
Oral Squamous Cell Carcinoma Contributes to Differentiation of Monocyte-Derived Tumor-Associated Macrophages via PAI-1 and IL-8 Production

 ,
,
Abstract
:1. Introduction
2. Results
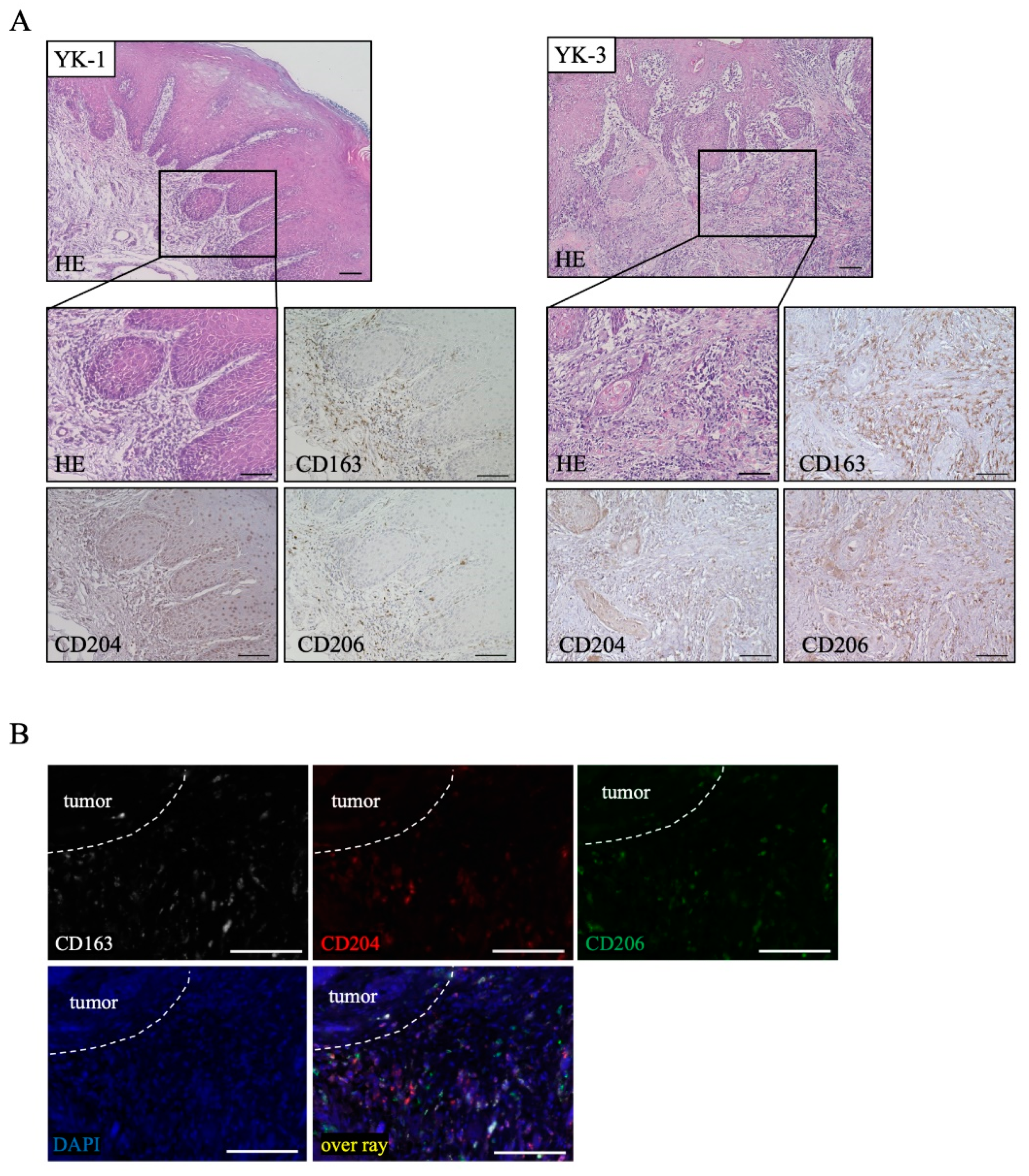
2.1. Distribution of TAM Markers in OSCC Tissues
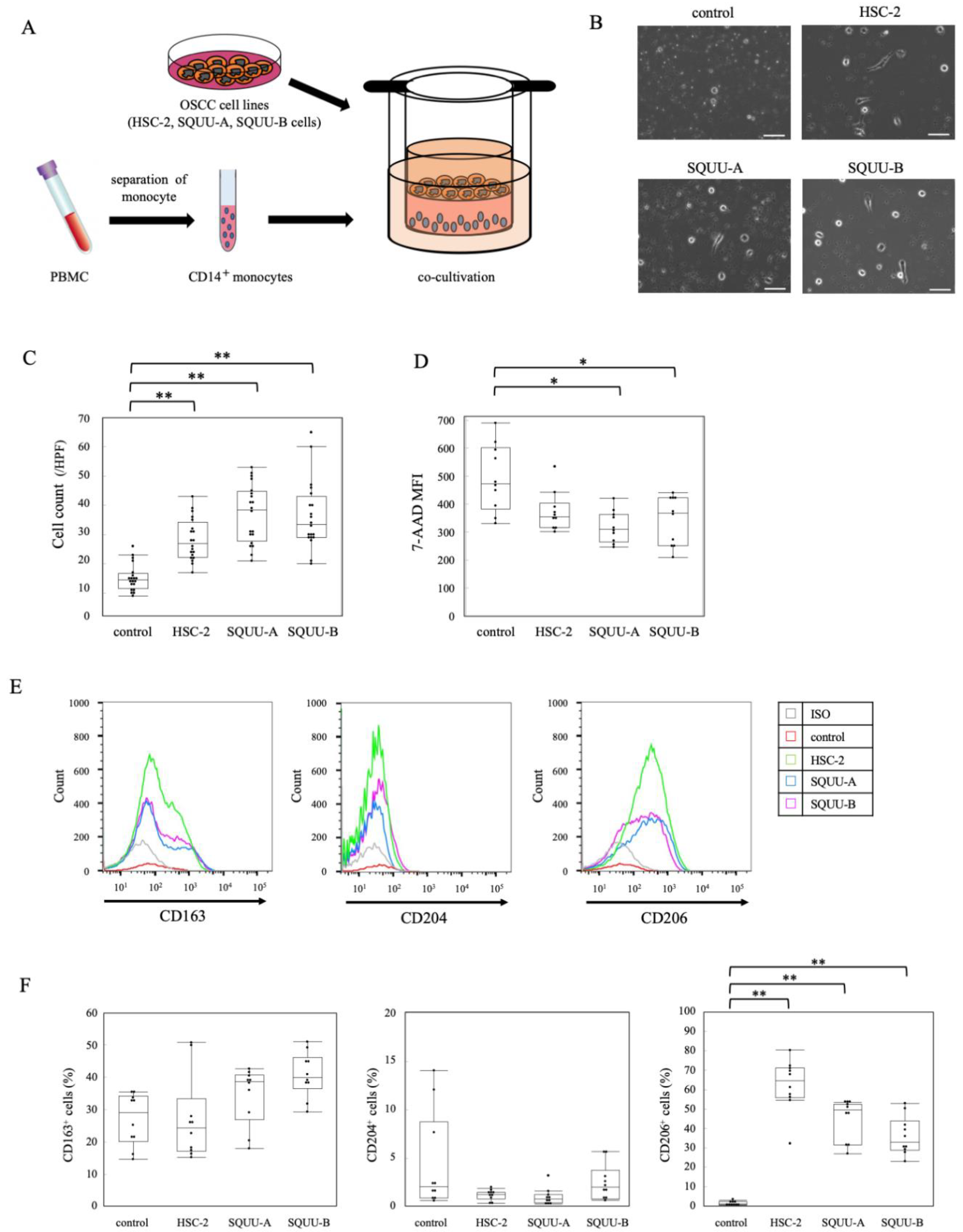
2.2. Effects of OSCC Cell Co-Culture on Apoptosis and Activation Status of CD14+ Monocytes
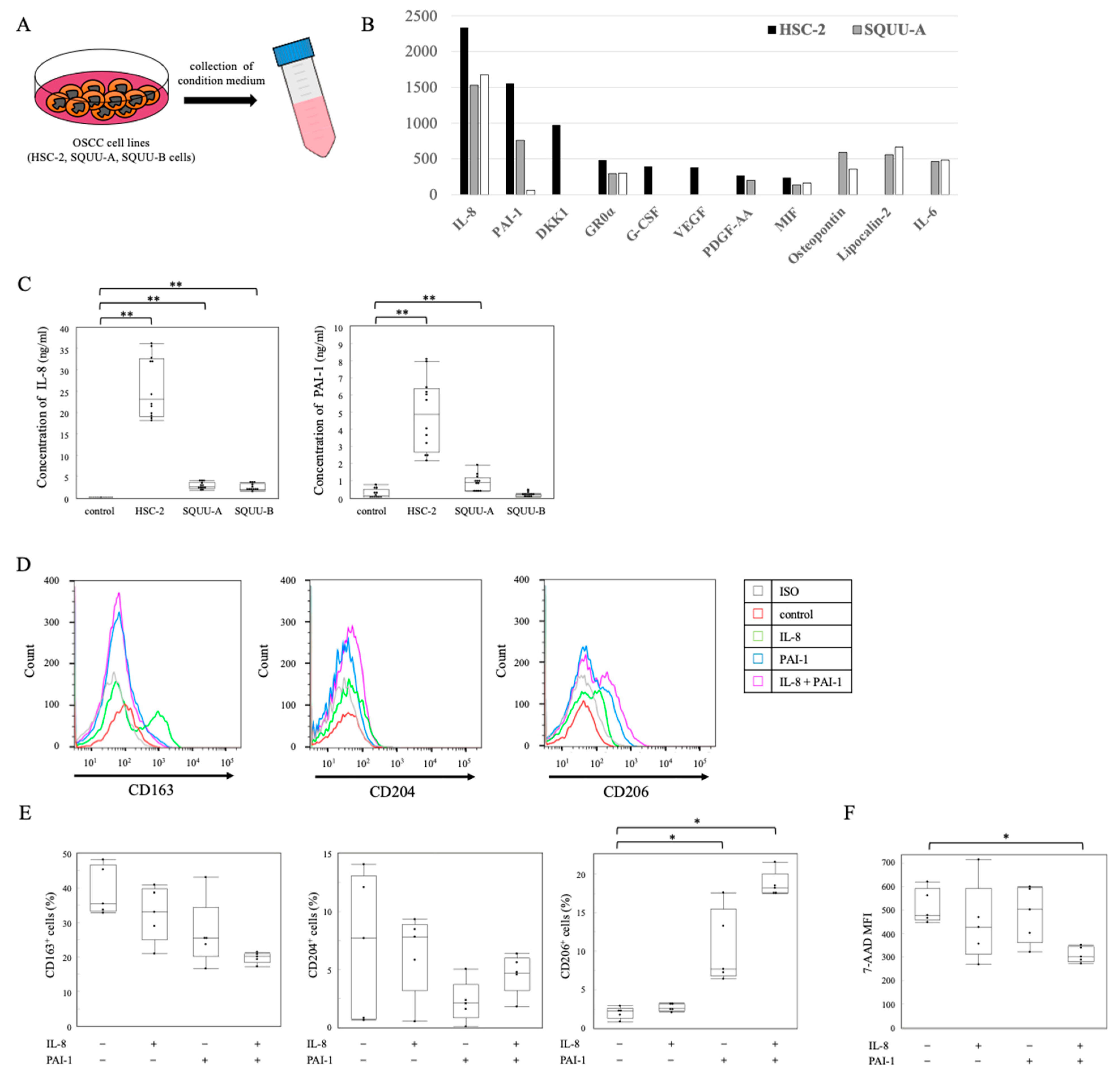
2.3. Association of OSCC Cell Lines with Differentiation of Monocytes to TAMs
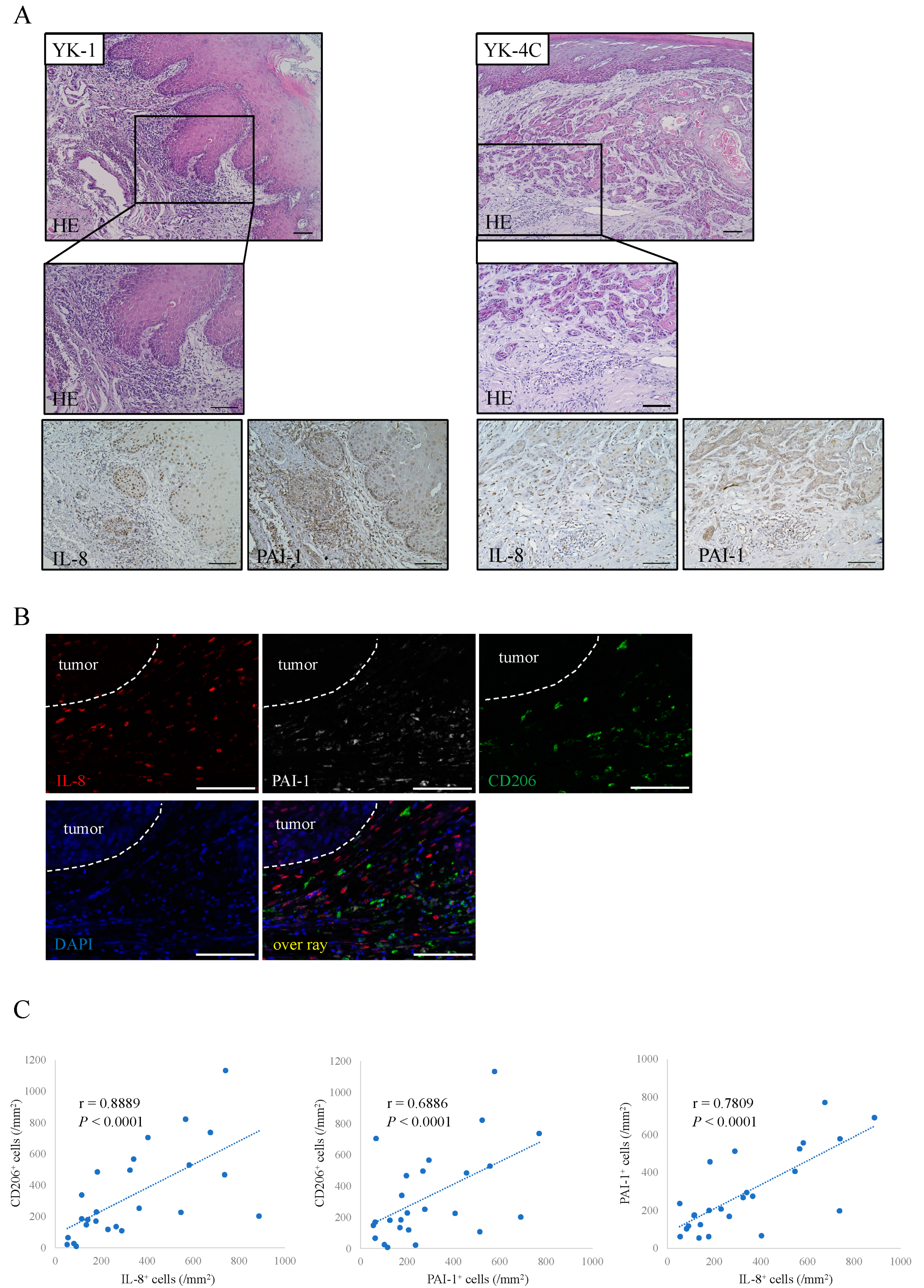
2.4. Distribution of IL-8 and PAI-1 in OSCC Tissues
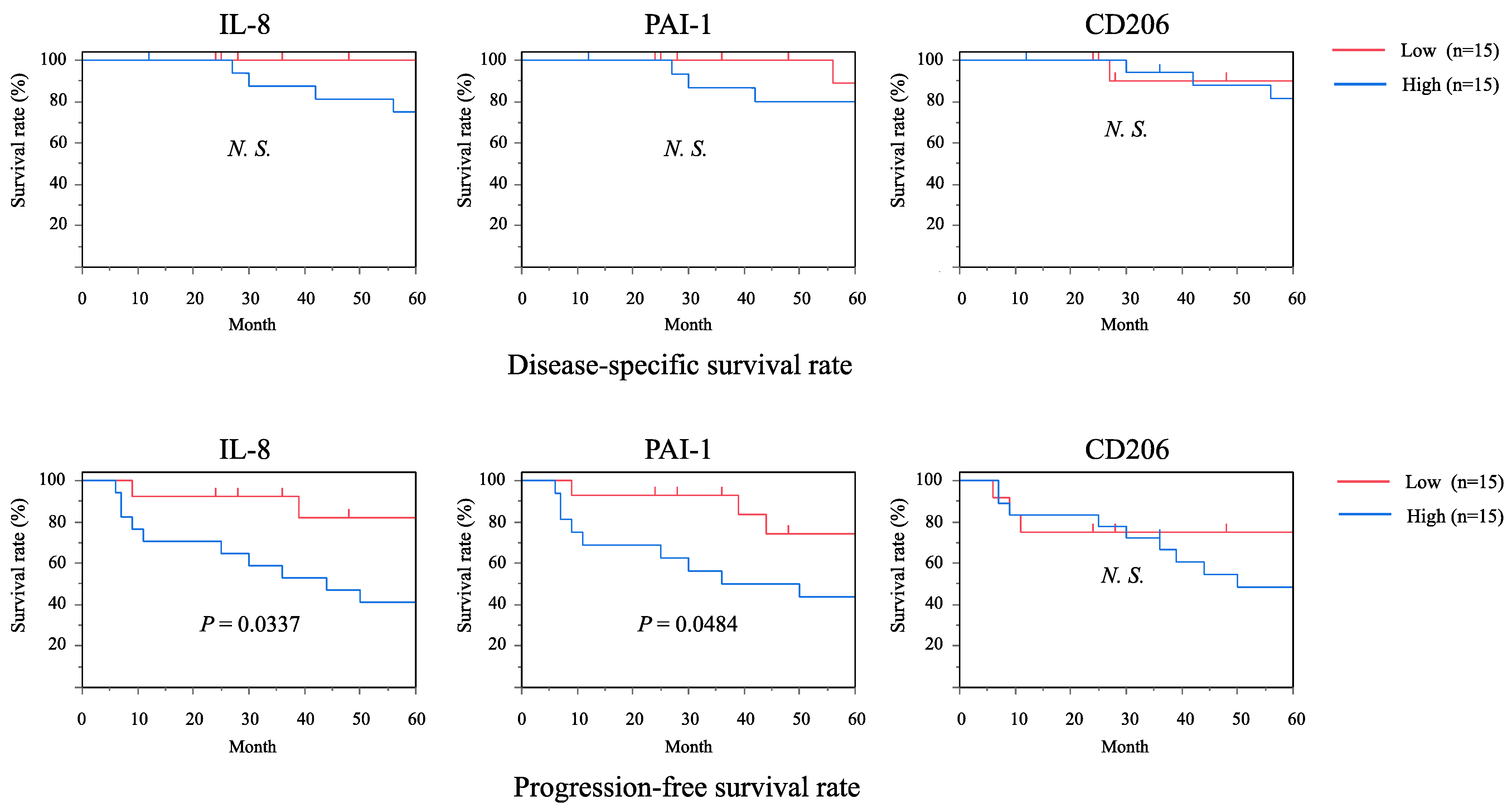
2.5. Associations of IL-8 and PAI-1 with Clinical Outcomes and Prognosis of OSCC Patients
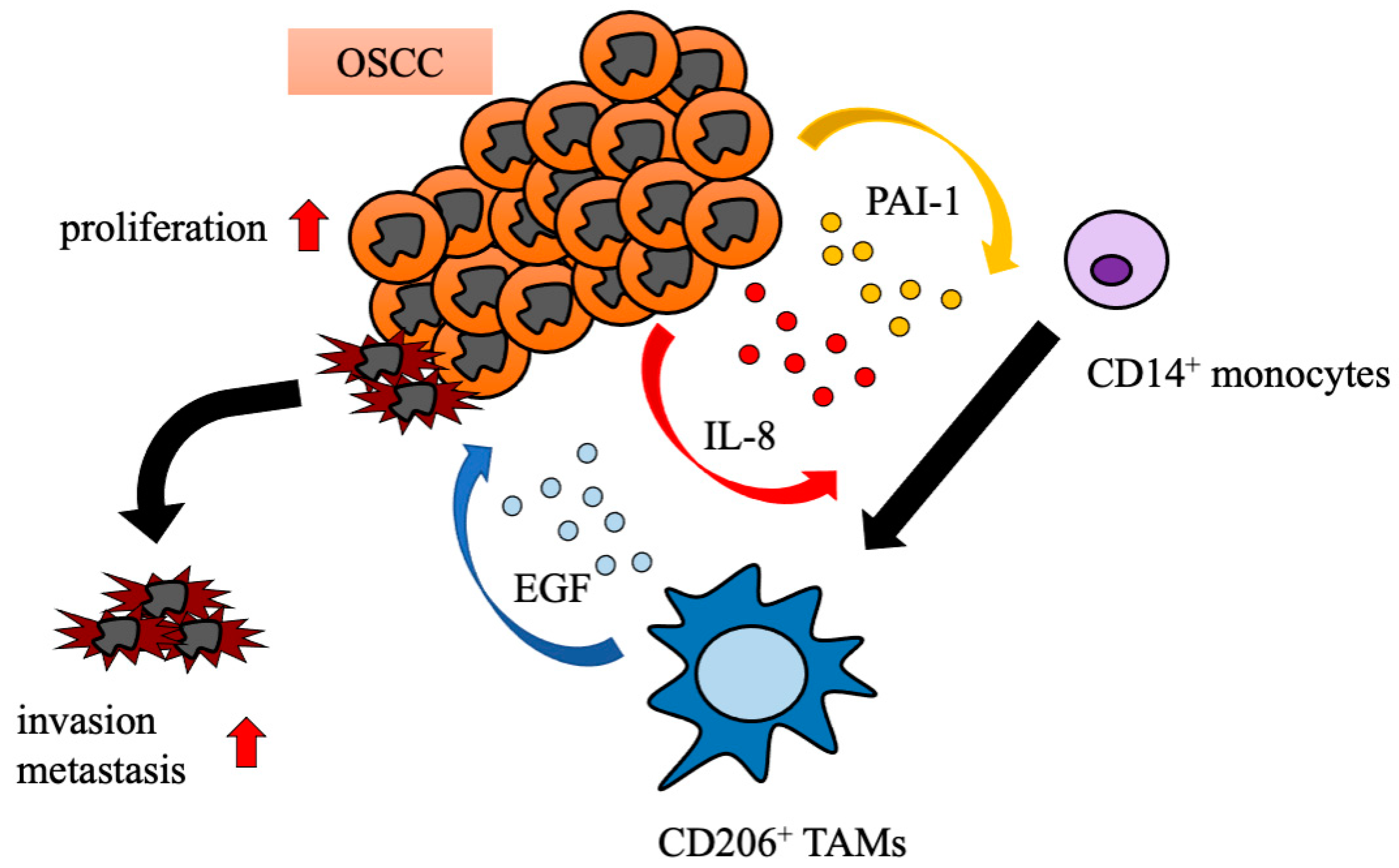
3. Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Principal Findings | References |
|---|---|
| CD206+ TAMs might play a key role in OSCC proliferation via EGF production | [23] |
| CAFs differentiate monocytes to TAMs via the tCXCL12/CXCR4 pathway and aid in the formation of CSC-like cells to enhance OSCC proliferation with reduced apoptosis | [28] |
| TAM and angiogenesis profiles imply a nonimmunosuppressive mechanism in young and elderly OSCC patients | [29] |
| PFKFB3 may promote angiogenesis in tumor progression and metastasis by regulating CD163+ TAM infiltration in OSCC | [30] |
| Infiltration by Tregs and M2 TAMs is associated with the progression of premalignant lesions to OSCC | [31] |
| CD163+- and CD8+-infiltrating cells influence early and subsequent stages of oral carcinogenesis | [32] |
| TGF-1β promotes OSCC-associated macrophages to secrete more VEGF via TβRII/Smad3 signaling | [33] |
| CD163+/CD204+ TAMs may affect OSCC invasion and metastasis by regulating T cells via IL-10 and PD-L1 | [19] |
| CD163+ TAMs promote lymphangiogenesis by expressing VEGF-C, which contributes to regional lymph node metastasis in OSCC | [34] |
| CAFs shape the OSCC immunosuppressive microenvironment by including the TAM protumoral phenotype | [35] |
| OSCCs directly suppress antitumor T cell immunity by conditioning TAMs | [36] |
| CD11b+ myeloid cells and CD206+ M2 macrophages increase during human OSCC recurrence after radiotherapy | [37] |
| TAMs promote the EMT of cancer cells, thereby leading to the progression of oral cancer | [38] |
| MMP and TAM expressions are inversely related in OSCC primary and metastatic regions | [39] |
| Axl signaling of OSCC is involved in polarizing TAMs toward the M2 phenotype | [40] |
| CD163+ cells may be effective predictors of OSCC prognosis | [41] |
| CAFs and TAMs expression may help guide treatment decisions to improve survival of OSCC patients | [42] |
| IL-8 affects the generation of CD163+ M2 macrophages in OSCC, which produces immune-suppressive cytokines such as IL-10 | [43] |
| TAMs have a protumor function in OSCC and likely promote tumor progression by activating Gas6/Axl-NF-κB signaling | [44] |
| TAM markers are associated with CSC markers and OSCC overall survival, suggesting their potential prognostic value in OSCC | [45] |
| TAMs and angiogenesis affect different histological grades of OSCC | [46] |
| CAFs and CD163+ macrophages may be potential prognostic predictors of OSCC | [47] |
| Infiltrated TAMs in OSCC have a M2 phenotype and may affect OSCC development and progression | [48] |
| Increased TAMs are associated with angiogenesis and higher histopathological grades in oral cancer | [49] |
| The presence of CD163 expression in oral tongue squamous cell carcinomas was associated with worse disease-free survival | [50] |
4. Materials and Methods
4.1. Isolation of Peripheral Blood Monocytes
4.2. Co-Culture of Monocytes and OSCC Cell Lines
4.3. Flow Cytometric Analysis
4.4. Cytokine Array
4.5. Enzyme-Linked Immunosorbent Assay (ELISA)
4.6. Cell Culture and Stimulation of Human Monocytes In Vitro
4.7. Immunohistochemical Analysis
4.8. Triple Immunofluorescence Analysis
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Quan, J.; Johnson, N.; Zhou, G.-B.; Parsons, P.; Boyle, G.; Gao, J. Potential molecular targets for inhibiting bone invasion by oral squamous cell carcinoma: A review of mechanisms. Cancer Metastasis Rev. 2011, 31, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Petersen, P.E. Oral cancer prevention and control—The approach of the World Health Organization. Oral Oncol 2009, 45, 454–460. [Google Scholar] [CrossRef]
- Anzai, H.; Yoshimoto, S.; Okamura, K.; Hiraki, A.; Hashimoto, S. IDO1-mediated Trp-kynurenine-AhR signal activation induces stemness and tumor dormancy in oral squamous cell carcinomas. Oral Sci. Int. 2021. [Google Scholar] [CrossRef]
- Meng, X.; Mori, T.; Matsumoto, F.; Miura, N.; Onidani, K.; Kobayashi, K.; Matsuzaki, Y.; Yoshimoto, S.; Ikeda, K.; Honda, K. Hepatitis B X-interacting protein, involved in increasing proliferation and cell migration, is a prognostic marker in head and neck squamous cell carcinoma. Oral Sci. Int. 2021. [Google Scholar] [CrossRef]
- Pollard, J.W. Tumour-educated macrophages promote tumour progression and metastasis. Nat. Rev. Cancer 2004, 4, 71–78. [Google Scholar] [CrossRef]
- Mantovani, A.; Sozzani, S.; Locati, M.; Allavena, P.; Sica, A. Macrophage polarization: Tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol. 2002, 23, 549–555. [Google Scholar] [CrossRef]
- Mills, C.D. M1 and M2 Macrophages: Oracles of Health and Disease. Crit. Rev. Immunol. 2012, 32, 463–488. [Google Scholar] [CrossRef] [Green Version]
- Gordon, S. Alternative activation of macrophages. Nat. Rev. Immunol. 2003, 3, 23–35. [Google Scholar] [CrossRef]
- Wynn, T.A.; Chawla, A.; Pollard, J.W. Macrophage biology in development, homeostasis and disease. Nature 2013, 496, 445–455. [Google Scholar] [CrossRef]
- Fujimura, T.; Kambayashi, Y.; Fujisawa, Y.; Hidaka, T.; Aiba, S. Tumor-Associated Macrophages: Therapeutic Targets for Skin Cancer. Front. Oncol. 2018, 8, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwadlo-Klarwasser, G.; Neubert, R.; Stahlmann, R.; Schmutzler, W. Influence of dexamethasone on the RM 3/1-positive macrophages in the peripheral blood and tissues of a New World monkey (the marmoset Callithrix jacchus). Int. Arch. Allergy Immunol. 1992, 97, 178–180. [Google Scholar] [CrossRef] [PubMed]
- Fabriek, B.O.; Dijkstra, C.D.; van den Berg, T.K. The macrophage scavenger receptor CD163. Immunobiology 2005, 210, 153–160. [Google Scholar] [CrossRef]
- Komohara, Y.; Niino, D.; Saito, Y.; Ohnishi, K.; Horlad, H.; Ohshima, K.; Takeya, M. Clinical significance of CD 163 + tumor-associated macrophages in patients with adult T-cell leukemia/lymphoma. Cancer Sci. 2013, 104, 945–951. [Google Scholar] [CrossRef] [PubMed]
- Berwin, B.; Hart, J.P.; Rice, S.; Gass, C.; Pizzo, S.V.; Post, S.; Nicchitta, C.V. Scavenger receptor-A mediates gp96/GRP94 and calreticulin internalization by antigen-presenting cells. EMBO J. 2003, 22, 6127–6136. [Google Scholar] [CrossRef]
- Krieger, M. The other side of scavenger receptors: Pattern recognition for host defense. Curr. Opin. Lipidol. 1997, 8, 275–280. [Google Scholar] [CrossRef]
- Ohnishi, K.; Komohara, Y.; Fujiwara, Y.; Takemura, K.; Lei, X.; Nakagawa, T.; Sakashita, N.; Takeya, M. Suppression of TLR4-mediated inflammatory response by macrophage class A scavenger receptor (CD204). Biochem. Biophys. Res. Commun. 2011, 411, 516–522. [Google Scholar] [CrossRef] [PubMed]
- Platt, N.; Gordon, S. Is the class A macrophage scavenger receptor (SR-A) multifunctional?—The mouse’s tale. J. Clin. Investig. 2001, 108, 649–654. [Google Scholar] [CrossRef] [PubMed]
- Kubota, K.; Moriyama, M.; Furukawa, S.; Rafiul, H.; Maruse, Y.; Jinno, T.; Tanaka, A.; Ohta, M.; Ishiguro, N.; Yamauchi, M.; et al. CD163(+)CD204(+) tumor-associated macrophages contribute to T cell regulation via interleukin-10 and PD-L1 production in oral squamous cell carcinoma. Sci. Rep. 2017, 7, 1755. [Google Scholar] [CrossRef]
- Azad, A.K.; Rajaram, M.V.S.; Schlesinger, L.S. Exploitation of the Macrophage Mannose Receptor (CD206) in Infectious Disease Diagnostics and Therapeutics. J. Cytol. Mol. Biol. 2014, 1, 1000003. [Google Scholar] [CrossRef]
- Hu, W.; Qian, Y.; Yu, F.; Liu, W.; Wu, Y.; Fang, X.; Hao, W. Alternatively activated macrophages are associated with metastasis and poor prognosis in prostate adenocarcinoma. Oncol. Lett. 2015, 10, 1390–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, P.; Ma, L.; Liu, L.; Zhao, G.; Zhang, S.; Dong, L.; Xue, R.; Chen, S. CD86+/CD206+, Diametrically Polarized Tumor-Associated Macrophages, Predict Hepatocellular Carcinoma Patient Prognosis. Int. J. Mol. Sci. 2016, 17, 320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haque, A.; Moriyama, M.; Kubota, K.; Ishiguro, N.; Sakamoto, M.; Chinju, A.; Mochizuki, K.; Sakamoto, T.; Kaneko, N.; Munemura, R.; et al. CD206(+) tumor-associated macrophages promote proliferation and invasion in oral squamous cell carcinoma via EGF production. Sci. Rep. 2019, 9, 14611. [Google Scholar] [CrossRef] [PubMed]
- Kelly, F.L.; Weinberg, K.E.; Nagler, A.E.; Nixon, A.B.; Star, M.D.; Todd, J.L.; Brass, D.M.; Palmer, S.M. EGFR-Dependent IL8 Production by Airway Epithelial Cells After Exposure to the Food Flavoring Chemical 2,3-Butanedione. Toxicol. Sci. 2019, 169, 534–542. [Google Scholar] [CrossRef]
- Freytag, J.; Wilkins-Port, C.E.; Higgins, C.E.; Higgins, S.P.; Samarakoon, R.; Higgins, P.J. PAI-1 mediates the TGF-beta1+EGF-induced “scatter” response in transformed human keratinocytes. J. Investig. Dermatol. 2010, 130, 2179–2190. [Google Scholar] [CrossRef] [Green Version]
- Qian, B.-Z.; Pollard, J.W. Macrophage Diversity Enhances Tumor Progression and Metastasis. Cell 2010, 141, 39–51. [Google Scholar] [CrossRef] [Green Version]
- Komohara, Y.; Jinushi, M.; Takeya, M. Clinical significance of macrophage heterogeneity in human malignant tumors. Cancer Sci. 2013, 105, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Bu, W.; Meng, L.; Liu, X.; Wang, S.; Jiang, L.; Ren, M.; Fan, Y.; Sun, H. CXCL12/CXCR4 pathway orchestrates CSC-like properties by CAF recruited tumor associated macrophage in OSCC. Exp. Cell Res. 2019, 378, 131–138. [Google Scholar] [CrossRef]
- Teixeira, L.R.; Almeida, L.Y.; Silva, R.N.; Mesquita, A.T.M.; Colturato, C.B.N.; Silveira, H.A.; Duarte, A.; Ribeiro-Silva, A.; Leon, J.E. Young and elderly oral squamous cell carcinoma patients present similar angiogenic profile and predominance of M2 macrophages: Comparative immunohistochemical study. Head Neck 2019, 41, 4111–4120. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-J.; Mao, X.-H.; Tian, T.; Wang, W.-M.; Su, T.; Jiang, C.-H.; Hu, C.-Y. Role of PFKFB3 and CD163 in Oral Squamous Cell Carcinoma Angiogenesis. Curr. Med. Sci. 2019, 39, 410–414. [Google Scholar] [CrossRef]
- Kouketsu, A.; Sato, I.; Oikawa, M.; Shimizu, Y.; Saito, H.; Tashiro, K.; Yamashita, Y.; Takahashi, T.; Kumamoto, H. Regulatory T cells and M2-polarized tumour-associated macrophages are associated with the oncogenesis and progression of oral squamous cell carcinoma. Int. J. Oral Maxillofac. Surg. 2019, 48, 1279–1288. [Google Scholar] [CrossRef]
- Stasikowska-Kanicka, O.; Wągrowska-Danilewicz, M.; Danilewicz, M. CD8+ and CD163+ infiltrating cells and PD-L1 immunoexpression in oral leukoplakia and oral carcinoma. APMIS 2018, 126, 732–738. [Google Scholar] [CrossRef]
- Sun, H.; Miao, C.; Liu, W.; Qiao, X.; Yang, W.; Li, L.; Li, C. TGF-beta1/TbetaRII/Smad3 signaling pathway promotes VEGF expression in oral squamous cell carcinoma tumor-associated macrophages. Biochem. Biophys. Res. Commun. 2018, 497, 583–590. [Google Scholar] [CrossRef]
- Yamagata, Y.; Tomioka, H.; Sakamoto, K.; Sato, K.; Harada, H.; Ikeda, T.; Kayamori, K. CD163-Positive Macrophages Within the Tumor Stroma Are Associated with Lymphangiogenesis and Lymph Node Metastasis in Oral Squamous Cell Carcinoma. J. Oral Maxillofac. Surg. 2017, 75, 2144–2153. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Sakakura, K.; Kudo, T.; Toyoda, M.; Kaira, K.; Oyama, T.; Chikamatsu, K. Cancer-associated fibroblasts promote an immunosuppressive microenvironment through the induction and accumulation of protumoral macrophages. Oncotarget 2016, 8, 8633–8647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, C.; Yuan, F.; Wang, J.; Wu, L. Oral squamous cell carcinoma suppressed antitumor immunity through induction of PD-L1 expression on tumor-associated macrophages. Immunobiology 2017, 222, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Okubo, M.; Kioi, M.; Nakashima, H.; Sugiura, K.; Mitsudo, K.; Aoki, I.; Taniguchi, H.; Tohnai, I. M2-polarized macro-phages contribute to neovasculogenesis, leading to relapse of oral cancer following radiation. Sci. Rep. 2016, 6, 27548. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; He, M.Y.; Zhu, L.F.; Yang, C.C.; Zhou, M.L.; Wang, Q.; Zhang, W.; Zheng, Y.Y.; Wang, D.M.; Xu, Z.Q.; et al. Tumor-associated macrophages correlate with the clinicopathological features and poor outcomes via inducing epithelial to mesenchymal transition in oral squamous cell carcinoma. J. Exp. Clin. Cancer Res. 2016, 35, 12. [Google Scholar] [CrossRef] [Green Version]
- Nishio, K.; Motozawa, K.; Omagari, D.; Gojoubori, T.; Ikeda, T.; Asano, M.; Gionhaku, N. Comparison of MMP2 and MMP9 expression levels between primary and metastatic regions of oral squamous cell carcinoma. J. Oral Sci. 2016, 58, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Chiu, K.-C.; Lee, C.-H.; Liu, S.-Y.; Chou, Y.-T.; Huang, R.-Y.; Huang, S.-M.; Shieh, Y.-S. Polarization of tumor-associated macrophages and Gas6/Axl signaling in oral squamous cell carcinoma. Oral Oncol. 2015, 51, 683–689. [Google Scholar] [CrossRef]
- Wang, S.; Sun, M.; Gu, C.; Wang, X.; Chen, D.; Zhao, E.; Jiao, X.; Zheng, J. Expression of CD163, interleukin-10, and interferon-gamma in oral squamous cell carcinoma: Mutual relationships and prognostic implications. Eur. J. Oral Sci. 2014, 122, 202–209. [Google Scholar] [CrossRef]
- Matsuoka, Y.; Yoshida, R.; Nakayama, H.; Nagata, M.; Hirosue, A.; Tanaka, T.; Kawahara, K.; Nakagawa, Y.; Sakata, J.; Arita, H.; et al. The tumour stromal features are associated with resistance to 5-FU-based chemoradiotherapy and a poor prognosis in patients with oral squamous cell carcinoma. APMIS 2015, 123, 205–214. [Google Scholar] [CrossRef]
- Fujita, Y.; Okamoto, M.; Goda, H.; Tano, T.; Nakashiro, K.-I.; Sugita, A.; Fujita, T.; Koido, S.; Homma, S.; Kawakami, Y.; et al. Prognostic Significance of Interleukin-8 and CD163-Positive Cell-Infiltration in Tumor Tissues in Patients with Oral Squamous Cell Carcinoma. PLoS ONE 2014, 9, e110378. [Google Scholar] [CrossRef]
- Lee, C.H.; Liu, S.Y.; Chou, K.C.; Yeh, C.T.; Shiah, S.G.; Huang, R.Y.; Cheng, J.C.; Yen, C.Y.; Shieh, Y.S. Tumor-associated macrophages promote oral cancer progression through activation of the Axl signaling pathway. Ann. Surg. Oncol. 2014, 21, 1031–1037. [Google Scholar] [CrossRef]
- He, K.-F.; Zhang, L.; Huang, C.-F.; Ma, S.-R.; Wang, Y.-F.; Wang, W.-M.; Zhao, Z.; Liu, B.; Zhao, Y.-F.; Zhang, W.-F.; et al. CD163+ Tumor-Associated Macrophages Correlated with Poor Prognosis and Cancer Stem Cells in Oral Squamous Cell Carcinoma. BioMed Res. Int. 2014, 2014, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Boas, D.S.; Takiya, C.M.; Gurgel, C.A.; Cabral, M.G.; Santos, J.N. Tumor-infiltrating macrophage and microvessel density in oral squamous cell carcinoma. Braz. Dent. J. 2013, 24, 194–199. [Google Scholar] [CrossRef]
- Fujii, N.; Shomori, K.; Shiomi, T.; Nakabayashi, M.; Takeda, C.; Ryoke, K.; Ito, H. Cancer-associated fibroblasts and CD163-positive macrophages in oral squamous cell carcinoma: Their clinicopathological and prognostic significance. J. Oral Pathol. Med. 2012, 41, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Mori, K.; Hiroi, M.; Shimada, J.; Ohmori, Y. Infiltration of m2 tumor-associated macrophages in oral squamous cell carcinoma correlates with tumor malignancy. Cancers 2011, 3, 3726–3739. [Google Scholar] [CrossRef] [PubMed]
- El-Rouby, D.H. Association of macrophages with angiogenesis in oral verrucous and squamous cell carcinomas. J. Oral Pathol. Med. 2010, 39, 559–564. [Google Scholar] [CrossRef]
- Agarbati, S.; Mascitti, M.; Paolucci, E.; Togni, L.; Santarelli, A.; Rubini, C.; Fazioli, F. Prognostic Relevance of Macrophage Phenotypes in High-grade Oral Tongue Squamous Cell Carcinomas. Appl. Immunohistochem. Mol. Morphol. 2021, 29, 359–365. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kai, K.; Moriyama, M.; Haque, A.S.M.R.; Hattori, T.; Chinju, A.; Hu, C.; Kubota, K.; Miyahara, Y.; Kakizoe-Ishiguro, N.; Kawano, S.; et al. Oral Squamous Cell Carcinoma Contributes to Differentiation of Monocyte-Derived Tumor-Associated Macrophages via PAI-1 and IL-8 Production. Int. J. Mol. Sci. 2021, 22, 9475. https://doi.org/10.3390/ijms22179475
Kai K, Moriyama M, Haque ASMR, Hattori T, Chinju A, Hu C, Kubota K, Miyahara Y, Kakizoe-Ishiguro N, Kawano S, et al. Oral Squamous Cell Carcinoma Contributes to Differentiation of Monocyte-Derived Tumor-Associated Macrophages via PAI-1 and IL-8 Production. International Journal of Molecular Sciences. 2021; 22(17):9475. https://doi.org/10.3390/ijms22179475
Chicago/Turabian StyleKai, Kazuki, Masafumi Moriyama, A. S. M. Rafiul Haque, Taichi Hattori, Akira Chinju, Chen Hu, Keigo Kubota, Yuka Miyahara, Noriko Kakizoe-Ishiguro, Shintaro Kawano, and et al. 2021. "Oral Squamous Cell Carcinoma Contributes to Differentiation of Monocyte-Derived Tumor-Associated Macrophages via PAI-1 and IL-8 Production" International Journal of Molecular Sciences 22, no. 17: 9475. https://doi.org/10.3390/ijms22179475
APA StyleKai, K., Moriyama, M., Haque, A. S. M. R., Hattori, T., Chinju, A., Hu, C., Kubota, K., Miyahara, Y., Kakizoe-Ishiguro, N., Kawano, S., & Nakamura, S. (2021). Oral Squamous Cell Carcinoma Contributes to Differentiation of Monocyte-Derived Tumor-Associated Macrophages via PAI-1 and IL-8 Production. International Journal of Molecular Sciences, 22(17), 9475. https://doi.org/10.3390/ijms22179475