
Targeting Cytokines, Pathogen-Associated Molecular Patterns, and Damage-Associated Molecular Patterns in Sepsis via Blood Purification


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
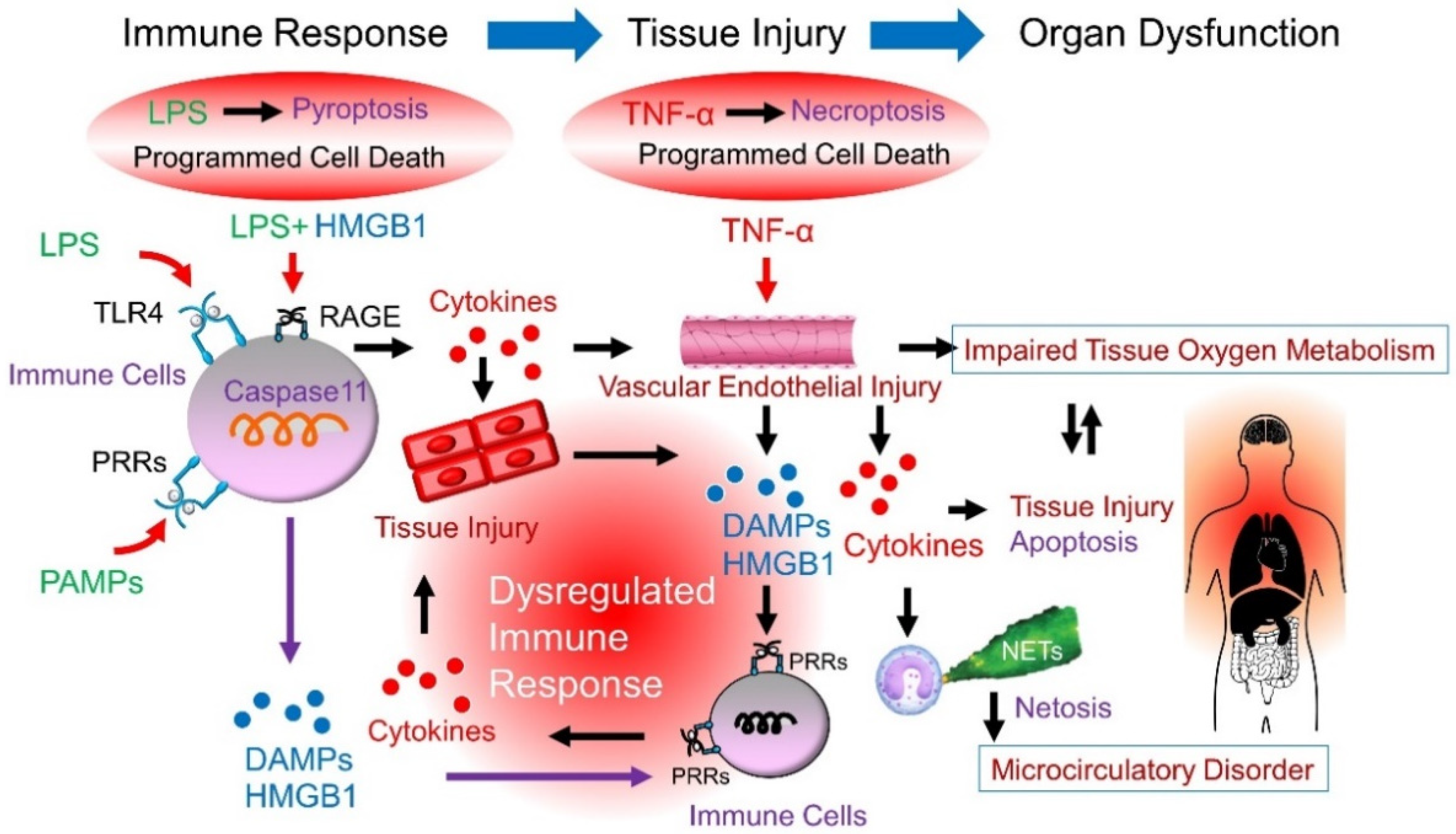
:1. Introduction
2. LPS Recognition Systems
3. LPS-Induced Vascular Endothelial Injury
4. Cytokines, Neutrophils, and DAMPs in Sepsis
4.1. Cytokines
4.2. Neutrophils
4.3. DAMPs
5. Extracorporeal Blood Purification Techniques
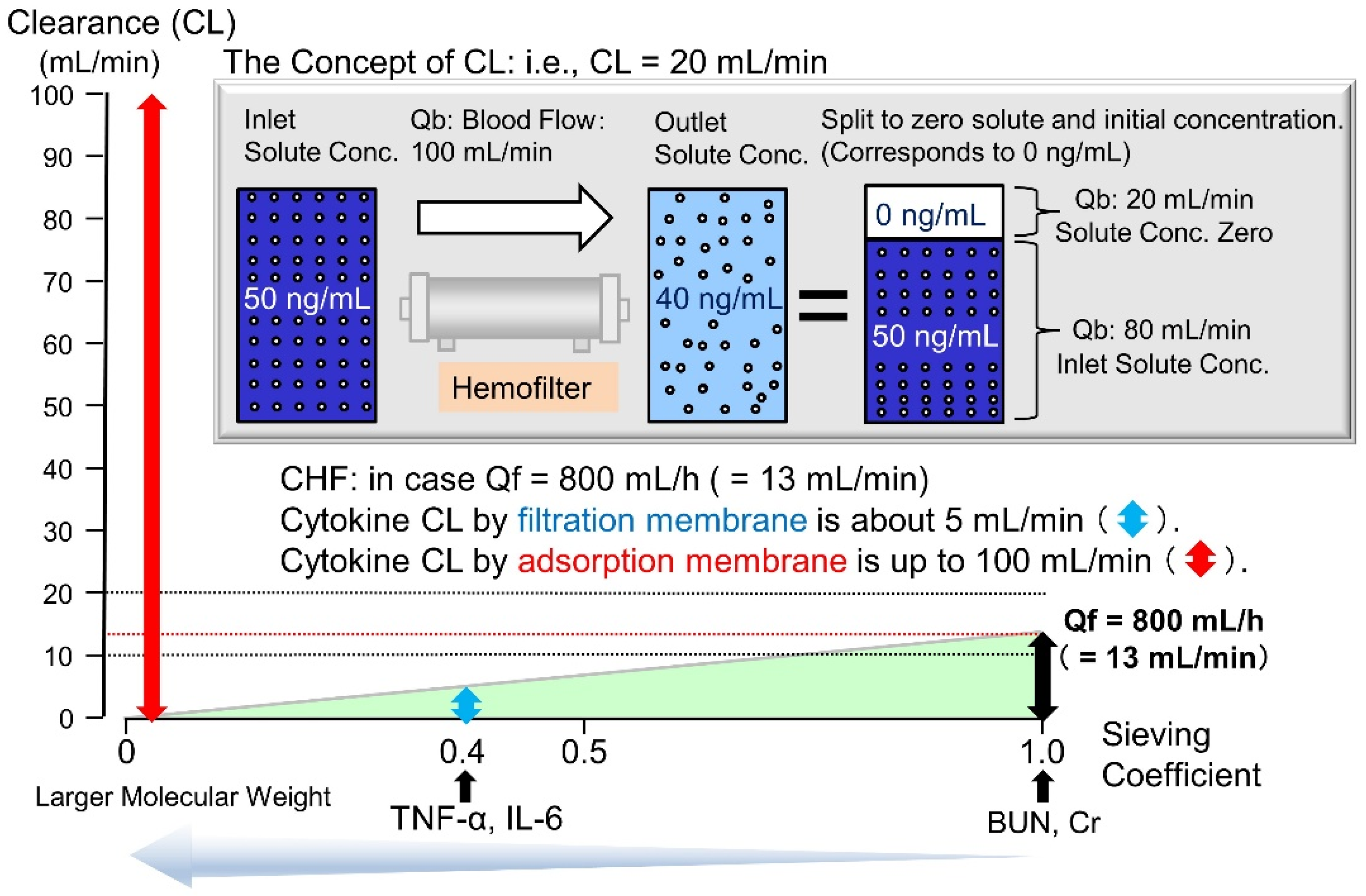
5.1. Rationale
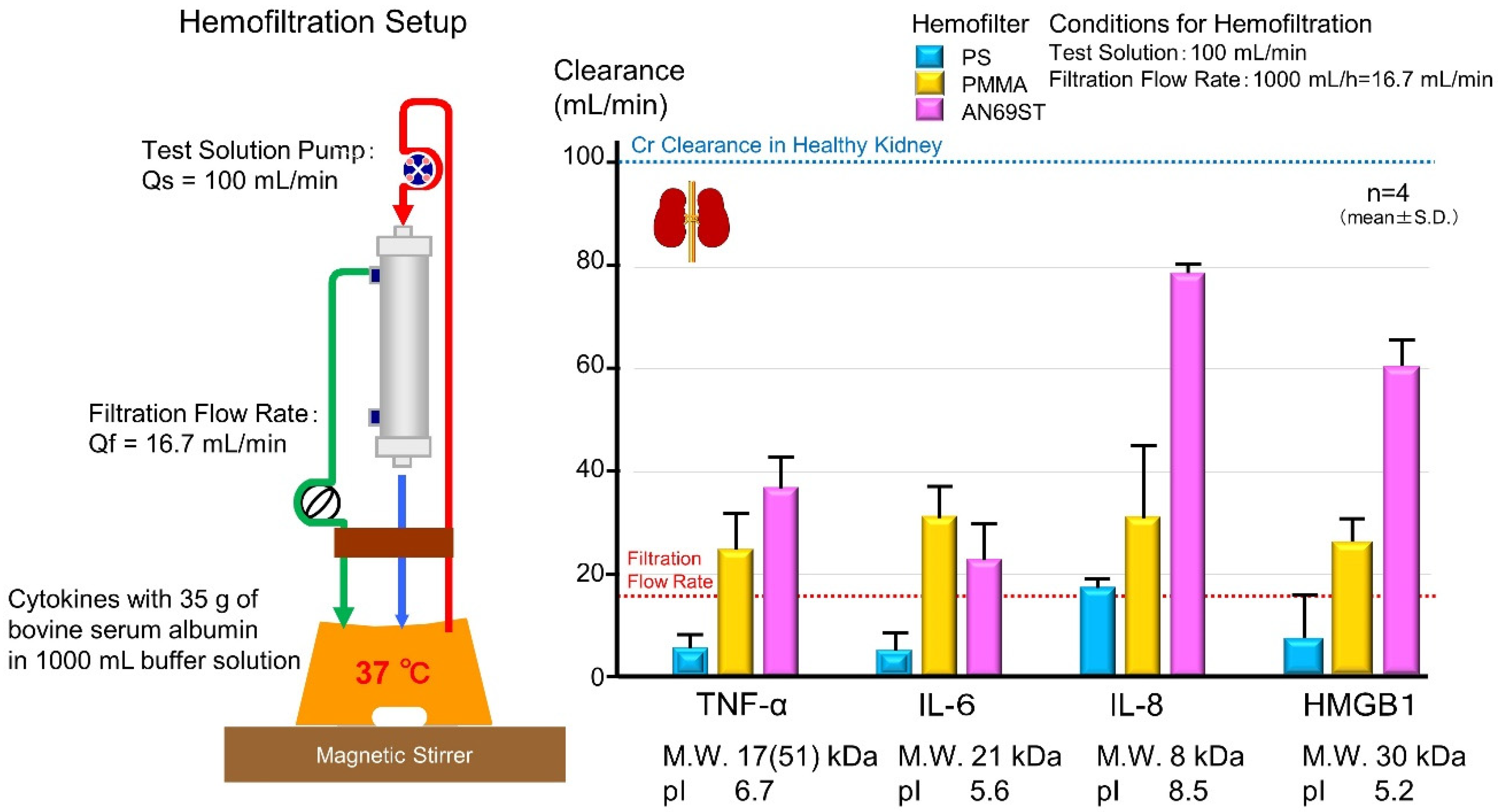
5.2. Cytokine-Adsorbing Hemofilter
5.3. LPS Absorptive Column
5.4. Cytokine Adsorbing Column
5.5. Immune-Modulation Therapy
6. Perspectives and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cecconi, M.; Evans, L.; Levy, M.; Rhodes, A. Sepsis and septic shock. Lancet 2018, 392, 75–87. [Google Scholar] [CrossRef]
- Rivera, A.; Siracusa, M.C.; Yap, G.S.; Gause, W.C. Innate cell communication kick-starts pathogen-specific immunity. Nat. Immunol. 2016, 17, 356–363. [Google Scholar] [CrossRef]
- Van der Poll, T.; van de Veerdonk, F.L.; Scicluna, B.P.; Netea, M.G. The immunopathology of sepsis and potential therapeutic targets. Nat. Rev. Immunol. 2017, 17, 407–420. [Google Scholar] [CrossRef]
- Kieser, K.J.; Kagan, J.C. Multi-receptor detection of individual bacterial products by the innate immune system. Nat. Rev. Immunol. 2017, 17, 376–390. [Google Scholar] [CrossRef]
- Savva, A.; Roger, T. Targeting toll-like receptors: Promising therapeutic strategies for the management of sepsis-associated pathology and infectious diseases. Front. Immunol. 2013, 4, 387. [Google Scholar] [CrossRef] [Green Version]
- Kayagaki, N.; Wong, M.T.; Stowe, I.B.; Ramani, S.R.; Gonzalez, L.C.; Akashi-Takamura, S.; Miyake, K.; Zhang, J.; Lee, W.P.; Muszyński, A.; et al. Noncanonical inflammasome activation by intracellular LPS independent of TLR4. Science 2013, 341, 1246–1249. [Google Scholar] [CrossRef]
- Hagar, J.A.; Powell, D.A.; Aachoui, Y.; Ernst, R.K.; Miao, E.A. Cytoplasmic LPS activates caspase-11: Implications in TLR4-independent endotoxic shock. Science 2013, 341, 1250–1253. [Google Scholar] [CrossRef] [Green Version]
- Cheng, K.T.; Xiong, S.; Ye, Z.; Hong, Z.; Di, A.; Tsang, K.M.; Gao, X.; An, S.; Mittal, M.; Vogel, S.M.; et al. Caspase-11–mediated endothelial pyroptosis underlies endotoxemia-induced lung injury. J. Clin. Investig. 2017, 127, 4124–4135. [Google Scholar] [CrossRef]
- Skirecki, T.; Cavaillon, J.M. Inner sensors of endotoxin–implications for sepsis research and therapy. FEMS Microbiol. Rev. 2019, 43, 239–256. [Google Scholar] [CrossRef]
- Gabarin, R.S.; Li, M.; Zimmel, P.A.; Marshall, J.C.; Li, Y.; Zhang, H. Intracellular and extracellular lipopolysaccharide signaling in sepsis: Avenues for novel therapeutic strategies. J. Innate Immun. 2021, 1–10. [Google Scholar] [CrossRef]
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.D.; Coopersmith, C.M.; et al. The third international consensus definition for sepsis and septic shock (Sepsis-3). JAMA 2016, 315, 801–810. [Google Scholar] [CrossRef] [PubMed]
- McCulloh, R.J.; Opal, S.M. Sepsis management: Importance of the pathogen. In Handbook of Sepsis; Springer: Cham, Switzerland, 2018; pp. 159–184. [Google Scholar]
- Darden, D.B.; Kelly, L.S.; Fenner, B.P.; Moldawer, L.L.; Mohr, A.M.; Efron, P.A. Dysregulated immunity and immunotherapy after sepsis. J. Clin. Med. 2021, 10, 1742. [Google Scholar] [CrossRef]
- Huber-Lang, M.; Lambris, J.D.; Ward, P.A. Innate immune responses to trauma. Nat. Immunol. 2018, 19, 327–341. [Google Scholar] [CrossRef] [PubMed]
- Lelubre, C.; Vincent, J.L. Mechanisms and treatment of organ failure in sepsis. Nat. Rev. Nephrol. 2018, 14, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Luhr, R.; Cao, Y.; Soederquist, B.; Cajander, S. Trends in sepsis mortality over time in randomised sepsis trials: A systematic literature review and meta-analysis of mortality in the control arm, 2002–2016. Crit. Care 2019, 23, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zhang, H.; Chen, Q.; Jiao, F.; Shi, C.; Pei, M.; Lv, J.; Zhang, H.; Wang, L.; Gong, Z. TNF-α/HMGB1 inflammation signalling pathway regulates pyroptosis during liver failure and acute kidney injury. Cell Prolif. 2020, 53, e12829. [Google Scholar] [CrossRef]
- Cutuli, S.L.; Carelli, S.; Grieco, D.L.; De Pascale, G. Immune modulation in critically ill septic patients. Medicina 2021, 57, 552. [Google Scholar] [CrossRef]
- Honore´, P.M.; Hoste, E.; Molnár, Z.; Jacobs, R.; Joannes-Boyau, O.; Malbrain, M.L.; Forni, L.G. Cytokine removal in human septic shock: Where are we and where are we going? Ann. Intensive Care 2019, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Cutuli, S.; Grieco, D.; De Pascale, G.; Antonelli, M. Hemadsorption. Curr. Opin. Anaesthesiol. 2021, 34, 113–118. [Google Scholar] [CrossRef]
- Hirasawa, H.; Oda, S.; Nakamura, M.; Watanabe, E.; Shiga, H.; Matsuda, K. Continuous hemodiafiltration with a cytokine-adsorbing hemofilter for sepsis. Blood Purif. 2012, 34, 164–170. [Google Scholar] [CrossRef]
- Cohen, J. The immunopathogenesis of sepsis. Nature 2002, 420, 885–891. [Google Scholar] [CrossRef]
- Vázquez-Carballo, C.; Guerrero-Hue, M.; García-Caballero, C.; Rayego-Mateos, S.; Opazo-Ríos, L.; Morgado-Pascual, J.L.; Herencia-Bellido, C.; Vallejo-Mudarra, M.; Cortegano, I.; Luisa Gaspar, M.; et al. Toll-like receptors in acute kidney injury. Int. J. Mol. Sci. 2021, 22, 816. [Google Scholar] [CrossRef]
- Mazgaeen, L.; Gurung, P. Recent advances in lipopolysaccharide recognition systems. Int. J. Mol. Sci. 2020, 21, 379. [Google Scholar] [CrossRef] [Green Version]
- Opal, S.M. The host response to endotoxin, anti-lipopolysaccharide strategies, and the management of severe sepsis. Int. J. Med. Microbiol. 2007, 297, 365–377. [Google Scholar] [CrossRef]
- Opal, S.M.; Laterre, P.F.; Francois, B.; LaRosa, S.P.; Angus, D.C.; Mira, J.P.; Wittebole, X.; Dugernier, T.; Perrotin, D.; Tidswell, M.; et al. Effect of eritoran, an antagonist of MD2-TLR4, on mortality in patients with severe sepsis: The ACCESS randomized trial. JAMA 2013, 309, 1154–1162. [Google Scholar] [CrossRef] [Green Version]
- Tse, M.T. Sepsis study failure highlights need for trial design rethink. Nat. Rev. Drug Discov. 2013, 12, 334–335. [Google Scholar] [CrossRef]
- Rathinam, V.A.; Zhao, Y.; Shao, F. Innate immunity to intracellular LPS. Nat. Immunol. 2019, 20, 527–533. [Google Scholar] [CrossRef]
- Kesavardhana, S.; Malireddi, R.S.; Kanneganti, T.D. Caspases in cell death, inflammation, and pyroptosis. Annu. Rev. Immunol. 2020, 38, 567–595. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Gao, W.; Shao, F. Pyroptosis: Gasdermin-mediated programmed necrotic cell death. Trends Biochem. Sci. 2017, 42, 245–254. [Google Scholar] [CrossRef]
- Stowe, I.; Lee, B.; Kayagaki, N. Caspase-11: Arming the guards against bacterial infection. Immunol. Rev. 2015, 265, 75–84. [Google Scholar]
- Wang, J.; Sahoo, M.; Lantier, L.; Warawa, J.; Cordero, H.; Deobald, K.; Re, F. Caspase-11-dependent pyroptosis of lung epithelial cells protects from melioidosis while caspase-1 mediates macrophage pyroptosis and production of IL-18. PLoS Pathog. 2018, 14, e1007105. [Google Scholar] [CrossRef]
- Zamyatina, A.; Heine, H. Lipopolysaccharide recognition in the crossroads of TLR4 and caspase-4/11 mediated inflammatory pathways. Front. Immunol. 2020, 11, 585146. [Google Scholar] [CrossRef]
- Wu, C.; Lu, W.; Zhang, Y.; Zhang, G.; Shi, X.; Hisada, Y.; Grover, S.P.; Zhang, X.; Li, L.; Xiang, B.; et al. Inflammasome activation triggers blood clotting and host death through pyroptosis. Immunity 2019, 50, 1401–1411. [Google Scholar] [CrossRef]
- Yang, X.; Cheng, X.; Tang, Y.; Qiu, X.; Wang, Y.; Kang, H.; Wu, J.; Wang, Z.; Liu, Y.; Chen, F.; et al. Bacterial endotoxin activates the coagulation cascade through gasdermin D-dependent phosphatidylserine exposure. Immunity 2019, 51, 983–996. [Google Scholar] [CrossRef]
- Pfalzgraff, A.; Weindl, G. Intracellular Lipopolysaccharide sensing as a potential therapeutic target for sepsis. Trends Pharmacol. Sci. 2019, 40, 187–197. [Google Scholar] [CrossRef]
- Matthay, M.A.; Zemans, R.L.; Zimmerman, G.A.; Arabi, Y.M.; Beitler, J.R.; Herridge, M.; Randolph, A.G.; Carolyn, S.; Calfee, C.S. Acute respiratory distress syndrome. Nat. Rev. Dis. Primers 2019, 5, 1–22. [Google Scholar] [CrossRef]
- Miao, N.; Yin, F.; Xie, H.; Wang, Y.; Xu, Y.; Shen, Y.; Xu, D.; Yin, J.; Wang, B.; Zhou, Z.; et al. The cleavage of gasdermin D by caspase-11 promotes tubular epithelial cell pyroptosis and urinary IL-18 excretion in acute kidney injury. Kidney Int. 2019, 96, 1105–1120. [Google Scholar] [CrossRef]
- Ye, B.; Chen, X.; Dai, S.; Han, J.; Liang, X.; Lin, S.; Cai, X.; Huang, Z.; Huang, W. Emodin alleviates myocardial ischemia/reperfusion injury by inhibiting gasdermin D-mediated pyroptosis in cardiomyocytes. Drug Des. Dev. Ther. 2019, 13, 975–990. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Meng, R.; Wang, X.; Xu, Y.; Tang, Y.; Wu, J.; Xue, Q.; Yu, S.; Duan, M.; Shan, D.; et al. Caspase-11 signaling enhances graft-versus-host disease. Nat. Commun. 2019, 10, 4044. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Sun, B. Neutrophil pyroptosis: New perspectives on sepsis. Cell. Mol. Life Sci. 2019, 76, 2031–2042. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Hernández, E.G.; Delgado-Coello, B.; Luna-Reyes, I.; Mas-Oliva, J. New insights into lipopolysaccharide inactivation mechanisms in sepsis. Biomed. Pharmacother. 2021, 141, 111890. [Google Scholar] [CrossRef]
- Yamashita, C.; Moriyama, K.; Hasegawa, D.; Hara, Y.; Kuriyama, N.; Nakamura, T.; Shibata, J.; Komura, H.; Nishida, O. Evidence and perspectives on the use of polymyxin B-immobilized fiber column hemoperfusion among critically ill patients. In Recent Advances in Dialysis Therapy in Japan; Karger Publishers: Basel, Switzerland, 2018; Volume 196, pp. 215–222. [Google Scholar]
- Vanaja, S.K.; Russo, A.J.; Behl, B.; Banerjee, I.; Yankova, M.; Deshmukh, S.D.; Rathinam, V.A. Bacterial outer membrane vesicles mediate cytosolic localization of LPS and caspase-11 activation. Cell 2016, 165, 1106–1119. [Google Scholar] [CrossRef] [Green Version]
- Kopp, F.; Kupsch, S.; Schromm, A.B. Lipopolysaccharide-binding protein is bound and internalized by host cells and colocalizes with LPS in the cytoplasm: Implications for a role of LBP in intracellular LPS-signaling. Biochim. Biophys. Acta 2016, 1863, 660–672. [Google Scholar] [CrossRef]
- Deng, M.; Tang, Y.; Li, W.; Wang, X.; Zhang, R.; Zhang, X.; Zhao, X.; Liu, J.; Tang, C.; Liu, Z.; et al. The Endotoxin Delivery Protein HMGB1 Mediates Caspase-11-Dependent Lethality in Sepsis. Immunity 2018, 49, 740–753. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Wang, H.; Andersson, U. Targeting inflammation driven by HMGB1. Front. Immunol. 2020, 11, 484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vijayakumar, E.C.; Bhatt, L.K.; Prabhavalkar, K.S. High Mobility Group Box-1 (HMGB1): A potential target in therapeutics. Curr. Drug Targets 2019, 20, 1474–1485. [Google Scholar] [CrossRef]
- Deng, C.; Zhao, L.; Yang, Z.; Shang, J.J.; Wang, C.Y.; Shen, M.Z.; Jiang, S.; Li, T.; Di, W.C.; Chen, Y. Targeting HMGB1 for the treatment of sepsis and sepsis-induced organ injury. Acta Pharmacol. Sin. 2021, 1–9. [Google Scholar] [CrossRef]
- Yoo, H.; Im, Y.; Ko, R.E.; Lee, J.Y.; Park, J.; Jeon, K. Association of plasma level of high-mobility group box-1 with necroptosis and sepsis outcomes. Sci. Rep. 2021, 11, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Fajgenbaum, D.C.; June, C.H. Cytokine storm. N. Engl. J. Med. 2020, 383, 2255–2273. [Google Scholar] [CrossRef] [PubMed]
- Peerapornratana, S.; Manrique-Caballero, C.L.; Gómez, H.; Kellum, J.A. Acute kidney injury from sepsis: Current concepts, epidemiology, pathophysiology, prevention and treatment. Kidney Int. 2019, 96, 1083–1099. [Google Scholar] [CrossRef]
- Kellum, J.A.; Prowle, J.R. Paradigms of acute kidney injury in the intensive care setting. Nat. Rev. Nephrol. 2018, 14, 217. [Google Scholar] [CrossRef] [PubMed]
- Drosatos, K.; Lymperopoulos, A.; Kennel, P.J.; Pollak, N.; Schulze, P.C.; Goldberg, I.J. Pathophysiology of sepsis-related cardiac dysfunction: Driven by inflammation, energy mismanagement, or both? Heart Fail. Rep. 2015, 12, 130–140. [Google Scholar] [CrossRef] [Green Version]
- Widmann, C.N.; Heneka, M.T. Long-term cerebral consequences of sepsis. Lancet Neurol. 2014, 13, 630–636. [Google Scholar] [CrossRef]
- Juffermans, N.P.; van den Brom, C.E.; Kleinveld, D.J. Targeting endothelial dysfunction in acute critical illness to reduce organ failure. Anesth. Analg. 2020, 131, 1708–1720. [Google Scholar] [CrossRef]
- Lee, W.L.; Slutsky, A.S. Sepsis and endothelial permeability. N. Engl. J. Med. 2010, 363, 689–691. [Google Scholar] [CrossRef] [Green Version]
- Iba, T.; Levy, J.H. Derangement of the endothelial glycocalyx in sepsis. J. Thromb. Haemost. 2019, 17, 283–294. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, E.P.; Yang, Y.; Janssen, W.J.; Gandjeva, A.; Perez, M.J.; Barthel, L.; Zemans, R.L.; Bowman, J.C.; Koyanagi, D.E.; Yunt, Z.X. The pulmonary endothelial glycocalyx regulates neutrophil adhesion and lung injury during experimental sepsis. Nat. Med. 2012, 18, 1217–1223. [Google Scholar] [CrossRef] [Green Version]
- Dolmatova, E.V.; Wang, K.; Mandavilli, R.; Griendling, K.K. The effects of sepsis on endothelium and clinical implications. Cardiovasc. Res. 2021, 117, 60–73. [Google Scholar] [CrossRef]
- Kruttgen, A.; Rose-John, S. Interleukin-6 in sepsis and capillary leakage syndrome. J. Interferon Cytokine Res. 2012, 32, 60–65. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Liu, Y.; Li, L.; Zhang, R.; Liu, W.; Wu, J.; Mao, E.; Tang, Y. HMGB1 increases permeability of the endothelial cell monolayer via RAGE and Src family tyrosine kinase pathways. Inflammation 2012, 35, 350–362. [Google Scholar] [CrossRef]
- Goligorsky, M.S.; Sun, D. Glycocalyx in endotoxemia and sepsis. Am. J. Pathol. 2020, 190, 791–798. [Google Scholar] [CrossRef]
- Joffre, J.; Hellman, J.; Ince, C.; Ait-Oufella, H. Endothelial responses in sepsis. Am. J. Respir. Crit. Care Med. 2020, 202, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Wake, H.; Gao, Y.; Wang, D.; Mori, S.; Liu, K.; Teshigawara, K.; Takahashi, H.; Nishibori, M. Histidine-rich glycoprotein ameliorates endothelial barrier dysfunction through regulation of NF-kB and MAPK signal pathway. Br. J. Pharmacol. 2019, 176, 2808–2824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iba, T.; Levy, J.H.; Hirota, T.; Hiki, M.; Sato, K.; Murakami, T.; Nagaoka, I. Protection of the endothelial glycocalyx by antithrombin in an endotoxin-induced rat model of sepsis. Throm. Res. 2018, 171, 1–6. [Google Scholar] [CrossRef]
- Villa, G.; Husain-Syed, F.; Saitta, T.; Degl’Innocenti, D.; Barbani, F.; Resta, M.; Castellani, G.; Romagnoli, S. Hemodynamic instability during acute kidney injury and acute renal replacement therapy: Pathophysiology and clinical implications. Blood Purif. 2021, 1–11. [Google Scholar] [CrossRef]
- El-Benna, J.; Hurtado-Nedelec, M.; Marzaioli, V.; Marie, J.C.; Gougerot-Pocidalo, M.A.; Dang, P.M.C. Priming of the neutrophil respiratory burst: Role in host defense and inflammation. Immunol. Rev. 2016, 273, 180–193. [Google Scholar] [CrossRef]
- Stiel, L.; Meziani, F.; Helms, J. Neutrophil activation during septic shock. Shock 2018, 49, 371–384. [Google Scholar] [CrossRef]
- Shen, X.F.; Cao, K.; Jiang, J.P.; Guan, W.X.; Du, J.F. Neutrophil dysregulation during sepsis: An overview and update. J. Cell Mol. Med. 2017, 21, 1687–1697. [Google Scholar] [CrossRef] [Green Version]
- Sonego, F.; Castanheira, F.V.; Ferreira, R.G.; Kanashiro, A.; Leite, C.A.; Nascimento, D.C.; Colon, D.F.; Borges Vde, F.; Alves-Filho, J.C.; Cunha, F.Q. Paradoxical roles of the neutrophil in sepsis: Protective and deleterious. Front. Immunol. 2016, 7, 155. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Moreno, D.; Adrover, J.M.; Hidalgo, A. Neutrophils as effectors of vascular inflammation. Eur. J. Clin. Investig. 2018, 48, e12940. [Google Scholar] [CrossRef] [Green Version]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, H.; Qu, M.; Nan, K.; Cao, H.; Cata, J.P.; Chen, W.; Miao, C. The emerging role of neutrophil extracellular traps in sepsis and sepsis-associated thrombosis. Front. Cell. Infect. Microbiol. 2021, 11, 653228. [Google Scholar] [CrossRef] [PubMed]
- Iba, T.; Levy, J.H. Inflammation and thrombosis: Roles of neutrophils, platelets and endothelial cells and their interactions in thrombus formation during sepsis. J. Thromb. Haemost. 2018, 16, 231–241. [Google Scholar] [CrossRef]
- Kato, Y.; Nishida, O.; Kuriyama, N.; Nakamura, T.; Kawaji, T.; Onouchi, T.; Hasegawa, D.; Shimomura, Y. Effects of thrombomodulin in reducing lethality and suppressing neutrophil extracellular trap formation in the lungs and liver in a lipopolysaccharide-induced murine septic shock model. Int. J. Mol. Sci. 2021, 22, 4933. [Google Scholar] [CrossRef]
- Alves-Filho, J.C.; de Freitas, A.; Spiller, F.; Souto, F.O.; Cunha, F.Q. The role of neutrophils in severe sepsis. Shock 2008, 30, 3–9. [Google Scholar] [CrossRef] [PubMed]
- McDonald, B. Neutrophils in critical illness. Cell Tissue Res. 2018, 373, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.A.; Brain, S.D.; Pearson, J.D.; Edgeworth, J.D.; Lewis, S.M.; Treacher, D.F. Neutrophils in development of multiple organ failure in sepsis. Lancet 2006, 368, 157–169. [Google Scholar] [CrossRef]
- Németh, T.; Sperandio, M.; Mócsai, A. Neutrophils as emerging therapeutic targets. Nat. Rev. Drug. Discov. 2020, 19, 253–275. [Google Scholar] [CrossRef]
- Ma, S.; Xu, Q.; Deng, B.; Zheng, Y.; Tian, H.; Wang, L.; Ding, F. Granulocyte and monocyte adsorptive apheresis ameliorates sepsis in rats. Intensive Care Med. Exp. 2017, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Karki, R.; Kanneganti, T.D. The ‘cytokine storm’: Molecular mechanisms and therapeutic prospects. Trends Immunol. 2021, 42, 681–705. [Google Scholar] [CrossRef]
- Eppensteiner, J.; Kwun, J.; Scheuermann, U.; Barbas, A.; Limkakeng, A.T.; Kuchibhatla, M.; Elster, E.A.; Kirk, A.D.; Lee, J. Damage- and pathogen-associated molecular patterns play differential roles in late mortality after critical illness. JCI Insight 2019, 4, e127925. [Google Scholar] [CrossRef]
- Malireddi, R.K.; Kesavardhana, S.; Kanneganti, T.D. ZBP1 and TAK1: Master regulators of NLRP3 inflammasome/pyroptosis, apoptosis, and necroptosis (PAN-optosis). Front. Cell. Infect. Microbiol. 2019, 9, 406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fritsch, M.; Günther, S.D.; Schwarzer, R.; Albert, M.C.; Schorn, F.; Werthenbach, J.P.; Schiffmann, L.M.; Stair, N.; Stocks, H.; Seeger, J.M.; et al. Caspase-8 is the molecular switch for apoptosis, necroptosis and pyroptosis. Nature 2019, 575, 683–687. [Google Scholar] [CrossRef] [PubMed]
- Duprez, L.; Takahashi, N.; van Hauwermeiren, F.; van Dendriessche, B.; Goossens, V.; Vanden Berghe, T.; Declercq, W.; Libert, C.; Cauwels, A.; Vandenabeele, P. RIP kinase-dependent necrosis drives lethal systemic inflammatory response syndrome. Immunity 2011, 35, 908–918. [Google Scholar] [CrossRef] [Green Version]
- Pasparakis, M.; Vandenabeele, P. Necroptosis and its role in inflammation. Nature 2015, 517, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Wallach, D.; Kang, T.B.; Dillon, C.P.; Green, D.R. Programmed necrosis in inflammation: Toward identification of the effector molecules. Science 2016, 352, aaf2154. [Google Scholar] [CrossRef]
- Lord, J.M.; Midwinter, M.J.; Chen, Y.F.; Belli, A.; Brohi, K.; Kovacs, E.J.; Koenderman, L.; Kubes, P.; Lilford, R.J. The systemic immune response to trauma: An overview of pathophysiology and treatment. Lancet 2014, 384, 1455–1465. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.H.; Wong, S.W.; Martinez, J. Programmed necrosis and disease: We interrupt your regular programming to bring you necroinflammation. Cell. Death Differ. 2019, 26, 25–40. [Google Scholar] [CrossRef] [Green Version]
- Nedeva, C.; Menassa, J.; Puthalakath, H. Sepsis: Inflammation is a necessary evil. Front. Cell Dev. Biol. 2019, 7, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monard, C.; Rimmelé, T.; Ronco, C. Extracorporeal Blood Purification Therapies for Sepsis. Blood Purif. 2019, 47, 1–14. [Google Scholar] [CrossRef]
- Zhang, L.; Feng, Y.; Fu, P. Blood purification for sepsis: An overview. Precis. Clin. Med. 2021, 4, 45–55. [Google Scholar] [CrossRef]
- Girardot, T.; Schneider, A.; Rimmelé, T. Blood purification techniques for sepsis and septic AKI. Semin. Nephrol. 2019, 39, 505–514. [Google Scholar] [CrossRef]
- Pickkers, P.; Vassiliou, T.; Liguts, V.; Prato, F.; Tissieres, P.; Kloesel, S.; Turani, F.; Popevski, D.; Broman, M.; Gindac, C.M.; et al. Sepsis management with a blood purification membrane: European experience. Blood Purif. 2019, 47, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Gruda, M.C.; Ruggeberg, K.G.; O’Sullivan, P.; Guliashvili, T.; Scheirer, A.R.; Golobish, T.D.; Capponi, V.J.; Chan, P.P. Broad adsorption of sepsis-related PAMP and DAMP molecules, mycotoxins, and cytokines from whole blood using CytoSorb(R) sorbent porous polymer beads. PLoS ONE 2018, 13, e0191676. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gallagher, M.; Li, Q.; Lo, S.; Cass, A.; Finfer, S.; Myburgh, J.; Bouman, C.; Faulhaber-Walter, R.; Kellum, J.A.; et al. Renal replacement therapy intensity for acute kidney injury and recovery to dialysis independence: A systematic review and individual patient data meta-analysis. Nephrol. Dial. Trans. 2017, 33, 1017–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joannes-Boyau, O.; Honoré, P.M.; Perez, P.; Bagshaw, S.M.; Grand, H.; Canivet, J.L.; Dewitte, A.; Flamens, C.; Pujol, W.; Grandoulier, A.S. High-volume versus standard-volume haemofiltration for septic shock patients with acute kidney injury (IVOIRE study): A multicentre randomized controlled trial. Intensive Care Med. 2013, 39, 1535–1546. [Google Scholar] [CrossRef]
- Atan, R.; Peck, L.; Prowle, J.; Licari, E.; Eastwood, G.M.; Storr, M.; Hermann, G.; Bellomo, R. A double-blind randomized controlled trial of high cutoff versus standard hemofiltration in critically ill patients with acute kidney injury. Crit. Care Med. 2018, 46, e988–e994. [Google Scholar] [CrossRef]
- Putzu, A.; Schorer, R.; Lopez-Delgado, J.C.; Cassina, T.; Landoni, G. Blood purification and mortality in sepsis and septic shock: A systematic review and meta-analysis of randomized trials. Anesthesiology 2019, 131, 580–593. [Google Scholar] [CrossRef]
- Moriyama, K.; Soejima, Y. Continuous hemodialfiltration using PMMA membrane: Clinical efficacy and its mechanisms. Contrib. Nephrol. 1999, 125, 222–232. [Google Scholar]
- Florens, N.; Guebre-Egziabher, F.; Juillard, L. Reconsidering adsorption in hemodialysis: Is it just an epiphenomenon? A narrative review. J. Nephrol. 2021, 1–9. [Google Scholar] [CrossRef]
- Joannes-Boyau, O.; Honore, P.M.; Boer, W.; Collin, V. Are the synergistic effects of high-volume haemofiltration and enhanced adsorption the missing key in sepsis modulation? Nephrol. Dial. Trans. 2009, 24, 354–357. [Google Scholar] [CrossRef] [Green Version]
- Hattori, N.; Oda, S. Cytokine-adsorbing hemofilter: Old but new modality for septic acute kidney injury. Ren. Replace. Ther. 2016, 2, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Shiga, H.; Hirasawa, H.; Nishida, O.; Oda, S.; Nakamura, M.; Mashiko, K.; Matsuda, K.; Kitamura, N.; Kikuchi, Y.; Fuke, N. Continuous hemodiafiltration with a cytokine-adsorbing hemofilter in patients with septic shock: A preliminary report. Blood Purif. 2014, 38, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.; Moriyama, K.; Ledebo, I. AN69: Evolution of the world’s first high permeability membrane. In High-Performance Membrane Dialyzers; Karger Publishers: Basel, Switzerland, 2011; pp. 119–129. [Google Scholar]
- Nishida, O.; Nakamura, T.; Kuriyama, N.; Hara, Y.; Yumoto, M.; Shimomura, Y.; Moriyama, K. Sustained high-efficiency daily diafiltration using a mediator-adsorbing membrane (SHEDD-fA) in the treatment of patients with severe sepsis. In High-Performance Membrane Dialyzers; Karger Publishers: Basel, Switzerland, 2011; pp. 172–181. [Google Scholar]
- Honore, P.M.; Jacobs, R.; Joannes-Boyau, O.; De Regt, J.; De Waele, E.; Van Gorp, V.; Boer, W.; Verfaillie, L.; Spapen, H.D. Newly designed CRRT membranes for sepsis and SIRS—A pragmatic approach for bedside intensivists summarizing the more recent advances: A systematic structured review. ASAIO J. 2013, 59, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Arimura, T.; Abe, M.; Shiga, H.; Katayama, H.; Kaizu, K.; Oda, S. Clinical study of blood purification therapy in critical care in Japan: Results from the survey research of the Japan Society for Blood Purification in Critical Care in 2013. J. Artif. Organs 2017, 20, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Yumoto, M.; Nishida, O.; Moriyama, K.; Shimomura, Y.; Nakamura, T.; Kuriyama, N.; Hara, Y.; Yamada, S. In vitro evaluation of high mobility group box 1 protein removal with various membranes for continuous hemofiltration. Ther. Apher. Dial. 2011, 15, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, K.; Kato, Y.; Hasegawa, D.; Kurimoto, Y.; Kawaji, T.; Nakamura, T.; Kuriyama, N.; Shimomura, Y.; Nishida, O. Involvement of ionic interactions in cytokine adsorption of polyethyleneimine-coated polyacrylonitrile and polymethyl methacrylate membranes in vitro. J. Artif. Organs 2020, 23, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Moriyama, K.; Shimomura, Y.; Kato, Y.; Kuriyama, N.; Hara, Y.; Yamada, S.; Nishida, O. Adsorption kinetics of high mobility group box 1 protein in a polyacrylonitrile hemofiltration membrane. Ther. Apher. Dial. 2021, 25, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Moriyama, K.; Hara, Y.; Hinoue, T.; Kato, Y.; Hasegawa, D.; Kuriyama, N.; Nakamura, T.; Komatsu, S.; Yamashita, C.; et al. Comparison of myoglobin clearance in three types of blood purification modalities. Ther. Apher. Dial. 2021, 25, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Hiraiwa, T.; Moriyama, K.; Matsumoto, K.; Shimomura, Y.; Kato, Y.; Yamashita, C.; Hara, Y.; Kawaji, T.; Kurimoto, Y.; Nakamura, T.; et al. In vitro evaluation of linezolid and doripenem clearance with different hemofilters. Blood Purif. 2020, 49, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Doi, K.; Nishida, O.; Shigematsu, T.; Sadahiro, T.; Itami, N.; Iseki, K.; Yuzawa, Y.; Okada, H.; Koya, D.; Kiyomoto, H.; et al. The Japanese clinical practice guideline for acute kidney injury 2016. Ren. Replace. Ther. 2018, 4, 1–55. [Google Scholar] [CrossRef] [Green Version]
- Kobashi, S.; Maruhashi, T.; Nakamura, T.; Hatabayashi, E.; Kon, A. The 28-day survival rates of two cytokine-adsorbing hemofilters for continuous renal replacement therapy: A single-center retrospective comparative study. Acute Med. Surg. 2019, 6, 60–67. [Google Scholar] [CrossRef] [Green Version]
- Doi, K.; Iwagami, M.; Yoshida, E.; Marshall, M.R. Associations of polyethylenimine-coated AN69ST membrane in continuous renal replacement therapy with the intensive care outcomes: Observations from a claims database from Japan. Blood Purif. 2017, 44, 184–192. [Google Scholar] [CrossRef] [Green Version]
- Shoji, H.; Opal, S.M. Therapeutic Rationale for Endotoxin Removal with Polymyxin B Immobilized Fiber Column (PMX) for Septic Shock. Int. J. Mol. Sci. 2021, 22, 2228. [Google Scholar] [CrossRef]
- Vincent, J.L.; Laterre, P.F.; Cohen, J.; Burchardi, H.; Bruining, H.; Lerma, F.A.; Wittebole, X.; Backer, D.D.; Brett, S.; Marzo, D.; et al. A pilot-controlled study of a polymyxin B-immobilized hemoperfusion cartridge in patients with severe sepsis secondary to intra-abdominal infection. Shock 2005, 23, 400–405. [Google Scholar] [CrossRef] [Green Version]
- Cruz, D.N.; Antonelli, M.; Fumagalli, R.; Foltran, F.; Brienza, N.; Donati, A.; Malcangi, V.; Petrini, F.; Volta, G.; Pallavicini, F.M.B.; et al. Early use of polymyxin B hemoperfusion in abdominal septic shock The EUPHAS randomized controlled trial. JAMA 2009, 301, 2445–2452. [Google Scholar] [CrossRef] [Green Version]
- Payen, D.M.; Guilhot, J.; Launey, Y.; Lukaszewicz, A.C.; Kaaki, M.; Veber, B.; Pottecher, J.; Joannes-Boyau, O.; Martin-Lefevre, L.; Jabaudon, M.; et al. Early use of polymyxin B hemoperfusion in patients with septic shock due to peritonitis: A multicenter randomized control trial. Intensive Care Med. 2015, 41, 975–984. [Google Scholar] [CrossRef] [Green Version]
- Dellinger, R.P.; Bagshaw, S.M.; Antonelli, M.; Foster, D.M.; Klein, D.J.; Marshall, J.C.; Palevsky, P.M.; Weisberg, L.S.; Schorr, C.A.; Trzeciak, S.; et al. Effect of targeted Polymyxin B hemoperfusion on 28-day mortality in patients with septic shock and elevated endotoxin level the EUPHRATES randomized clinical trial. JAMA 2018, 320, 1455–1463. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, C.; Moriyama, K.; Hasegawa, D.; Kato, Y.; Sakai, T.; Kawaji, T.; Shimomura, Y.; Kunimoto, Y.; Nagata, M.; Nishida, O. In vitro study of endotoxin adsorption by a polymyxin B-immobilized fiber column. Blood Purif. 2018, 46, 269–273. [Google Scholar] [CrossRef]
- Rachoin, J.S.; Foster, D.; Giese, R.; Weisberg, L.S.; Klein, D.J. Importance of endotoxin clearance in endotoxemic septic shock: An analysis from the Evaluating Use of PolymyxinB Hemoperfusion in a Randomized Controlled Trial of Adults Treated for Endotoxemic Septic Shock (EUPHRATES) trial. Crit. Care Explor. 2020, 2, e0083. [Google Scholar] [CrossRef] [Green Version]
- Malard, B.; Lambert, C.; Kellum, J.A. In vitro comparison of the adsorption of inflammatory mediators by blood purification devices. Intensive Care Med. Exp. 2018, 6, 12. [Google Scholar] [CrossRef] [Green Version]
- Bonavia, A.; Groff, A.; Karamchandani, K.; Singbartl, K. Clinical utility of extracorporeal cytokine hemoadsorption therapy: A literature review. Blood Purif. 2018, 46, 337–349. [Google Scholar] [CrossRef] [PubMed]
- Scharf, C.; Schroeder, I.; Paal, M.; Winkels, M.; Irlbeck, M.; Zoller, M.; Liebchen, U. Can the cytokine adsorber CytoSorb® help to mitigate cytokine storm and reduce mortality in critically ill patients? A propensity score matching analysis. Ann. Intensive Care 2021, 11, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kogelmann, K.; Jarczak, D.; Scheller, M.; Drüner, M. Hemoadsorption by CytoSorb in septic patients: A case series. Crit. Care 2017, 21, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schädler, D.; Pausch, C.; Heise, D.; Meier-Hellmann, A.; Brederlau, J.; Weiler, N.; Marx, G.; Putensen, C.; Spies, C.; Jörres, A.; et al. The effect of a novel extracorporeal cytokine hemoadsorption device on IL-6 elimination in septic patients: A randomized controlled trial. PLoS ONE 2017, 12, e0187015. [Google Scholar] [CrossRef] [Green Version]
- Rimmelé, T.; Payen, D.; Cantaluppi, V.; Marshall, J.; Gomez, H.; Gomez, A.; Murray, P.; Kellum, J.A.; Workgroup ADQIXIV. Immune cell phenotype and function in sepsis. Shock 2016, 45, 282–291. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.; Cao, K.; Zhao, Y.; Du, J. Targeting Neutrophils in Sepsis: From Mechanism to Translation. Front. Pharmacol. 2020, 12, 644270. [Google Scholar] [CrossRef]
- Steinhagen, F.; Schmidt, S.; Schewe, J.C.; Peukert, K.; Klinman, D.M.; Bode, C. Immunotherapy in sepsis-brake or accelerate? Pharmacol. Ther. 2020, 208, 107476. [Google Scholar] [CrossRef]
- Hara, Y.; Shimomura, Y.; Nakamura, T.; Kuriyama, N.; Yamashita, C.; Kato, Y.; Miyasho, T.; Sakai, T.; Yamada, S.; Moriyama, K.; et al. Novel blood purification system for regulating excessive immune reactions in severe sepsis and septic shock: An ex vivo pilot study. Ther. Apher. Dial. 2015, 19, 308–315. [Google Scholar] [CrossRef]
- Takeda, Y.; Ohba, M.; Ueno, M.; Saniabadi, A.R.; Wakabayashi, I. Cellulose acetate beads activate the complement system but inactivate the anaphylatoxins generated. Artif. Organs 2010, 34, 1144–1149. [Google Scholar] [CrossRef]
- Hiraishi, K.; Takeda, Y.; Shiobara, N.; Shibusawa, H.; Jimma, F.; Kashiwagi, N.; Saniabadi, A.R.; Adachi, M. Studies on the mechanisms of leukocyte adhesion to cellulose acetate beads: An in vitro model to assess the efficacy of cellulose acetate carrier-based granulocyte and monocyte adsorptive apheresis. Ther. Apher. Dial. 2003, 7, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Hanai, H.; Takeda, Y.; Eberhardson, M.; Gruber, R.; Saniabadi, A.R.; Winqvist, O.; Lofberg, R. The mode of actions of the Adacolumn therapeutic leucocytapheresis in patients with inflammatory bowel disease: A concise review. Clin. Exp. Immunol. 2011, 163, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Cai, S.; Su, J. The pathogenesis of sepsis and potential therapeutic targets. Int. J. Mol. Sci. 2019, 20, 5376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romagnoli, S.; Ricci, Z.; Ronco, C. CRRT for sepsis-induced acute kidney injury. Curr. Opin. Crit. Care 2018, 24, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Cavaillon, J.M.; Singer, M.; Skirecki, T. Sepsis therapies: Learning from 30 years of failure of translational research to propose new leads. EMBO Mol. Med. 2020, 12, e10128. [Google Scholar] [CrossRef] [PubMed]
- Honoré, P.M.; De Bels, D.; Gutierrez, L.B.; Spapen, H.D. Hemoadsorption therapy in the critically ill: Solid base but clinical haze. Ann. Intensive Care 2019, 9, 22. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moriyama, K.; Nishida, O. Targeting Cytokines, Pathogen-Associated Molecular Patterns, and Damage-Associated Molecular Patterns in Sepsis via Blood Purification. Int. J. Mol. Sci. 2021, 22, 8882. https://doi.org/10.3390/ijms22168882
Moriyama K, Nishida O. Targeting Cytokines, Pathogen-Associated Molecular Patterns, and Damage-Associated Molecular Patterns in Sepsis via Blood Purification. International Journal of Molecular Sciences. 2021; 22(16):8882. https://doi.org/10.3390/ijms22168882
Chicago/Turabian StyleMoriyama, Kazuhiro, and Osamu Nishida. 2021. "Targeting Cytokines, Pathogen-Associated Molecular Patterns, and Damage-Associated Molecular Patterns in Sepsis via Blood Purification" International Journal of Molecular Sciences 22, no. 16: 8882. https://doi.org/10.3390/ijms22168882
APA StyleMoriyama, K., & Nishida, O. (2021). Targeting Cytokines, Pathogen-Associated Molecular Patterns, and Damage-Associated Molecular Patterns in Sepsis via Blood Purification. International Journal of Molecular Sciences, 22(16), 8882. https://doi.org/10.3390/ijms22168882