
Trace Amine-Associated Receptor 1 Contributes to Diverse Functional Actions of O-Phenyl-Iodotyramine in Mice but Not to the Effects of Monoamine-Based Antidepressants

 ,
, 
Abstract
:1. Introduction
2. Results
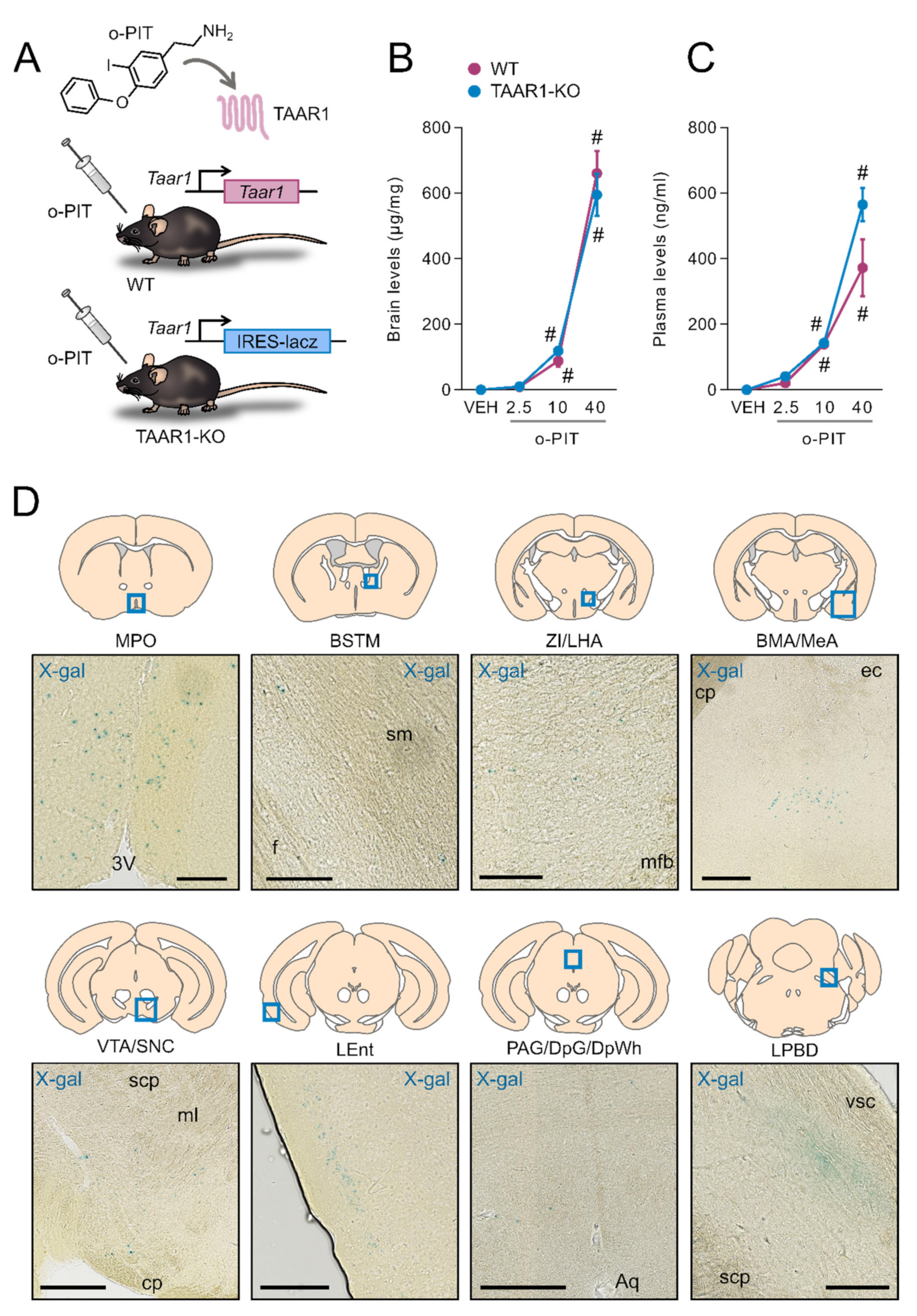
2.1. Exposure to O-PIT and Histoenzymological Detection of β-Galactosidase in TAAR1-KO Mice
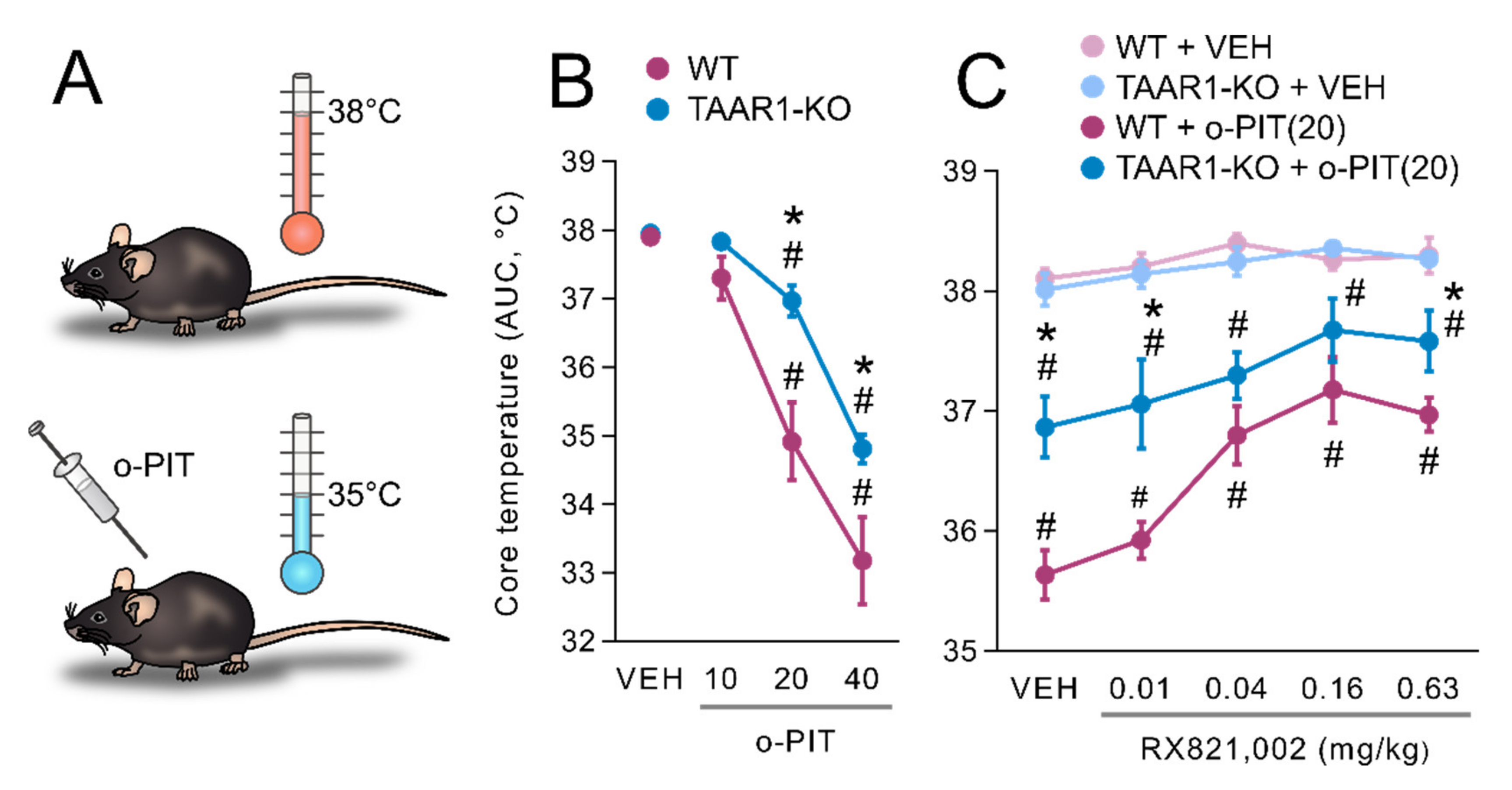
2.2. Influence of O-PIT on Core Temperature in TAAR1-KO
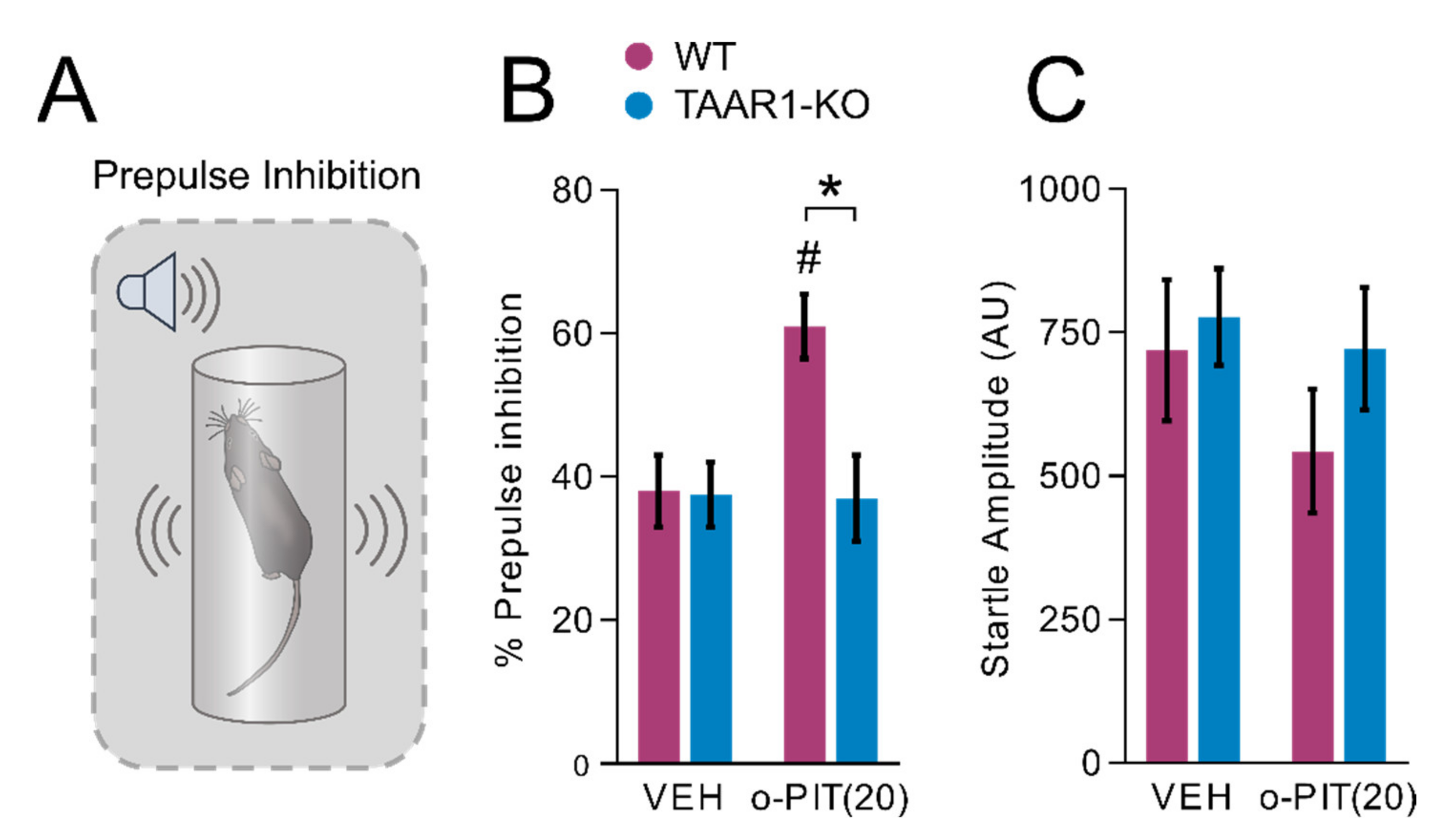
2.3. Influence of O-PIT on Prepulse Inhibition in TAAR1-KO Mice
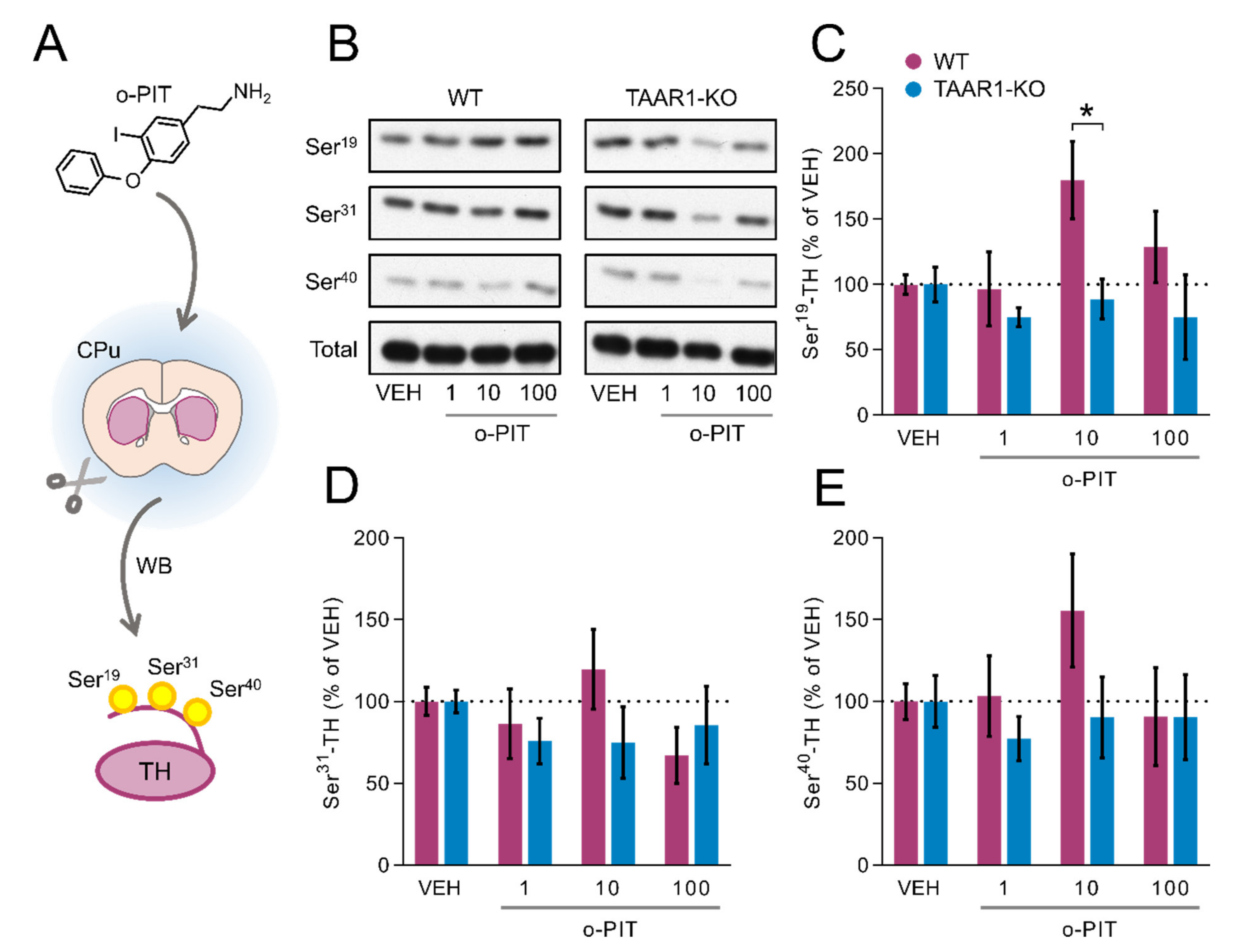
2.4. Induction by O-PIT of Tyrosine Hydroxylase Phosphorylation (TH) and Activity in the Caudoputamen (CPu)
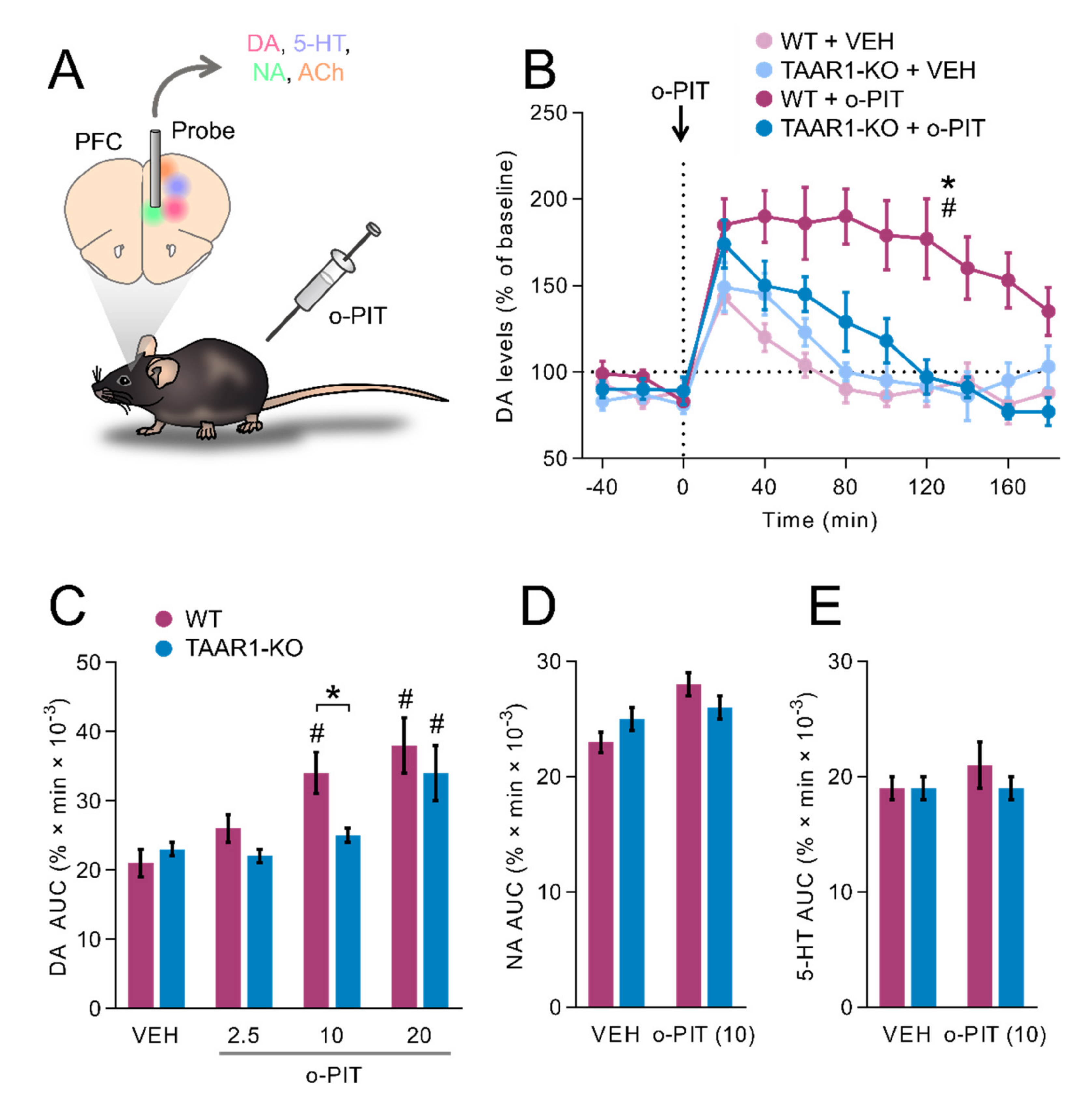
2.5. Influence of O-PIT upon Dialysis Levels of DA, NA and 5-HT in the mPFC of Freely Moving TAAR1-KO Mice
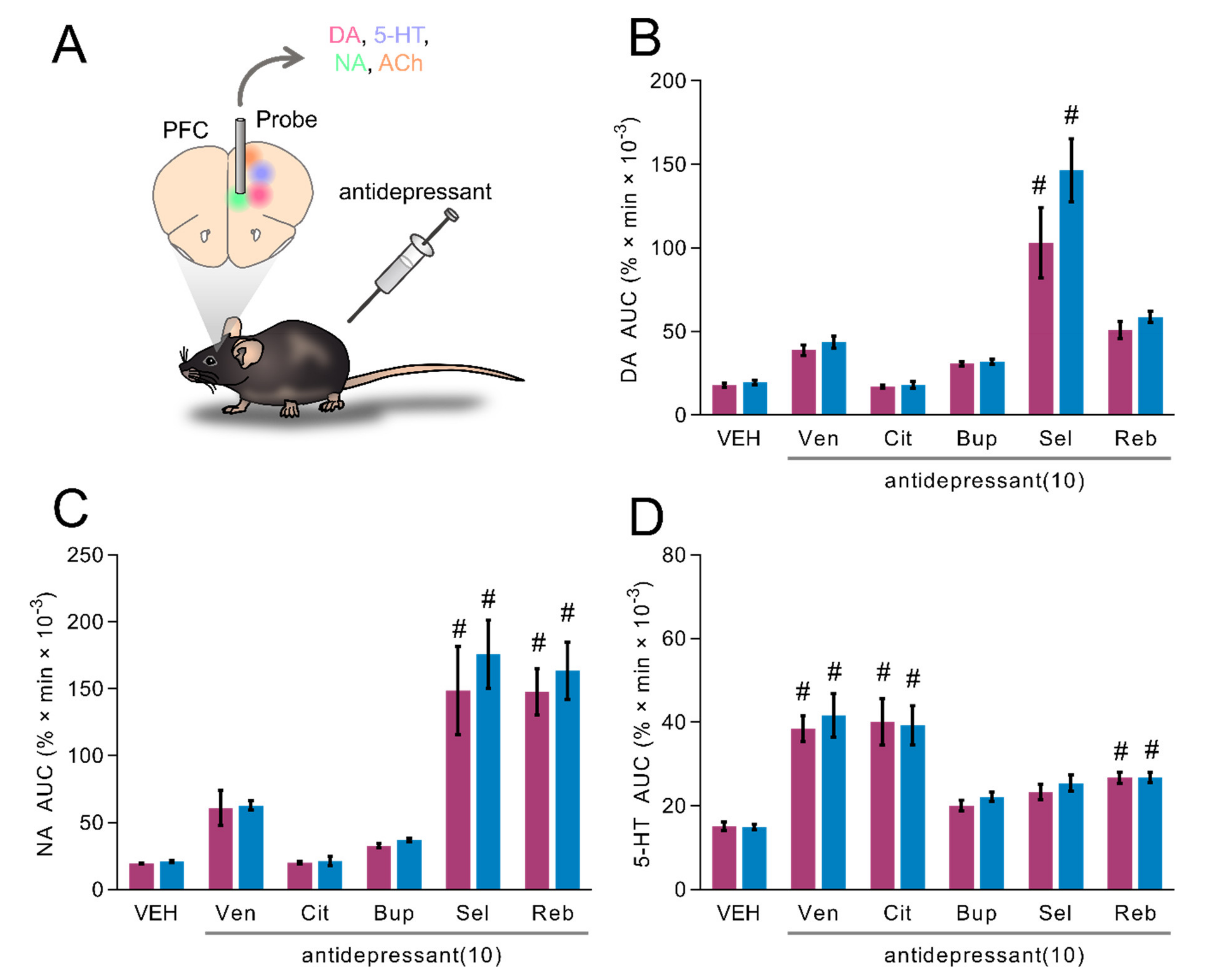
2.6. Influence of Classical Antidepressants upon Dialysis Levels of DA, NA and 5-HT in the mPFC of Freely Moving TAAR1-KO Mice
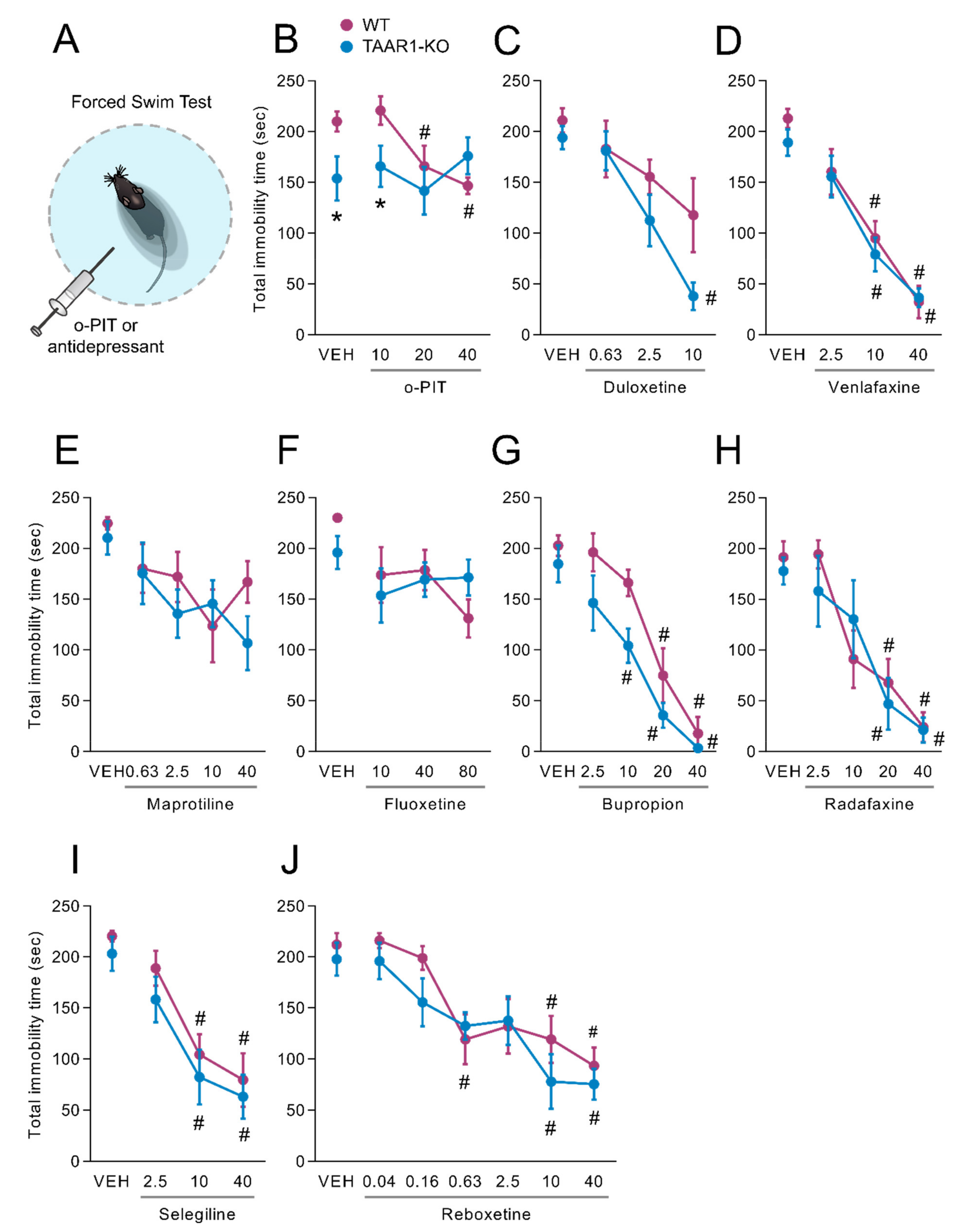
2.7. Influence of O-PIT and Commonly Used Antidepressants on the Forced-Swim Test in TAAR1-KO Mice
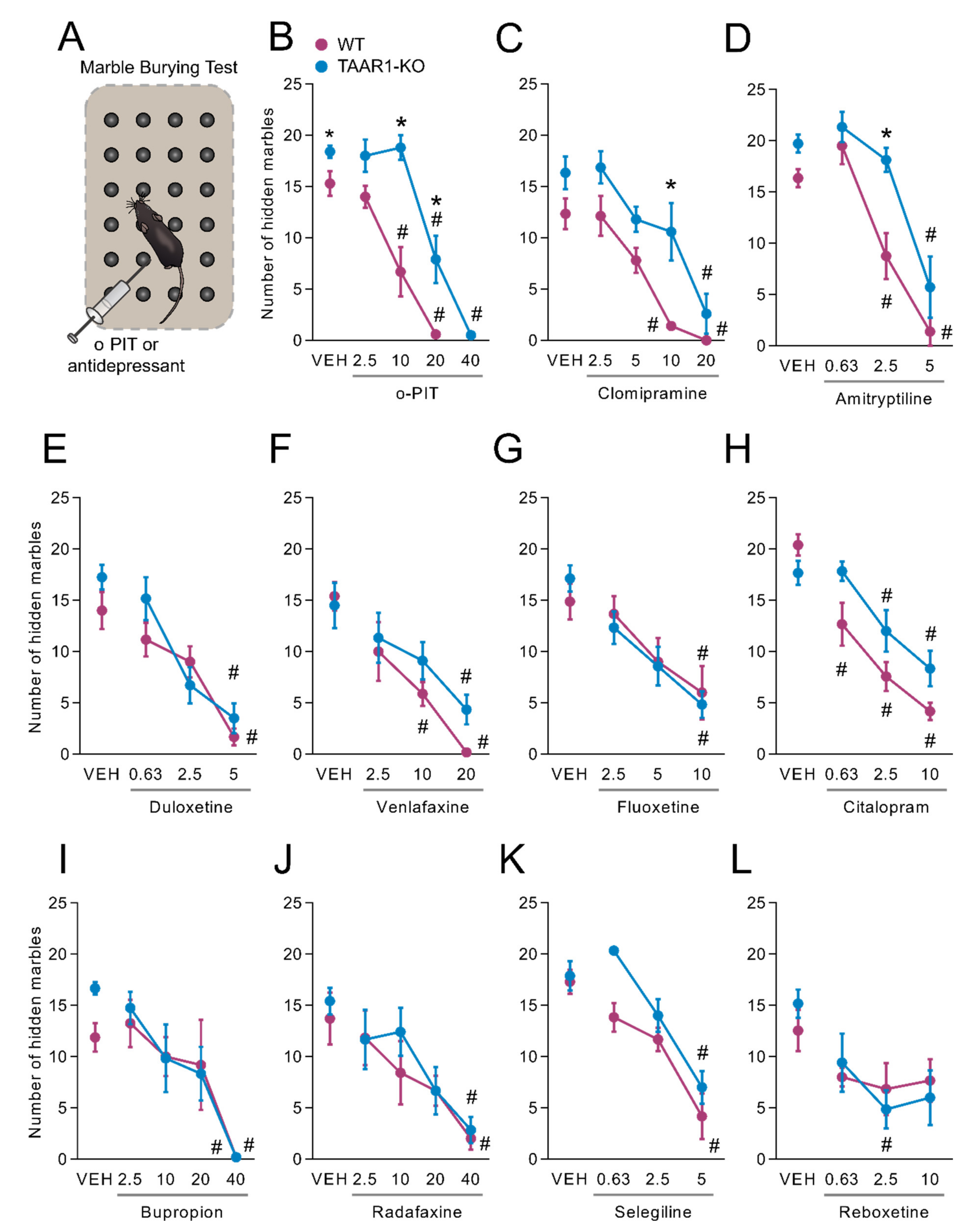
2.8. Influence of O-PIT and Commonly Used Antidepressants on Marble-Burying Test in TAAR1-KO Mice
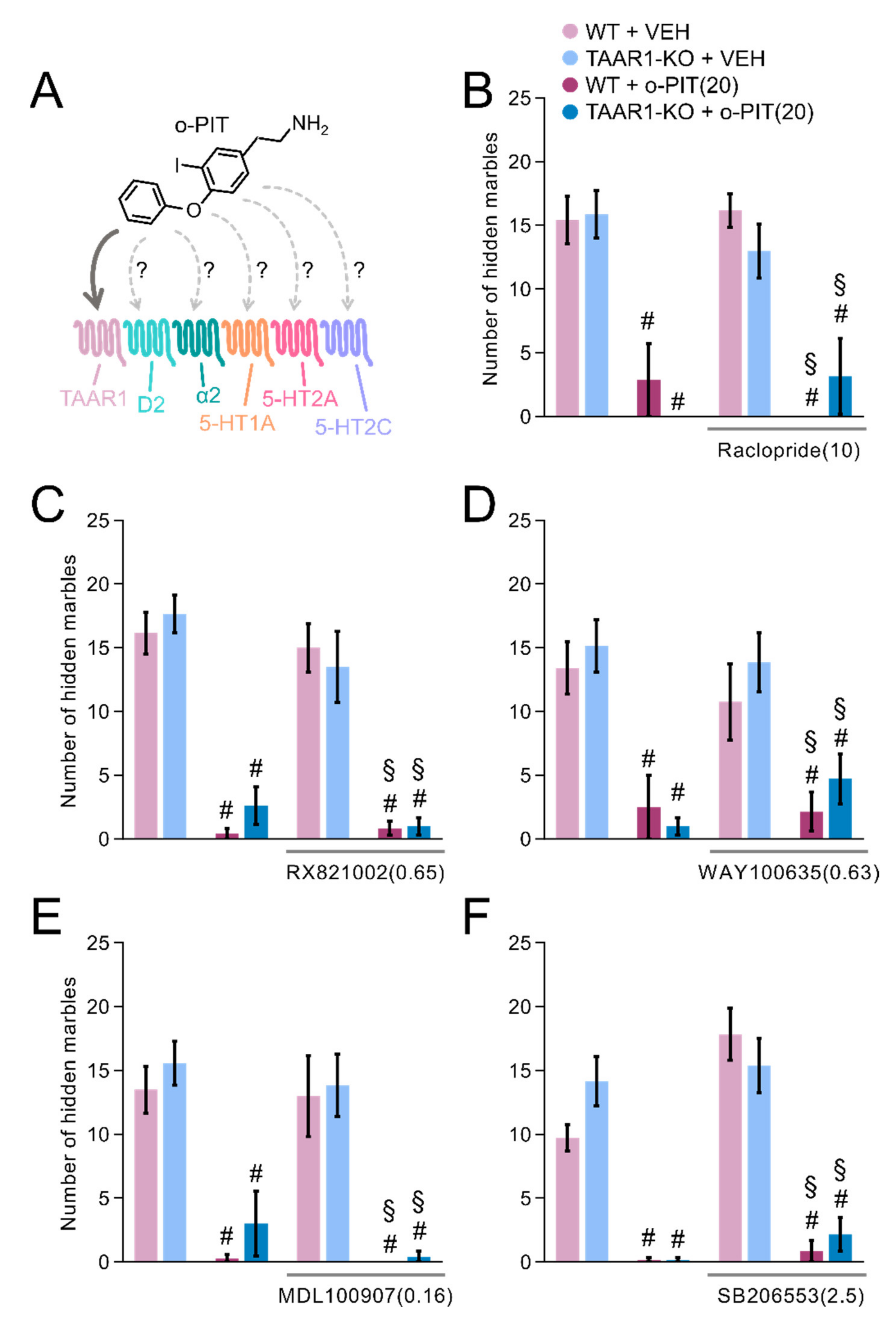
2.9. Influence of O-PIT and Monoaminergic Receptor Antagonists on Marble-Burying Test in TAAR1-KO Mice
3. Discussion
3.1. Hypothermia Elicited by O-PIT Is Partially Mediated by TAAR1
3.2. O-PIT Enhancement of Prepulse Inhibition Is TAAR1-Dependent
3.3. O-PIT Increases TH Phosphorylation through TAAR1 in the Striatum
3.4. O-PIT Stimulates DA Release in mPFC through TAAR1
3.5. A Differential Role of TAAR1 in the Antidepressant Actions of O-PIT vs. Clinically used Classes of Antidepressants
4. Conclusions
5. Material and Methods
5.1. Animals
5.2. Drugs
5.3. Plasma and Brain Levels of O-PIT
5.4. Histoenzymological Detection of β-Galactosidase
5.5. Core Temperature Measurement
5.6. Acoustic Startle and Prepulse Inhibition
5.7. Tyrosine Hydroxylase Phosphorylation in Brain Slices
5.8. Microdialysis in Freely Moving Mice
5.9. Forced-Swim Test
5.10. Marble-Burying Behaviour
5.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gainetdinov, R.R.; Hoener, M.C.; Berry, M.D. Trace amines and their receptors. Pharmacol. Rev. 2018, 70, 549–620. [Google Scholar] [CrossRef] [Green Version]
- Pretorius, L.; Smith, C. The trace aminergic system: A gender-sensitive therapeutic target for IBS? J. Biomed. Sci. 2020, 27, 95. [Google Scholar] [CrossRef]
- Zucchi, R.; Chiellini, G.; Scanlan, T.S.; Grandy, D.K. Trace amine-associated receptors and their ligands. Br. J. Pharmacol. 2006, 149, 967–978. [Google Scholar] [CrossRef] [Green Version]
- Accorroni, A.; Rutigliano, G.; Sabatini, M.; Frascarelli, S.; Borsò, M.; Novelli, E.; Bandini, L.; Ghelardoni, S.; Saba, A.; Zucchi, R.; et al. Exogenous 3-iodothyronamine rescues the entorhinal cortex from β-amyloid toxicity. Thyroid 2020, 30, 147–160. [Google Scholar] [CrossRef] [PubMed]
- Rutigliano, G.; Zucchi, R. Molecular variants in human trace amine-associated receptors and their implications in mental and metabolic disorders. Cell. Mol. Neurobiol. 2020, 40, 239–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Köhrle, J.; Biebermann, H. 3-Iodothyronamine-A thyroid hormone metabolite with distinct target profiles and mode of action. Endocr. Rev. 2019, 40, 620–630. [Google Scholar] [CrossRef] [PubMed]
- Berry, M.D. Mammalian central nervous system trace amines. Pharmacologic amphetamines, physiologic neuromodulators. J. Neurochem. 2004, 90, 257–271. [Google Scholar] [CrossRef] [PubMed]
- Van Nguyen, T.; Juorio, A.V. Down-regulation of tryptamine binding sitess following chronic molindone administration: A comparison with responses of dopamine and 5-hydroxytryptamine receptors. Naunyn Schmiedebergs Arch. Pharmacol. 1989, 340, 366–371. [Google Scholar] [CrossRef]
- Borowsky, B.; Adham, N.; Jones, K.A.; Raddatz, R.; Artymyshyn, R.; Ogozalek, K.L.; Durkin, M.M.; Lakhlani, P.P.; Bonini, J.A.; Pathirana, S.; et al. Trace amines: Identification of a family of mammalian G protein-coupled receptors. Proc. Natl. Acad. Sci. USA 2001, 98, 8966–8971. [Google Scholar] [CrossRef] [Green Version]
- Bunzow, J.R.; Sonders, M.S.; Arttamangkul, S.; Harrison, L.M.; Zhang, G.; Quigley, D.I.; Darland, T.; Suchland, K.L.; Pasumamula, S.; Kennedy, J.L.; et al. Amphetamine, 3,4-methylenedioxymethamphetamine, lysergic acid diethylamide, and metabolites of the catecholamine neurotransmitters are agonists of a rat trace amine receptor. Mol. Pharmacol. 2001, 60, 1181–1188. [Google Scholar] [CrossRef]
- Maguire, J.J.; Parker, W.A.E.; Foord, S.M.; Bonner, T.I.; Neubig, R.R.; Davenport, A.P. International union of pharmacology. LXXII. Recommendations for trace amine receptor nomenclature. Pharmacol. Rev. 2009, 61, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liberles, S.D. Trace amine-associated receptors: Ligands, neural circuits, and behaviors. Curr. Opin. Neurobiol. 2015, 34, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutigliano, G.; Accorroni, A.; Zucchi, R. The case for TAAR1 as a modulator of central nervous system function. Front. Pharmacol. 2018, 8, 987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berry, M.D.; Gainetdinov, R.R.; Hoener, M.C.; Shahid, M. Pharmacology of human trace amine-associated receptors: Therapeutic opportunities and challenges. Pharmacol. Ther. 2017, 180, 161–180. [Google Scholar] [CrossRef] [PubMed]
- Lindemann, L.; Meyer, C.A.; Jeanneau, K.; Bradaia, A.; Ozmen, L.; Bluethmann, H.; Bettler, B.; Wettstein, J.G.; Borroni, E.; Moreau, J.L.; et al. Trace amine-associated receptor 1 modulates dopaminergic activity. J. Pharmacol. Exp. Ther. 2008, 324, 948–956. [Google Scholar] [CrossRef] [Green Version]
- Mantas, I.; Vallianatou, T.; Yang, Y.; Shariatgorji, M.; Kalomoiri, M.; Fridjonsdottir, E.; Millan, M.J.; Zhang, X.; Andrén, P.E.; Svenningsson, P. TAAR1-dependent and -independent actions of tyramine in interaction with glutamate underlie central Effects of monoamine oxidase inhibition. Biol. Psychiatry 2021, 90, 16–27. [Google Scholar] [CrossRef]
- Underhill, S.; Amara, S. MDMA and TAAR1-mediated RhoA activation in serotonin neurons. FASEB J. 2020, 34, 1. [Google Scholar] [CrossRef]
- Di Cara, B.; Maggio, R.; Aloisi, G.; Rivet, J.M.; Lundius, E.G.; Yoshitake, T.; Svenningsson, P.; Brocco, M.; Gobert, A.; de Groote, L.; et al. Genetic deletion of trace amine 1 receptors reveals their role in auto-inhibiting the actions of ecstasy (MDMA). J. Neurosci. 2011, 31, 16928–16940. [Google Scholar] [CrossRef]
- Underhill, S.M.; Hullihen, P.D.; Chen, J.; Fenollar-Ferrer, C.; Rizzo, M.A.; Ingram, S.L.; Amara, S.G. Amphetamines signal through intracellular TAAR1 receptors coupled to Gα13 and GαS in discrete subcellular domains. Mol. Psychiatry 2019, 26, 1208–1223. [Google Scholar] [CrossRef] [Green Version]
- Bradaia, A.; Trube, G.; Stalder, H.; Norcross, R.D.; Ozmen, L.; Wettstein, J.G.; Pinard, A.; Buchy, D.; Gassmann, M.; Hoener, M.C.; et al. The selective antagonist EPPTB reveals TAAR1-mediated regulatory mechanisms in dopaminergic neurons of the mesolimbic system. Proc. Natl. Acad. Sci. USA 2009, 106, 20081–20086. [Google Scholar] [CrossRef] [Green Version]
- Revel, F.G.; Moreau, J.L.; Gainetdinov, R.R.; Bradaia, A.; Sotnikova, T.D.; Mory, R.; Durkin, S.; Zbinden, K.G.; Norcross, R.; Meyer, C.A.; et al. TAAR1 activation modulates monoaminergic neurotransmission, preventing hyperdopaminergic and hypoglutamatergic activity. Proc. Natl. Acad. Sci. USA 2011, 108, 8485–8490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolinsky, T.D.; Swanson, C.J.; Smith, K.E.; Zhong, H.; Borowsky, B.; Seeman, P.; Branchek, T.; Gerald, C.P. The Trace Amine 1 receptor knockout mouse: An animal model with relevance to schizophrenia. Genes Brain Behav. 2007, 6, 628–639. [Google Scholar] [CrossRef]
- Revel, F.G.; Moreau, J.L.; Pouzet, B.; Mory, R.; Bradaia, A.; Buchy, D.; Metzler, V.; Chaboz, S.; Groebke Zbinden, K.; Galley, G.; et al. A new perspective for schizophrenia: TAAR1 agonists reveal antipsychotic- and antidepressant-like activity, improve cognition and control body weight. Mol. Psychiatry 2013, 18, 543–556. [Google Scholar] [CrossRef] [PubMed]
- Dedic, N.; Jones, P.G.; Hopkins, S.C.; Lew, R.; Shao, L.; Campbell, J.E.; Spear, K.L.; Large, T.H.; Campbell, U.C.; Hanania, T.; et al. SEP-363856, a novel psychotropic agent with a unique, non-D2 receptor mechanism of action. J. Pharmacol. Exp. Ther. 2019, 371, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Tonelli, M.; Cichero, E. Trace amine associated receptor 1 (TAAR1) modulators: A patent review (2010-present). Expert Opin. Ther. Pat. 2020, 30, 137–145. [Google Scholar] [CrossRef]
- Koblan, K.S.; Kent, J.; Hopkins, S.C.; Krystal, J.H.; Cheng, H.; Goldman, R.; Loebel, A. A Non-D2-Receptor-Bindingdrug for the treatment of schizophrenia. New Engl. J. Med. 2020, 382, 1497–1506. [Google Scholar] [CrossRef]
- Correll, C.U.; Koblan, K.S.; Hopkins, S.C.; Kent, J.; Cheng, H.; Goldman, R.; Loebel, A. Safety and effectiveness of SEP-363856 in schizophrenia: Results of a 6-month, open-label extension study. CNS Spectr. 2021, 26, 148–149. [Google Scholar] [CrossRef]
- Sabelli, H.C.; Mosnaim, A.D. Phenylethylamine hypothesis of affective behavior. Am. J. Psychiatry 1974, 131, 695–699. [Google Scholar] [CrossRef]
- Davis, B.A.; Boulton, A.A. The trace amines and their acidic metabolites in depression—An overview. Prog. Neuropsychopharmacol. Biol. Psychiatry 1994, 18, 17–45. [Google Scholar] [CrossRef]
- Holschneider, D.P.; Chen, K.; Seif, I.; Shih, J.C. Biochemical, behavioral, physiologic, and neurodevelopmental changes in mice deficient in monoamine oxidase A or B. Brain Res. Bull. 2001, 56, 453–462. [Google Scholar] [CrossRef] [Green Version]
- Espinoza, S.; Sukhanov, I.; Efimova, E.V.; Kozlova, A.; Antonova, K.A.; Illiano, P.; Leo, D.; Merkulyeva, N.; Kalinina, D.; Musienko, P.; et al. Trace amine-associated receptor 5 provides olfactory input into limbic brain areas and modulates emotional behaviors and serotonin transmission. Front. Mol. Neurosci. 2020, 13, 18. [Google Scholar] [CrossRef] [PubMed]
- Hackenmueller, S.A.; Marchini, M.; Saba, A.; Zucchi, R.; Scanlan, T.S. Biosynthesis of 3-Iodothyronamine (T1AM) is dependent on the sodium-iodide symporter and thyroperoxidase but does not involve extrathyroidal metabolism of T4. Endocrinology 2012, 153, 5659–5667. [Google Scholar] [CrossRef] [Green Version]
- Hoefig, C.S.; Renko, K.; Piehl, S.; Scanlan, T.S.; Bertoldi, M.; Opladen, T.; Hoffmann, G.F.; Klein, J.; Blankenstein, O.; Schweizer, U.; et al. Does the aromatic l-amino acid decarboxylase contribute to thyronamine biosynthesis? Mol. Cell. Endocrinol. 2012, 349, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Chiellini, G.; Bellusci, L.; Sabatini, M.; Zucchi, R. Thyronamines and analogues-the route from rediscovery to translational research on thyronergic amines. Mol. Cell. Endocrinol. 2017, 458, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Piehl, S.; Hoefig, C.S.; Scanlan, T.S.; Köhrle, J. Thyronamines—Past, present, and future. Endocr. Rev. 2011, 32, 64–80. [Google Scholar] [CrossRef] [Green Version]
- Rutigliano, G.; Bandini, L.; Sestito, S.; Chiellini, G. 3-iodothyronamine and derivatives: New allies against metabolic syndrome? Int. J. Mol. Sci. 2020, 21, 2005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurino, A.; Landucci, E.; Raimondi, L. Central effects of 3-Iodothyronamine reveal a novel role for mitochondrial monoamine oxidases. Front. Endocrinol. 2018, 9, 6. [Google Scholar] [CrossRef]
- Zhang, X.; Mantas, I.; Alvarsson, A.; Yoshitake, T.; Shariatgorji, M.; Pereira, M.; Nilsson, A.; Kehr, J.; Andrén, P.E.; Millan, M.J.; et al. Striatal tyrosine hydroxylase is stimulated via TAAR1 by 3-iodothyronamine, but not by tyramine or β-phenylethylamine. Front. Pharmacol. 2018, 9, 166. [Google Scholar] [CrossRef] [Green Version]
- Hart, M.E.; Suchland, K.L.; Miyakawa, M.; Bunzow, J.R.; Grandy, D.K.; Scanlan, T.S. Trace amine-associated receptor agonists: Synthesis and evaluation of thyronamines and related analogues. J. Med. Chem. 2006, 49, 1101–1112. [Google Scholar] [CrossRef]
- Scanlan, T.S.; Suchland, K.L.; Hart, M.E.; Chiellini, G.; Huang, Y.; Kruzich, P.J.; Frascarelli, S.; Crossley, D.A.; Bunzow, J.R.; Ronca-Testoni, S.; et al. 3-Iodothyronamine is an endogenous and rapid-acting derivative of thyroid hormone. Nat. Med. 2004, 10, 638–642. [Google Scholar] [CrossRef]
- Gachkar, S.; Oelkrug, R.; Martinez-Sanchez, N.; Rial-Pensado, E.; Warner, A.; Hoefig, C.S.; López, M.; Mittag, J. 3-Iodothyronamine induces tail vasodilation through central action in male mice. Endocrinology 2017, 158, 1977–1984. [Google Scholar] [CrossRef] [PubMed]
- Boulant, J.A. Role of the preoptic-anterior hypothalamus in thermoregulation and fever. Clin. Infect. Dis. 2000, 31, S157–S161. [Google Scholar] [CrossRef]
- Szymusiak, R.; Satinoff, E. Acute thermoregulatory effects of unilateral electrolytic lesions of the medial and lateral preoptic area in rats. Physiol. Behav. 1982, 28, 161–170. [Google Scholar] [CrossRef]
- Tan, C.L.; Cooke, E.K.; Leib, D.E.; Lin, Y.C.; Daly, G.E.; Zimmerman, C.A.; Knight, Z.A. Warm-sensitive neurons that control body temperature. Cell 2016, 167, 47–59.e15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panas, H.N.; Lynch, L.J.; Vallender, E.J.; Xie, Z.; Chen, G.; Lynn, S.K.; Scanlan, T.S.; Miller, G.M. Normal thermoregulatory responses to 3-iodothyronamine, trace amines and amphetamine-like psychostimulants in trace amine associated receptor 1 knockout mice. J. Neurosci. Res. 2010, 88, 1962–1969. [Google Scholar] [CrossRef] [Green Version]
- Feleder, C.; Perlik, V.; Blatteis, C.M. Preoptic α1- and α2-noradrenergic agonists induce, respectively, PGE2 -independent and PGE2-dependent hyperthermic responses in guinea pigs. Am. J. Physiol. Integr. Comp. Physiol. 2004, 286, R1156–R1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, T.D.; Moffett, S.X.; Scanlan, T.S.; Martin, J.V. Effects of acute microinjections of the thyroid hormone derivative 3-iodothyronamine to the preoptic region of adult male rats on sleep, thermoregulation and motor activity. Horm. Behav. 2013, 64, 81–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wadenberg, M.L.G.; Soliman, A.; VanderSpek, S.C.; Kapur, S. Dopamine D2 receptor occupancy is a common mechanism underlying animal models of antipsychotics and their clinical effects. Neuropsychopharmacology 2001, 25, 633–641. [Google Scholar] [CrossRef]
- Creese, I.; Burt, D.R.; Snyder, S.H. Dopamine receptor binding predicts clinical and pharmacological potencies of antischizophrenic drugs. Science 1976, 192, 481–483. [Google Scholar] [CrossRef]
- Seeman, P.; Lee, T.; Chau-Wong, M.; Wong, K. Antipsychoticdrug doses and neuroleptic/dopamine receptors. Nature 1976, 261, 717–719. [Google Scholar] [CrossRef]
- Kumari, V.; Soni, W.; Mathew, V.M.; Sharma, T. Prepulse inhibition of the startle response in men with schizophrenia: Effects of age of onset of illness, symptoms, and medication. Arch. Gen. Psychiatry 2000, 57, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Goto, K.; Ueki, A.; Iso, H.; Morita, Y. Reduced prepulse inhibition in rats with entorhinal cortex lesions. Behav. Brain Res. 2002, 134, 201–207. [Google Scholar] [CrossRef]
- Goto, K.; Ueki, A.; Iso, H.; Morita, Y. Involvement of nucleus accumbens dopaminergic transmission in acoustic startle: Observations concerning prepulse inhibition in rats with entorhinal cortex lesions. Psychiatry Clin. Neurosci. 2004, 58, 441–445. [Google Scholar] [CrossRef] [PubMed]
- Tozzi, F.; Rutigliano, G.; Borsò, M.; Falcicchia, C.; Zucchi, R.; Origlia, N. T1AM-TAAR1 signalling protects against OGD-induced synaptic dysfunction in the entorhinal cortex. Neurobiol. Dis. 2021, 151, 105271. [Google Scholar] [CrossRef] [PubMed]
- Dunkley, P.R.; Bobrovskaya, L.; Graham, M.E.; Von Nagy-Felsobuki, E.I.; Dickson, P.W. Tyrosine hydroxylase phosphorylation: Regulation and consequences. J. Neurochem. 2004, 91, 1025–1043. [Google Scholar] [CrossRef] [PubMed]
- Parker, E.M.; Cubeddu, L.X. Comparative effects of amphetamine, phenylethylamine and related drugs on dopamine efflux, dopamine uptake and mazindol binding. J. Pharmacol. Exp. Ther. 1988, 245, 199–210. [Google Scholar] [PubMed]
- Sulzer, D.; Rayport, S. Amphetamine and other psychostimulants reduce pH gradients in midbrain dopaminergic neurons and chromaffin granules: A mechanism of action. Neuron 1990, 5, 797–808. [Google Scholar] [CrossRef]
- Borison, R.L.; Mosnaim, A.D.; Sabelli, H.C. Brain 2-phenylethylamine as a major mediator for the central actions of amphetamine and methylphenidate. Life Sci. 1975, 17, 1331–1343. [Google Scholar] [CrossRef]
- Krishnan, V.; Nestler, E.J. The molecular neurobiology of depression. Nature 2008, 455, 894–902. [Google Scholar] [CrossRef]
- Millan, M.J.; Di Cara, B.; Dekeyne, A.; Panayi, F.; De Groote, L.; Sicard, D.; Cistarelli, L.; Billiras, R.; Gobert, A. Selective blockade of dopamine D3 versus D2 receptors enhances frontocortical cholinergic transmission and social memory in rats: A parallel neurochemical and behavioural analysis. J. Neurochem. 2007, 100, 1047–1061. [Google Scholar] [CrossRef]
- Belujon, P.; Grace, A.A. Dopamine system dysregulation in major depressive disorders. Int. J. Neuropsychopharmacol. 2017, 20, 1036–1046. [Google Scholar] [CrossRef] [Green Version]
- Millan, M.J.; Audinot, V.; Rivet, J.M.; Gobert, A.; Vian, J.; Prost, J.F.; Spedding, M.; Peglion, J.L. S 14297, a novel selective ligand at cloned human dopamine D3 receptors, blocks 7-OH-DPAT-induced hypothermia in rats. Eur. J. Pharmacol. 1994, 260, R3–R5. [Google Scholar] [CrossRef]
- Groenink, L.; Dirks, A.; Verdouw, P.M.; de Graaff, M.; Peeters, B.W.; Millan, M.J.; Olivier, B. CRF1 Not glucocorticoid receptors mediate prepulse inhibition deficits in mice overexpressing CRF. Biol. Psychiatry 2008, 63, 360–368. [Google Scholar] [CrossRef]
- Gobert, A.; Di Cara, B.; Cistarelli, L.; Millan, M.J. Piribedil enhances frontocortical and hippocampal release of acetylcholine in freely moving rats by blockade of α2A-adrenoceptors: A dialysis comparison to talipexole and quinelorane in the absence of acetylcholinesterase inhibitors. J. Pharmacol. Exp. Ther. 2003, 305, 338–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millan, M.J.; Dekeyne, A.; Papp, M.; Drieu La Rochelle, C.; Macsweeny, C.; Peglion, J.L.; Brocco, M. S33005, a novel ligand at both serotonin and norepinephrine transporters: II. Behavioral profile in comparison with venlafaxine, reboxetine, citalopram, and clomipramine. J. Pharmacol. Exp. Ther. 2001, 298, 581–591. [Google Scholar] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Resting Levels | o-PIT (10 mg/kg i.p.) [AUC; % of Vehicle] | ||||
|---|---|---|---|---|---|
| (Units) | WT | TAAR1-KO | WT | TAAR1-KO | |
| DA | (pM) | 238.87 ± 30.13 | 203.49 ± 23.84 | 160 ± 13 | 109 ± 6 * |
| NA | (pM) | 445.49 ± 25.79 | 411.87 ± 22.47 | 115 ± 5 | 106 ± 4 |
| 5-HT | (pM) | 188.29 ± 15.19 | 209.08 ± 22.40 | 109 ± 8 | 102 ± 4 |
| Acetylcholine | (nM) | 0.92 ± 0.10 | 1.13 ± 0.14 | 98 ± 7 | 107 ± 20 |
| Alanine | (µM) | 0.90 ± 0.12 | 1.18 ± 0.21 | 94 ± 3 | 84 ± 4 |
| Arginine | (µM) | 0.38 ± 0.07 | 0.48 ± 0.10 | 89 ± 2 | 91 ± 4 |
| Aspartate | (µM) | 70.83 ± 20.45 | 69.98 ± 20.50 | 103 ± 5 | 105 ± 6 |
| GABA | (µM) | 10.74 ± 1.82 | 13.24 ± 3.19 | 87 ± 8 | 82 ± 8 |
| Glutamate | (µM) | 2.05 ± 0.71 | 1.64 ± 0.83 | 106 ± 3 | 101 ± 3 |
| Glutamine | (µM) | 9.55 ± 2.26 | 13.61 ± 3.73 | 106 ± 5 | 101 ± 5 |
| Glycine | (µM) | 1.08 ± 0.10 | 1.53 ± 0.29 | 93 ± 2 | 90 ± 2 |
| Phenethylamine | (µM) | 0.40 ± 0.08 | 0.32 ± 0.09 | 108 ± 3 | 114 ± 6 |
| L-serine | (µM) | 1.31 ± 0.18 | 1.58 ± 0.25 | 96 ± 2 | 96 ± 5 |
| D-serine | (µM) | 0.45 ± 0.09 | 0.35 ± 0.06 | 95 ± 4 | 107 ± 4 |
| Taurine | (µM) | 6.92 ± 1.64 | 6.99 ± 3.20 | 116 ± 6 | 105 ± 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mantas, I.; Millan, M.J.; Di Cara, B.; Groenink, L.; Veiga, S.; Cistarelli, L.; Brocco, M.; Bertrand, M.; Svenningsson, P.; Zhang, X. Trace Amine-Associated Receptor 1 Contributes to Diverse Functional Actions of O-Phenyl-Iodotyramine in Mice but Not to the Effects of Monoamine-Based Antidepressants. Int. J. Mol. Sci. 2021, 22, 8907. https://doi.org/10.3390/ijms22168907
Mantas I, Millan MJ, Di Cara B, Groenink L, Veiga S, Cistarelli L, Brocco M, Bertrand M, Svenningsson P, Zhang X. Trace Amine-Associated Receptor 1 Contributes to Diverse Functional Actions of O-Phenyl-Iodotyramine in Mice but Not to the Effects of Monoamine-Based Antidepressants. International Journal of Molecular Sciences. 2021; 22(16):8907. https://doi.org/10.3390/ijms22168907
Chicago/Turabian StyleMantas, Ioannis, Mark J. Millan, Benjamin Di Cara, Lucianne Groenink, Sylvie Veiga, Laetitia Cistarelli, Mauricette Brocco, Marc Bertrand, Per Svenningsson, and Xiaoqun Zhang. 2021. "Trace Amine-Associated Receptor 1 Contributes to Diverse Functional Actions of O-Phenyl-Iodotyramine in Mice but Not to the Effects of Monoamine-Based Antidepressants" International Journal of Molecular Sciences 22, no. 16: 8907. https://doi.org/10.3390/ijms22168907
APA StyleMantas, I., Millan, M. J., Di Cara, B., Groenink, L., Veiga, S., Cistarelli, L., Brocco, M., Bertrand, M., Svenningsson, P., & Zhang, X. (2021). Trace Amine-Associated Receptor 1 Contributes to Diverse Functional Actions of O-Phenyl-Iodotyramine in Mice but Not to the Effects of Monoamine-Based Antidepressants. International Journal of Molecular Sciences, 22(16), 8907. https://doi.org/10.3390/ijms22168907