
The Role of the Second Extracellular Loop of Norepinephrine Transporter, Neurotrophin-3 and Tropomyosin Receptor Kinase C in T Cells: A Peripheral Biomarker in the Etiology of Schizophrenia

 ,
,  ,
, 

Abstract
:1. Introduction
2. Results
2.1. Demographic and Clinical Data Analysis
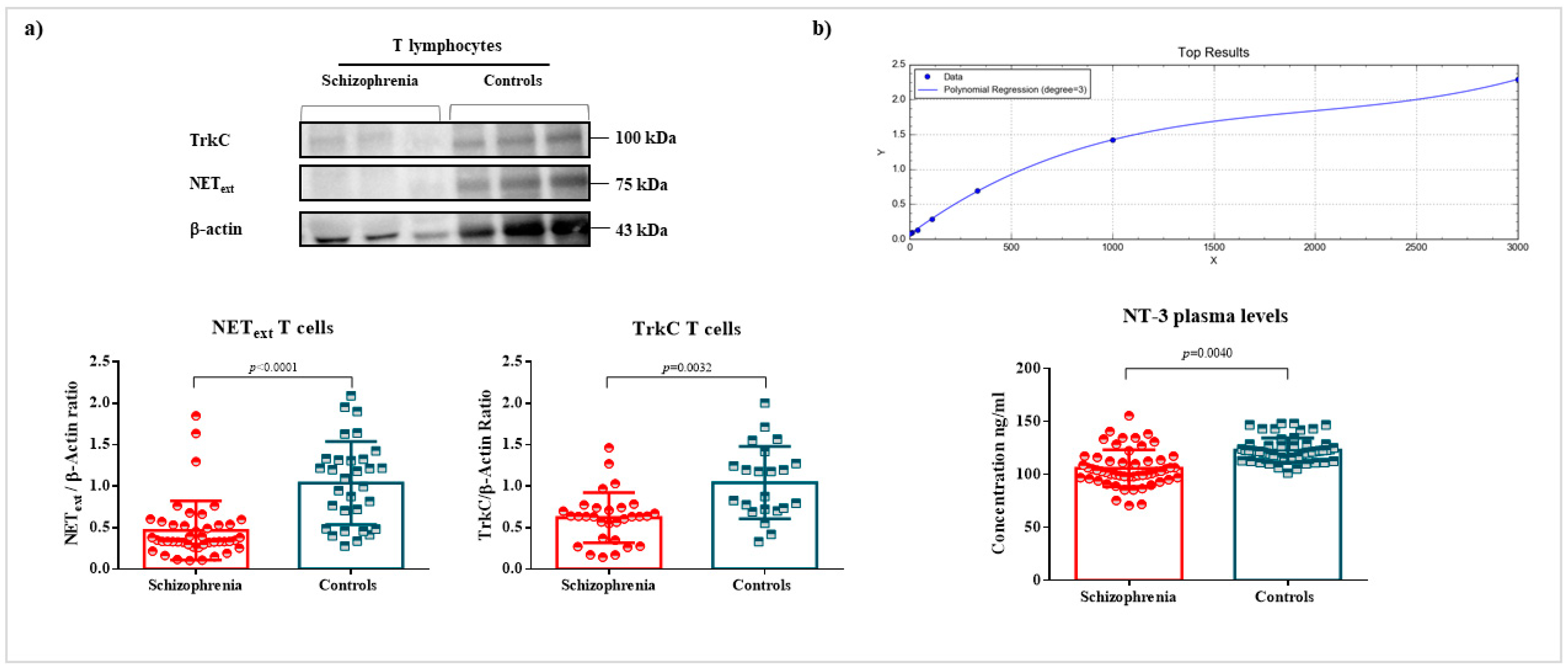
2.2. Quantification of NET, TrkC, and NT-3 Levels
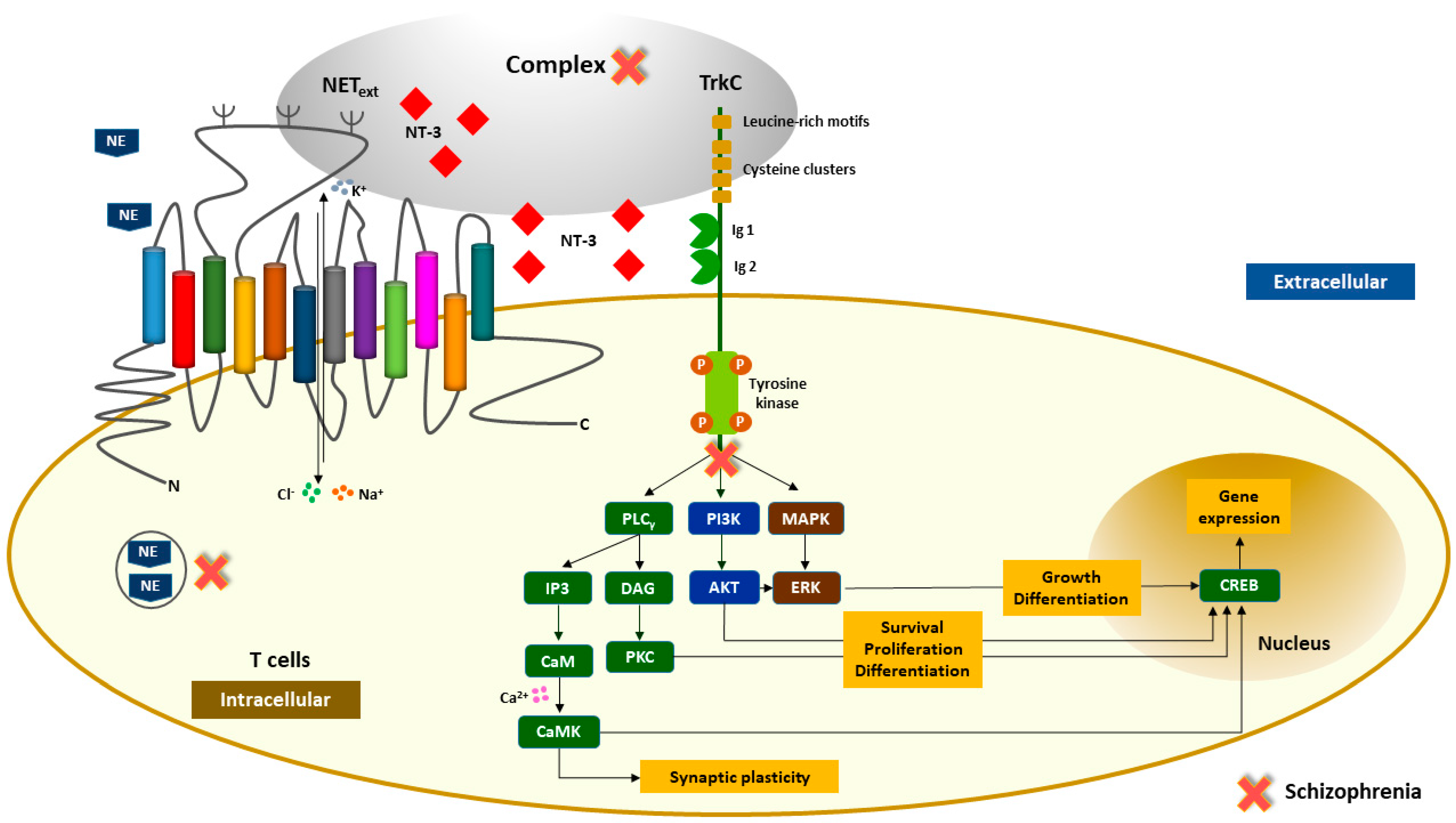
2.3. Identification of an Interaction between the Second Extracellular Loop of NET and NT-3 or TrkC in T Lymphocytes
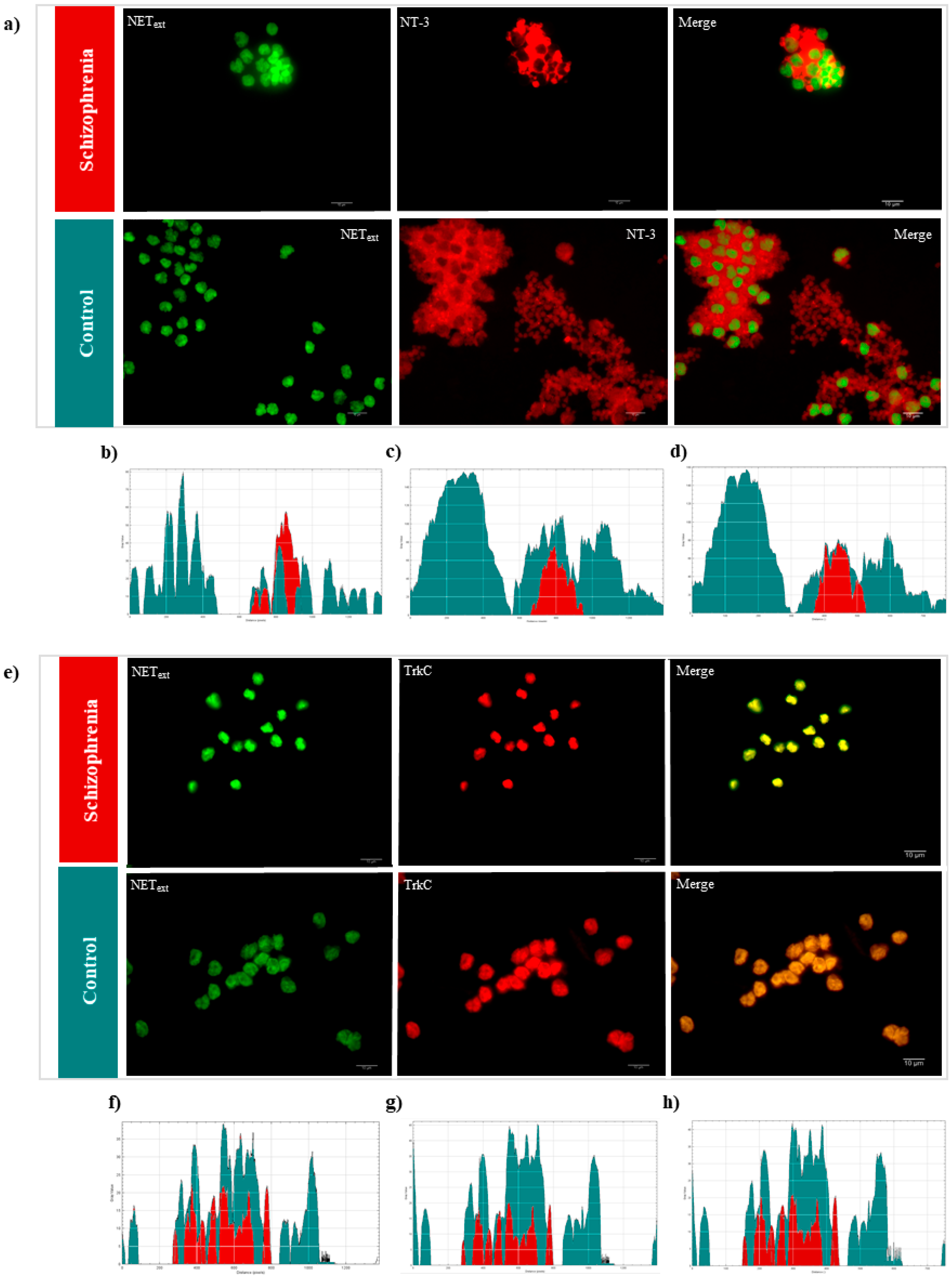
2.4. Immunofluorescence Microscopy
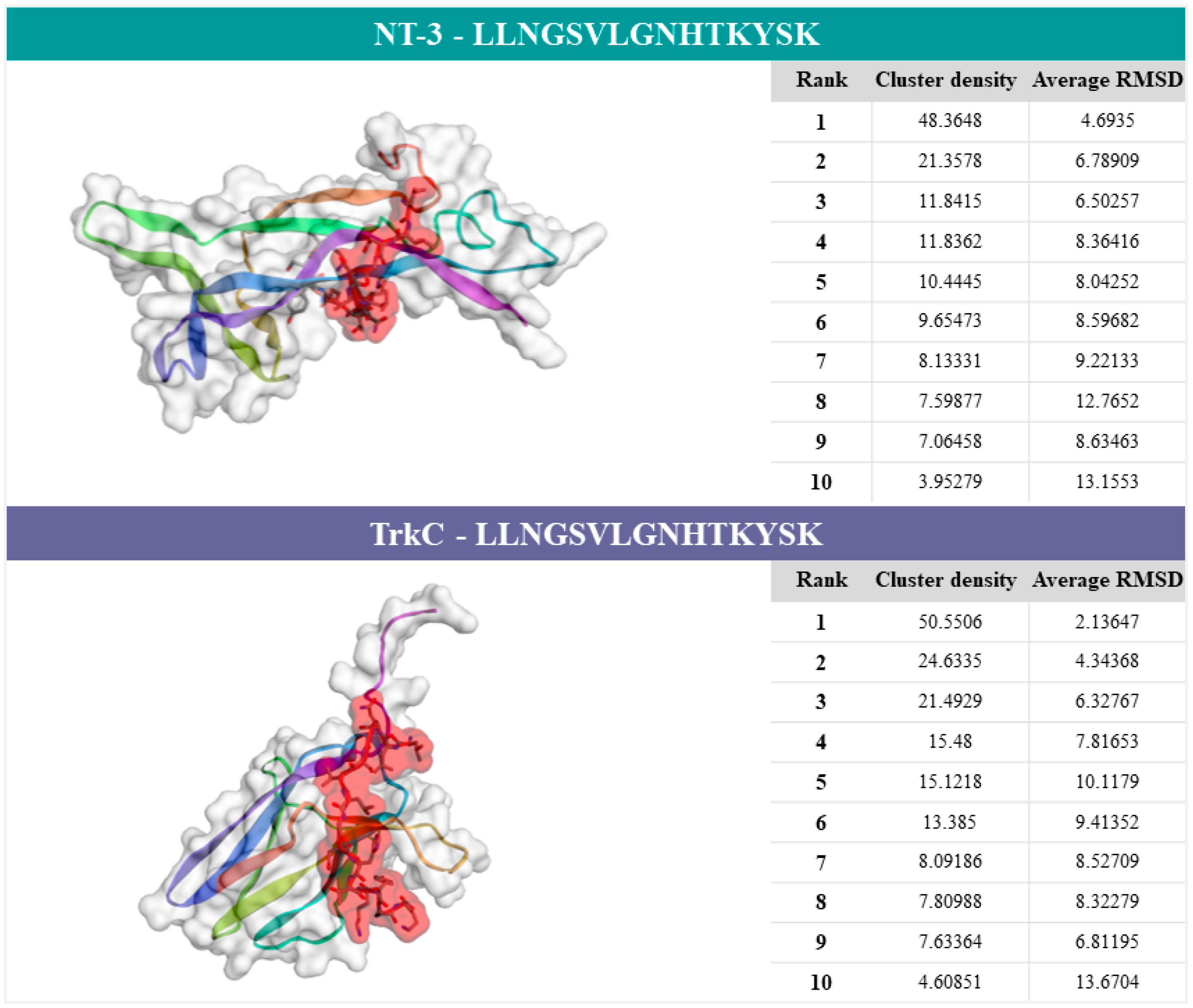
2.5. Characterization of NT-3–NEText and TrkC–NEText Docking Predictions
3. Discussion
4. Materials and Methods
4.1. Subjects and Samples
4.2. Clinical and Demographic Assessment
4.3. Sample Preparation
4.4. Immunoprecipitation (IP) and Western Blot
4.5. Immunofluorescence Microscopy
4.6. NT-3 Plasma Quantification
4.7. NT-3 or TrkC Proteins and NEText Peptide Docking Prediction
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Joyce, E.M.; Roiser, J.P. Cognitive heterogeneity in schizophrenia. Curr. Opin. Psychiatry 2007, 20, 268–272. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.R.; Cherian, J.; Gohil, K.; Atkinson, D. Schizophrenia: Overview and treatment options. Pharm. Ther. 2014, 39, 638–645. [Google Scholar]
- Horváth, S.; Mirnics, K. Schizophrenia as a disorder of molecular pathways. Biol. Psychiatry 2015, 77, 22–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, M.; Shirakawa, O.; Toyooka, K.; Kitamura, N.; Hashimoto, T.; Maeda, K.; Koizumi, S.; Wakabayashi, K.; Takahashi, H.; Someya, T.; et al. Abnormal expression of brain-derived neurotrophic factor and its receptor in the corticolimbic system of schizophrenic patients. Mol. Psychiatry 2000, 5, 293–300. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues-Amorim, D.; Rivera-Baltanás, T.; Bessa, J.; Sousa, N.; de Carmen Vallejo-Curto, M.; Rodríguez-Jamardo, C.; de las Heras, M.E.; Díaz, R.; Agís-Balboa, R.C.; Olivares, J.M.; et al. The neurobiological hypothesis of neurotrophins in the pathophysiology of schizophrenia: A meta-analysis. J. Psychiatr. Res. 2018, 106, 43–53. [Google Scholar] [CrossRef]
- Buckley, P.F.; Mahadik, S.; Pillai, A.; Terry, A. Neurotrophins and schizophrenia. Schizophr. Res. 2007, 94, 1–11. [Google Scholar] [CrossRef]
- Shoval, G.; Weizman, A. The possible role of neurotrophins in the pathogenesis and therapy of schizophrenia. Eur. Neuropsychopharmacol. 2005, 15, 319–329. [Google Scholar] [CrossRef]
- Durany, N.; Michel, T.; Zöchling, R.; Boissl, K.W.; Cruz-Sánchez, F.F.; Riederer, P.; Thome, J. Brain-derived neurotrophic factor and neurotrophin 3 in schizophrenic psychoses. Schizophr. Res. 2001, 52, 79–86. [Google Scholar] [CrossRef]
- Huang, E.J.; Reichardt, L.F. Neurotrophins: Roles in neuronal development and function. Annu. Rev. Neurosci. 2001, 24, 677–736. [Google Scholar] [CrossRef] [Green Version]
- Keefe, K.M.; Sheikh, I.S.; Smith, G.M. Targeting neurotrophins to specific populations of neurons: NGF, BDNF, and NT-3 and their relevance for treatment of spinal cord injury. Int. J. Mol. Sci. 2017, 18, 548. [Google Scholar] [CrossRef]
- Chao, M.V. Neurotrophins and their receptors: A convergence point for many signalling pathways. Nat. Rev. Neurosci. 2003, 4, 299–309. [Google Scholar] [CrossRef]
- Huang, E.J.; Reichardt, L.F. Trk Receptors: Roles in Neuronal Signal Transduction. Annu. Rev. Biochem. 2003, 72, 609–642. [Google Scholar] [CrossRef] [Green Version]
- Bermingham, D.P.; Blakely, R.D. Kinase-dependent regulation of monoamine neurotransmitter transporters. Pharmacol. Rev. 2016, 68, 888–953. [Google Scholar] [CrossRef] [Green Version]
- Angelucci, F.; Brenè, S.; Mathé, A.A. BDNF in schizophrenia, depression and corresponding animal models. Mol. Psychiatry 2005, 10, 345–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Kammen, D.P.; Peters, J.; Yao, J.; Van Kammen, W.B.; Neylan, T.; Shaw, D.; Linnoila, M. norepinephrine in acute exacerbations of chronic schizophrenia: Negative symptoms revisited. Arch. Gen. Psychiatry 1990, 47, 161–168. [Google Scholar] [CrossRef]
- Mäki-Marttunen, V.; Andreassen, O.A.; Espeseth, T. The role of norepinephrine in the pathophysiology of schizophrenia. Neurosci. Biobehav. Rev. 2020, 118, 298–314. [Google Scholar] [CrossRef]
- Kerage, D.; Sloan, E.K.; Mattarollo, S.R.; McCombe, P.A. Interaction of neurotransmitters and neurochemicals with lymphocytes. J. Neuroimmunol. 2019, 332, 99–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slota, C.; Shi, A.; Chen, G.; Bevans, M.; Weng, N. ping Norepinephrine preferentially modulates memory CD8 T cell function inducing inflammatory cytokine production and reducing proliferation in response to activation. Brain Behav. Immun. 2015, 46, 168–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takenaka, M.C.; Araujo, L.P.; Maricato, J.T.; Nascimento, V.M.; Guereschi, M.G.; Rezende, R.M.; Quintana, F.J.; Basso, A.S. Norepinephrine Controls Effector T Cell Differentiation through β 2 -Adrenergic Receptor–Mediated Inhibition of NF-κB and AP-1 in Dendritic Cells. J. Immunol. 2016, 196, 637–644. [Google Scholar] [CrossRef] [Green Version]
- Akbarian, S.; Bates, B.; Liu, R.J.; Skirboll, S.L.; Pejchal, T.; Coppola, V.; Sun, L.D.; Fan, G.; Kucera, J.; Wilson, M.A.; et al. Neurotrophin-3 modulates noradrenergic neuron function and opiate withdrawal. Mol. Psychiatry 2001, 6, 593–604. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues-Amorim, D.; Rivera-Baltanás, T.; Spuch, C.; Caruncho, H.J.; González-Fernandez, Á.; Olivares, J.M.; Agís-Balboa, R.C. Cytokines dysregulation in schizophrenia: A systematic review of psychoneuroimmune relationship. Schizophr. Res. 2017. [Google Scholar] [CrossRef]
- Khandaker, G.M.; Cousins, L.; Deakin, J.; Lennox, B.R.; Yolken, R.; Jones, P.B. Inflammation and immunity in schizophrenia: Implications for pathophysiology and treatment. Lancet Psychiatry 2015, 2, 258–270. [Google Scholar] [CrossRef] [Green Version]
- Van Kesteren, C.F.M.G.; Gremmels, H.; De Witte, L.D.; Hol, E.M.; Van Gool, A.R.; Falkai, P.G.; Kahn, R.S.; Sommer, I.E.C. Immune involvement in the pathogenesis of schizophrenia: A meta-analysis on postmortem brain studies. Transl. Psychiatry 2017, 7, e1075. [Google Scholar] [CrossRef] [PubMed]
- Debnath, M. Adaptive Immunity in schizophrenia: Functional Implications of T cells in the etiology, course and treatment. J. Neuroimmune Pharmacol. 2015, 10, 610–619. [Google Scholar] [CrossRef] [PubMed]
- Khandaker, G.M.; Dantzer, R.; Jones, P.B. Immunopsychiatry: Important facts. Psychol. Med. 2017, 47, 2229–2237. [Google Scholar] [CrossRef] [Green Version]
- Rivera-Baltanas, T.; Olivares, J.M.; Calado-Otero, M.; Kalynchuk, L.E.; Martinez-Villamarin, J.R.; Caruncho, H.J. Serotonin transporter clustering in blood lymphocytes as a putative biomarker of therapeutic efficacy in major depressive disorder. J. Affect. Disord. 2012, 137, 46–55. [Google Scholar] [CrossRef]
- Lago, S.G.; Tomasik, J.; van Rees, G.F.; Ramsey, J.M.; Haenisch, F.; Cooper, J.D.; Broek, J.A.; Suarez-Pinilla, P.; Ruland, T.; Auyeug, B.; et al. Exploring the neuropsychiatric spectrum using high-content functional analysis of single-cell signaling networks. Mol. Psychiatry 2020, 25, 2355–2372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, S.; Martínez-Cengotitabengoa, M.; Yao, Y.; Guo, Z.; Xu, Q.; Li, S.; Zhou, X.; Zhang, F. Peripheral blood nerve growth factor levels in major psychiatric disorders. J. Psychiatr. Res. 2017, 86, 39–45. [Google Scholar] [CrossRef]
- Ajami, A.; Hosseini, S.H.; Taghipour, M.; Khalilian, A. Changes in serum levels of brain derived neurotrophic factor and nerve growth factor-beta in schizophrenic patients before and after treatment. Scand. J. Immunol. 2014, 80, 36–42. [Google Scholar] [CrossRef]
- Bakirhan, A.; Sahiner, S.Y.; Sahiner, I.V.; Safak, Y.; Goka, E. Association of serum brain derived neurotropic factor with duration of drug-naive period and positive-negative symptom scores in drug naive schizophrenia. PLoS ONE 2017, 12, e0189373. [Google Scholar] [CrossRef] [Green Version]
- Blaszczyk, M.; Kurcinski, M.; Kouza, M.; Wieteska, L.; Debinski, A.; Kolinski, A.; Kmiecik, S. Modeling of protein-peptide interactions using the CABS-dock web server for binding site search and flexible docking. Methods 2016, 93, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Kroken, R.A.; Sommer, I.E.; Steen, V.M.; Dieset, I.; Johnsen, E. Constructing the immune signature of schizophrenia for clinical use and research; an integrative review translating descriptives into diagnostics. Front. Psychiatry 2019, 753, 1–16. [Google Scholar] [CrossRef]
- Vega, J.A.; García-Suárez, O.; Hannestad, J.; Pérez-Pérez, M.; Germanà, A. Neurotrophins and the immune system. J. Anat. 2003, 203, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Meredith, E.J.; Chamba, A.; Holder, M.J.; Barnes, N.M.; Gordon, J. Close encounters of the monoamine kind: Immune cells betray their nervous disposition. Immunology 2005, 115, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Ip, N.Y.; Li, Y.; Yancopoulos, G.D.; Lindsay, R.M. Cultured hippocampal neurons show responses to BDNF, NT-3, and NT-4, but not NGF. J. Neurosci. 1993, 13, 3394–3405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rangasamy, S.B.; Soderstrom, K.; Bakay, R.A.E.; Kordower, J.H. Neurotrophic factor therapy for Parkinson’s disease. Prog. Brain Res. 2010, 184, 237–264. [Google Scholar] [CrossRef] [PubMed]
- Meeker, R.B.; Williams, K.S. The p75 neurotrophin receptor: At the crossroad of neural repair and death. Neural Regen. Res. 2015, 10, 721–725. [Google Scholar] [CrossRef]
- Gómez-Pineda, V.G.; Torres-Cruz, F.M.; Vivar-Cortés, C.I.; Hernández-Echeagaray, E. Neurotrophin-3 restores synaptic plasticity in the striatum of a mouse model of Huntington’s disease. CNS Neurosci. Ther. 2018, 24, 353–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, M.A. Identification and characterization of compounds that potentiate NT-3-mediated trk receptor activity. Mol. Pharmacol. 2006, 69, 1396–1404. [Google Scholar] [CrossRef] [Green Version]
- Mandela, P.; Ordway, G.A. The norepinephrine transporter and its regulation. J. Neurochem. 2006, 97, 310–333. [Google Scholar] [CrossRef] [PubMed]
- Andersen, J.; Ringsted, K.B.; Bang-Andersen, B.; Strømgaard, K.; Kristensen, A.S. Binding site residues control inhibitor selectivity in the human norepinephrine transporter but not in the human dopamine transporter. Sci. Rep. 2015, 5, 15650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, S.J.; Kavanaugh, M.P.; Sonders, M.S.; Amara, S.G.; Zahniser, N.R. Activation of protein kinase C inhibits uptake, currents and binding associated with the human dopamine transporter expressed in Xenopus oocytes. J. Pharmacol. Exp. Ther. 1997, 282, 1358–1365. [Google Scholar] [PubMed]
- Hu, Y.F.; Caron, M.G.; Sieber-Blum, M. Norepinephrine transport-mediated gene expression in noradrenergic neurogenesis. BMC Genom. 2009, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Sieber-Blum, M.; Ren, Z. Norepinephrine transporter expression and function in noradrenergic cell differentiation. Mol. Cell. Biochem. 2000, 212, 61–70. [Google Scholar] [CrossRef]
- Poyurovsky, M.; Faragian, S.; Fuchs, C.; Pashinian, A. Effect of the selective norepinephrine reuptake inhibitor reboxetine on cognitive dysfunction in schizophrenia patients: An add-on, double-blind placebo-controlled study. Isr. J. sychiatry 2009, 46, 213–220. [Google Scholar]
- Uhl, G.R.; Johnson, P.S. Neurotransmitter transporters: Three important gene families for neuronal function. J. Exp. Biol. 1994, 196, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Choo, M.; Hwang, J.-A.; Jeon, S.W.; Oh, S.-Y.; Yoon, H.-K.; Lee, H.-J.; Kim, Y.-K. Association study between norepinephrine transporter gene polymorphism and schizophrenia in a Korean population. Psychiatry Investig. 2015, 12, 551–558. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, K.; Hornykiewicz, O. Proposal for a noradrenaline hypothesis of schizophrenia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2004, 28, 913–922. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Schizophrenia | Controls | p-Value |
|---|---|---|---|
| Total Number (N) | 54 | 54 | - |
| Age (mean ± SD) | 39.93 ± 12.29 | 44.61 ± 12.84 | 0.1310 1 |
| Gender (M/F) | 30/24 | 31/23 | 1.0000 2 |
| Neutrophil–lymphocyte ratio | 2.57 ± 1.29 | 2.81 ± 1.84 | 0.6881 3 |
| Illness onset (mean ± SD) 4 | 29.69 ± 9.70 | - | - |
| Duration of illness (mean ± SD) 4 | 10.22 ± 8.14 | - | - |
| PANSS (mean ± SD) | |||
| PANSS Positive | 20.85 ± 5.44 | - | - |
| PANSS Negative | 38.59 ± 6.67 | - | - |
| PANSS General | 36.81 ± 7.36 | - | - |
| PANSS Total | 85.89 ± 13.95 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodrigues-Amorim, D.; Iglesias-Martínez-Almeida, M.; Rivera-Baltanás, T.; Fernández-Palleiro, P.; Freiría-Martínez, L.; Rodríguez-Jamardo, C.; Comís-Tuche, M.; Vallejo-Curto, M.d.C.; Álvarez-Ariza, M.; López-García, M.; et al. The Role of the Second Extracellular Loop of Norepinephrine Transporter, Neurotrophin-3 and Tropomyosin Receptor Kinase C in T Cells: A Peripheral Biomarker in the Etiology of Schizophrenia. Int. J. Mol. Sci. 2021, 22, 8499. https://doi.org/10.3390/ijms22168499
Rodrigues-Amorim D, Iglesias-Martínez-Almeida M, Rivera-Baltanás T, Fernández-Palleiro P, Freiría-Martínez L, Rodríguez-Jamardo C, Comís-Tuche M, Vallejo-Curto MdC, Álvarez-Ariza M, López-García M, et al. The Role of the Second Extracellular Loop of Norepinephrine Transporter, Neurotrophin-3 and Tropomyosin Receptor Kinase C in T Cells: A Peripheral Biomarker in the Etiology of Schizophrenia. International Journal of Molecular Sciences. 2021; 22(16):8499. https://doi.org/10.3390/ijms22168499
Chicago/Turabian StyleRodrigues-Amorim, Daniela, Marta Iglesias-Martínez-Almeida, Tania Rivera-Baltanás, Patricia Fernández-Palleiro, Luis Freiría-Martínez, Cynthia Rodríguez-Jamardo, María Comís-Tuche, María del Carmen Vallejo-Curto, María Álvarez-Ariza, Marta López-García, and et al. 2021. "The Role of the Second Extracellular Loop of Norepinephrine Transporter, Neurotrophin-3 and Tropomyosin Receptor Kinase C in T Cells: A Peripheral Biomarker in the Etiology of Schizophrenia" International Journal of Molecular Sciences 22, no. 16: 8499. https://doi.org/10.3390/ijms22168499
APA StyleRodrigues-Amorim, D., Iglesias-Martínez-Almeida, M., Rivera-Baltanás, T., Fernández-Palleiro, P., Freiría-Martínez, L., Rodríguez-Jamardo, C., Comís-Tuche, M., Vallejo-Curto, M. d. C., Álvarez-Ariza, M., López-García, M., de las Heras, E., García-Caballero, A., Olivares, J. M., & Spuch, C. (2021). The Role of the Second Extracellular Loop of Norepinephrine Transporter, Neurotrophin-3 and Tropomyosin Receptor Kinase C in T Cells: A Peripheral Biomarker in the Etiology of Schizophrenia. International Journal of Molecular Sciences, 22(16), 8499. https://doi.org/10.3390/ijms22168499