
Neural Contributions of the Hypothalamus to Parental Behaviour

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Parental Behaviour in Male Mice
3. Evidences of Neuromolecular Regulation of Parental Behaviour
4. Involvement of MCH Neurons in Parental Care
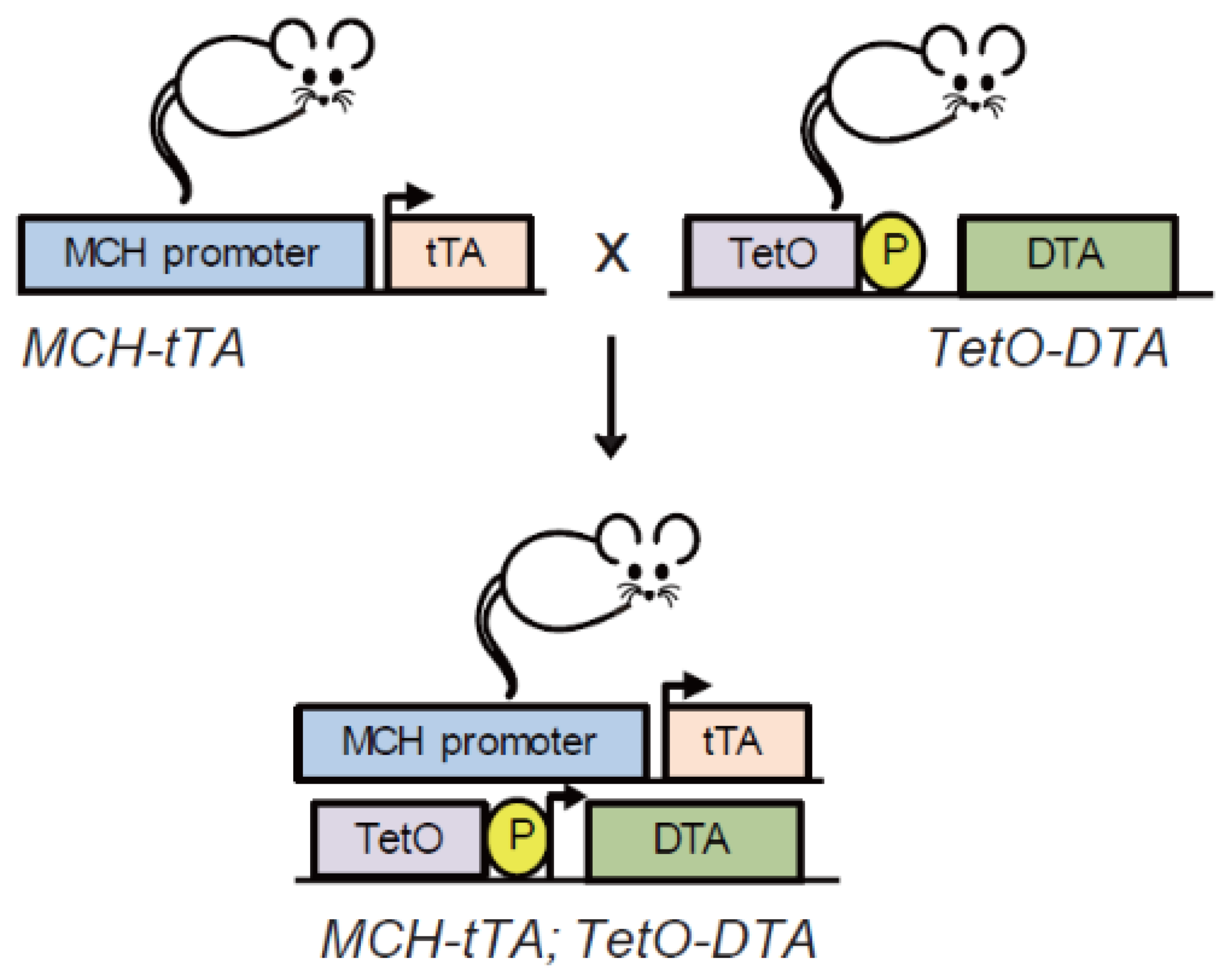
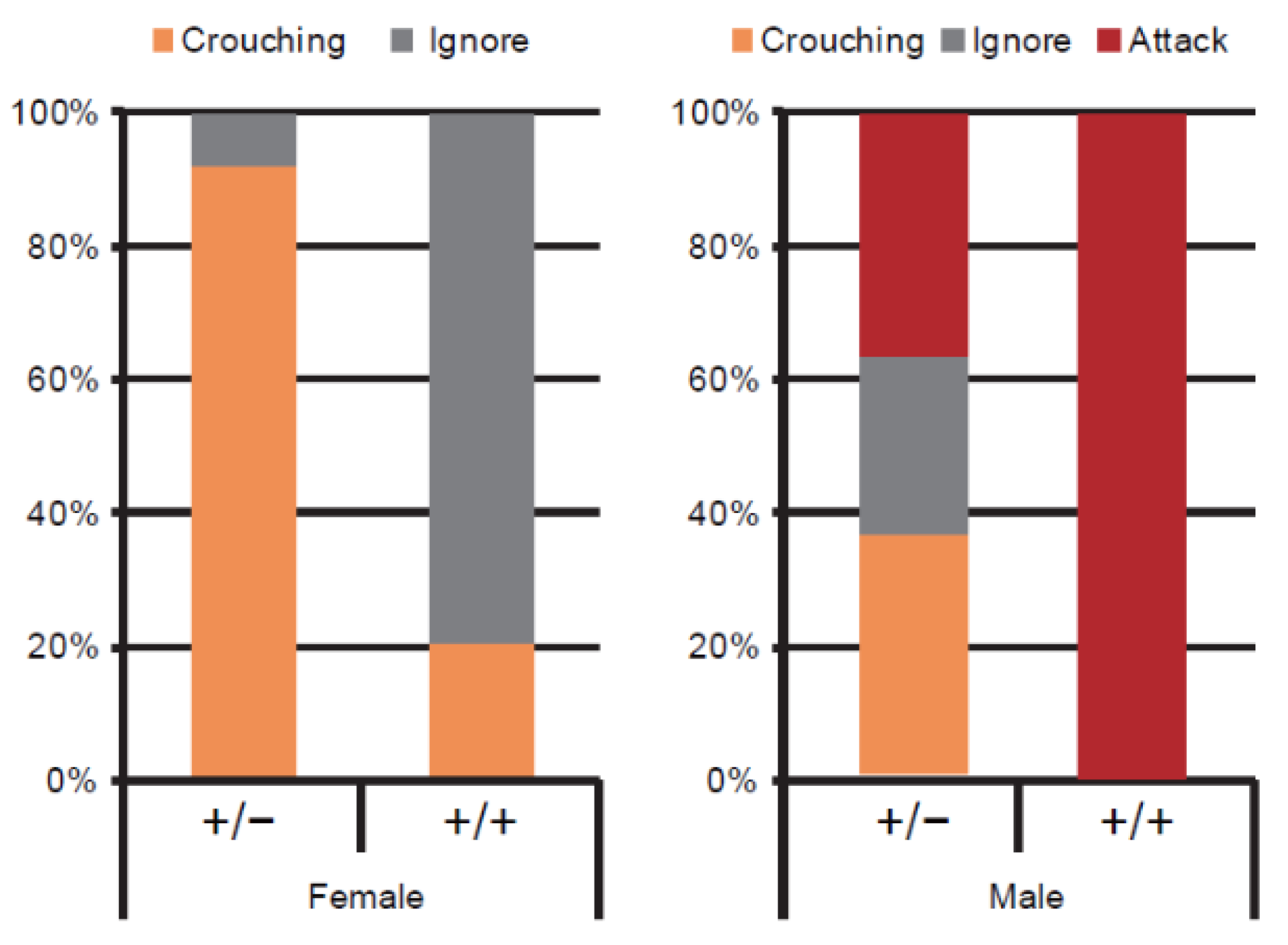
5. Effect of Congenital Ablation of MCH on Nursing Behaviour
6. Effect of Optogenetic Stimulation Intensity on Behaviour
7. MCH Neural Relay in PVN Oxytocin Neurons Is Involved in Nursing Behaviour
8. OT Enhances the Neural Circuits of Rewarding from Pups
9. Social Isolation Modifies GABAergic Transmission
10. Aggressive Behaviour towards Pups
11. Parental Licking Behaviour
12. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Numan, M. The Parental Behavior: Mechanisms, Development, and Evolution; Oxford University Press: Oxford, UK, 2020. [Google Scholar]
- Tachikawa, K.S.; Yoshihara, Y.; Kuroda, K.O. Behavioral transition from attack to parenting in male Mice: A crucial role of the vomeronasal system. J. Neurosci. 2013, 33, 5120–5126. [Google Scholar] [CrossRef]
- Wu, Z.; Autry, A.E.; Bergan, J.F.; Watabe-Uchida, M.; Dulac, C.G. Galanin neurons in the medial preoptic area govern parental behaviour. Nature 2014, 509, 325–330. [Google Scholar] [CrossRef] [Green Version]
- Marrocco, J.; McEwen, B.S. Sex in the brain: Hormones and sex differences. Dialogues Clin. Neurosci. 2016, 18, 373–383. [Google Scholar] [PubMed]
- Ubuka, T.; Trudeau, V.L.; Parhar, I. Editorial: Steroids and the Brain. Front. Endocrinol. 2020, 11, 366. [Google Scholar] [CrossRef]
- Blakemore, J.; Naftolin, F. Aromatase: Contributions to Physiology and Disease in Women and Men. Physiology 2016, 31, 258–269. [Google Scholar] [CrossRef] [Green Version]
- McEwen, B.S. Hormones and behavior and the integration of brain-body science. Horm. Behav. 2020, 119, 104619. [Google Scholar] [CrossRef]
- Arnold, A.P. A general theory of sexual differentiation. J. Neurosci. Res. 2017, 95, 291–300. [Google Scholar] [CrossRef] [Green Version]
- Bakker, J. The Sexual Differentiation of the Human Brain: Role of Sex Hormones Versus Sex Chromosomes. In Neuroendocrine Regulation of Behavior; Springer: Cham, Switzerland, 2018; Volume 43, pp. 45–67. [Google Scholar]
- McCarthy, M.M.; Wright, C.L. Convergence of Sex Differences and the Neuroimmune System in Autism Spectrum Disorder. Biol. Psychiatry 2017, 81, 402–410. [Google Scholar] [CrossRef] [Green Version]
- Forger, N.G. Epigenetic mechanisms in sexual differentiation of the brain and behaviour. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150114. [Google Scholar] [CrossRef] [Green Version]
- Döhler, K.D.; Coquelin, A.; Davis, F.; Hines, M.; Shryne, J.E.; Gorski, R.A. Differentiation of the sexually dimorphic nucleus in the preoptic area of the rat brain is determined by the perinatal hormone environment. Neurosci. Lett. 1982, 33, 295–298. [Google Scholar] [CrossRef]
- Mhaouty-Kodja, S.; Naulé, L.; Capela, D. Sexual Behavior: From Hormonal Regulation to Endocrine Disruption. Neuroendocrinology 2018, 107, 400–416. [Google Scholar] [CrossRef]
- Ribeiro, A.C.; Musatov, S.; Shteyler, A.; Simanduyev, S.; Arrieta-Cruz, I.; Ogawa, S.; Pfaff, D.W. siRNA silencing of estrogen receptor-α expression specifically in medial preoptic area neurons abolishes maternal care in female mice. Proc. Natl. Acad. Sci. USA 2012, 109, 16324–16329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orikasa, C.; Nagaoka, K.; Katsumata, H.; Sato, M.; Kondo, Y.; Minami, S.; Sakuma, Y. Social isolation prompts maternal behavior in sexually naïve male ddN mice. Physiol. Behav. 2015, 151, 9–15. [Google Scholar] [CrossRef]
- Mumtaz, F.; Khan, M.I.; Zubair, M.; Dehpour, A.R. Neurobiology and consequences of social isolation stress in animal model-A comprehensive review. Biomed. Pharm. 2018, 105, 1205–1222. [Google Scholar] [CrossRef]
- Buwalda, B.; Geerdink, M.; Vidal, J.; Koolhaas, J.M. Social behavior and social stress in adolescence: A focus on animal models. Neurosci. Biobehav. Rev. 2011, 35, 1713–1721. [Google Scholar] [CrossRef]
- Farbstein, D.; Hollander, N.; Peled, O.; Apter, A.; Fennig, S.; Haberman, Y.; Gitman, H.; Yaniv, I.; Shkalim, V.; Pick, C.G.; et al. Social isolation in mice: Behavior, immunity, and tumor growth. Stress 2021, 24, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Takeda, A.; Tamano, H.; Kan, F.; Hanajima, T.; Yamada, K.; Oku, N. Enhancement of social isolation-induced aggressive behavior of young mice by zinc deficiency. Life Sci. 2008, 82, 909–914. [Google Scholar] [CrossRef]
- O’Keefe, L.M.; Doran, S.J.; Mwilambwe-Tshilobo, L.; Conti, L.H.; Venna, V.R.; McCullough, L.D. Social isolation after stroke leads to depressive-like behavior and decreased BDNF levels in mice. Behav. Brain Res. 2014, 260, 162–170. [Google Scholar] [CrossRef] [Green Version]
- Koike, H.; Ibi, D.; Mizoguchi, H.; Nagai, T.; Nitta, A.; Takuma, K.; Nabeshima, T.; Yoneda, Y.; Yamada, K. Behavioral abnormality and pharmacologic response in social isolation reared mice. Behav. Brain Res. 2009, 202, 114–121. [Google Scholar] [CrossRef]
- Kozhemyakina, R.V.; Shikhevich, S.G.; Konoshenko, M.Y.; Gulevich, R.G. Adolescent oxytocin treatment affects resident behavior in aggressive but not tame adult rats. Physiol. Behav. 2020, 224, 113046. [Google Scholar] [CrossRef]
- Tan, O.; Musullulu, H.; Raymond, J.S.; Wilson, B.; Langguth, M.; Bowen, M.T. Oxytocin and vasopressin inhibit hyper-aggressive behaviour in socially isolated mice. Neuropharmacology. 2019, 156, 107573. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, J.R.; McMahon, E.K.; Boner, W.; Haussmann, M.F. Oxytocin administration prevents cellular aging caused by social isolation. Psychoneuroendocrinology 2019, 103, 52–60. [Google Scholar] [CrossRef]
- Ferdman, N.; Murmu, R.P.; Bock, J.; Braun, K.; Leshem, M. Weaning age, social isolation, and gender, interact to determine adult explorative and social behavior, and dendritic and spine morphology in prefrontal cortex of rats. Behav. Brain Res. 2007, 180, 174–182. [Google Scholar] [CrossRef]
- Du Preez, A.; Onorato, D.; Eiben, I.; Musaelyan, K.; Egeland, M.; Zunszain, P.A.; Fernandes, C.; Thuret, S.; Pariante, C.M. Chronic stress followed by social isolation promotes depressive-like behaviour, alters microglial and astrocyte biology and reduces hippocampal neurogenesis in male mice. Brain Behav. Immun. 2021, 91, 24–47. [Google Scholar] [CrossRef] [PubMed]
- Deguchi, Y.; Harada, M.; Shinohara, R.; Lazarus, M.; Cherasse, Y.; Urade, Y.; Yamada, D.; Sekiguchi, M.; Watanabe, D.; Furuyashiki, T.; et al. mDia and ROCK Mediate Actin-Dependent Presynaptic Remodeling Regulating Synaptic Efficacy and Anxiety. Cell Rep. 2016, 17, 2405–2417. [Google Scholar] [CrossRef] [Green Version]
- Matthews, G.A.; Edward, H.; Nieh, E.H.; Weele, C.M.V.; Halbert, S.A.; Pradhan, R.V.; Yosafat, A.S.; Glober, G.F.; Izadmehr, E.M.; Thomas, R.E.; et al. Dorsal Raphe Dopamine Neurons Represent the Experience of Social Isolation. Cell 2016, 164, 617–631. [Google Scholar] [CrossRef] [Green Version]
- Scott, N.; Prigge, M.; Yizhar, O.; Kimchi, T. A sexually dimorphic hypothalamic circuit controls maternal care and oxytocin secretion. Nature 2015, 525, 519–522. [Google Scholar] [CrossRef] [PubMed]
- Kohl, J.; Babayan, B.M.; Rubinstein, N.D.; Autry, A.E.; Marin-Rodriguez, B.; Kapoor, V.; Miyamishi, K.; Zweifel, L.S.; Luo, L.; Uchida, N.; et al. Functional circuit architecture underlying parental behaviour. Nature 2018, 556, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Marlin, B.J.; Mitre, M.; D’amour, J.A.; Chao, M.V.; Froemke, R.C. Oxytocin enables maternal behaviour by balancing cortical inhibition. Nature 2015, 520, 499–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watarai, A.; Tsutaki, S.; Nishimori, K.; Okuyama, T.; Mogi, K.; Kikusui, T. The blockade of oxytocin receptors in the paraventricular thalamus reduces maternal crouching behavior over pups in lactating mice. Neurosci. Lett. 2020, 720, 134761. [Google Scholar] [CrossRef] [PubMed]
- Yoshihar, C.; Numan, M.; Kuroda, K.O. Oxytocin and Parental Behaviors. Curr. Top. Behav. Neurosci. 2018, 35, 119–153. [Google Scholar]
- Rassovsky, Y.; Harwood, A.; Zagoory-Sharon, O.; Feldman, R. Martial arts increase oxytocin production. Sci. Rep. 2019, 9, 12980. [Google Scholar] [CrossRef]
- Tse, W.S.; Siu, A.F.Y.; Zhang, Q.; Chan, H.Y.E. Maternal oxytocin responsiveness improves specificity of positive social memory recall. Psychoneuroendocrinology 2018, 98, 148–152. [Google Scholar] [CrossRef]
- Bartz, J.A.; Zaki, J.; Ochsner, K.N.; Bolger, N.; Kolevzon, A.; Ludwig, N.; Lydon, J.E. Effects of oxytocin on recollections of maternal care and closeness. Proc. Natl. Acad. Sci. USA 2010, 107, 21371–21375. [Google Scholar] [CrossRef] [Green Version]
- Brill-Maoz, N.; Maroun, M. Extinction of fear is facilitated by social presence: Synergism with prefrontal oxytocin. Psychoneuroendocrinology 2016, 66, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Fu, L.Y.; Zhang, X.; van den Pol, A.N. Vasopressin and oxytocin excite MCH neurons, but not other lateral hypothalamic GABA neurons. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 302, R815–R824. [Google Scholar] [CrossRef] [Green Version]
- Qu, D.; Ludwig, D.S.; Gammeltoft, S.; Piper, M.; Pelleymounter, M.A.; Cullen, M.J.; Mathes, W.F.; Przypek, R.; Kanarek, R.; Maratos-Flier, E. A role for melanin-concentrating hormone in the central regulation of feeding behaviour. Nature 1996, 380, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Shimada, M.; Tritos, N.A.; Lowell, B.B.; Flier, J.S.; Maratos-Flier, E. Mice lacking melanin-concentrating hormone are hypophagic and lean. Nature 1998, 396, 670–674. [Google Scholar] [CrossRef]
- Marsh, D.J.; Weingarth, D.T.; Novi, D.E.; Chen, H.Y.; Trumbauer, M.E.; Chen, A.S.; Guan, X.M.; Jiang, M.M.; Feng, Y.; Camacho, R.E.; et al. Melanin-concentrating hormone 1 receptor-deficient mice are social isolation stress rodent lean, hyperactive, and hyperphagic and have altered metabolism. Proc. Natl. Acad. Sci. USA 2002, 99, 3240–3245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsunematsu, T.; Ueno, T.; Tabuchi, S.; Inutsuka, A.; Tanaka, K.F.; Hasuwa, H.; Kilduff, T.S.; Terao, A.; Yamanaka, A. Optogenetic manipulation of activity and temporally controlled cell-specific ablation reveal a role for MCH meurons in sleep/wake regulation. J. Neurosci. 2014, 34, 6896–6909. [Google Scholar] [CrossRef] [Green Version]
- Alhassen, L.; Phan, A.; Alhassen, W.; Nguyen, P.; Lo, A.; Shaharuddin, H.; Sanathara, N.; Civelli, O.; Alachkar, A. The role of Olfaction in MCH-regulated spontaneous maternal responses. Brain Res. 2019, 1719, 71–76. [Google Scholar] [CrossRef]
- Concetti, C.; Bracey, E.F.; Peleg-Raibstein, D.; Burdakov, D. Control of fear extinction by hypothalamic melanin-concentrating hormone-expressing neurons. Proc. Natl. Acad. Sci. USA 2020, 117, 22514–22521. [Google Scholar] [CrossRef]
- García-Fuster, M.J.; Parks, G.S.; Clinton, S.M.; Watson, S.J.; Akil, H.; Civelli, O. The melanin-concentrating hormone (MCH) system in an animal model of depression-like behavior. Eur. Neuropsychopharmacol. 2012, 22, 607–613. [Google Scholar] [CrossRef] [Green Version]
- Blouin, A.M.; Fried, I.; Wilson, C.L.; Staba, R.J.; Behnke, E.J.; Lam, H.A.; Maidment, N.T.; Karlsson, K.Æ.; Lapierre, J.L.; Siegel, J.M. Human hypocretin and melanin-concentrating hormone levels are linked to emotion and social interaction. Nat. Commun. 2013, 4, 1547. [Google Scholar] [CrossRef]
- Roy, M.; David, N.; Cueva, M.; Giorgetti, M. A study of the involvement of melanin-concentrating hormone receptor 1 (MCHR1) in murine models of depression. Biol. Psychiatry. 2007, 61, 174–180. [Google Scholar] [CrossRef]
- Sherwood, A.; Wosiski-Kuhn, M.; Nguyen, T.; Holland, P.C.; Lakaye, B.; Adamantidis, A.; Johnson, A.W. The role of melanin-concentrating hormone in conditioned reward learning. Eur. J. Neurosci. 2012, 36, 3126–3133. [Google Scholar] [CrossRef] [Green Version]
- Izawa, S.; Chowdhury, S.; Miyazaki, T.; Mukai, Y.; Ono, D.; Inoue, R.; Ohmura, Y.; Mizoguchi, H.; Kimura, K.; Yoshioka, M.; et al. REM sleep-active MCH neurons are involved in forgetting hippocampus-dependent memories. Science 2019, 365, 1308–1313. [Google Scholar] [CrossRef]
- Komagata, N.; Latifi, B.; Rusterholz, T.; Bassetti, C.L.A.; Adamantidis, A.; Schmidt, M.H. Dynamic REM Sleep Modulation by Ambient Temperature and the Critical Role of the Melanin-Concentrating Hormone System. Curr. Biol. 2019, 29, 1976–1987.e4. [Google Scholar] [CrossRef]
- Saito, Y.; Cheng, M.; Leslie, F.M.; Civelli, O. Expression of the melanin-concentrating hormone (MCH) receptor mRNA in the rat brain. J. Comp. Neurol. 2001, 435, 26–40. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.H.; Park, J.Y.; Oh, J.Y.; Bae, S.J.; Jang, H.; Jeon, S.; Kim, J.; Park, H.J. Novel analgesic effects of melanin-concentrating hormone on persistent neuropathic and inflammatory pain in mice. Sci. Rep. 2018, 8, 707. [Google Scholar] [CrossRef]
- Teixeira, P.D.S.; Wasinski, F.; Lima, L.B.; Frazão, R.; Bittencourt, J.C.; Donato, J., Jr. Regulation and neurochemical identity of melanin-concentrating hormone neurones in the preoptic area of lactating mice. J. Neuroendocrinol. 2020, 32, e12818. [Google Scholar] [CrossRef] [PubMed]
- Kawata, Y.; Okuda, S.; Hotta, N.; Igawa, H.; Takahashi, M.; Ikoma, M.; Kasai, S.; Ando, A.; Satomi, Y.; Nishida, M.; et al. A novel and selective melanin-concentrating hormone receptor 1 antagonist ameliorates obesity and hepatic steatosis in diet-induced obese rodent models. Eur. J. Pharmacol. 2017, 796, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Sanathara, N.M.; Garau, C.; Alachkar, A.; Wang, L.; Wang, Z.; Nishimori, K.; Xu, X.; Civelli, O. Melanin concentrating hormone modulates oxytocin-mediated marble burying. Neuropharmacology 2018, 128, 22–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawauchi, H.; Kawazoe, I.; Tsubokawa, M.; Kishida, M.; Baker, B.I. Characterization of melanin-concentrating hormone in chum salmon pituitaries. Nature 1983, 305, 321–323. [Google Scholar] [CrossRef]
- Bittencourt, J.C.; Presse, F.; Arias, C.; Peto, C.; Vaughan, J.; Nahon, J.L.; Vale, W.; Sawchenko, P.E. The melanin-concentrating hormone system of the rat brain: An immuno- and hybridization histochemical characterization. J. Comp. Neurol. 1992, 319, 218–245. [Google Scholar] [CrossRef]
- Alachkar, A.; Alhassen, L.; Wang, Z.; Wang, L.; Onouye, K.; Sanathara, N.; Civelli, O. Inactivation of the melanin concentrating hormone system impairs maternal behavior. Eur. Neuropsychopharmacol. 2016, 26, 1826–1835. [Google Scholar] [CrossRef] [Green Version]
- Kato, Y.; Katsumata, H.; Inutsuka, A.; Yamanaka, A.; Onaka, T.; Minami, S.; Orikasa, C. Involvement of MCH-oxytocin neural relay within the hypothalamus in murine nursing behavior. Sci. Rep. 2021, 11, 3348. [Google Scholar] [CrossRef]
- Kokkotou, E.; Jeon, J.Y.; Wang, X.; Marino, F.E.; Carlson, M.; Trombly, D.J.; Maratos-Flier, E. Mice with MCH ablation resist diet-induced obesity through strain-specific mechanisms. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 289, R117–R124. [Google Scholar] [CrossRef]
- Remedios, R.; Kennedy, A.; Zelikowsky, M.; Grewe, B.F.; Schnitzer, M.J.; Anderson, D.J. Social behaviour shapes hypothalamic neural ensemble representations of conspecific sex. Nature 2017, 550, 388–392. [Google Scholar] [CrossRef]
- Lee, H.; Kim, D.-W.; Remedios, R.; Anthony, T.E.; Chang, A.; Madisen, L.; Hongkui, Z.; Anderson, D.J. Scalable control of mounting and attack by Esr1+ neurons in the ventromedial hypothalamus. Nature 2014, 509, 627–632. [Google Scholar] [CrossRef]
- Costa, H.C.; Da-Silva, J.M.; Diniz, G.B.; Motta-Teixeira, L.C.; Da-Silva, R.J.; Battagello, D.S.; Sita, L.V.; de Moraes, M.C.; Horta-Júnior, J.A.C.; Bittencourt, J.C. Characterisation and origins of melanin-concentrating hormone immunoreactive fibres of the posterior lobe of the pituitary and median eminence during lactation in the Long-Evans rat. J. Neuroendocrinol. 2019, 31, e12723. [Google Scholar] [CrossRef]
- Lonstein, J.S.; Stern, J.M. Site and behavioral specificity of periaqueductal gray lesions on postpartum sexual, maternal, andaggressive behaviors in rats. Brain Res. 1998, 804, 21–35. [Google Scholar] [CrossRef]
- Salzberg, H.C.; Lonstein, J.S.; Stern, J.M. GABA(A) receptor regulation of kyphotic nursing and female sexual behavior in thecaudal ventrolateral periaqueductal gray of postpartum rats. Neuroscience 2002, 114, 675–687. [Google Scholar] [CrossRef]
- Rossier, D.; Franca, V.L.; Salemi, T.; Natale, S.; Gross, C.T. A neural circuit for competing approach and defense underlying prey capture. Proc. Natl. Acad. Sci. USA 2021, 118, e2013411118. [Google Scholar] [CrossRef]
- Tsuneoka, Y.; Maruyama, T.; Yoshida, S.; Nishimori, K.; Kato, T.; Numan, M.; Kuroda, K.O. Functional, anatomical, and neurochemical differentiation of medial preoptic area subregions in relation to maternal behavior in the mouse. J. Comp. Neurol. 2013, 521, 1633–1663. [Google Scholar] [CrossRef]
- Fang, Y.Y.; Yamaguchi, T.; Song, S.C.; Tritsch, N.X.; Lin, D.A. Hypothalamic Midbrain Pathway Essential for Driving Maternal Behaviors. Neuron 2018, 98, 192–207.e10. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.-C.; Wang, S.-R.; Jiao, Z.-L.; Zhang, W.; Lin, J.-K.; Li, X.-Y.; Li, S.-S.; Zhang, X.; Xu, X.-H. Medial preoptic area in mice is capable of mediating sexually dimorphic behaviors regardless of gender. Nat. Commun. 2018, 18, 279. [Google Scholar] [CrossRef]
- Numan, M. Medial preoptic area and maternal behavior in the female rat. J. Comp. Physiol. Psychol. 1974, 87, 746–759. [Google Scholar] [CrossRef]
- Young, L.J.; Wang, Z. The neurobiology of pair bonding. Nat. Neurosci. 2004, 7, 1048–1054. [Google Scholar] [CrossRef]
- Kelly, A.M.; Hiura, L.C.; Saunders, A.G.; Ophir, A.G. Oxytocin Neurons Exhibit Extensive Functional Plasticity Due To Offspring Age in Mothers and Fathers. Integr. Comp. Biol. 2017, 57, 603–618. [Google Scholar] [CrossRef]
- Caitlin Post, C.; Leuner, B. The Maternal Reward System in Postpartum Depression. Arch. Womens Ment. Health 2019, 22, 417–429. [Google Scholar] [CrossRef] [PubMed]
- Del Cid-Pellitero, E.; Jones, B.E. Immunohistochemical evidence for synaptic release of GABA from melanin-concentrating hormone containing varicosities in the locus coeruleus. Neuroscience 2012, 223, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Elias, C.F.; Lee, C.E.; Kelly, J.F.; Ahima, R.S.; Kuhar, M.; Saper, C.B.; Elmquist, J.K. Characterization of CART neurons in the rat and human hypothalamus. J. Comp. Neurol. 2001, 432, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Harthoorn, L.F.; Sañé, A.; Nethe, M.; Heerikhuize, J.J.V. Multi-transcriptional profiling of melanin-concentrating hormone and orexin-containing neurons. Cell Mol. Neurobiol. 2005, 25, 1209–1223. [Google Scholar] [CrossRef]
- Abrahamson, E.E.; Leak, R.K.; Moore, R.Y. The suprachiasmatic nucleus projects to posterior hypothalamic arousal systems. Neuroreport 2001, 12, 435–440. [Google Scholar] [CrossRef]
- Chee, M.J.S.; Arrigoni, E.; Maratos-Flier, E. Melanin-concentrating hormone neurons release glutamate for feedforward inhibition of the lateral septum. J. Neurosci. 2015, 35, 3644–3651. [Google Scholar] [CrossRef] [PubMed]
- Jennings, J.H.; Sparta, D.R.; Stamatakis, A.M.; Ung, R.L.; Pleil, K.E.; Kash, T.L.; Stuber, G.D. Distinct extended amygdala circuits for divergent motivational states. Nature 2013, 496, 224–228. [Google Scholar] [CrossRef]
- McHenry, J.A.; Otis, J.M.; Rossi, M.A.; Robinson, J.E.; Kosyk, O.; Miller, N.W.; McElligott, Z.A.; Budygin, E.A.; Rubinow, D.R.; Stuber, G.D. Hormonal gain control of a medial preoptic area social reward circuit. Nat. Neurosci. 2017, 20, 449–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gies, U.; Theodosis, D.T. Synaptic plasticity in the rat supraoptic nucleus during lactation involves GABA innervation and oxytocin neurons: A quantitative immunocytochemical analysis. J. Neurosci. 1994, 14 Pt 1, 2861–2869. [Google Scholar] [CrossRef]
- Li, Y.; Gao, X.B.; Sakurai, T.; van den Pol, A.N. Hypocretin/orexin excites hypocretin neurons via a local glutamate neuron-a potential mechanism for orchestrating the hypothalamic arousal system. Neuron 2002, 36, 1169–1181. [Google Scholar] [CrossRef] [Green Version]
- Marty, A.; Llano, I. Excitatory effects of GABA in established brain networks. Trends Neurosci. 2005, 28, 284–289. [Google Scholar] [CrossRef]
- Kim, J.S.; Kim, W.B.; Kim, Y.-B.; Lee, Y.; Kim, Y.S.; Shen, F.-Y.; Lee, S.W.; Park, D.; Choi, H.-J.; Hur, J.; et al. Chronic hyperosmotic stress converts GABAergic inhibition into excitation in vasopressin and oxytocin neurons in the rat. J. Neurosci. 2011, 31, 13312–13322. [Google Scholar] [CrossRef]
- Lee, S.W.; Kim, Y.B.; Kim, J.S.; Kim, W.B.; Kim, Y.S.; Han, H.C.; Colwell, C.S.; Cho, Y.W.; Kim, Y.I. GABAergic inhibition is weakened or converted into excitation in the oxytocin and vasopressin neurons of the lactating rat. Mol. Brain 2015, 8, 34. [Google Scholar] [CrossRef] [Green Version]
- Choi, K.; Lee, Y.; Lee, C.; Hong, S.; Lee, S.; Kang, S.J.; Shin, K.S. Optogenetic activation of septal GABAergic afferents entrains neuronal firing in the medial habenula. Sci. Rep. 2016, 6, 34800. [Google Scholar] [CrossRef]
- Tirindelli, R.; Dibattista, M.; Pifferi, S.; Menini, A. From pheromones to behavior. Physiol. Rev. 2009, 89, 921–956. [Google Scholar] [CrossRef]
- Brennan, P.A.; Zufall, F. Pheromonal communication in vertebrates. Nature 2006, 444, 308–315. [Google Scholar] [CrossRef]
- Gutiérrez-Castellanos, N.; Martínez-Marcos, A.; Martínez-García, F.; Lanuza, E. Chemosensory function of the amygdala. Vitam. Horm. 2010, 83, 165–196. [Google Scholar]
- Trainor, B.C.; Rowland, M.R.; Nelson, R.J. Photoperiod affects estrogen receptor alpha, estrogen receptor beta and aggressive behavior. Eur. J. Neurosci. 2007, 26, 207–218. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Coats, J.K.; Yang, C.F.; Wang, A.; Ahmed, O.M.; Alvarado, M.; Izumi, T.; Shah, N.M. Modular genetic control of sexually dimorphic behaviors. Cell 2012, 148, 596–607. [Google Scholar] [CrossRef] [Green Version]
- Trouillet, A.-C.; Keller, M.; Jan Weiss, J.; Leinders-Zufall, T.; Birnbaumer, L.; Frank Zufall, F.; Pablo Chamero, P. Central role of G protein Gαi2 and Gαi2 + vomeronasal neurons in balancing territorial and infant-directed aggression of male mice. Proc. Natl. Acad. Sci. USA 2019, 116, 5135–5143. [Google Scholar] [CrossRef] [Green Version]
- Chen, A.-X.; Yan, J.-J.; Zhang, W.; Wang, L.; Yu, Z.-X.; Ding, X.-J.; Wang, D.-Y.; Zhang, M.; Zhang, Y.-L.; Song, N.; et al. Specific Hypothalamic Neurons Required for Sensing Conspecific Male Cues Relevant to Inter-male Aggression. Neuron 2020, 108, 763–774.e6. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, S.D.; Robertson, D.H.L.; Cheetham, S.A.; Hurst, J.L.; Beynon, R.J. Structural and functional differences in isoforms of mouse major urinary proteins: A male-specific protein that preferentially binds a male pheromone. Biochem. J. 2005, 391 Pt 2, 343–350. [Google Scholar] [CrossRef] [Green Version]
- Tolokh, I.I.; Fu, X.; Holy, T.E. Reliable sex and strain discrimination in the mouse vomeronasal organ and accessory olfactory bulb. J. Neurosci. 2013, 33, 13903–13913. [Google Scholar] [CrossRef] [Green Version]
- Isoga, Y.; Si, S.; Pont-Lezica, L.; Tan, T.; Kapoor, V.; Murthy, V.N.; Dulac, C. Molecular organization of vomeronasal chemoreception. Nature 2011, 478, 241–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimoto, H.; Sato, K.; Nodari, F.; Haga, S.; Holy, T.E.; Touhara, K. Sex- and strain-specific expression and vomeronasal activity of mouse ESP family peptides. Curr. Biol. 2007, 21, 1879–1884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Dudley, C.A.; Moss, R.L. Functional dichotomy within the vomeronasal system: Distinct zones of neuronal activity in the accessory olfactory bulb correlate with sex-specific behaviors. J. Neurosci. 1999, 19, RC32. [Google Scholar] [CrossRef]
- Leypold, B.G.; Yu, C.R.; Leinders-Zufall, T.; Kim, M.M.; Zufall, F.; Axel, R. Altered sexual and social behaviors in trp2 mutant mice. Proc. Natl. Acad. Sci. USA 2002, 99, 6376–6381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stowers, L.; Holy, T.E.; Meister, M.; Dulac, C.; Koentges, G. Loss of sex discrimination and male-male aggression in mice deficient for TRP2. Science 2002, 295, 1493–1500. [Google Scholar] [CrossRef]
- Orikasa, C.; Kondo, Y.; Katsumata, H.; Terada, M.; Akimoto, T.; Sakuma, Y.; Minami, S. Vomeronasal signal deficiency enhances parental behavior in socially isolated male mice. Physiol. Behav. 2017, 168, 98–102. [Google Scholar] [CrossRef]
- Stolzenberg, D.S.; Mayer, H.S. Experience-dependent mechanisms in the regulation of parental care. Front. Neuroendocrinol. 2019, 54, 100745. [Google Scholar] [CrossRef] [PubMed]
- Fleming, A.S.; Vaccarino, F.; Luebke, C. Amygdaloid inhibition of maternal behavior in the nulliparous female rat. Physiol. Behav. 1980, 25, 731–743. [Google Scholar] [CrossRef]
- Numan, M.; Numan, M.J.; English, J.B. Excitotoxic amino acid injections into the medial amygdala facilitate maternal behavior in virgin female rats. Horm. Behav. 1993, 27, 56–81. [Google Scholar] [CrossRef]
- Sheehan, T.; Paul, M.; Amaral, E.; Numan, M.J.; Numan, M. Evidence that the medial amygdala projects to the anterior/ventromedial hypothalamic nuclei to inhibit maternal behavior in rats. Neuroscience 2001, 106, 341–356. [Google Scholar] [CrossRef]
- Numan, M.; Young, L.J. Neural mechanisms of mother-infant bonding and pair bonding: Similarities, differences, and broader implications. Horm. Behav. 2016, 77, 98–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, J.G.; Yokota, S.; Tsumori, T.; Oka, T.; Yasui, Y. Projections from the anterior basomedial and anteriorcortical amygdaloid nuclei to melanin-concentrating hormone-containing neurons in the lateral hypothalamus of the rat. Brain Res. 2012, 1479, 31–43. [Google Scholar] [CrossRef]
- De Jong, T.R.; Neumann, I.D. Oxytocin and Aggression. Curr. Top. Behav. Neurosci. 2018, 35, 175–192. [Google Scholar]
- Pedersen, C.A.; Caldwell, J.D.; Walker, C.; Ayers, G.; Mason, G.A. Oxytocin activates the postpartum onset of rat maternal behavior in the ventral tegmental and medial preoptic areas. Behav. Neurosci. 1994, 108, 1163–1171. [Google Scholar] [CrossRef]
- Zhang, G.-W.; Shen, L.; Tao, C.; Jung, A.-H.; Peng, B.; Li, Z.; Zhang, L.I.; Tao, H.W. Medial preoptic area antagonistically mediates stress-induced anxiety and parental behavior. Nat. Neurosci. 2021, 24, 516–528. [Google Scholar] [CrossRef]
- Anpilov, S.; Shemesh, Y.; Eren, N.; Harony-Nicolas, H.; Benjamin, A.; Dine, J.; Oliveira, V.E.M.; Forkosh, O.; Karamihalev, S.; Hüttl, R.E.; et al. Wireless Optogenetic Stimulation of Oxytocin Neurons in a Semi-natural Setup Dynamically Elevates Both Pro-social and Agonistic Behaviors. Neuron 2020, 107, 644–655.e7. [Google Scholar] [CrossRef]
- Shamay-Tsoory, S.G.; Abu-Akel, A. The Social Salience Hypothesis of Oxytocin. Biol. Psychiatry 2016, 79, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Steinman, M.Q.; Duque-Wilckens, N.; Trainor, B.C. Complementary Neural Circuits for Divergent Effects of Oxytocin: Social Approach Versus Social Anxiety. Biol. Psychiatry 2019, 85, 792–801. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, V.; Maltese, F.; Contarini, G.; Nigro, M.; Bonavia, A.; Huang, H.; Gigliucci, V.; Morelli, G.; Scheggia, D.; Managò, F.; et al. Oxytocin Signaling in the Central Amygdala Modulates Emotion Discrimination in Mice. Curr. Biol. 2019, 29, 1938–1953.e6. [Google Scholar] [CrossRef] [PubMed]
- Caldji, C.; Diorio, J.; Meaney, M.J. Variations in maternal care in infancy regulate the development of stress reactivity. Biol. Psychiatry 2000, 48, 1164–1174. [Google Scholar] [CrossRef]
- Francis, D.D.; Szegda, K.; Campbell, G.; Martin, W.D.; Insel, T.R. Epigenetic sources of behavioral differences in mice. Nat. Neurosci. 2003, 6, 445–446. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orikasa, C. Neural Contributions of the Hypothalamus to Parental Behaviour. Int. J. Mol. Sci. 2021, 22, 6998. https://doi.org/10.3390/ijms22136998
Orikasa C. Neural Contributions of the Hypothalamus to Parental Behaviour. International Journal of Molecular Sciences. 2021; 22(13):6998. https://doi.org/10.3390/ijms22136998
Chicago/Turabian StyleOrikasa, Chitose. 2021. "Neural Contributions of the Hypothalamus to Parental Behaviour" International Journal of Molecular Sciences 22, no. 13: 6998. https://doi.org/10.3390/ijms22136998
APA StyleOrikasa, C. (2021). Neural Contributions of the Hypothalamus to Parental Behaviour. International Journal of Molecular Sciences, 22(13), 6998. https://doi.org/10.3390/ijms22136998