
Could the Epigenetics of Eosinophils in Asthma and Allergy Solve Parts of the Puzzle?



Abstract
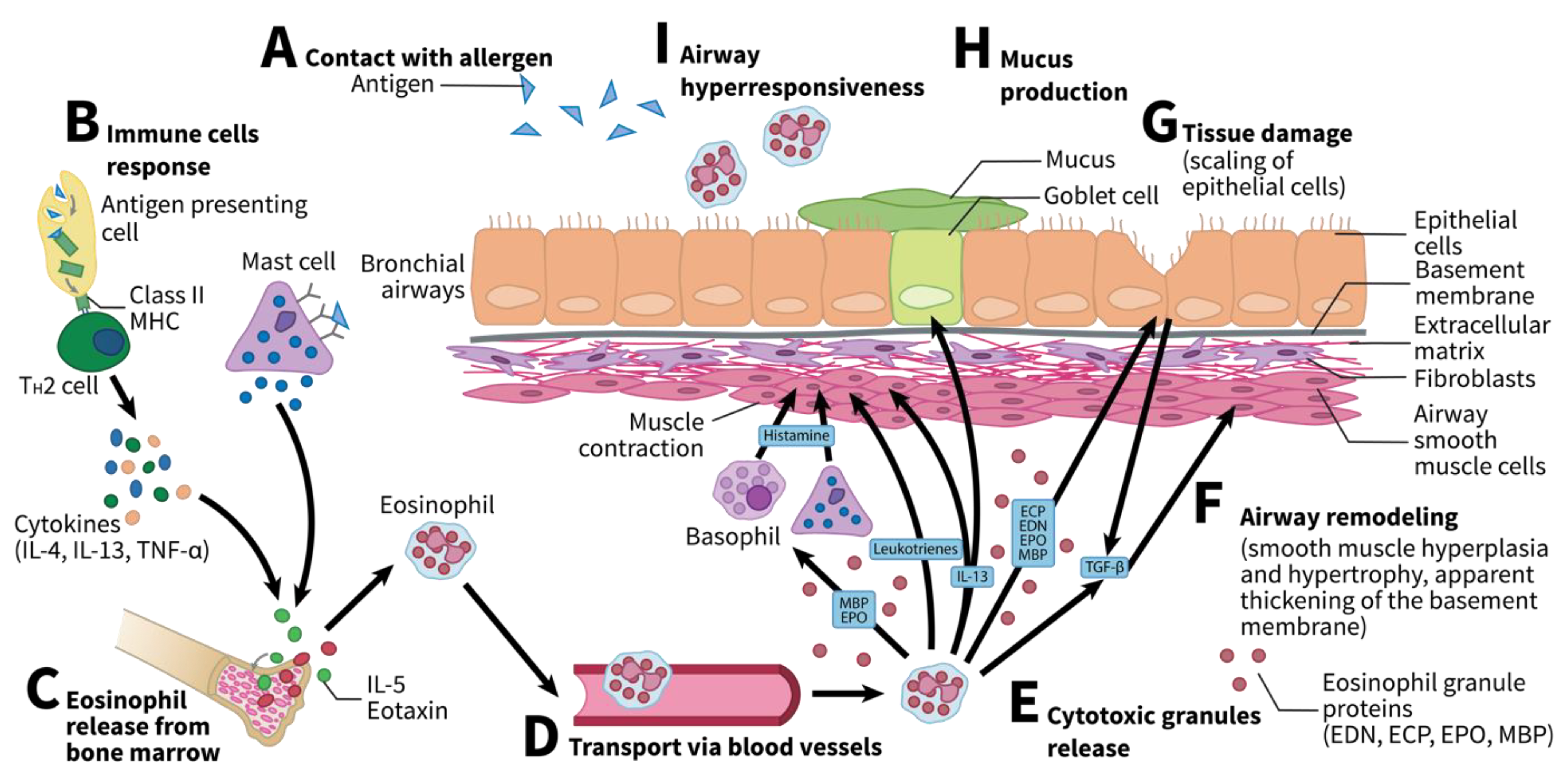
:1. Introduction
2. Eosinophils in Human Diseases
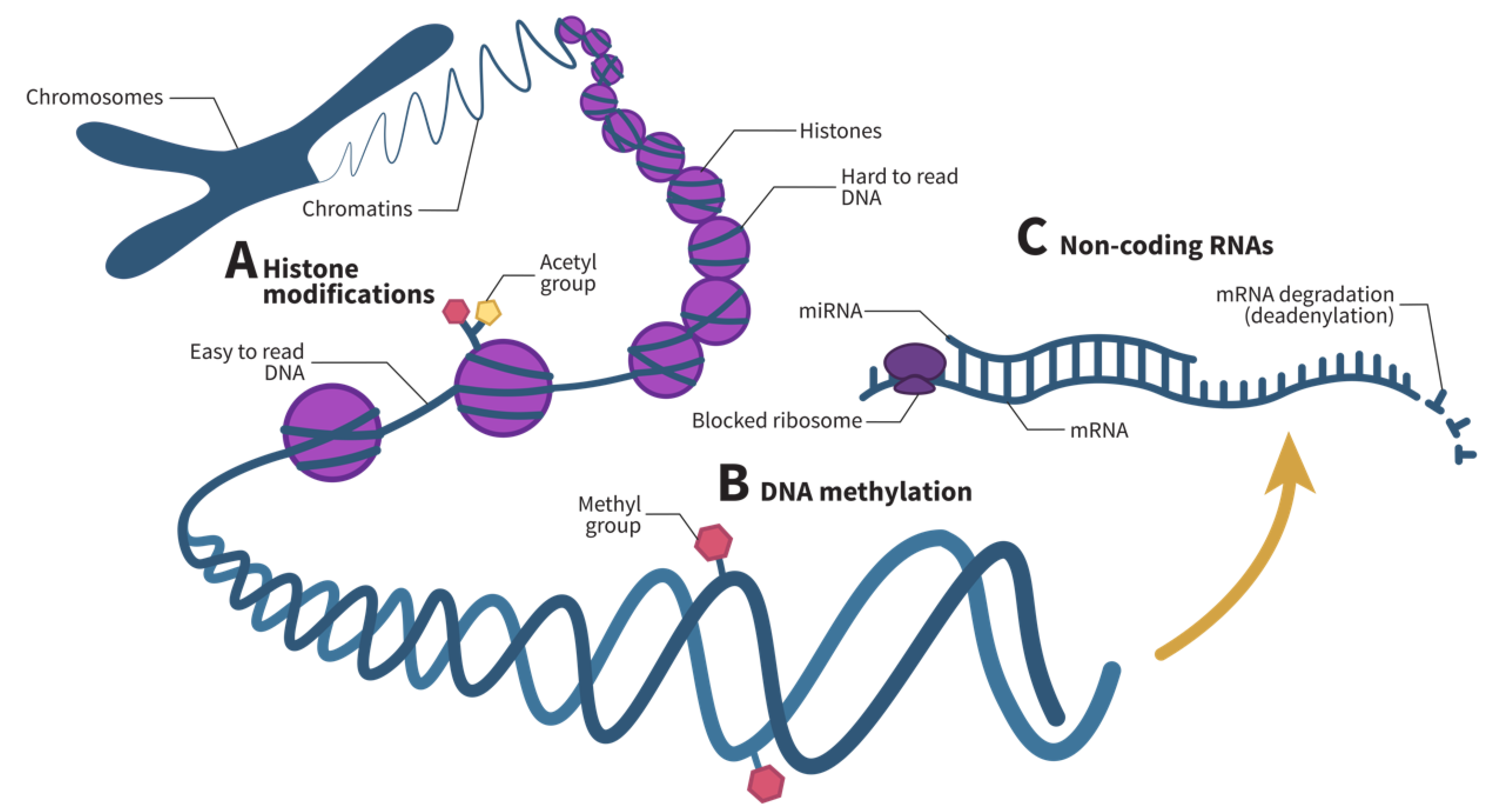
3. Epigenetic Mechanisms
4. Methylation Profile of Eosinophils
5. microRNA Profile of Eosinophils
6. Other Epigenetic Mechanisms of Eosinophils
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mims, J.W. Asthma: Definitions and pathophysiology. Int. Forum. Allergy. Rhinol. 2015, 5, S2–S6. [Google Scholar] [CrossRef]
- Kay, A.B.; Corrigan, C.J. Asthma. Eosinophils and neutrophils. Br. Med. Bull. 1992, 48, 51–64. [Google Scholar] [CrossRef] [PubMed]
- Kay, A.B. The role of eosinophils in the pathogenesis of asthma. Trends Mol. Med. 2005, 11, 148–152. [Google Scholar] [CrossRef]
- Zhang, L.; Lu, Q.; Chang, C. Epigenetics in Health and Disease. Adv. Exp. Med. Biol. 2020, 1253, 3–55. [Google Scholar] [CrossRef] [PubMed]
- DeVries, A.; Vercelli, D. Epigenetic Mechanisms in Asthma. Ann. Am. Thorac. Soc. 2016, 13, S48–S50. [Google Scholar] [CrossRef] [PubMed]
- Gleich, G.J. Historical Overview and Perspective on the Role of the Eosinophil in Health and Disease; Academic Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Jacobsen, E.A.; Helmers, R.A.; Lee, J.J.; Lee, N.A. The expanding role(s) of eosinophils in health and disease. Blood 2012, 120, 3882–3890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bochner, B.S. The eosinophil: For better or worse, in sickness and in health. Ann. Allergy Asthma Immunol. 2018, 121, 150–155. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.J.; Jacobsen, E.A.; McGarry, M.P.; Schleimer, R.P.; Lee, N.A. Eosinophils in health and disease: The LIAR hypothesis. Clin. Exp. Allergy 2010, 40, 563–575. [Google Scholar] [CrossRef] [Green Version]
- O’Connell, A.E.; Hess, J.A.; Santiago, G.A.; Nolan, T.J.; Lok, J.B.; Lee, J.J.; Abraham, D. Major basic protein from eosinophils and myeloperoxidase from neutrophils are required for protective immunity to Strongyloides stercoralis in mice. Infect. Immun. 2011, 79, 2770–2778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCormick, M.L.; Metwali, A.; Railsback, M.A.; Weinstock, J.V.; Britigan, B.E. Eosinophils from schistosome-induced hepatic granulomas produce superoxide and hydroxyl radical. J. Immunol. 1996, 157, 5009–5015. [Google Scholar]
- Liao, W.; Long, H.; Chang, C.C.; Lu, Q. The Eosinophil in Health and Disease: From Bench to Bedside and Back. Clin. Rev. Allergy Immunol. 2016, 50, 125–139. [Google Scholar] [CrossRef] [PubMed]
- Fainardi, V.; Saglani, S. The need to differentiate between adults and children when treating severe asthma. Expert Rev. Respir. Med. 2015, 9, 419–428. [Google Scholar] [CrossRef] [PubMed]
- McBrien, C.N.; Menzies-Gow, A. The Biology of Eosinophils and Their Role in Asthma. Front. Med. 2017, 4, 93. [Google Scholar] [CrossRef] [PubMed]
- Gundel, R.H.; Letts, L.G.; Gleich, G.J. Human eosinophil major basic protein induces airway constriction and airway hyperresponsiveness in primates. J. Clin. Investig. 1991, 87, 1470–1473. [Google Scholar] [CrossRef] [PubMed]
- Coyle, A.J.; Ackerman, S.J.; Burch, R.; Proud, D.; Irvin, C.G. Human eosinophil-granule major basic protein and synthetic polycations induce airway hyperresponsiveness in vivo dependent on bradykinin generation. J. Clin. Investig. 1995, 95, 1735–1740. [Google Scholar] [CrossRef] [PubMed]
- Piliponsky, A.M.; Gleich, G.J.; Nagler, A.; Bar, I.; Levi-Schaffer, F. Non-IgE-dependent activation of human lung- and cord blood-derived mast cells is induced by eosinophil major basic protein and modulated by the membrane form of stem cell factor. Blood 2003, 101, 1898–1904. [Google Scholar] [CrossRef]
- Ben-Zimra, M.; Bachelet, I.; Seaf, M.; Gleich, G.J.; Levi-Schaffer, F. Eosinophil major basic protein activates human cord blood mast cells primed with fibroblast membranes by integrin-β1. Allergy 2013, 68, 1259–1268. [Google Scholar] [CrossRef]
- Grünig, G.; Warnock, M.; Wakil, A.E.; Venkayya, R.; Brombacher, F.; Rennick, D.M.; Sheppard, D.; Mohrs, M.; Donaldson, D.D.; Locksley, R.M.; et al. Requirement for IL-13 independently of IL-4 in experimental asthma. Science 1998, 282, 2261–2263. [Google Scholar] [CrossRef] [Green Version]
- Hallstrand, T.S.; Henderson, W.R., Jr. An update on the role of leukotrienes in asthma. Curr. Opin. Allergy Clin. Immunol. 2010, 10, 60–66. [Google Scholar] [CrossRef] [Green Version]
- Frigas, E.; Gleich, G.J. The eosinophil and the pathophysiology of asthma. J. Allergy Clin. Immunol. 1986, 77, 527–537. [Google Scholar] [CrossRef]
- Slifman, N.R.; Loegering, D.A.; McKean, D.J.; Gleich, G.J. Ribonuclease activity associated with human eosinophil-derived neurotoxin and eosinophil cationic protein. J. Immunol. 1986, 137, 2913–2917. [Google Scholar]
- Navarro, S.; Aleu, J.; Jiménez, M.; Boix, E.; Cuchillo, C.M.; Nogués, M.V. The cytotoxicity of eosinophil cationic protein/ribonuclease 3 on eukaryotic cell lines takes place through its aggregation on the cell membrane. Cell Mol. Life Sci. 2008, 65, 324–337. [Google Scholar] [CrossRef]
- Durack, D.T.; Ackerman, S.J.; Loegering, D.A.; Gleich, G.J. Purification of human eosinophil-derived neurotoxin. Proc. Natl. Acad. Sci. USA 1981, 78, 5165–5169. [Google Scholar] [CrossRef] [Green Version]
- Fredens, K.; Dahl, R.; Venge, P. The Gordon phenomenon induced by the eosinophil cationic protein and eosinophil protein X. J. Allergy Clin. Immunol. 1982, 70, 361–366. [Google Scholar] [CrossRef]
- van Dalen, C.J.; Kettle, A.J. Substrates and products of eosinophil peroxidase. Biochem. J. 2001, 358, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Birkland, T.P.; Cheavens, M.D.; Pincus, S.H. Human eosinophils stimulate DNA synthesis and matrix production in dermal fibroblasts. Arch. Dermatol. Res. 1994, 286, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Makinde, T.; Murphy, R.F.; Agrawal, D.K. The regulatory role of TGF-beta in airway remodeling in asthma. Immunol. Cell Biol. 2007, 85, 348–356. [Google Scholar] [CrossRef] [PubMed]
- Levi-Schaffer, F.; Garbuzenko, E.; Rubin, A.; Reich, R.; Pickholz, D.; Gillery, P.; Emonard, H.; Nagler, A.; Maquart, F.A. Human eosinophils regulate human lung- and skin-derived fibroblast properties in vitro: A role for transforming growth factor beta (TGF-beta). Proc. Natl. Acad. Sci. USA 1999, 96, 9660–9665. [Google Scholar] [CrossRef] [Green Version]
- Possa, S.S.; Leick, E.A.; Prado, C.M.; Martins, M.A.; Tibério, I.F. Eosinophilic inflammation in allergic asthma. Front. Pharmacol. 2013, 4, 46. [Google Scholar] [CrossRef] [Green Version]
- Green, R.H.; Brightling, C.E.; McKenna, S.; Hargadon, B.; Parker, D.; Bradding, P.; Wardlaw, A.J.; Pavord, I.D. Asthma exacerbations and sputum eosinophil counts: A randomised controlled trial. Lancet 2002, 360, 1715–1721. [Google Scholar] [CrossRef]
- Price, D.B.; Rigazio, A.; Campbell, J.D.; Bleecker, E.R.; Corrigan, C.J.; Thomas, M.; Wenzel, S.E.; Wilson, A.M.; Small, M.B.; Gopalan, G.; et al. Blood eosinophil count and prospective annual asthma disease burden: A UK cohort study. Lancet Respir. Med. 2015, 3, 849–858. [Google Scholar] [CrossRef]
- Denlinger, L.C.; Phillips, B.R.; Ramratnam, S.; Ross, K.; Bhakta, N.R.; Cardet, J.C.; Castro, M.; Peters, S.P.; Phipatanakul, W.; Aujla, S.; et al. Inflammatory and Comorbid Features of Patients with Severe Asthma and Frequent Exacerbations. Am. J. Respir. Crit. Care Med. 2017, 195, 302–313. [Google Scholar] [CrossRef] [PubMed]
- Nakagome, K.; Nagata, M. Involvement and possible role of eosinophils in asthma exacerbation. Front. Immunol. 2018, 9, 2220. [Google Scholar] [CrossRef] [PubMed]
- Kelly, E.A.; Esnault, S.; Liu, L.Y.; Evans, M.D.; Johansson, M.W.; Mathur, S.; Mosher, D.F.; Denlinger, L.C.; Jarjour, N.N. Mepolizumab Attenuates Airway Eosinophil Numbers, but Not Their Functional Phenotype, in Asthma. Am. J. Respir. Crit. Care Med. 2017, 196, 1385–1395. [Google Scholar] [CrossRef] [PubMed]
- Haldar, P.; Brightling, C.E.; Hargadon, B.; Gupta, S.; Monteiro, W.; Sousa, A.; Marshall, R.P.; Bradding, P.; Green, R.H.; Wardlaw, A.J.; et al. Mepolizumab and exacerbations of refractory eosinophilic asthma. N. Engl. J. Med. 2009, 360, 973–984. [Google Scholar] [CrossRef] [Green Version]
- Hanania, N.A.; Noonan, M.; Corren, J.; Korenblat, P.; Zheng, Y.; Fischer, S.K.; Cheu, M.; Putnam, W.S.; Murray, E.; Scheerens, H.; et al. Lebrikizumab in moderate-to-severe asthma: Pooled data from two randomised placebo-controlled studies. Thorax 2015, 70, 748–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wenzel, S.; Castro, M.; Corren, J.; Maspero, J.; Wang, L.; Zhang, B.; Pirozzi, G.; Sutherland, E.R.; Evans, R.R.; Joish, V.N.; et al. Dupilumab efficacy and safety in adults with uncontrolled persistent asthma despite use of medium-to-high-dose inhaled corticosteroids plus a long-acting β2 agonist: A randomised double-blind placebo-controlled pivotal phase 2b dose-ranging trial. Lancet 2016, 388, 31–44. [Google Scholar] [CrossRef]
- Bel, E.H.; Ten Brinke, A. New Anti-Eosinophil Drugs for Asthma and COPD: Targeting the Trait! Chest 2017, 152, 1276–1282. [Google Scholar] [CrossRef]
- Gauvreau, G.M.; O’Byrne, P.M.; Boulet, L.P.; Wang, Y.; Cockcroft, D.; Bigler, J.; FitzGerald, J.M.; Boedigheimer, M.; Davis, B.E.; Dias, C.; et al. Effects of an anti-TSLP antibody on allergen-induced asthmatic responses. N. Engl. J. Med. 2014, 370, 2102–2110. [Google Scholar] [CrossRef]
- Tammen, S.A.; Friso, S.; Choi, S.W. Epigenetics: The link between nature and nurture. Mol. Asp. Med. 2013, 34, 753–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.C.; Lu, Q. Epigenetics in Allergy and Autoimmunity; Springer: Singapore, 2020. [Google Scholar] [CrossRef]
- Zhang, R. The Epigenetics of Autoimmunity; Academic Press, An Imprint of Elsevier: London, UK, 2018. [Google Scholar]
- Portela, A.; Esteller, M. Epigenetic modifications and human disease. Nat. Biotechnol. 2010, 28, 1057–1068. [Google Scholar] [CrossRef] [PubMed]
- Milavetz, B.I.; Balakrishnan, L. Viral epigenetics. Methods Mol. Biol. 2015, 1238, 569–596. [Google Scholar] [CrossRef] [PubMed]
- Esteller, M. Epigenetic gene silencing in cancer: The DNA hypermethylome. Hum. Mol. Genet. 2007, 16, R50–R59. [Google Scholar] [CrossRef]
- Lopez-Serra, L.; Esteller, M. Proteins that bind methylated DNA and human cancer: Reading the wrong words. Br. J. Cancer 2008, 98, 1881–1885. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, A.; Rauch, T.A.; Todorov, I.; Ku, H.T.; Al-Abdullah, I.H.; Kandeel, F.; Mullen, Y.; Pfeifer, G.P.; Ferreri, K. Insulin gene expression is regulated by DNA methylation. PLoS ONE 2009, 4, e6953. [Google Scholar] [CrossRef]
- Calle-Fabregat, C.; Morante-Palacios, O.; Ballestar, E. Understanding the Relevance of DNA Methylation Changes in Immune Differentiation and Disease. Genes 2020, 11, 110. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Wu, W.; Chen, Q.; Chen, M. Non-Coding RNAs and their Integrated Networks. J. Integr. Bioinform. 2019, 16. [Google Scholar] [CrossRef] [PubMed]
- Mohr, A.M.; Mott, J.L. Overview of microRNA biology. Semin. Liver Dis. 2015, 35, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Gavrilov, K.; Saltzman, W.M. Therapeutic siRNA: Principles, challenges, and strategies. Yale J. Biol. Med. 2012, 85, 187–200. [Google Scholar]
- Holoch, D.; Moazed, D. RNA-mediated epigenetic regulation of gene expression. Nat. Rev. Genet. 2015, 16, 71–84. [Google Scholar] [CrossRef]
- Sartorelli, V.; Lauberth, S.M. Enhancer RNAs are an important regulatory layer of the epigenome. Nat. Struct. Mol. Biol. 2020, 27, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-C.; Lee, Y.-C.; Chang, K.-L.; Hsiao, K.-Y. Analysis of common targets for circular RNAs. BMC Bioinform. 2019, 20, 372. [Google Scholar] [CrossRef]
- Kowalski, M.P.; Krude, T. Functional roles of non-coding Y RNAs. Int. J. Biochem. Cell Biol. 2015, 66, 20–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Backes, C.; Meese, E.; Keller, A. Specific miRNA Disease Biomarkers in Blood, Serum and Plasma: Challenges and Prospects. Mol. Diagn. Ther. 2016, 20, 509–518. [Google Scholar] [CrossRef]
- Rupaimoole, R.; Slack, F.J. microRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.J.; Söderhäll, C.; Bustamante, M.; Baïz, N.; Gruzieva, O.; Gehring, U.; Mason, D.; Chatzi, L.; Basterrechea, M.; Llop, S.; et al. DNA methylation in childhood asthma: An epigenome-wide meta-analysis. Lancet Respir. Med. 2018, 6, 379–388. [Google Scholar] [CrossRef] [Green Version]
- Arathimos, R.; Suderman, M.; Sharp, G.C.; Burrows, K.; Granell, R.; Tilling, K.; Gaunt, T.R.; Henderson, J.; Ring, S.; Richmond, R.C.; et al. Epigenome-wide association study of asthma and wheeze in childhood and adolescence. Clin. Epigenetics 2017, 9, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, I.V.; Pedersen, B.S.; Liu, A.; O’Connor, G.T.; Teach, S.J.; Kattan, M.; Misiak, R.T.; Gruchalla, R.; Steinbach, S.F.; Szefler, S.J.; et al. DNA methylation and childhood asthma in the inner city. J. Allergy Clin. Immunol. 2015, 136, 69–80. [Google Scholar] [CrossRef] [Green Version]
- Gunawardhana, L.P.; Baines, K.J.; Mattes, J.; Murphy, V.E.; Simpson, J.L.; Gibson, P.G. Differential DNA methylation profiles of infants exposed to maternal asthma during pregnancy. Pediatr. Pulmonol. 2014, 49, 852–862. [Google Scholar] [CrossRef] [PubMed]
- Forno, E.; Wang, T.; Yan, Q.; Brehm, J.; Acosta-Perez, E.; Colon-Semidey, A.; Alvarez, M.; Boutaoui, N.; Cloutier, M.M.; Alcorn, J.F.; et al. A Multiomics Approach to Identify Genes Associated with Childhood Asthma Risk and Morbidity. Am. J. Respir. Cell Mol. Biol. 2017, 57, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Adalsteinsson, B.T.; Gudnason, H.; Aspelund, T.; Harris, T.B.; Launer, L.J.; Eiriksdottir, G.; Smith, A.V.; Gudnason, V. Heterogeneity in white blood cells has potential to confound DNA methylation measurements. PLoS ONE 2012, 7, e46705. [Google Scholar] [CrossRef] [Green Version]
- Liang, L.; Willis-Owen, S.A.G.; Laprise, C.; Wong, K.C.C.; Davies, G.A.; Hudson, T.J.; Binia, A.; Hopkin, J.M.; Yang, I.V.; Grundberg, E.; et al. An epigenome-wide association study of total serum immunoglobulin E concentration. Nature 2015, 520, 670–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reese, S.E.; Xu, C.J.; den Dekker, H.T.; Lee, M.K.; Sikdar, S.; Ruiz-Arenas, C.; Merid, S.K.; Rezwan, F.I.; Page, C.M.; Ullemar, V.; et al. Epigenome-wide meta-analysis of DNA methylation and childhood asthma. J. Allergy Clin. Immunol. 2019, 143, 2062–2074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.J.; Gruzieva, O.; Qi, C.; Esplugues, A.; Gehring, U.; Bergström, A.; Mason, D.; Chatzi, L.; Porta, D.; Lodrup Carlsen, K.C.; et al. Shared DNA methylation signatures in childhood allergy: The MeDALL study. J. Allergy Clin. Immunol. 2021, 147, 1031–1040. [Google Scholar] [CrossRef]
- Kitazawa, H.; Masuko, H.; Kanazawa, J.; Shigemasa, R.; Hyodo, K.; Yamada, H.; Yatagai, Y.; Kaneko, Y.; Iijima, H.; Naito, T.; et al. ORMDL3/GSDMB genotype as a risk factor for early-onset adult asthma is linked to total serum IgE levels but not to allergic sensitization. Allergol. Int. 2021, 70, 55–60. [Google Scholar] [CrossRef]
- Madore, A.-M.; Pain, L.; Boucher-Lafleur, A.-M.; Morin, A.; Meloche, J.; Simon, M.-M.; Ge, B.; Kwan, T.; Cheung, W.A.; Pastinen, T.; et al. Asthma-associated polymorphisms in 17q12-21 locus modulate methylation and gene expression of GSDMA in naïve CD4+ T cells. J. Genet. Genom. 2020, 47, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Grozdanovic, M.; Laffey, K.G.; Abdelkarim, H.; Hitchinson, B.; Harijith, A.; Moon, H.G.; Park, G.Y.; Rousslang, L.K.; Masterson, J.C.; Furuta, G.T.; et al. Novel peptide nanoparticle-biased antagonist of CCR3 blocks eosinophil recruitment and airway hyperresponsiveness. J. Allergy Clin. Immunol. 2019, 143, 669–680.e612. [Google Scholar] [CrossRef] [Green Version]
- Uhm, T.G.; Lee, S.K.; Kim, B.S.; Kang, J.H.; Park, C.S.; Rhim, T.Y.; Chang, H.S.; Kim, D.J.; Chung, I.Y. CpG methylation at GATA elements in the regulatory region of CCR3 positively correlates with CCR3 transcription. Exp. Mol. Med. 2012, 44, 268–280. [Google Scholar] [CrossRef] [Green Version]
- Rodrigo-Muñoz, J.M.; Cañas, J.A.; Sastre, B.; Rego, N.; Greif, G.; Rial, M.; Mínguez, P.; Mahíllo-Fernández, I.; Fernández-Nieto, M.; Mora, I.; et al. Asthma diagnosis using integrated analysis of eosinophil microRNAs. Allergy 2019, 74, 507–517. [Google Scholar] [CrossRef]
- Bélanger, É.; Madore, A.M.; Boucher-Lafleur, A.M.; Simon, M.M.; Kwan, T.; Pastinen, T.; Laprise, C. Eosinophil microRNAs Play a Regulatory Role in Allergic Diseases Included in the Atopic March. Int. J. Mol. Sci. 2020, 21, 9011. [Google Scholar] [CrossRef]
- Allantaz, F.; Cheng, D.T.; Bergauer, T.; Ravindran, P.; Rossier, M.F.; Ebeling, M.; Badi, L.; Reis, B.; Bitter, H.; D’Asaro, M.; et al. Expression profiling of human immune cell subsets identifies miRNA-mRNA regulatory relationships correlated with cell type specific expression. PLoS ONE 2012, 7, e29979. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Ju, J.; Liu, J.; Li, D. Aberrant expression of miR-663 and transforming growth factor-β1 in nasal polyposis in children. Exp. Ther. Med. 2018, 15, 4550–4556. [Google Scholar] [CrossRef] [Green Version]
- Wong, C.K.; Lau, K.M.; Chan, I.H.; Hu, S.; Lam, Y.Y.; Choi, A.O.; Lam, C.W. microRNA-21* regulates the prosurvival effect of GM-CSF on human eosinophils. Immunobiology 2013, 218, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Grausenburger, R.; Bilic, I.; Boucheron, N.; Zupkovitz, G.; El-Housseiny, L.; Tschismarov, R.; Zhang, Y.; Rembold, M.; Gaisberger, M.; Hartl, A.; et al. Conditional deletion of histone deacetylase 1 in T cells leads to enhanced airway inflammation and increased Th2 cytokine production. J. Immunol. 2010, 185, 3489–3497. [Google Scholar] [CrossRef] [PubMed]
- Koyanagi, K.O. Inferring changes in histone modification during cell differentiation by ancestral state estimation based on phylogenetic trees of cell types: Human hematopoiesis as a model case. Gene X 2019, 3, 100021. [Google Scholar] [CrossRef] [PubMed]
- Miao, R.; Dong, X.; Gong, J.; Wang, Y.; Guo, X.; Li, Y.; Li, J.; Yang, S.; Kuang, T.; Wan, J.; et al. Possible immune regulation mechanisms for the progression of chronic thromboembolic pulmonary hypertension. Thromb. Res. 2021, 198, 122–131. [Google Scholar] [CrossRef]
- Huang, Z.; Cao, Y.; Zhou, M.; Qi, X.; Fu, B.; Mou, Y.; Wu, G.; Xie, J.; Zhao, J.; Xiong, W. Hsa_circ_0005519 increases IL-13/IL-6 by regulating hsa-let-7a-5p in CD4(+) T cells to affect asthma. Clin. Exp. Allergy 2019, 49, 1116–1127. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.H.; Liao, B.; Liu, K.; Liu, Y.H. Effect of RNA interference therapy on the mice eosinophils CCR3 gene and granule protein in the murine model of allergic rhinitis. Asian Pac. J. Trop. Med. 2014, 7, 226–230. [Google Scholar] [CrossRef] [Green Version]
- Errahali, Y.J.; Taka, E.; Abonyo, B.O.; Heiman, A.S. CCL26-targeted siRNA treatment of alveolar type II cells decreases expression of CCR3-binding chemokines and reduces eosinophil migration: Implications in asthma therapy. J. Interferon Cytokine Res. 2009, 29, 227–239. [Google Scholar] [CrossRef]
- Zhang, S.; Yang, R.; Zheng, Y. The effect of siRNA-mediated lymphocyte-specific protein tyrosine kinase (Lck) inhibition on pulmonary inflammation in a mouse model of asthma. Int. J. Clin. Exp. Med. 2015, 8, 15146–15154. [Google Scholar]
- Zafra, M.P.; Cañas, J.A.; Mazzeo, C.; Gámez, C.; Sanz, V.; Fernández-Nieto, M.; Quirce, S.; Barranco, P.; Ruiz-Hornillos, J.; Sastre, J.; et al. SOCS3 silencing attenuates eosinophil functions in asthma patients. Int. J. Mol. Sci. 2015, 16, 5434–5451. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Chen, H.; Li, Y.M.; Wang, S.Y.; Diao, X.; Liu, K.G. Intranasal sirna targeting c-kit reduces airway inflammation in experimental allergic asthma. Int. J. Clin. Exp. Pathol. 2014, 7, 5505–5514. [Google Scholar] [PubMed]
- Suzuki, M.; Yokota, M.; Matsumoto, T.; Ozaki, S. Synergic Effects of CD40 and CD86 Silencing in Dendritic Cells on the Control of Allergic Diseases. Int. Arch. Allergy Immunol. 2018, 177, 87–96. [Google Scholar] [CrossRef]
- Liu, D.J.; Leal, S.M. A novel adaptive method for the analysis of next-generation sequencing data to detect complex trait associations with rare variants due to gene main effects and interactions. PLoS Genet. 2010, 6, e1001156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darcan-Nicolaisen, Y.; Meinicke, H.; Fels, G.; Hegend, O.; Haberland, A.; Kühl, A.; Loddenkemper, C.; Witzenrath, M.; Kube, S.; Henke, W.; et al. Small interfering RNA against transcription factor STAT6 inhibits allergic airway inflammation and hyperreactivity in mice. J. Immunol. 2009, 182, 7501–7508. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, R.; Wu, J.; Hu, H. Knockdown of neurokinin-1 receptor expression by small interfering RNA prevents the development of allergic rhinitis in rats. Inflamm. Res. 2013, 62, 903–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Abbreviation | Full Name | Size | Function | Reference |
|---|---|---|---|---|
| miRNA | microRNA | 21–23 nt | Represses target mRNA by accelerating deadenylation and blocking ribosome | [51] |
| siRNA | Small interfering RNA | 20–25 nt | Degrades target mRNA after its transcription | [52] |
| piRNA | Piwi-interacting RNA | 26–32 nt | Binds to piwi proteins to silence transposons | [53] |
| eRNA | Enhancer RNA | 50–2000 nt | Influences gene expression by modulating chromatin structure | [54] |
| lncRNA | Long noncoding RNA | >200 nt | Induces changes in chromatin structure | [53] |
| circRNA | Circular RNA | 100–10,000 nt | Regulates gene expression by binding to miRNAs, preventing them from binding to mRNA | [55] |
| Y RNA | Y RNA | 80–120 nt | Inhibits DNA replication | [56] |
| Total Samples n | Eosinophil Samples n | Mechanism | Disease | Country of Eosinophil Sample Collection | Author, PMID |
|---|---|---|---|---|---|
| 664 | 24 | Methylation | Asthma | Canada | Liang et al., 25707804 |
| 1850 | 24 | Methylation | Childhood asthma | Canada | Reese et al., 30579849 |
| 6539 | 24 | Methylation | Childhood asthma | Canada | Xu et al., 2018, 29496485 |
| 5826 | 24 | Methylation | Childhood allergy | Canada | Xu et al., 2020, 33338541 |
| 315 | 140 | Methylation | Allergic asthma | Canada | Madore et al., 32312674 |
| N/A | N/A | Methylation | N/A | Korea (commercial cell lines) | Uhm et al., 22217447 |
| Eosinophil Samples n | miRNAs Studied n | Mechanism | Disease | Country of Eosinophil Sample Collection | Author, PMID |
|---|---|---|---|---|---|
| 44 | 14 | miRNA | Asthma | Spain | Rodrigo-Muñoz et al., 30040124 |
| 145 | 18 | miRNA | Atopic march | Canada | Bélanger et al., 33260893 |
| N/A | 18 | miRNA | N/A | France | Allantaz et al., 22276136 |
| 81 | 1 | miRNA | Nasal polyposis | China | Yu et al., 29849780 |
| N/A | 1 | miRNA | N/A; eosinophil survival | China | Wong et al., 22698984 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bélanger, É.; Laprise, C. Could the Epigenetics of Eosinophils in Asthma and Allergy Solve Parts of the Puzzle? Int. J. Mol. Sci. 2021, 22, 8921. https://doi.org/10.3390/ijms22168921
Bélanger É, Laprise C. Could the Epigenetics of Eosinophils in Asthma and Allergy Solve Parts of the Puzzle? International Journal of Molecular Sciences. 2021; 22(16):8921. https://doi.org/10.3390/ijms22168921
Chicago/Turabian StyleBélanger, Émile, and Catherine Laprise. 2021. "Could the Epigenetics of Eosinophils in Asthma and Allergy Solve Parts of the Puzzle?" International Journal of Molecular Sciences 22, no. 16: 8921. https://doi.org/10.3390/ijms22168921
APA StyleBélanger, É., & Laprise, C. (2021). Could the Epigenetics of Eosinophils in Asthma and Allergy Solve Parts of the Puzzle? International Journal of Molecular Sciences, 22(16), 8921. https://doi.org/10.3390/ijms22168921