
The Role of Neuropeptide Y in the Nucleus Accumbens

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
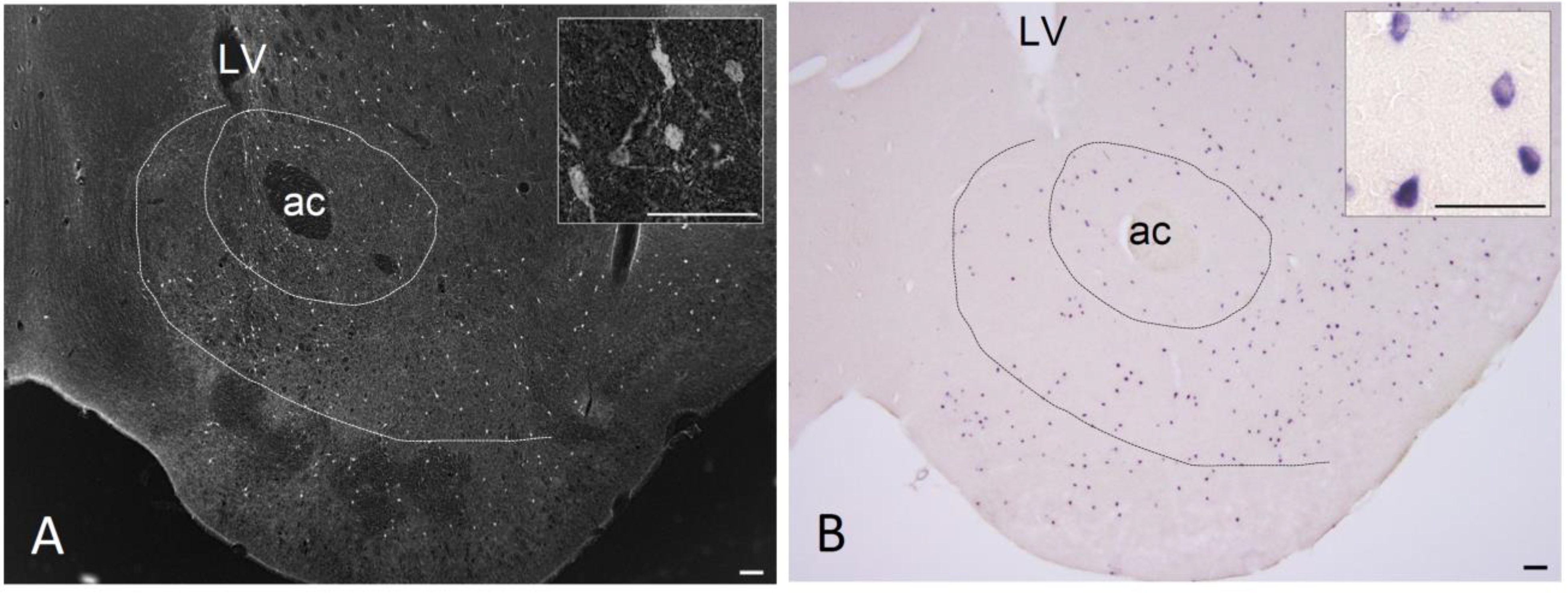
2. Expression of NPY in the NAc
Expression of NPY Is Altered by Drug Treatments
3. NPY and Alcohol Intake in the NAc
4. NPY and Feeding Behavior in the NAc
5. NPY and Reward/Addiction in the NAc
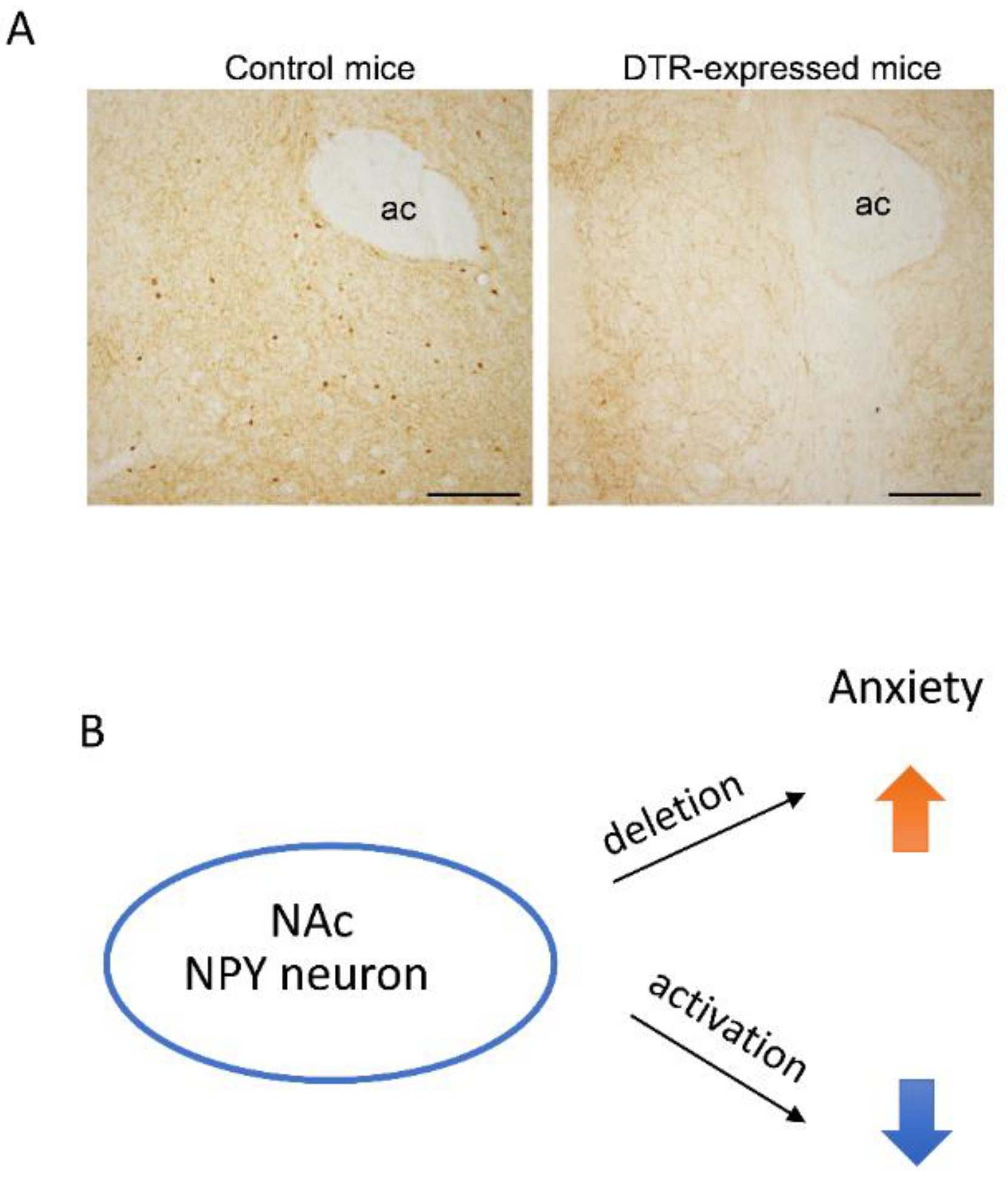
6. NPY and Emotional Behavior in the NAc
7. Other Functions
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Tatemoto, K.; Carlquist, M.; Mutt, V. Neuropeptide Y—A novel brain peptide with structural similarities to peptide YY and pancreatic polypeptide. Nature 1982, 296, 659–660. [Google Scholar] [CrossRef]
- Tatemoto, K.; Siimesmaa, S.; Jörnvall, H.; Allen, J.M.; Polak, J.M.; Bloom, S.R.; Mutt, V. Isolation and characterization of neuropeptide Y from porcine intestine. FEBS Lett. 1985, 179, 181–184. [Google Scholar] [CrossRef] [Green Version]
- Adrian, T.E.; Allen, J.M.; Bloom, S.R.; Ghatei, M.A.; Rossor, M.N.; Roberts, G.W.; Crow, T.J.; Tatemoto, K.; Polak, J.M. Neuropeptide Y distribution in human brain. Nature 1983, 306, 584–586. [Google Scholar] [CrossRef] [PubMed]
- Wahlestedt, C.; Ekman, R.; Widerlöv, E. Neuropeptide Y (NPY) and the central nervous system: Distribution effects and possible relationship to neurological and psychiatric disorders. Prog. Neuropsychopharmacol. Biol. Psychiatry 1989, 13, 31–54. [Google Scholar] [CrossRef]
- Yamazoe, M.; Shiosaka, S.; Emson, P.C.; Tohyama, M. Distribution of neuropeptide Y in the lower brainstem: An immunohistochemical analysis. Brain Res. 1985, 335, 109–120. [Google Scholar] [CrossRef]
- de Quidt, M.E.; Emson, P.C. Distribution of neuropeptide Y-like immunoreactivity in the rat central nervous system--I. Radioimmunoassay and chromatographic characterisation. Neuroscience 1986, 18, 527–543. [Google Scholar] [CrossRef]
- Gray, T.S.; Morley, J.E. Neuropeptide Y: Anatomical distribution and possible function in mammalian nervous system. Life Sci. 1986, 38, 389–401. [Google Scholar] [CrossRef]
- Allen, J.M.; Ferrier, I.N.; Roberts, G.W.; Cross, A.J.; Adrian, T.E.; Crow, T.J.; Bloom, S.R. Elevation of neuropeptide Y (NPY) in substantia innominata in Alzheimer’s type dementia. J. Neurol. Sci. 1984, 64, 325–331. [Google Scholar] [CrossRef]
- Gustafson, E.L.; Card, J.P.; Moore, R.Y. Neuropeptide Y localization in the rat amygdaloid complex. J. Comp. Neurol. 1986, 251, 349–362. [Google Scholar] [CrossRef]
- Chronwall, B.M.; DiMaggio, D.A.; Massari, V.J.; Pickel, V.M.; Ruggiero, D.A.; O’Donohue, T.L. The anatomy of neuropeptide-Y-containing neurons in rat brain. Neuroscience 1985, 15, 1159–1181. [Google Scholar] [CrossRef]
- Lundberg, J.M.; Terenius, L.; Hökfelt, T.; Martling, C.R.; Tatemoto, K.; Mutt, V.; Polak, J.; Bloom, S.; Goldstein, M. Neuropeptide Y (NPY)-like immunoreactivity in peripheral noradrenergic neurons and effects of NPY on sympathetic function. Acta Physiol. Scand. 1982, 116, 477–480. [Google Scholar] [CrossRef]
- Sundler, F.; Håkanson, R.; Ekblad, E.; Uddman, R.; Wahlestedt, C. Neuropeptide Y in the peripheral adrenergic and enteric nervous systems. Int. Rev. Cytol. 1986, 102, 243–269. [Google Scholar]
- Hirsch, D.; Zukowska, Z. NPY and stress 30 years later: The peripheral view. Cell Mol. Neurobiol. 2012, 32, 645–659. [Google Scholar] [CrossRef] [Green Version]
- Benque, I.J.; Xia, P.; Shannon, R.; Ng, T.F.; Taylor, A.W. The Neuropeptides of Ocular Immune Privilege, alpha-MSH and NPY, Suppress Phagosome Maturation in Macrophages. Immunohorizons 2018, 2, 314–323. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.; Shin, M.K.; Kim, E.Y.; Park, J.E.; Lee, H.; Kim, S.W.; Song, J.K.; Chang, E.J. Elevated Neuropeptide Y in Endothelial Dysfunction Promotes Macrophage Infiltration and Smooth Muscle Foam Cell Formation. Front. Immunol. 2019, 10, 1701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bar, F.; Foh, B.; Pagel, R.; Schroder, T.; Schlichting, H.; Hirose, M.; Lemcke, S.; Klinger, A.; Konig, P.; Karsten, C.M. Carboxypeptidase E modulates intestinal immune homeostasis and protects against experimental colitis in mice. PLoS ONE 2014, 9, e102347. [Google Scholar]
- Schwarz, H.; Villiger, P.M.; von Kempis, J.; Lotz, M. Neuropeptide Y is an inducible gene in the human immune system. J. Neuroimmunol. 1994, 51, 53–61. [Google Scholar] [CrossRef]
- Medina, S.; Del Río, M.; Hernanz, A.; De la Fuente, M. The NPY effects on murine leukocyte adherence and chemotaxis change with age. Adherent cell implication. Regul. Pept. 2000, 95, 35–45. [Google Scholar] [CrossRef]
- Chen, W.C.; Liu, Y.B.; Liu, W.F.; Zhou, Y.Y.; He, H.F.; Lin, S. Neuropeptide Y Is an Immunomodulatory Factor: Direct and Indirect. Front. Immunol. 2020, 11, 580378. [Google Scholar] [CrossRef] [PubMed]
- Hörsten, S.; Hoffmann, T.; Alfalah, M.; Wrann, C.D.; Karl, T.; Pabst, R.; Bedoui, C. PP, PYY and NPY: Synthesis, Storage, Release and Degradation. In Neuropeptide Y and RElated Peptides; Michel, M.C., Ed.; Springer: Berlin, Germany, 2004; pp. 23–44. [Google Scholar]
- Wahlestedt, C.; Grundemar, L.; Håkanson, R.; Heilig, M.; Shen, G.H.; Zukowska-Grojec, Z.; Reis, D.J. Neuropeptide Y receptor subtypes, Y1 and Y2. Ann. N. Y. Acad. Sci. 1990, 611, 7–26. [Google Scholar] [CrossRef]
- Larhammar, D.; Blomqvist, A.G.; Yee, F.; Jazin, E.; Yoo, H.; Wahlested, C. Cloning and functional expression of a human neuropeptide Y/peptide YY receptor of the Y1 type. J. Biol. Chem. 1992, 267, 10935–10938. [Google Scholar] [CrossRef]
- Dumont, Y.; Fournier, A.; St-Pierre, S.; Quirion, R. Characterization of neuropeptide Y binding sites in rat brain membrane preparations using [125I] [Leu31,Pro34] peptide YY and [125I] peptide YY3-36 as selective Y1 and Y2 radioligands. J. Pharmacol. Exp. Ther. 1995, 272, 673–680. [Google Scholar] [PubMed]
- Brothers, S.P.; Wahlestedt, C. Therapeutic potential of neuropeptide Y (NPY) receptor ligands. EMBO Mol. Med. 2010, 2, 429–439. [Google Scholar] [CrossRef]
- Reichmann, F.; Holzer, P. Neuropeptide Y: A stressful review. Neuropeptides 2016, 55, 99–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, S.P.; Christopoulos, A.; Davenport, A.P.; Kelly, E.; Marrion, N.V.; Peters, J.A.; Faccenda, E.; Harding, S.D.; Pawson, A.J.; Sharman, J.L. The concise guide to pharmacology 2017/18: G protein-coupled receptors. Br. J. Pharmacol. 2017, 174 (Suppl. 1), S17–S129. [Google Scholar] [CrossRef]
- Eva, C.; Oberto, A.; Sprengel, R.; Genazzani, E. The murine NPY-1 receptor gene. Structure and delineation of tissue-specific expression. FEBS Lett. 1992, 314, 285–288. [Google Scholar] [CrossRef] [Green Version]
- Rose, P.M.; Fernandes, P.; Lynch, J.S.; Frazier, S.T.; Fisher, S.M.; Kodukula, K.; Kienzle, B.; Seethala, R. Cloning and functional expression of a cDNA encoding a human type 2 neuropeptide Y receptor. J. Biol. Chem. 1995, 270, 22661–22664. [Google Scholar] [CrossRef] [Green Version]
- Gerald, C.; Walker, M.W.; Criscione, L.; Gustafson, E.L.; Batzl-Hartmann, C.; Smith, K.E.; Vaysse, P.; Durkin, M.M.; Laz, T.M.; Linemeyer, D.L. A receptor subtype involved in neuropeptide-Y-induced food intake. Nature 1996, 382, 168–171. [Google Scholar] [CrossRef]
- Michel, M.C. Receptors for neuropeptide Y: Multiple subtypes and multiple second messengers. Trends Pharmacol. Sci. 1991, 12, 389–394. [Google Scholar] [CrossRef]
- Stanić, D.; Brumovsky, P.; Fetissov, S.; Shuster, S.; Herzog, H.; Hökfelt, T. Characterization of neuropeptide Y2 receptor protein expression in the mouse brain. I. Distribution in cell bodies and nerve terminals. J. Comp. Neurol 2006, 499, 357–390. [Google Scholar] [CrossRef]
- Decressac, M.; Barker, R.A. Neuropeptide Y and its role in CNS disease and repair. Exp. Neurol. 2012, 238, 265–272. [Google Scholar] [CrossRef]
- Stanić, D.; Mulder, J.; Watanabe, M.; Hökfelt, T. Characterization of NPY Y2 receptor protein expression in the mouse brain. II. Coexistence with NPY, the Y1 receptor, and other neurotransmitter-related molecules. J. Comp. Neurol. 2011, 519, 1219–1257. [Google Scholar] [CrossRef]
- Pickel, V.M.; Beck-Sickinger, A.G.; Chan, J.; Weiland, H.A. Y1 receptors in the nucleus accumbens: Ultrastructural localization and association with neuropeptide Y. J. Neurosci. Res. 1998, 52, 54–68. [Google Scholar] [CrossRef]
- Härfstrand, A.; Fredholm, B.; Fuxe, K. Inhibitory effects of neuropeptide Y on cyclic AMP accumulation in slices of the nucleus tractus solitarius region of the rat. Neurosci. Lett. 1987, 76, 185–190. [Google Scholar] [CrossRef]
- Sun, Q.Q.; Huguenard, J.R.; Prince, D.A. Neuropeptide Y receptors differentially modulate G-protein-activated inwardly rectifying K+ channels and high-voltage-activated Ca2+ channels in rat thalamic neurons. J. Physiol. 2001, 531 Pt 1, 67–79. [Google Scholar] [CrossRef]
- Acuna-Goycolea, C.; Tamamaki, N.; Yanagawa, Y.; Obata, K.; van den Pol, A.N. Mechanisms of neuropeptide Y, peptide YY, and pancreatic polypeptide inhibition of identified green fluorescent protein-expressing GABA neurons in the hypothalamic neuroendocrine arcuate nucleus. J. Neurosci. 2005, 25, 7406–7419. [Google Scholar] [CrossRef] [Green Version]
- Sheriff, S.; Quershy, A.F.; Chance, W.T.; Kasckow, J.W.; Balasubramaniam, A. Predominant role by CaM kinase in NPY Y(1) receptor signaling: Involvement of CREB and Ambikaipakan. Peptides 2002, 23, 87–96. [Google Scholar] [CrossRef]
- Mullins, D.E.; Zhang, X.; Hawes, B.E. Activation of extracellular signal regulated protein kinase by neuropeptide Y and pancreatic polypeptide in CHO cells expressing the NPY Y(1), Y(2), Y(4) and Y(5) receptor subtypes. Regul. Pept. 2002, 105, 65–73. [Google Scholar] [CrossRef]
- Kask, A.; Harro, J.; von Horsten, S.; Redrobe, J.P.; Dumont, Y.; Quirion, R. The neurocircuitry and receptor subtypes mediating anxiolytic-like effects of neuropeptide Y. Neurosci. Biobehav. Rev. 2002, 26, 259–283. [Google Scholar] [CrossRef]
- Kopp, J.; Xu, Z.Q.; Zhang, X.; Pedrazzini, T.; Herzog, H.; Kresse, A.; Wong, H.; Walsh, J.H.; Hökfelt, T. Expression of the neuropeptide Y Y1 receptor in the CNS of rat and of wild-type and Y1 receptor knock-out mice. Focus on immunohistochemical localization. Neuroscience 2002, 111, 443–532. [Google Scholar] [CrossRef]
- Wolak, M.L.; DeJoseph, M.R.; Cator, A.D.; Mokashi, A.S.; Brownfield, M.S.; Urban, J.H. Comparative distribution of neuropeptide Y Y1 and Y5 receptors in the rat brain by using immunohistochemistry. J. Comp. Neurol. 2003, 464, 285–311. [Google Scholar] [CrossRef] [PubMed]
- Oberto, A.; Acquadro, E.; Bus, T.; Sprengel, R.; Eva, C. Expression patterns of promoters for NPY Y(1) and Y(5) receptors in Y(5)RitTA and Y(1)RVenus BAC-transgenic mice. Eur. J. Neurosci. 2007, 26, 155–170. [Google Scholar] [CrossRef]
- Mercer, R.E.; Chee, M.J.; Colmers, W.F. The role of NPY in hypothalamic mediated food intake. Front. Neuroendocrinol. 2011, 32, 398–415. [Google Scholar] [CrossRef] [PubMed]
- Tasan, R.O.; Verma, D.; Wood, J.; Lach, G.; Hormer, B.; de Lima, T.C.; Herzog, H.; Sperk, G. The role of Neuropeptide Y in fear conditioning and extinction. Neuropeptides 2016, 55, 111–126. [Google Scholar] [CrossRef] [Green Version]
- Gotzsche, C.R.; Woldbye, D.P. The role of NPY in learning and memory. Neuropeptides 2016, 55, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Tural, U.; Iosifescu, D.V. Neuropeptide Y in PTSD, MDD, and chronic stress: A systematic review and meta-analysis. J. Neurosci. Res. 2020, 98, 950–963. [Google Scholar] [CrossRef]
- Morales-Medina, J.C.; Dumont, Y.; Quirion, R. A possible role of neuropeptide Y in depression and stress. Brain Res. 2010, 1314, 194–205. [Google Scholar] [CrossRef]
- Comeras, L.B.; Herzog, H.; Tasan, R.O. Neuropeptides at the crossroad of fear and hunger: A special focus on neuropeptide Y. Ann. N. Y. Acad. Sci. 2019, 1455, 59–80. [Google Scholar] [CrossRef]
- Horn, S.R.; Charney, D.S.; Feder, A. Understanding resilience: New approaches for preventing and treating PTSD. Exp. Neurol. 2016, 284 Pt B, 119–132. [Google Scholar] [CrossRef]
- Schmeltzer, S.N.; Herman, J.P.; Sah, R. Neuropeptide Y (NPY) and posttraumatic stress disorder (PTSD): A translational update. Exp. Neurol. 2016, 284 Pt B, 196–210. [Google Scholar] [CrossRef]
- Sayed, S.; Van Dam, N.T.; Horn, S.R.; Kautz, M.M.; Parides, M.; Costi, S.; Collins, K.A.; Iacoviello, B.; Iosifescu, D.V.; Mathe, A.A. A Randomized Dose-Ranging Study of Neuropeptide Y in Patients with Posttraumatic Stress Disorder. Int. J. Neuropsychopharmacol. 2018, 21, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Aveleira, C.A.; Botelho, M.; Carmo-Silva, S.; Pascoal, J.F.; Ferreira-Marques, M.; Nóbrega, C.; Cortes, L.; Valero, J.; Sousa-Ferreira, L.; Álvaro, A.R. Neuropeptide Y stimulates autophagy in hypothalamic neurons. Proc. Natl. Acad. Sci. USA 2015, 112, E1642–E1651. [Google Scholar] [CrossRef] [Green Version]
- Croce, N.; Ciotti, M.T.; Gelfo, F.; Cortelli, S.; Federici, G.; Caltagirone, C.; Bernardini, S.; Angelucci, F. Neuropeptide Y protects rat cortical neurons against β-amyloid toxicity and re-establishes synthesis and release of nerve growth factor. ACS Chem. Neurosci. 2012, 3, 312–318. [Google Scholar] [CrossRef] [Green Version]
- Duarte-Neves, J.; Pereira de Almeida, L.; Cavadas, C. Neuropeptide Y (NPY) as a therapeutic target for neurodegenerative diseases. Neurobiol. Dis. 2016, 95, 210–224. [Google Scholar] [CrossRef]
- Baraban, S.C.; Hollopeter, G.; Erickson, J.C.; Schwartzkroin, P.A.; Palmiter, R.D. Knock-out mice reveal a critical antiepileptic role for neuropeptide Y. J. Neurosci. 1997, 17, 8927–8936. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.P.; Carvalho, A.P.; Carvalho, C.M.; Malva, J.O. Modulation of intracellular calcium changes and glutamate release by neuropeptide Y1 and Y2 receptors in the rat hippocampus: Differential effects in CA1, CA3 and dentate gyrus. J. Neurochem. 2001, 79, 286–296. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wu, X.; Liu, S.; Zhao, Y.; Zhu, J.; Liu, K. Roles of Neuropeptide Y in Neurodegenerative and Neuroimmune Diseases. Front. Neurosci. 2019, 13, 869. [Google Scholar] [CrossRef] [PubMed]
- Duarte-Neves, J.; Cavadas, C.; Pereira de Almeida, L. Neuropeptide Y (NPY) intranasal delivery alleviates Machado-Joseph disease. Sci. Rep. 2021, 11, 3345. [Google Scholar] [CrossRef]
- Clark, C.M.; Clark, R.M.; Hoyle, J.A.; Dickson, T.C. Pathogenic or protective? Neuropeptide Y in amyotrophic lateral sclerosis. J. Neurochem. 2021, 156, 273–289. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, R.M.; Phan, K.; Highton-Williamson, E.; Strikwerda-Brown, C.; Caga, J.; Ramsey, E.; Zoing, M.; Devenney, E.; Kim, W.S.; Hodges, J.R. Eating peptides: Biomarkers of neurodegeneration in amyotrophic lateral sclerosis and frontotemporal dementia. Ann. Clin. Transl. Neurol. 2019, 6, 486–495. [Google Scholar] [CrossRef] [Green Version]
- Meredith, G.E.; Agolia, R.; Arts, M.P.; Groenewegen, H.J.; Zahm, D.S. Morphological differences between projection neurons of the core and shell in the nucleus accumbens of the rat. Neuroscience 1992, 50, 149–162. [Google Scholar] [CrossRef]
- Záborszky, L.; Alheid, G.F.; Beinfeld, M.C.; Eiden, L.E.; Heimer, L.; Palkovits, M. Cholecystokinin innervation of the ventral striatum: A morphological and radioimmunological study. Neuroscience 1985, 14, 427–453. [Google Scholar] [CrossRef]
- Tripathi, A.; Prensa, L.; Cebrián, C.; Mengual, E. Axonal branching patterns of nucleus accumbens neurons in the rat. J. Comp. Neurol. 2010, 518, 4649–4673. [Google Scholar] [CrossRef] [PubMed]
- Zahm, D.S.; Brog, J.S. On the significance of subterritories in the “accumbens” part of the rat ventral striatum. Neuroscience 1992, 50, 751–767. [Google Scholar] [CrossRef]
- Ma, L.; Chen, W.; Yu, D.; Han, Y. Brain-Wide Mapping of Afferent Inputs to Accumbens Nucleus Core Subdomains and Accumbens Nucleus Subnuclei. Front. Syst. Neurosci. 2020, 14, 15. [Google Scholar] [CrossRef] [Green Version]
- van Dongen, Y.C.; Deniau, J.M.; Pennartz, C.M.; Galis-de Graaf, Y.; Voorn, P.; Thierry, A.M.; Groenewegen, H.J. Anatomical evidence for direct connections between the shell and core subregions of the rat nucleus accumbens. Neuroscience 2005, 136, 1049–1071. [Google Scholar] [CrossRef] [PubMed]
- Caberlotto, L.; Fuxe, K.; Sedvall, G.; Hurd, Y.L. Localization of neuropeptide Y Y1 mRNA in the human brain: Abundant expression in cerebral cortex and striatum. Eur. J. Neurosci 1997, 9, 1212–1225. [Google Scholar] [CrossRef]
- Aoki, C.; Pickel, V.M. Neuropeptide Y-containing neurons in the rat striatum: Ultrastructure and cellular relations with tyrosine hydroxylase- containing terminals and with astrocytes. Brain Res. 1988, 459, 205–225. [Google Scholar] [CrossRef]
- Massari, V.J.; Chan, J.; Chronwall, B.M.; O’Donohue, T.L.; Oertel, W.H.; Pickel, V.M. Neuropeptide Y in the rat nucleus accumbens: Ultrastructural localization in aspiny neurons receiving synaptic input from GABAergic terminals. J. Neurosci. Res. 1988, 19, 171–186. [Google Scholar] [CrossRef]
- Lüscher, C. The Emergence of a Circuit Model for Addiction. Annu. Rev. Neurosci. 2016, 39, 257–276. [Google Scholar] [CrossRef] [Green Version]
- Salin, P.; Kerkerian, L.; Nieoullon, A. Expression of neuropeptide Y immunoreactivity in the rat nucleus accumbens is under the influence of the dopaminergic mesencephalic pathway. Exp. Brain Res. 1990, 81, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Westwood, S.C.; Hanson, G.R. Effects of stimulants of abuse on extrapyramidal and limbic neuropeptide Y systems. J. Pharmacol. Exp. Ther. 1999, 288, 1160–1166. [Google Scholar]
- Wahlestedt, C.; Karoum, F.; Jaskiw, G.; Wyatt, R.J.; Larhammar, D.; Ekman, R.; Reis, D.J. Cocaine-induced reduction of brain neuropeptide Y synthesis dependent on medial prefrontal cortex. Proc. Natl. Acad. Sci. USA 1991, 88, 2078–2082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Midgley, L.P.; Bush, L.G.; Gibb, J.W.; Hanson, G.R. Differential regulation of neuropeptide Y systems in limbic structures of the rat. J. Pharmacol. Exp. Ther. 1993, 267, 707–713. [Google Scholar]
- Obuchowicz, E.; Turchan, J. Effects of acute or long-term treatment with chlorpromazine, haloperidol or sulpiride on neuropeptide Y-like immunoreactivity concentrations in the nucleus accumbens of rat. Eur. Neuropsychopharmacol. 1999, 9, 51–59. [Google Scholar] [CrossRef]
- Herman, Z.S. Neuropeptide Y (NPY) and its mRNA in discrete brain areas after subchronic administration of neuroleptics. Acta Neurobiol. Exp. 1996, 56, 55–61. [Google Scholar]
- Obuchowicz, E.; Turchan, J. Clozapine decreases neuropeptide Y-like immunoreactivity and neuropeptide Y mRNA levels in rat nucleus accumbens. Eur. Neuropsychopharmacol. 1999, 9, 329–335. [Google Scholar] [CrossRef]
- Krysiak, R.; Obuchowicz, E.; Herman, Z.S. Diazepam and buspirone alter neuropeptide Y-like immunoreactivity in rat brain. Neuropeptides 1999, 33, 542–549. [Google Scholar] [CrossRef]
- Dautan, D.; Huerta-Ocampo, I.; Witten, I.B.; Deisseroth, K.; Bolam, J.P.; Gerdjikov, T.; Mena-Segovia, J. A major external source of cholinergic innervation of the striatum and nucleus accumbens originates in the brainstem. J. Neurosci. 2014, 34, 4509–4518. [Google Scholar] [CrossRef] [Green Version]
- Pereira, P.A.; Vilela, M.; Sousa, S.; Neves, J.; Paula-Barbosa, M.M.; Madeira, M.D. Lesions of the laterodorsal tegmental nucleus alter the cholinergic innervation and neuropeptide Y expression in the medial prefrontal cortex and nucleus accumbens. Neuroscience 2015, 284, 707–718. [Google Scholar] [CrossRef]
- Robinson, S.L.; Thiele, T.E. The Role of Neuropeptide Y (NPY) in Alcohol and Drug Abuse Disorders. Int. Rev. Neurobiol. 2017, 136, 177–197. [Google Scholar]
- Thorsell, A.; Mathe, A.A. Neuropeptide Y in Alcohol Addiction and Affective Disorders. Front. Endocrinol. 2017, 8, 178. [Google Scholar] [CrossRef] [Green Version]
- Thiele, T.E.; Marsh, D.J.; Ste Marie, L.; Bernstein, I.L.; Palmiter, R.D. Ethanol consumption and resistance are inversely related to neuropeptide Y levels. Nature 1998, 396, 366–369. [Google Scholar] [CrossRef] [PubMed]
- Misra, K.; Pandey, S.C. Differences in basal levels of CREB and NPY in nucleus accumbens regions between C57BL/6 and DBA/2 mice differing in inborn alcohol drinking behavior. J. Neurosci. Res. 2003, 74, 967–975. [Google Scholar] [CrossRef] [PubMed]
- Hayes, D.M.; Knapp, D.J.; Breese, G.R.; Thiele, T.E. Comparison of basal neuropeptide Y and corticotropin releasing factor levels between the high ethanol drinking C57BL/6J and low ethanol drinking DBA/2J inbred mouse strains. Alcohol Clin. Exp. Res. 2005, 29, 721–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, S.C. Anxiety and alcohol abuse disorders: A common role for CREB and its target, the neuropeptide Y gene. Trends Pharmacol. Sci. 2003, 24, 456–460. [Google Scholar] [CrossRef]
- Czyzyk, T.A.; Sikorski, M.A.; Yang, L.; McKnight, G.S. Disruption of the RIIbeta subunit of PKA reverses the obesity syndrome of Agouti lethal yellow mice. Proc. Natl. Acad. Sci. USA 2008, 105, 276–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cummings, D.E.; Brandon, E.P.; Planas, J.V.; Motamed, K.; Idzerda, R.L.; McKnight, G.S. Genetically lean mice result from targeted disruption of the RII beta subunit of protein kinase A. Nature 1996, 382, 622–626. [Google Scholar] [CrossRef]
- Hayes, D.M.; Fee, J.R.; McCown, T.J.; Knapp, D.J.; Breese, G.R.; Cubero, I.; Carvajal, F.; Lerma-Cabrera, J.M.; Navarro, M.; Thiele, T.E. Neuropeptide Y signaling modulates the expression of ethanol-induced behavioral sensitization in mice. Addict. Biol. 2012, 17, 338–350. [Google Scholar] [CrossRef] [Green Version]
- Quadros, I.M.; Hipólide, D.C.; Frussa-Filho, R.; De Lucca, E.M.; Nobrega, J.N.; Souza-Formigoni, M.L. Resistance to ethanol sensitization is associated with increased NMDA receptor binding in specific brain areas. Eur. J. Pharmacol 2002, 442, 55–61. [Google Scholar] [CrossRef]
- Thiele, T.E.; Willis, B.; Stadler, J.; Reynolds, J.G.; Bernstein, I.L.; McKnight, G.S. High ethanol consumption and low sensitivity to ethanol-induced sedation in protein kinase A-mutant mice. J. Neurosci. 2000, 20, RC75. [Google Scholar] [CrossRef]
- Fee, J.R.; Knapp, D.J.; Sparta, D.R.; Breese, G.R.; Picker, M.J.; Thiele, T.E. Involvement of protein kinase A in ethanol-induced locomotor activity and sensitization. Neuroscience 2006, 140, 21–31. [Google Scholar] [CrossRef] [Green Version]
- Pereira, P.A.; Neves, J.; Vilela, M.; Sousa, S.; Cruz, C.; Madeira, M.D. Chronic alcohol consumption leads to neurochemical changes in the nucleus accumbens that are not fully reversed by withdrawal. Neurotoxicol. Teratol. 2014, 44, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Plescia, F.; Brancato, A.; Marino, R.A.; Vita, C.; Navarra, M.; Cannizzaro, C. Effect of Acetaldehyde Intoxication and Withdrawal on NPY Expression: Focus on Endocannabinoidergic System Involvement. Front. Psychiatry 2014, 5, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borkar, C.D.; Upadhya, M.A.; Shelkar, G.P.; Subhedar, N.K.; Kokare, D.M. Neuropeptide Y system in accumbens shell mediates ethanol self-administration in posterior ventral tegmental area. Addict. Biol. 2016, 21, 766–775. [Google Scholar] [CrossRef]
- Crabbe, J.C.; Metten, P.; Rhodes, J.S.; Yu, C.H.; Brown, L.L.; Phillips, T.J.; Finn, D.A. A line of mice selected for high blood ethanol concentrations shows drinking in the dark to intoxication. Biol. Psychiatry 2009, 65, 662–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barkley-Levenson, A.M.; Ryabinin, A.E.; Crabbe, J.C. Neuropeptide Y response to alcohol is altered in nucleus accumbens of mice selectively bred for drinking to intoxication. Behav. Brain Res. 2016, 302, 160–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brancato, A.; Castelli, V.; Lavanco, G.; Marino, R.A.M.; Cannizzaro, C. In utero Δ9-tetrahydrocannabinol exposure confers vulnerability towards cognitive impairments and alcohol drinking in the adolescent offspring: Is there a role for neuropeptide Y? J. Psychopharmacol. 2020, 34, 663–679. [Google Scholar] [CrossRef]
- Holderness, C.C.; Brooks-Gunn, J.; Warren, M.P. Co-morbidity of eating disorders and substance abuse review of the literature. Int. J. Eat. Disord. 1994, 16, 1–34. [Google Scholar] [CrossRef]
- Brown, C.M.; Coscina, D.V.; Fletcher, P.J. The rewarding properties of neuropeptide Y in perifornical hypothalamus vs. nucleus accumbens. Peptides 2000, 21, 1279–1287. [Google Scholar] [CrossRef]
- Maldonado-Irizarry, C.S.; Swanson, C.J.; Kelley, A.E. Glutamate receptors in the nucleus accumbens shell control feeding behavior via the lateral hypothalamus. J. Neurosci. 1995, 15, 6779–6788. [Google Scholar] [CrossRef]
- Stratford, T.R.; Swanson, C.J.; Kelley, A. Specific changes in food intake elicited by blockade or activation of glutamate receptors in the nucleus accumbens shell. Behav. Brain Res. 1998, 93, 43–50. [Google Scholar] [CrossRef]
- Stratford, T.R.; Kelley, A.E. GABA in the nucleus accumbens shell participates in the central regulation of feeding behavior. J. Neurosci. 1997, 17, 4434–4440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stratford, T.R.; Wirtshafter, D. NPY mediates the feeding elicited by muscimol injections into the nucleus accumbens shell. Neuroreport 2004, 15, 2673–2676. [Google Scholar] [CrossRef] [PubMed]
- Beck, B.; Jhanwar-Uniyal, M.; Burlet, A.; Chapleur-Chateau, M.; Leibowitz, S.F.; Burlet, C. Rapid and localized alterations of neuropeptide Y in discrete hypothalamic nuclei with feeding status. Brain Res. 1990, 528, 245–249. [Google Scholar] [CrossRef]
- Pandit, R.; Luijendijk, M.C.; Vanderschuren, L.J.; la Fleur, S.E.; Adan, R.A. Limbic substrates of the effects of neuropeptide Y on intake of and motivation for palatable food. Obesity 2014, 22, 1216–1219. [Google Scholar] [CrossRef] [Green Version]
- van den Heuvel, J.K.; Furman, K.; Gumbs, M.C.; Eggels, L.; Opland, D.M.; Land, B.B.; Kolk, S.M.; Narayanan, N.S.; Fliers, E.; Kalsbeek, A. Neuropeptide Y activity in the nucleus accumbens modulates feeding behavior and neuronal activity. Biol. Psychiatry 2015, 77, 633–641. [Google Scholar] [CrossRef] [Green Version]
- Pandit, R.; la Fleur, S.E.; Adan, R.A.H. The role of melanocortins and Neuropeptide Y in food reward. Eur. J. Pharmacol. 2013, 719, 208–214. [Google Scholar] [CrossRef]
- Kelley, A.E.; Will, M.J.; Steininger, T.L.; Zhang, M.; Haber, S.N. Restricted daily consumption of a highly palatable food (chocolate Ensure(R)) alters striatal enkephalin gene expression. Eur. J. Neurosci. 2003, 18, 2592–2598. [Google Scholar] [CrossRef]
- Zhang, M.; Gosnell, B.A.; Kelley, A.E. Intake of high-fat food is selectively enhanced by mu opioid receptor stimulation within the nucleus accumbens. J. Pharmacol. Exp. Ther. 1998, 285, 908–914. [Google Scholar]
- Meredith, G.E.; Pennartz, C.M.; Groenewegen, H.J. The cellular framework for chemical signalling in the nucleus accumbens. Prog. Brain Res. 1993, 99, 3–24. [Google Scholar]
- Zhou, L.; Furuta, T.; Kaneko, T. Chemical organization of projection neurons in the rat accumbens nucleus and olfactory tubercle. Neuroscience 2003, 120, 783–798. [Google Scholar] [CrossRef] [Green Version]
- Josselyn, S.A.; Beninger, R.J. Neuropeptide Y: Intraaccumbens injections produce a place preference that is blocked by cis-flupenthixol. Pharmacol. Biochem. Behav. 1993, 46, 543–552. [Google Scholar] [CrossRef]
- Sørensen, G.; Wegener, G.; Hasselstrøm, J.; Hansen, T.V.; Wörtwein, G.; Fink-Jensen, A.; Woldbye, D.P. Neuropeptide Y infusion into the shell region of the rat nucleus accumbens increases extracellular levels of dopamine. Neuroreport 2009, 20, 1023–1026. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, G.; Jensen, M.; Weikop, P.; Dencker, D.; Christiansen, S.H.; Loland, C.J.; Bengtsen, C.H.; Petersen, J.H.; Fink-Jensen, A.; Wörtwein, G. Neuropeptide Y Y5 receptor antagonism attenuates cocaine-induced effects in mice. Psychopharmacology 2012, 222, 565–577. [Google Scholar] [CrossRef] [PubMed]
- Desai, S.J.; Upadhya, M.A.; Subhedar, N.K.; Kokare, D.M. NPY mediates reward activity of morphine, via NPY Y1 receptors, in the nucleus accumbens shell. Behav. Brain Res. 2013, 247, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Tian, Z.; Ma, J.; Feng, Z.; Ou, Y.; Zhou, M.; Peng, J.; Lv, Y.; Gao, G.; Qi, S. NPY alterations induced by chronic morphine exposure affect the maintenance and reinstatement of morphine conditioned place preference. Neuropharmacology 2020, 181, 108350. [Google Scholar] [CrossRef]
- Warthen, K.G.; Sanford, B.; Walker, K.; Jones, K.G.; Angstadt, M.; Sripada, C.; Goldman, D.; Zubieta, J.K.; Welsh, R.C.; Burmeister, M. Neuropeptide Y and representation of salience in human nucleus accumbens. Neuropsychopharmacology 2019, 44, 495–502. [Google Scholar] [CrossRef] [Green Version]
- Rotzinger, S.; Lovejoy, D.A.; Tan, L.A. Behavioral effects of neuropeptides in rodent models of depression and anxiety. Peptides 2010, 31, 736–756. [Google Scholar] [CrossRef]
- Kautz, M.; Charney, D.S.; Murrough, J.W. Neuropeptide Y, resilience, and PTSD therapeutics. Neurosci. Lett. 2017, 649, 164–169. [Google Scholar] [CrossRef]
- Nahvi, R.J.; Sabban, E.L. Sex Differences in the Neuropeptide Y System and Implications for Stress Related Disorders. Biomolecules 2020, 10, 1248. [Google Scholar] [CrossRef] [PubMed]
- Caberlotto, L.; Fuxe, K.; Overstreet, D.H.; Gerrard, P.; Hurd, Y.L. Alterations in neuropeptide Y and Y1 receptor mRNA expression in brains from an animal model of depression: Region specific adaptation after fluoxetine treatment. Brain Res. Mol. Brain Res. 1998, 59, 58–65. [Google Scholar] [CrossRef]
- Krysiak, R.; Obuchowicz, E.; Herman, Z.S. Conditioned fear-induced changes in neuropeptide Y-like immunoreactivity in rats: The effect of diazepam and buspirone. Neuropeptides 2000, 34, 148–157. [Google Scholar] [CrossRef] [PubMed]
- van Megen, H.J.; Westenberg, H.G.; Den Boer, J.A.; Kahn, R.S. The panic-inducing properties of the cholecystokinin tetrapeptide CCK4 in patients with panic disorder. Eur. Neuropsychopharmacol. 1996, 6, 187–194. [Google Scholar] [CrossRef]
- Desai, S.J.; Borkar, C.D.; Nakhate, K.T.; Subhedar, N.K.; Kokare, D.M. Neuropeptide Y attenuates anxiety- and depression-like effects of cholecystokinin-4 in mice. Neuroscience 2014, 277, 818–830. [Google Scholar] [CrossRef]
- Nakhate, K.T.; Yedke, S.U.; Bharne, A.P.; Subhedar, N.K.; Kokare, D.M. Evidence for the involvement of neuropeptide Y in the antidepressant effect of imipramine in type 2 diabetes. Brain Res. 2016, 1646, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Aoki, M.; Watanabe, Y.; Yoshimoto, K.; Tsujimura, A.; Yamamoto, T.; Kanamura, N.; Tanaka, M. Involvement of serotonin 2C receptor RNA editing in accumbal neuropeptide Y expression and behavioural despair. Eur. J. Neurosci 2016, 43, 1219–1228. [Google Scholar] [CrossRef]
- Yamada, S.; Islam, M.S.; van Kooten, N.; Bovee, S.; Oh, Y.M.; Tsujimura, A.; Watanabe, Y.; Tanaka, M. Neuropeptide Y neurons in the nucleus accumbens modulate anxiety-like behavior. Exp. Neurol. 2020, 327, 113216. [Google Scholar] [CrossRef]
- Hökfelt, T.; Brumovsky, P.; Shi, T.; Pedrazzini, T.; Villar, M. NPY and pain as seen from the histochemical side. Peptides 2007, 28, 365–372. [Google Scholar] [CrossRef]
- Diaz-delCastillo, M.; Woldbye, D.P.D.; Heegaard, A.M. Neuropeptide Y and its Involvement in Chronic Pain. Neuroscience 2018, 387, 162–169. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, J.J.; Yu, L.C. Anti-nociceptive effect of neuropeptide Y in the nucleus accumbens of rats: An involvement of opioid receptors in the effect. Brain Res. 2002, 940, 69–78. [Google Scholar] [CrossRef]
- Allen, Y.S.; Bloom, S.R.; Polak, J.M. The neuropeptide Y-immunoreactive neuronal system: Discovery, anatomy and involvement in neurodegenerative disease. Hum. Neurobiol. 1986, 5, 227–234. [Google Scholar] [PubMed]
- Cannizzaro, C.; Tel, B.C.; Rose, S.; Zeng, B.Y.; Jenner, P. Increased neuropeptide Y mRNA expression in striatum in Parkinson’s disease. Brain Res. Mol. Brain Res. 2003, 110, 169–176. [Google Scholar] [CrossRef]
- Pereira, P.A.; Santos, D.; Neves, J.; Madeira, M.D.; Paula-Barbosa, M.M. Nerve growth factor retrieves neuropeptide Y and cholinergic immunoreactivity in the nucleus accumbens of old rats. Neurobiol. Aging 2013, 34, 1988–1995. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanaka, M.; Yamada, S.; Watanabe, Y. The Role of Neuropeptide Y in the Nucleus Accumbens. Int. J. Mol. Sci. 2021, 22, 7287. https://doi.org/10.3390/ijms22147287
Tanaka M, Yamada S, Watanabe Y. The Role of Neuropeptide Y in the Nucleus Accumbens. International Journal of Molecular Sciences. 2021; 22(14):7287. https://doi.org/10.3390/ijms22147287
Chicago/Turabian StyleTanaka, Masaki, Shunji Yamada, and Yoshihisa Watanabe. 2021. "The Role of Neuropeptide Y in the Nucleus Accumbens" International Journal of Molecular Sciences 22, no. 14: 7287. https://doi.org/10.3390/ijms22147287
APA StyleTanaka, M., Yamada, S., & Watanabe, Y. (2021). The Role of Neuropeptide Y in the Nucleus Accumbens. International Journal of Molecular Sciences, 22(14), 7287. https://doi.org/10.3390/ijms22147287