy, A.; Dey, S.; Hays, C.; Kateri, M.; Logan, D.; Marton, T.F.; Spehr, M.; et al. Murine Pheromone Proteins Constitute a Context-Dependent Combinatorial Code Governing Multiple Social Behaviors. Cell 2014, 157, 676–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chamero, P.; Marton, T.F.; Logan, D.; Flanagan, K.; Cruz, J.R.; Saghatelian, A.; Cravatt, B.F.; Stowers, L. Identification of protein pheromones that promote aggressive behaviour. Nature 2007, 450, 899–902. [Google Scholar] [CrossRef]
- Roberts, S.A.; Simpson, D.M.; Armstrong, S.D.; Davidson, A.J.; Robertson, D.H.; McLean, L.; Beynon, R.J.; Hurst, J.L. Darcin: A male pheromone that stimulates female memory and sexual attraction to an individual male’s odour. BMC Biol. 2010, 8, 75. [Google Scholar] [CrossRef] [Green Version]
- Demir, E.; Li, K.; Bobrowski-Khoury, N.; Sanders, J.I.; Beynon, R.J.; Hurst, J.L.; Kepecs, A.; Axel, R. The pheromone darcin drives a circuit for innate and reinforced behaviours. Nature 2020, 578, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Haga, S.; Hattori, T.; Sato, T.; Sato, K.; Matsuda, S.; Kobayakawa, R.; Sakano, H.; Yoshihara, Y.; Kikusui, T.; Touhara, K. The male mouse pheromone ESP1 enhances female sexual receptive behaviour through a specific vomeronasal receptor. Nature 2010, 466, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Stopka, P.; Kuntová, B.; Klempt, P.; Havrdová, L.; Černá, M.; Stopková, R. On the saliva proteome of the Eastern European house mouse (Mus musculus musculus) focusing on sexual signalling and immunity. Sci. Rep. 2016, 6, 32481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niimura, Y. Olfactory Receptor Multigene Family in Vertebrates: From the Viewpoint of Evolutionary Genomics. Curr. Genom. 2012, 13, 103–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liberles, S.D. Trace Amine-associated Receptors Are Olfactory Receptors in Vertebrates. Ann. N. Y. Acad. Sci. 2009, 1170, 168–172. [Google Scholar] [CrossRef]
- Li, Q.; Liberles, S.D. Aversion and Attraction through Olfaction. Curr. Biol. 2015, 25, R120–R129. [Google Scholar] [CrossRef] [Green Version]
- Liberles, S.D. Trace amine-associated receptors: Ligands, neural circuits, and behaviors. Curr. Opin. Neurobiol. 2015, 34, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harmeier, A.; Meyer, C.A.; Staempfli, A.; Casagrande, F.; Petrinovic, M.M.; Zhang, Y.-P.; Künnecke, B.; Iglesias, A.; Höner, O.P.; Hoener, M.C. How Female Mice Attract Males: A Urinary Volatile Amine Activates a Trace Amine-Associated Receptor That Induces Male Sexual Interest. Front. Pharmacol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Korzan, W.J.; Ferrero, D.M.; Chang, R.B.; Roy, D.S.; Buchi, M.; Lemon, J.K.; Kaur, A.W.; Stowers, L.; Fendt, M.; et al. Synchronous Evolution of an Odor Biosynthesis Pathway and Behavioral Response. Curr. Biol. 2013, 23, 11–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacifico, R.; Dewan, A.; Cawley, D.; Guo, C.; Bozza, T. An Olfactory Subsystem that Mediates High-Sensitivity Detection of Volatile Amines. Cell Rep. 2012, 2, 76–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, M.A.; Tsai, L.; Roy, D.S.; Valenzuela, D.H.; Mosley, C.; Magklara, A.; Lomvardas, S.; Liberles, S.D.; Barnea, G. Neurons expressing trace amine-associated receptors project to discrete glomeruli and constitute an olfactory subsystem. Proc. Natl. Acad. Sci. USA 2012, 109, 13410–13415. [Google Scholar] [CrossRef] [Green Version]
- Kobayakawa, K.; Kobayakawa, R.; Matsumoto, H.; Oka, Y.; Imai, T.; Ikawa, M.; Okabe, M.; Ikeda, T.; Itohara, S.; Kikusui, T.; et al. Innate versus learned odour processing in the mouse olfactory bulb. Nature 2007, 450, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Omura, M.; Mombaerts, P. Trpc2-Expressing Sensory Neurons in the Main Olfactory Epithelium of the Mouse. Cell Rep. 2014, 8, 583–595. [Google Scholar] [CrossRef] [Green Version]
- Orsulak, P.J.; Gawienowski, A.M. Olfactory Preferences for the Rat Preputial Gland1. Biol. Reprod. 1972, 6, 219–223. [Google Scholar] [CrossRef] [Green Version]
- Bronson, F.H.; Caroom, D. Preputial gland of the male mouse: Attractant function. Reproduction 1971, 25, 279–282. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.X.; Sun, L.; Zhang, J.H.; Feng, Z.Y. Sex- and gonad-affecting scent compounds and 3 male pheromones in the rat. Chem. Senses 2008, 33, 611–621. [Google Scholar] [CrossRef] [Green Version]
- Logan, D.W.; Marton, T.F.; Stowers, L. Species Specificity in Major Urinary Proteins by Parallel Evolution. PLoS ONE 2008, 3, e3280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, S.A.; Prescott, M.C.; Davidson, A.J.; McLean, L.; Beynon, R.J.; Hurst, J.L. Individual odour signatures that mice learn are shaped by involatile major urinary proteins (MUPs). BMC Biol. 2018, 16, 48. [Google Scholar] [CrossRef] [PubMed]
- Mucignat-Caretta, C.; Caretta, A.; Cavaggioni, A. Acceleration of puberty onset in female mice by male urinary proteins. J. Physiol. 1995, 486, 517–522. [Google Scholar] [CrossRef] [Green Version]
- Marchlewska-Koj, A.; Cavaggioni, A.; Mucignat-Caretta, C.; Olejniczak, P. Stimulation of Estrus in Female Mice by Male Urinary Proteins. J. Chem. Ecol. 2000, 26, 2355–2366. [Google Scholar] [CrossRef]
- Pankevich, D.E.; Cherry, J.A.; Baum, M.J. Effect of vomeronasal organ removal from male mice on their preference for and neural Fos responses to female urinary odors. Behav. Neurosci. 2006, 120, 925–936. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Ricós, J.; Agustín-Pavón, C.; Lanuza, E.; Martínez-García, F. Intraspecific Communication Through Chemical Signals in Female Mice: Reinforcing Properties of Involatile Male Sexual Pheromones. Chem. Senses 2006, 32, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Ganfornina, M.D.; Gutiérrez, G.; Bastiani, M. A Phylogenetic Analysis of the Lipocalin Protein Family. Mol. Biol. Evol. 2000, 17, 114–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timm, D.E.; Baker, L.; Mueller, H.; Zidek, L.; Novotny, M.V. Structural basis of pheromone binding to mouse major urinary protein (MUP-I). Protein Sci. 2001, 10, 997–1004. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, S.D.; Robertson, D.H.L.; Cheetham, S.A.; Hurst, J.L.; Beynon, R.J. Structural and functional differences in isoforms of mouse major urinary proteins: A male-specific protein that preferentially binds a male pheromone. Biochem. J. 2005, 391, 343–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jemiolo, B.; Alberts, J.; Sochinski-Wiggins, S.; Harvey, S.; Novotny, M. Behavioural and endocrine responses of female mice to synthetic analogues of volatile compounds in male urine. Anim. Behav. 1985, 33, 1114–1118. [Google Scholar] [CrossRef]
- Asaba, A.; Hattori, T.; Mogi, K.; Kikusui, T. Sexual attractiveness of male chemicals and vocalizations in mice. Front. Neurosci. 2014, 8, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korzan, W.J.; Freamat, M.; Johnson, A.G.; Cherry, J.A.; Baum, M.J. Either main or accessory olfactory system signaling can mediate the rewarding effects of estrous female chemosignals in sexually naive male mice. Behav. Neurosci. 2013, 127, 755–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Jiang, Y.; Si, Y.; Kim, J.-Y.; Chen, Z.-F.; Rao, Y. Molecular regulation of sexual preference revealed by genetic studies of 5-HT in the brains of male mice. Nature 2011, 472, 95–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, M.; Pierman, S.; Douhard, Q.; Baum, M.J.; Bakker, J. The vomeronasal organ is required for the expression of lordosis behaviour, but not sex discrimination in female mice. Eur. J. Neurosci. 2006, 23, 521–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bodo, C.; Rissman, E.F. The Androgen Receptor Is Selectively Involved in Organization of Sexually Dimorphic Social Behaviors in Mice. Endocrinology 2008, 149, 4142–4150. [Google Scholar] [CrossRef] [PubMed]
- Meerts, S.H.; Clark, A.S. Stimulus animal characteristics do not modulate the expression of partner preference by female rats. Physiol. Behav. 2006, 89, 623–626. [Google Scholar] [CrossRef]
- DiBenedictis, B.T.; Ingraham, K.L.; Baum, M.J.; Cherry, J.A. Disruption of urinary odor preference and lordosis behavior in female mice given lesions of the medial amygdala. Physiol. Behav. 2012, 105, 554–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DiBenedictis, B.T.; Olugbemi, A.O.; Baum, M.J.; Cherry, J.A. DREADD-Induced Silencing of the Medial Olfactory Tubercle Disrupts the Preference of Female Mice for Opposite-Sex Chemosignals. Eneuro 2015, 2. [Google Scholar] [CrossRef]
- Bakker, J.; Honda, S.-I.; Harada, N.; Balthazart, J. The Aromatase Knock-Out Mouse Provides New Evidence That Estradiol Is Required during Development in the Female for the Expression of Sociosexual Behaviors in Adulthood. J. Neurosci. 2002, 22, 9104–9112. [Google Scholar] [CrossRef]
- Gubits, R.M.; Lynch, K.R.; Kulkarni, A.B.; Dolan, K.P.; Gresik, E.W.; Hollander, P.; Feigelson, P. Differential regulation of alpha 2u globulin gene expression in liver, lachrymal gland, and salivary gland. J. Biol. Chem. 1984, 259, 12803–12809. [Google Scholar] [CrossRef]
- Shahan, K.; Denaro, M.; Gilmartin, M.; Shi, Y.; Derman, E. Expression of six mouse major urinary protein genes in the mammary, parotid, sublingual, submaxillary, and lachrymal glands and in the liver. Mol. Cell. Biol. 1987, 7, 1947–1954. [Google Scholar] [CrossRef] [PubMed]
- Dhungel, S.; Rai, D.; Terada, M.; Orikasa, C.; Nishimori, K.; Sakuma, Y.; Kondo, Y. Oxytocin is indispensable for conspecific-odor preference and controls the initiation of female, but not male, sexual behavior in mice. Neurosci. Res. 2019, 148, 34–41. [Google Scholar] [CrossRef]
- Achiraman, S.; Ponmanickam, P.; Ganesh, D.S.; Archunan, G. Detection of estrus by male mice: Synergistic role of olfactory–vomeronasal system. Neurosci. Lett. 2010, 477, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, M.C.; Nagata, K.; Sagoshi, S.; Ogawa, S. Estrogen and oxytocin involvement in social preference in male mice: A study using a novel long-term social preference paradigm with aromatase, estrogen receptor-α and estrogen receptor-β, oxytocin, and oxytocin receptor knockout male mice. Integr. Zoöl. 2018, 13, 698–710. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, H.; Kumagai, R.; Kondo, Y. Why does castrated male odor attract sexually active male rats?–Attractivity induced by hypothalamus-pituitary-gonad axis block. Physiol. Behav. 2021, 230, 113288. [Google Scholar] [CrossRef]
- Xiao, K.; Chiba, A.; Sakuma, Y.; Kondo, Y. Transient reversal of olfactory preference following castration in male rats: Implication for estrogen receptor involvement. Physiol. Behav. 2015, 152, 161–167. [Google Scholar] [CrossRef]
- Luo, M.; Fee, M.S.; Katz, L.C. Encoding Pheromonal Signals in the Accessory Olfactory Bulb of Behaving Mice. Science 2003, 299, 1196–1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammen, G.F.; Turaga, D.; Holy, T.E.; Meeks, J.P. Functional organization of glomerular maps in the mouse accessory olfactory bulb. Nat. Neurosci. 2014, 17, 953–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakker, J.; Honda, S.; Harada, N.; Balthazart, J. Restoration of male sexual behavior by adult exogenous estrogens in male aromatase knockout mice. Horm. Behav. 2004, 46, 1–10. [Google Scholar] [CrossRef]
- Vega Matuszczyk, J.; Larsson, K. Role of androgen, estrogen and sexual experience on the female rat’s partner preference. Physiol Behav 1991, 50, 139–142. [Google Scholar] [CrossRef]
- Brand, T.; Slob, A. Neonatal organization of adult partner preference behavior in male rats. Physiol. Behav. 1991, 49, 107–111. [Google Scholar] [CrossRef]
- Matuszczyk, J.V.; Larsson, K. Sexual Preference and Feminine and Masculine Sexual Behavior of Male Rats Prenatally Exposed to Antiandrogen or Antiestrogen. Horm. Behav. 1995, 29, 191–206. [Google Scholar] [CrossRef]
- Bakker, J.; Brand, T.; van Ophemert, J.; Slob, A.K. Hormonal regulation of adult partner preference behavior in neonatally ATD-treated male rats. Behav. Neurosci. 1993, 107, 480–487. [Google Scholar] [CrossRef]
- Houtsmuller, E.; Brand, T.; de Jonge, F.; Joosten, R.; van de Poll, N.; Slob, A. SDN-POA volume, sexual behavior, and partner preference of male rats affected by perinatal treatment with ATD. Physiol. Behav. 1994, 56, 535–541. [Google Scholar] [CrossRef] [Green Version]
- Cruz, C.D.; Pereira, O.C.M. Prenatal testosterone supplementation alters puberty onset, aggressive behavior, and partner preference in adult male rats. J. Physiol. Sci. 2012, 62, 123–131. [Google Scholar] [CrossRef]
- Henley, C.; Nunez, A.; Clemens, L. Exogenous androgen during development alters adult partner preference and mating behavior in gonadally intact male rats. Horm. Behav. 2010, 57, 488–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henley, C.; Nunez, A.; Clemens, L. Estrogen treatment during development alters adult partner preference and reproductive behavior in female laboratory rats. Horm. Behav. 2009, 55, 68–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Jonge, F.H.; Muntjewerff, J.W.; Louwerse, A.L.; Van De Poll, N.E. Sexual behavior and sexual orientation of the female rat after hormonal treatment during various stages of development. Horm. Behav. 1988, 22, 100–115. [Google Scholar] [CrossRef]
- Wersinger, S.R.; Rissman, E.F. Oestrogen receptor alpha is essential for female-directed chemo-investigatory behaviour but is not required for the pheromone-induced luteinizing hormone surge in male mice. J. Neuroendocr. 2000, 12, 103–110. [Google Scholar] [CrossRef]
- Bodo, C.; Rissman, E.F. New roles for estrogen receptor beta in behavior and neuroendocrinology. Front. Neuroendocrinol. 2006, 27, 217–232. [Google Scholar] [CrossRef]
- Bakker, J.; Honda, S.; Harada, N.; Balthazart, J. Sexual Partner Preference Requires a Functional Aromatase (Cyp19) Gene in Male Mice. Horm. Behav. 2002, 42, 158–171. [Google Scholar] [CrossRef] [PubMed]
- Bodo, C.; Rissman, E.F. Androgen receptor is essential for sexual differentiation of responses to olfactory cues in mice. Eur. J. Neurosci. 2007, 25, 2182–2190. [Google Scholar] [CrossRef] [PubMed]
- Raskin, K.; De Gendt, K.; Duittoz, A.; Liere, P.; Verhoeven, G.; Tronche, F.; Mhaouty-Kodja, S. Conditional Inactivation of Androgen Receptor Gene in the Nervous System: Effects on Male Behavioral and Neuroendocrine Responses. J. Neurosci. 2009, 29, 4461–4470. [Google Scholar] [CrossRef]
- Brock, O.; Douhard, Q.; Baum, M.J.; Bakker, J. Reduced Prepubertal Expression of Progesterone Receptor in the Hypothalamus of Female Aromatase Knockout Mice. Endocrinology 2010, 151, 1814–1821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brock, O.; Baum, M.J.; Bakker, J. The Development of Female Sexual Behavior Requires Prepubertal Estradiol. J. Neurosci. 2011, 31, 5574–5578. [Google Scholar] [CrossRef]
- Mazzucco, C.A.; Walker, H.A.; Pawluski, J.; Lieblich, S.E.; Galea, L.A. ERα, but not ERβ, mediates the expression of sexual behavior in the female rat. Behav. Brain Res. 2008, 191, 111–117. [Google Scholar] [CrossRef]
- Ogawa, S.; Lubahn, D.B.; Korach, K.; Pfaff, D.W. Behavioral effects of estrogen receptor gene disruption in male mice. Proc. Natl. Acad. Sci. USA 1997, 94, 1476–1481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imwalle, D.B.; Scordalakes, E.M.; Rissman, E.F. Estrogen Receptor α Influences Socially Motivated Behaviors. Horm. Behav. 2002, 42, 484–491. [Google Scholar] [CrossRef]
- Honda, S.; Harada, N.; Ito, S.; Takagi, Y.; Maeda, S. Disruption of sexual behavior in male aromatase-deficient mice lacking exons 1 and 2 of the cyp19 gene. Biochem. Biophys. Res. Commun. 1998, 252, 445–449. [Google Scholar] [CrossRef]
- Kondo, Y.; Tomihara, K.; Sakuma, Y. Sensory requirements for noncontact penile erection in the rat. Behav. Neurosci. 1999, 113, 1062–1070. [Google Scholar] [CrossRef]
- Bialy, M.; Nikolaev-Diak, A.; Kalata, U.; Nikolaev, E. Blockade of androgen receptor in the medial amygdala inhibits noncontact erections in male rats. Physiol. Behav. 2011, 103, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Bialy, M.; Sachs, B.D. Androgen Implants in Medial Amygdala Briefly Maintain Noncontact Erection in Castrated Male Rats. Horm. Behav. 2002, 42, 345–355. [Google Scholar] [CrossRef]
- Manzo, J.; Cruz, M.; Hernandez, M.E.; Pacheco, P.; Sachs, B. Regulation of Noncontact Erection in Rats by Gonadal Steroids. Horm. Behav. 1999, 35, 264–270. [Google Scholar] [CrossRef]
- Seo, S.I.; Kim, S.W.; Paick, J.S. The effects of androgen on penile reflex, erectile response to electrical stimulation and penile NOS activity in the rat. Asian J. Androl. 1999, 1, 169–174. [Google Scholar]
- Muroi, Y.; Ishii, T.; Komori, S.; Nishimura, M. A competitive effect of androgen signaling on male mouse attraction to volatile female mouse odors. Physiol. Behav. 2006, 87, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Schellino, R.; Trova, S.; Cimino, I.; Farinetti, A.; Jongbloets, B.C.; Pasterkamp, R.J.; Panzica, G.; Giacobini, P.; De Marchis, S.; Peretto, P. Opposite-sex attraction in male mice requires testosterone-dependent regulation of adult olfactory bulb neurogenesis. Sci. Rep. 2016, 6, 36063. [Google Scholar] [CrossRef]
- Eliasson, M.; Meyerson, B.J. Sexual preference in female rats during estrous cycle, pregnancy and lactation. Physiol. Behav. 1975, 14, 705–710. [Google Scholar] [CrossRef]
- Clark, A.S.; Kelton, M.C.; Guarraci, F.A.; Clyons, E.Q. Hormonal status and test condition, but not sexual experience, modulate partner preference in female rats. Horm. Behav. 2004, 45, 314–323. [Google Scholar] [CrossRef]
- Monchobogani, J.; Lanuza, E.; Lorente, M.; Martinez-Garcia, F. Attraction to male pheromones and sexual behaviour show different regulatory mechanisms in female mice. Physiol. Behav. 2004, 81, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Lukas, M.; Neumann, I.D. Social preference and maternal defeat-induced social avoidance in virgin female rats: Sex differences in involvement of brain oxytocin and vasopressin. J. Neurosci. Methods 2014, 234, 101–107. [Google Scholar] [CrossRef]
- Yao, S.; Bergan, J.; Lanjuin, A.; Dulac, C. Oxytocin signaling in the medial amygdala is required for sex discrimination of social cues. eLife 2017, 6, e31373. [Google Scholar] [CrossRef] [PubMed]
- Eidson, L.N.; Maras, P.M.; Epperson, E.; Petrulis, A. Female hamster preference for odors is not regulated by circulating gonadal hormones. Physiol. Behav. 2007, 91, 134–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chamero, P.; Leinders-Zufall, T.; Zufall, F. From genes to social communication: Molecular sensing by the vomeronasal organ. Trends Neurosci. 2012, 35, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Wysocki, C.J.; Lepri, J.J. Consequences of removing the vomeronasal organ. J. Steroid Biochem. Mol. Biol. 1991, 39, 661–669. [Google Scholar] [CrossRef]
- Powers, J.; Winans, S. Vomeronasal organ: Critical role in mediating sexual behavior of the male hamster. Science 1975, 187, 961–963. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.R. Copulatory Behavior of Male Rats Paired with Natural Proestrous and Hormone-treated Ovariectomized Females. Exp. Anim. 1987, 36, 91–93. [Google Scholar] [CrossRef] [Green Version]
- Kondo, Y.; Sudo, T.; Tomihara, K.; Sakuma, Y. Activation of accessory olfactory bulb neurons during copulatory behavior after deprivation of vomeronasal inputs in male rats. Brain Res. 2003, 962, 232–236. [Google Scholar] [CrossRef]
- Dhungel, S.; Masaoka, M.; Rai, D.; Kondo, Y.; Sakuma, Y. Both olfactory epithelial and vomeronasal inputs are essential for activation of the medial amygdala and preoptic neurons of male rats. Neuroscience 2011, 199, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Pankevich, D.E.; Baum, M.J.; Cherry, J.A. Olfactory Sex Discrimination Persists, Whereas the Preference for Urinary Odorants from Estrous Females Disappears in Male Mice after Vomeronasal Organ Removal. J. Neurosci. 2004, 24, 9451–9457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cross, S.K.J.; Martin, Y.H.; Salia, S.; Gamba, I.; Major, C.A.; Hassan, S.; Parsons, K.A.; Swift-Gallant, A. Puberty is a Critical Period for Vomeronasal Organ Mediation of Socio-sexual Behavior in Mice. Front. Behav. Neurosci. 2021, 14. [Google Scholar] [CrossRef]
- Stowers, L.; Holy, T.E.; Meister, M.; Dulac, C.; Koentges, G. Loss of Sex Discrimination and Male-Male Aggression in Mice Deficient for TRP2. Science 2002, 295, 1493–1500. [Google Scholar] [CrossRef] [PubMed]
- Beny, Y.; Kimchi, T. Conditioned odor aversion induces social anxiety towards females in wild-type and TrpC2 knockout male mice. Genes Brain Behav. 2016, 15, 722–732. [Google Scholar] [CrossRef]
- Beny-Shefer, Y.; Zilkha, N.; Lavi-Avnon, Y.; Bezalel, N.; Rogachev, I.; Brandis, A.; Dayan, M.; Kimchi, T. Nucleus Accumbens Dopamine Signaling Regulates Sexual Preference for Females in Male Mice. Cell Rep. 2017, 21, 3079–3088. [Google Scholar] [CrossRef] [Green Version]
- Kimchi, T.; Xu, J.; Dulac, C. A functional circuit underlying male sexual behaviour in the female mouse brain. Nature 2007, 448, 1009–1014. [Google Scholar] [CrossRef]
- Martel, K.L.; Baum, M.J. Adult Testosterone Treatment But Not Surgical Disruption of Vomeronasal Function Augments Male-Typical Sexual Behavior in Female Mice. J. Neurosci. 2009, 29, 7658–7666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, B.A. Trpc2. Handb. Exp. Pharmacol. 2014, 222, 53–65. [Google Scholar]
- Keller, M.; Douhard, Q.; Baum, M.J.; Bakker, J. Sexual Experience Does Not Compensate for the Disruptive Effects of Zinc Sulfate—Lesioning of the Main Olfactory Epithelium on Sexual Behavior in Male Mice. Chem. Senses 2006, 31, 753–762. [Google Scholar] [CrossRef] [Green Version]
- Keller, M.; Douhard, Q.; Baum, M.J.; Bakker, J. Destruction of the Main Olfactory Epithelium Reduces Female Sexual Behavior and Olfactory Investigation in Female Mice. Chem. Senses 2006, 31, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, T.; Hattori, T.; Asaba, A.; Inoue, N.; Kanomata, N.; Kikusui, T.; Kobayakawa, R.; Kobayakawa, K. Genetic dissection of pheromone processing reveals main olfactory system-mediated social behaviors in mice. Proc. Natl. Acad. Sci. USA 2015, 112, E311–E320. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.-M.; Kim, S.-S.; Choi, C.-I.; Cha, H.-L.; Oh, H.-H.; Ghil, S.; Lee, Y.-D.; Birnbaumer, L.; Suh-Kim, H. Development of the main olfactory system and main olfactory epithelium-dependent male mating behavior are altered in Go-deficient mice. Proc. Natl. Acad. Sci. USA 2016, 113, 10974–10979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimomi, Y.; Kondo, Y. Blunt olfaction in sexually sluggish male rats. Exp. Anim. Jpn. Assoc. Lab. Anim. Sci. 2020, 69, 441–447. [Google Scholar] [CrossRef]
- Portillo, W.; Paredes, R.G. Sexual incentive motivation, olfactory preference, and activation of the vomeronasal projection pathway by sexually relevant cues in non-copulating and naive male rats. Horm. Behav. 2004, 46, 330–340. [Google Scholar] [CrossRef]
- McCarthy, E.A.; Kunkhyen, T.; Korzan, W.J.; Naik, A.; Maqsudlu, A.; Cherry, J.A.; Baum, M.J. A comparison of the effects of male pheromone priming and optogenetic inhibition of accessory olfactory bulb forebrain inputs on the sexual behavior of estrous female mice. Horm. Behav. 2017, 89, 104–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakupovic, J.; Kang, N.; Baum, M.J. Effect of bilateral accessory olfactory bulb lesions on volatile urinary odor discrimination and investigation as well as mating behavior in male mice. Physiol. Behav. 2008, 93, 467–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martel, K.L.; Baum, M.J. Sexually dimorphic activation of the accessory, but not the main, olfactory bulb in mice by urinary volatiles. Eur. J. Neurosci. 2007, 26, 463–475. [Google Scholar] [CrossRef] [Green Version]
- Vargas-Barroso, V.; Ordaz-Sanchez, B.; Pena-Ortega, F.; Larriva-Sahd, J.A. Electrophysiological Evidence for a Direct Link between the Main and Accessory Olfactory Bulbs in the Adult Rat. Front. Neurosci. 2015, 9, 518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas-Barroso, V.; Peña-Ortega, F.; Larriva-Sahd, J.A. Olfaction and Pheromones: Uncanonical Sensory Influences and Bulbar Interactions. Front. Neuroanat. 2017, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunkhyen, T.; McCarthy, E.A.; Korzan, W.J.; Doctor, D.; Han, X.; Baum, M.J.; Cherry, J.A. Optogenetic Activation of Accessory Olfactory Bulb Input to the Forebrain Differentially Modulates Investigation of Opposite versus Same-Sex Urinary Chemosignals and Stimulates Mating in Male Mice. Eneuro 2017, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, N.; Baum, M.J.; Cherry, J.A. A direct main olfactory bulb projection to the ‘vomeronasal’ amygdala in female mice selectively responds to volatile pheromones from males. Eur. J. Neurosci. 2009, 29, 624–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martel, K.L.; Baum, M.J. A centrifugal pathway to the mouse accessory olfactory bulb from the medial amygdala conveys gender-specific volatile pheromonal signals. Eur. J. Neurosci. 2009, 29, 368–376. [Google Scholar] [CrossRef] [Green Version]
- Stowers, L.; Marton, T.F. What Is a Pheromone? Mammalian Pheromones Reconsidered. Neuron 2005, 46, 699–702. [Google Scholar] [CrossRef] [Green Version]
- Lin, D.Y.; Zhang, S.-Z.; Block, E.; Katz, L.C. Encoding social signals in the mouse main olfactory bulb. Nature 2005, 434, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Inokuchi, K.; Imamura, F.; Takeuchi, H.; Kim, R.; Okuno, H.; Nishizumi, H.; Bito, H.; Kikusui, T.; Sakano, H. Nrp2 is sufficient to instruct circuit formation of mitral-cells to mediate odour-induced attractive social responses. Nat. Commun. 2017, 8, 15977. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, E.A.; Maqsudlu, A.; Bass, M.; Georghiou, S.; Cherry, J.A.; Baum, M.J. DREADD-induced silencing of the medial amygdala reduces the preference for male pheromones and the expression of lordosis in estrous female mice. Eur. J. Neurosci. 2017, 46, 2035–2046. [Google Scholar] [CrossRef] [PubMed]
- Billing, A.; Correia, M.H.; Kelly, D.A.; Li, G.-L.; Bergan, J.F. Synaptic Connections of Aromatase Circuits in the Medial Amygdala Are Sex Specific. Eneuro 2020, 7. [Google Scholar] [CrossRef] [PubMed]
- Bergan, J.F.; Ben-Shaul, Y.; Dulac, C. Sex-specific processing of social cues in the medial amygdala. eLife 2014, 3, e02743. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.M.; Marcotulli, D.; Shen, A.; Zweifel, L.S. Divergent medial amygdala projections regulate approach–avoidance conflict behavior. Nat. Neurosci. 2019, 22, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Iurilli, G.; Datta, S.R. Population Coding in an Innately Relevant Olfactory Area. Neuron 2017, 93, 1180–1197. [Google Scholar] [CrossRef] [Green Version]
- Root, C.M.; Denny, C.A.; Hen, R.; Axel, R. The participation of cortical amygdala in innate, odour-driven behaviour. Nature 2014, 515, 269–273. [Google Scholar] [CrossRef] [Green Version]
- Pardo-Bellver, C.; Martínez-Bellver, S.; Martínez-García, F.; Lanuza, E.; Teruel-Martí, V. Synchronized Activity in The Main and Accessory Olfactory Bulbs and Vomeronasal Amygdala Elicited by Chemical Signals in Freely Behaving Mice. Sci. Rep. 2017, 7, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Maras, P.; Petrulis, A. The posteromedial cortical amygdala regulates copulatory behavior, but not sexual odor preference, in the male Syrian hamster (Mesocricetus auratus). Neuroscience 2008, 156, 425–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maras, P.M.; Petrulis, A. The anterior medial amygdala transmits sexual odor information to the posterior medial amygdala and related forebrain nuclei. Eur. J. Neurosci. 2010, 32, 469–482. [Google Scholar] [CrossRef] [PubMed]
- Maras, P.M.; Petrulis, A. Lesions that functionally disconnect the anterior and posterodorsal sub-regions of the medial amygdala eliminate opposite-sex odor preference in male Syrian hamsters (Mesocricetus auratus). Neuroscience 2010, 165, 1052–1062. [Google Scholar] [CrossRef] [Green Version]
- Vaz, R.P.; Cardoso, A.; Sá, S.; Pereira, P.; Madeira, M.D. The integrity of the nucleus of the lateral olfactory tract is essential for the normal functioning of the olfactory system. Brain Struct. Funct. 2017, 222, 3615–3637. [Google Scholar] [CrossRef] [Green Version]
- Kondo, Y.; Arai, Y. Functional association between the medial amygdala and the medial preoptic area in regulation of mating behavior in the male rat. Physiol. Behav. 1995, 57, 69–73. [Google Scholar] [CrossRef]
- Kondo, Y.; Yamanouchi, K. The possible involvement of the nonstrial pathway of the amygdala in neural control of sexual behavior in male rats. Brain Res. Bull. 1995, 38, 37–40. [Google Scholar] [CrossRef]
- Been, L.E.; Petrulis, A. Dissociated functional pathways for appetitive and consummatory reproductive behaviors in male Syrian hamsters. Horm. Behav. 2012, 61, 204–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Been, L.E.; Petrulis, A. Lesions of the posterior bed nucleus of the stria terminalis eliminate opposite-sex odor preference and delay copulation in male Syrian hamsters: Role of odor volatility and sexual experience. Eur. J. Neurosci. 2010, 32, 483–493. [Google Scholar] [CrossRef] [Green Version]
- Dhungel, S.; Urakawa, S.; Kondo, Y.; Sakuma, Y. Olfactory preference in the male rat depends on multiple chemosensory inputs converging on the preoptic area. Horm. Behav. 2011, 59, 193–199. [Google Scholar] [CrossRef]
- Hurtazo, H.A.; Paredes, R.G. Olfactory preference and Fos expression in the accessory olfactory system of male rats with bilateral lesions of the medial preoptic area/anterior hypothalamus. Neuroscience 2005, 135, 1035–1044. [Google Scholar] [CrossRef] [PubMed]
- Hurtazo, H.; Paredes, R.; Ågmo, A. Inactivation of the medial preoptic area/anterior hypothalamus by lidocaine reduces male sexual behavior and sexual incentive motivation in male rats. Neuroscience 2008, 152, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Guarraci, F.A.; Clark, A.S. Ibotenic acid lesions of the medial preoptic area disrupt the expression of partner preference in sexually receptive female rats. Brain Res. 2006, 1076, 163–170. [Google Scholar] [CrossRef]
- Martinez, L.A.; Petrulis, A. The medial preoptic area is necessary for sexual odor preference, but not sexual solicitation, in female Syrian hamsters. Horm. Behav. 2013, 63, 606–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McHenry, J.A.; Otis, J.M.; Rossi, M.A.; Robinson, J.E.; Kosyk, O.; Miller, N.W.; McElligott, Z.A.; Budygin, E.A.; Rubinow, D.R.; Stuber, G.D. Hormonal gain control of a medial preoptic area social reward circuit. Nat. Neurosci. 2017, 20, 449–458. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Catalán, M.J.; Orrico, A.; Hipólito, L.; Zornoza, T.; Polache, A.; Lanuza, E.; Martínez-García, F.; Granero, L.; Agustín-Pavón, C. Glutamate and Opioid Antagonists Modulate Dopamine Levels Evoked by Innately Attractive Male Chemosignals in the Nucleus Accumbens of Female Rats. Front. Neuroanat. 2017, 11, 8. [Google Scholar] [CrossRef] [Green Version]
- Wesson, D.W.; Wilson, D. Sniffing out the contributions of the olfactory tubercle to the sense of smell: Hedonics, sensory integration, and more? Neurosci. Biobehav. Rev. 2011, 35, 655–668. [Google Scholar] [CrossRef] [Green Version]
- Pardo-Bellver, C.; Cádiz-Moretti, B.; Novejarque, A.; Martínez-García, F.; Lanuza, E. Differential efferent projections of the anterior, posteroventral, and posterodorsal subdivisions of the medial amygdala in mice. Front. Neuroanat. 2012, 6, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DiBenedictis, B.T.; Helfand, A.I.; Baum, M.J.; Cherry, J.A. A quantitative comparison of the efferent projections of the anterior and posterior subdivisions of the medial amygdala in female mice. Brain Res. 2014, 1543, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Agustín-Pavón, C.; Martinez-Garcia, F.; Lanuza, E. Focal lesions within the ventral striato-pallidum abolish attraction for male chemosignals in female mice. Behav. Brain Res. 2014, 259, 292–296. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Liu, Y.; Rao, Y. Serotonin signaling in the brain of adult female mice is required for sexual preference. Proc. Natl. Acad. Sci. USA 2013, 110, 9968–9973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angoa-Pérez, M.; Herrera-Mundo, N.; Kane, M.J.; Sykes, C.E.; Anneken, J.H.; Francescutti, D.M.; Kuhn, D.M. Brain Serotonin Signaling Does Not Determine Sexual Preference in Male Mice. PLoS ONE 2015, 10, e0118603. [Google Scholar] [CrossRef]
- Sachs, B.D. Erection evoked in male rats by airborne scent from estrous females. Physiol. Behav. 1997, 62, 921–924. [Google Scholar] [CrossRef]
- Lydell, K.; Doty, R.L. Male rat of odor preferences for female urine as a function of sexual experience, urine age, and urine source. Horm. Behav. 1972, 3, 205–212. [Google Scholar] [CrossRef]
- Olvera-Hernández, S.; Hernández, A.; Reyes, R.; Fernández-Guasti, A. Establishment of partner preference in male rats: Effect of prenatal letrozole and sexual experience. Horm. Behav. 2019, 109, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Pankevich, D.E.; Cherry, J.A.; Baum, M.J. Accessory olfactory neural Fos responses to a conditioned environment are blocked in male mice by vomeronasal organ removal. Physiol. Behav. 2006, 87, 781–788. [Google Scholar] [CrossRef] [Green Version]
- Ballard, C.L.; Wood, R.I. Partner preference in male hamsters: Steroids, sexual experience and chemosensory cues. Physiol. Behav. 2007, 91, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maras, P.M.; Petrulis, A. Olfactory experience and the development of odor preference and vaginal marking in female Syrian hamsters. Physiol. Behav. 2008, 94, 545–551. [Google Scholar] [CrossRef] [Green Version]
- Been, L.E.; Petrulis, A. The role of the medial preoptic area in appetitive and consummatory reproductive behaviors depends on sexual experience and odor volatility in male Syrian hamsters. Neuroscience 2010, 170, 1120–1132. [Google Scholar] [CrossRef] [Green Version]
- Van Der Linden, C.; Jakob, S.; Gupta, P.; Dulac, C.; Santoro, S.W. Sex separation induces differences in the olfactory sensory receptor repertoires of male and female mice. Nat. Commun. 2018, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Dillon, T.S.; Fox, L.C.; Han, C.; Linster, C. 17beta-estradiol enhances memory duration in the main olfactory bulb in CD-1 mice. Behav Neurosci 2013, 127, 923–931. [Google Scholar] [CrossRef] [Green Version]
- Shea, S.D.; Katz, L.C.; Mooney, R. Noradrenergic Induction of Odor-Specific Neural Habituation and Olfactory Memories. J. Neurosci. 2008, 28, 10711–10719. [Google Scholar] [CrossRef]
- Unda, N.M.; Eportillo, W.; Corona, R.; Paredes, R.G. Sexual Stimulation Increases the Survival of New Cells in the Accessory Olfactory Bulb of the Male Rat. Front. Neurosci. 2016, 10, 65. [Google Scholar] [CrossRef] [Green Version]
- Swaney, W.T.; Curley, J.P.; Champagne, F.A.; Keverne, E.B. The paternally expressed gene Peg3 regulates sexual experience-dependent preferences for estrous odors. Behav. Neurosci. 2008, 122, 963–973. [Google Scholar] [CrossRef]
- Li, Y.; Mathis, A.; Grewe, B.; Osterhout, J.A.; Ahanonu, B.; Schnitzer, M.J.; Murthy, V.; Dulac, C. Neuronal Representation of Social Information in the Medial Amygdala of Awake Behaving Mice. Cell 2017, 171, 1176–1190. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, J.M.; Brann, J.H.; Gil, M.; Hull, E.M. Sexual experience increases nitric oxide synthase in the medial preoptic area of male rats. Behav. Neurosci. 2006, 120, 1389–1394. [Google Scholar] [CrossRef] [Green Version]
- Swaney, W.; Dubose, B.N.; Curley, J.P.; Champagne, F.A. Sexual experience affects reproductive behavior and preoptic androgen receptors in male mice. Horm. Behav. 2012, 61, 472–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maejima, S.; Abe, Y.; Yamaguchi, S.; Musatov, S.; Ogawa, S.; Kondo, Y.; Tsukahara, S. VGF in the Medial Preoptic Nucleus Increases Sexual Activity Following Sexual Arousal Induction in Male Rats. Endocrinology 2018, 159, 3993–4005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, J.E.; Brameld, J.M.; Jethwa, P.H. Neuroendocrine Role for VGF. Front. Endocrinol. 2015, 6, 3. [Google Scholar] [CrossRef] [Green Version]
- Succu, S.; Mascia, M.S.; Melis, T.; Sanna, F.; Melis, M.R.; Possenti, R.; Argiolas, A. Pro-VGF-derived peptides induce penile erection in male rats: Involvement of paraventricular nitric oxide. Neuropharmacology 2005, 49, 1017–1025. [Google Scholar] [CrossRef] [PubMed]
- Bell, M.R.; Meerts, S.H.; Sisk, C.L. Male Syrian hamsters demonstrate a conditioned place preference for sexual behavior and female chemosensory stimuli. Horm. Behav. 2010, 58, 410–414. [Google Scholar] [CrossRef] [Green Version]
- Gadziola, M.A.; Tylicki, K.A.; Christian, D.L.; Wesson, D.W. The Olfactory Tubercle Encodes Odor Valence in Behaving Mice. J. Neurosci. 2015, 35, 4515–4527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Liu, Q.; Wen, P.; Zhang, J.; Rao, X.; Zhou, Z.; Zhang, H.; He, X.; Li, J.; Zhou, Z.; et al. Activation of the dopaminergic pathway from VTA to the medial olfactory tubercle generates odor-preference and reward. eLife 2017, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, H.H.; Ettenberg, A. Exposure to female rats produces differences in c-fos induction between sexually-naive and experienced male rats. Brain Res. 2002, 947, 57–66. [Google Scholar] [CrossRef]
- Hosokawa, N.; Chiba, A. Effects of sexual experience on conspecific odor preference and estrous odor-induced activation of the vomeronasal projection pathway and the nucleus accumbens in male rats. Brain Res. 2005, 1066, 101–108. [Google Scholar] [CrossRef]
- Fujiwara, M.; Chiba, A. Sexual odor preference and dopamine release in the nucleus accumbens by estrous olfactory cues in sexually naïve and experienced male rats. Physiol. Behav. 2018, 185, 95–102. [Google Scholar] [CrossRef]
- Moncho-Bogani, J.; Martinez-Garcia, F.; Novejarque, A.; Lanuza, E. Attraction to sexual pheromones and associated odorants in female mice involves activation of the reward system and basolateral amygdala. Eur. J. Neurosci. 2005, 21, 2186–2198. [Google Scholar] [CrossRef]
- Moncho-Bogani, J.; Lanuza, E.; Hernandez, A.; Novejarque, A.; Martinez-Garcia, F. Attractive properties of sexual pheromones in mice: Innate or learned? Physiol. Behav. 2002, 77, 167–176. [Google Scholar] [CrossRef]
- Ramm, S.; Cheetham, S.A.; Hurst, J.L. Encoding choosiness: Female attraction requires prior physical contact with individual male scents in mice. Proc. R. Soc. B Biol. Sci. 2008, 275, 1727–1735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarthy, E.A.; Naik, A.S.; Coyne, A.F.; Cherry, J.A.; Baum, M.J. Effect of Ovarian Hormones and Mating Experience on the Preference of Female Mice to Investigate Male Urinary Pheromones. Chem. Senses 2017, 43, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, N.; Chiba, A. Effects of sexual experience on conspecific odor preference and male odor-induced activation of the vomeronasal projection pathway and the nucleus accumbens in female rats. Brain Res. 2007, 1175, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Meerts, S.H.; Park, S.; Sekhawat, R. Sexual experience modulates partner preference and mPOA nitric oxide synthase in female rats. Behav. Neurosci. 2016, 130, 490–499. [Google Scholar] [CrossRef] [PubMed]
- Piergies, A.M.; Hicks, M.E., Jr.; Schwartz, J.P.; Meerts, S.H. Sexually experienced, but not naïve, female rats show a conditioned object preference (COP) for mating after a single training trial. Physiol. Behav. 2019, 198, 42–47. [Google Scholar] [CrossRef]
- Kudwa, A.E.; López, F.J.; McGivern, R.F.; Handa, R.J. A Selective Androgen Receptor Modulator Enhances Male-Directed Sexual Preference, Proceptive Behavior, and Lordosis Behavior in Sexually Experienced, But Not Sexually Naive, Female Rats. Endocrinology 2010, 151, 2659–2668. [Google Scholar] [CrossRef] [PubMed]
- Mitre, M.; Marlin, B.J.; Schiavo, J.K.; Morina, E.; Norden, S.E.; Hackett, T.A.; Aoki, C.J.; Chao, M.V.; Froemke, R.C. A Distributed Network for Social Cognition Enriched for Oxytocin Receptors. J. Neurosci. 2016, 36, 2517–2535. [Google Scholar] [CrossRef] [PubMed]
- Acevedo-Rodriguez, A.; Mani, S.K.; Handa, R.J. Oxytocin and Estrogen Receptor beta in the Brain: An Overview. Front. Endocrinol. 2015, 6, 160. [Google Scholar] [CrossRef] [Green Version]
- Vaccari, C.; Lolait, S.J.; Ostrowski, N.L. Comparative Distribution of Vasopressin V1b and Oxytocin Receptor Messenger Ribonucleic Acids in Brain1. Endocrinology 1998, 139, 5015–5033. [Google Scholar] [CrossRef] [PubMed]
- Wacker, D.; Ludwig, M. The role of vasopressin in olfactory and visual processing. Cell Tissue Res. 2018, 375, 201–215. [Google Scholar] [CrossRef]
- Ferguson, J.N.; Young, L.J.; Hearn, E.F.; Matzuk, M.M.; Insel, T.R.; Winslow, J.T. Social amnesia in mice lacking the oxytocin gene. Nat. Genet. 2000, 25, 284–288. [Google Scholar] [CrossRef]
- Ferguson, J.N.; Aldag, J.M.; Insel, T.R.; Young, L.J. Oxytocin in the Medial Amygdala is Essential for Social Recognition in the Mouse. J. Neurosci. 2001, 21, 8278–8285. [Google Scholar] [CrossRef]
- Oettl, L.-L.; Ravi, N.; Schneider, M.; Scheller, M.F.; Schneider, P.; Mitre, M.; da Silva Gouveia, M.; Froemke, R.; Chao, M.; Young, W.S.; et al. Oxytocin Enhances Social Recognition by Modulating Cortical Control of Early Olfactory Processing. Neuron 2016, 90, 609–621. [Google Scholar] [CrossRef] [Green Version]
- Wersinger, S.R.; Ginns, E.I.; O’Carroll, A.M.; Lolait, S.J.; Young, W.S., 3rd. Vasopressin V1b receptor knockout reduces aggressive behavior in male mice. Mol. Psychiatry 2002, 7, 975–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De la Zerda, S.H.; Netser, S.; Magalnik, H.; Wagner, S. Impaired sex preference, but not social and social novelty preferences, following systemic blockade of oxytocin receptors in adult male mice. Psychoneuroendocrinology 2020, 116, 104676. [Google Scholar] [CrossRef]
- Blitzer, D.; Wells, T.; Hawley, W. Administration of an oxytocin receptor antagonist attenuates sexual motivation in male rats. Horm. Behav. 2017, 94, 33–39. [Google Scholar] [CrossRef]
- Kent, K.; Arientyl, V.; Khachatryan, M.M.; Wood, R.I. Oxytocin Induces a Conditioned Social Preference in Female Mice. J. Neuroendocr. 2013, 25, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Rio, R.T.-D.; Silvaran, M.B.T.; Díaz-Estrada, V.X.; Herrera-Covarrubias, D.; Corona-Morales, A.A.; Pfaus, J.G.; Coria-Avila, G.A. Conditioned same-sex partner preference in male rats is facilitated by oxytocin and dopamine: Effect on sexually dimorphic brain nuclei. Behav. Brain Res. 2015, 283, 69–77. [Google Scholar] [CrossRef]
- Shimizu, K.; Nakamura, K.; Yokosuka, M.; Kondo, Y. Modulation of male mouse sociosexual and anxiety-like behaviors by vasopressin receptors. Physiol. Behav. 2018, 197, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Swaab, D.; Slob, A.; Houtsmuller, E.; Brand, T.; Zhou, J. Increased number of vasopressin neurons in the suprachiasmatic nucleus (SCN) of ‘bisexual’ adult male rats following perinatal treatment with the aromatase blocker ATD. Dev. Brain Res. 1995, 85, 273–279. [Google Scholar] [CrossRef] [Green Version]
- Wersinger, S.R.; Kelliher, K.R.; Zufall, F.; Lolait, S.J.; O’Carroll, A.M.; Young, W.S., 3rd. Social motivation is reduced in vasopressin 1b receptor null mice despite normal performance in an olfactory discrimination task. Horm. Behav. 2004, 46, 638–645. [Google Scholar] [CrossRef] [Green Version]
- DiBenedictis, B.T.; Cheung, H.K.; Nussbaum, E.R.; Veenema, A. Involvement of ventral pallidal vasopressin in the sex-specific regulation of sociosexual motivation in rats. Psychoneuroendocrinology 2020, 111, 104462. [Google Scholar] [CrossRef]
- Bakker, J.; Pierman, S.; Gonzales-Martinez, D. Effects of aromatase mutation (ArKO) on the sexual differentiation of kisspeptin neuronal numbers and their activation by same versus opposite sex urinary pheromones. Horm. Behav. 2010, 57, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Hellier, V.; Brock, O.; Candlish, M.; Desroziers, E.; Aoki, M.; Mayer, C.; Piet, R.; Herbison, A.; Colledge, W.H.; Prevot, V.; et al. Female sexual behavior in mice is controlled by kisspeptin neurons. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Adekunbi, D.; Li, X.F.; Lass, G.; Shetty, K.; Adegoke, O.A.; Yeo, S.H.; Colledge, W.H.; Lightman, S.L.; O’Byrne, K.T. Kisspeptin neurones in the posterodorsal medial amygdala modulate sexual partner preference and anxiety in male mice. J. Neuroendocr. 2018, 30, e12572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gresham, R.; Li, S.; Adekunbi, D.; Hu, M.; Li, X.F.; O’Byrne, K.T. Kisspeptin in the medial amygdala and sexual behavior in male rats. Neurosci. Lett. 2016, 627, 13–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kondo, Y.; Hayashi, H. Neural and Hormonal Basis of Opposite-Sex Preference by Chemosensory Signals. Int. J. Mol. Sci. 2021, 22, 8311. https://doi.org/10.3390/ijms22158311
Kondo Y, Hayashi H. Neural and Hormonal Basis of Opposite-Sex Preference by Chemosensory Signals. International Journal of Molecular Sciences. 2021; 22(15):8311. https://doi.org/10.3390/ijms22158311
Chicago/Turabian StyleKondo, Yasuhiko, and Himeka Hayashi. 2021. "Neural and Hormonal Basis of Opposite-Sex Preference by Chemosensory Signals" International Journal of Molecular Sciences 22, no. 15: 8311. https://doi.org/10.3390/ijms22158311
APA StyleKondo, Y., & Hayashi, H. (2021). Neural and Hormonal Basis of Opposite-Sex Preference by Chemosensory Signals. International Journal of Molecular Sciences, 22(15), 8311. https://doi.org/10.3390/ijms22158311