
Simple and Rapid Method for Wogonin Preparation and Its Biotransformation

 , , ,
, , ,
Abstract
:1. Introduction
2. Results and Discussion
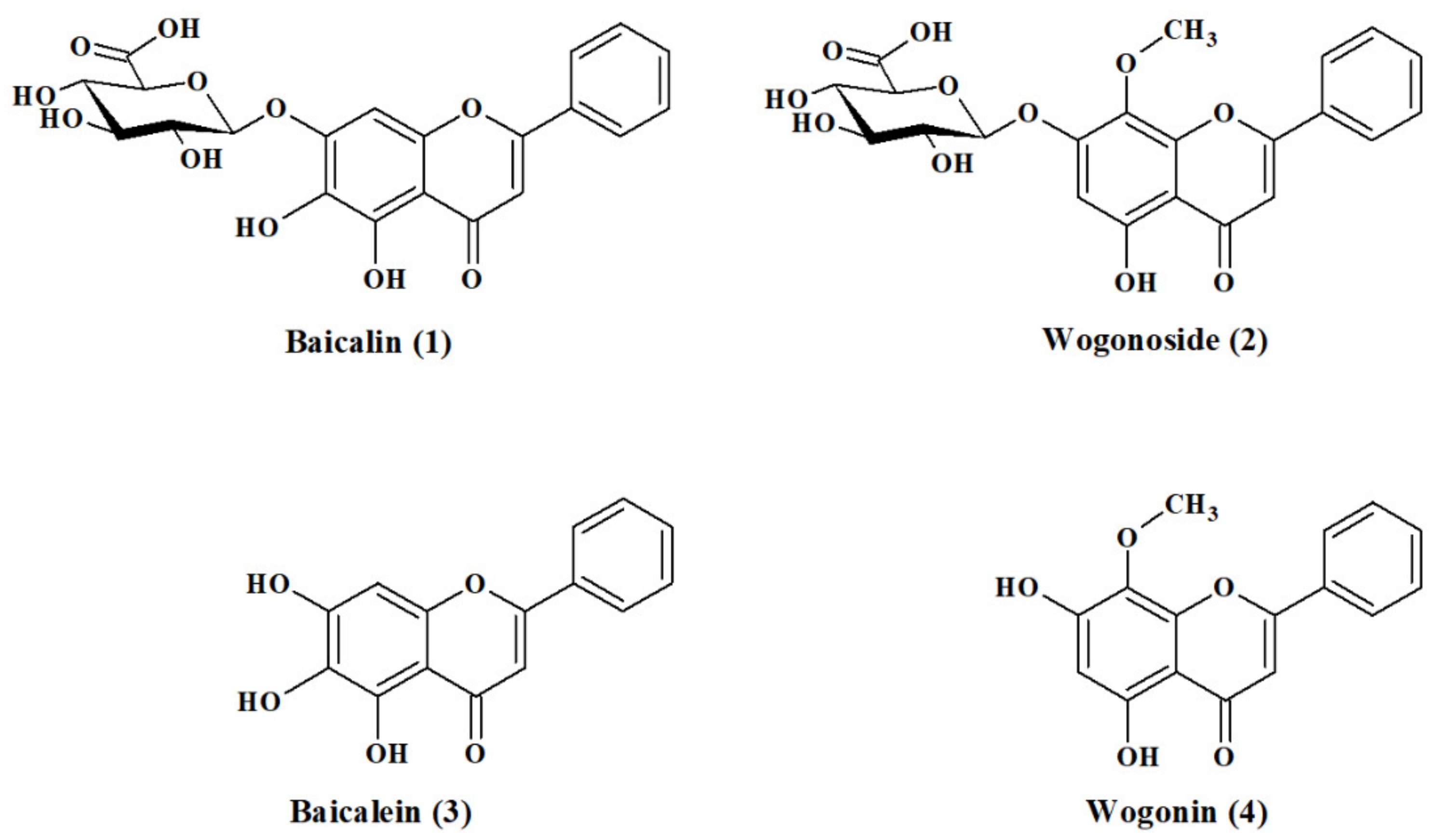
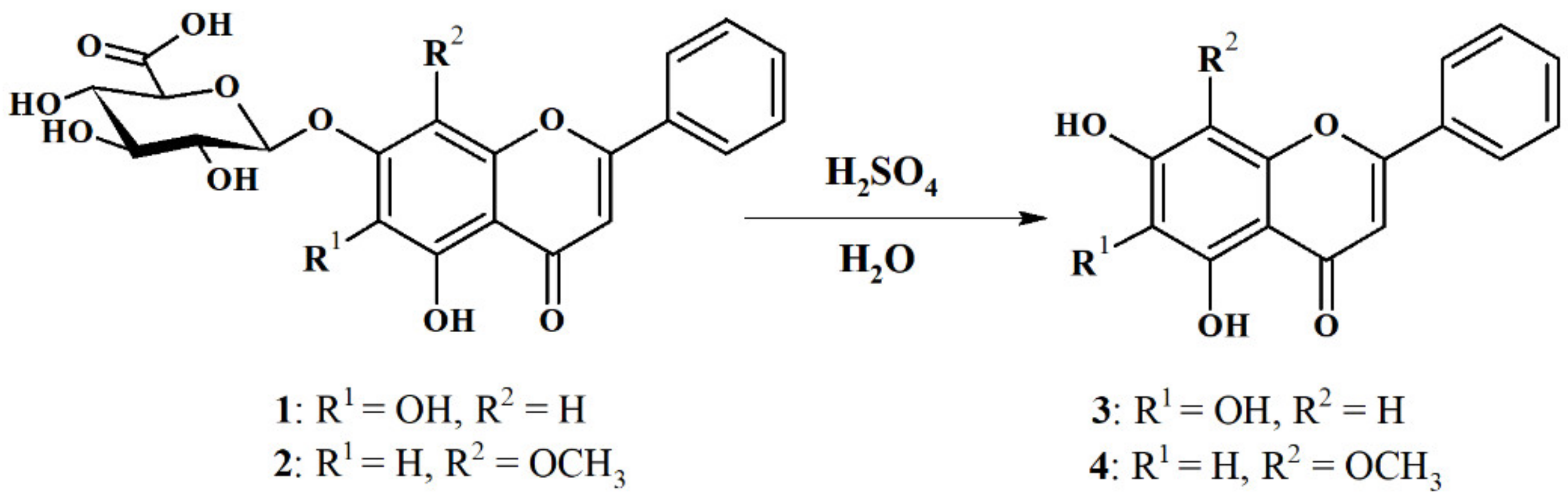
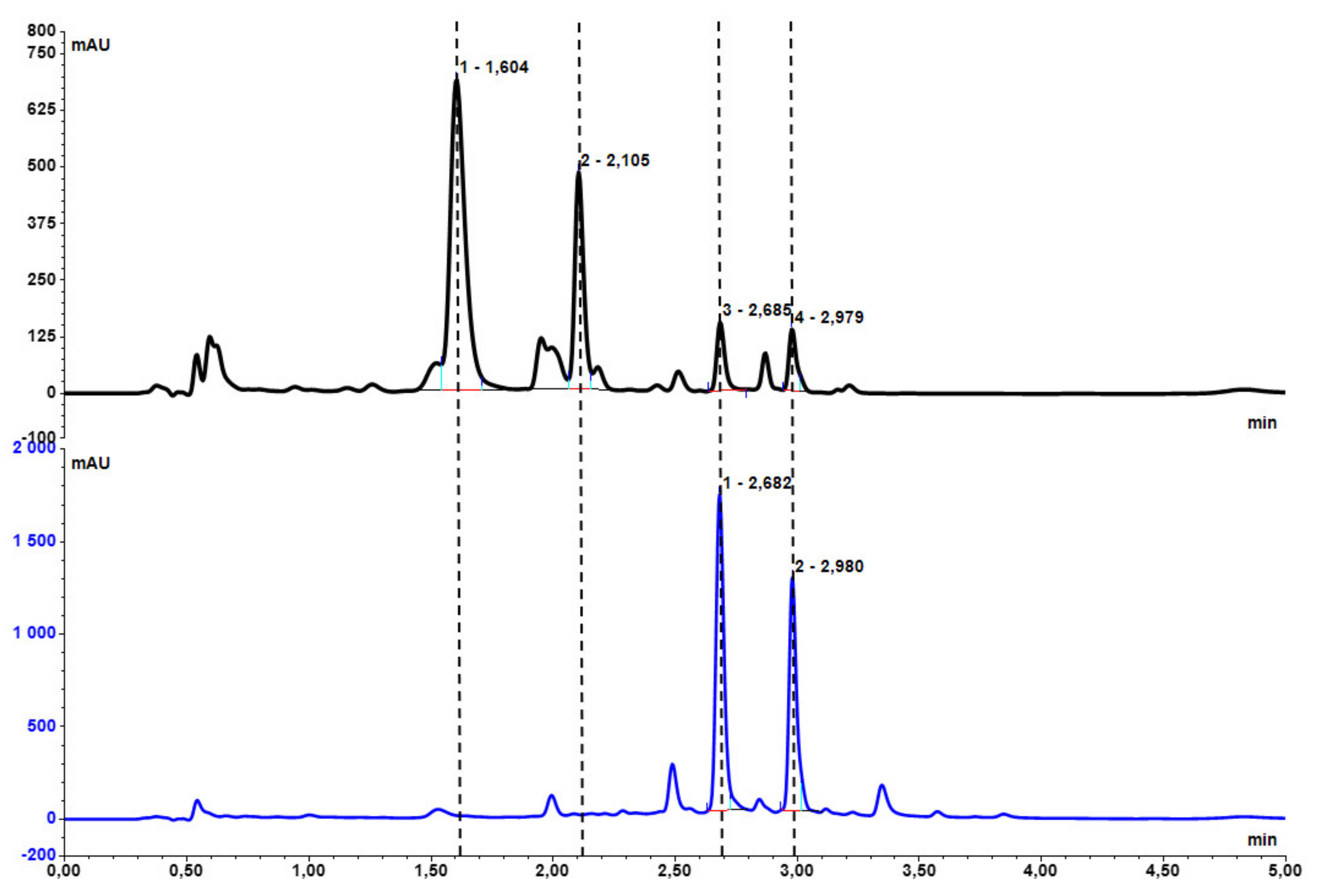
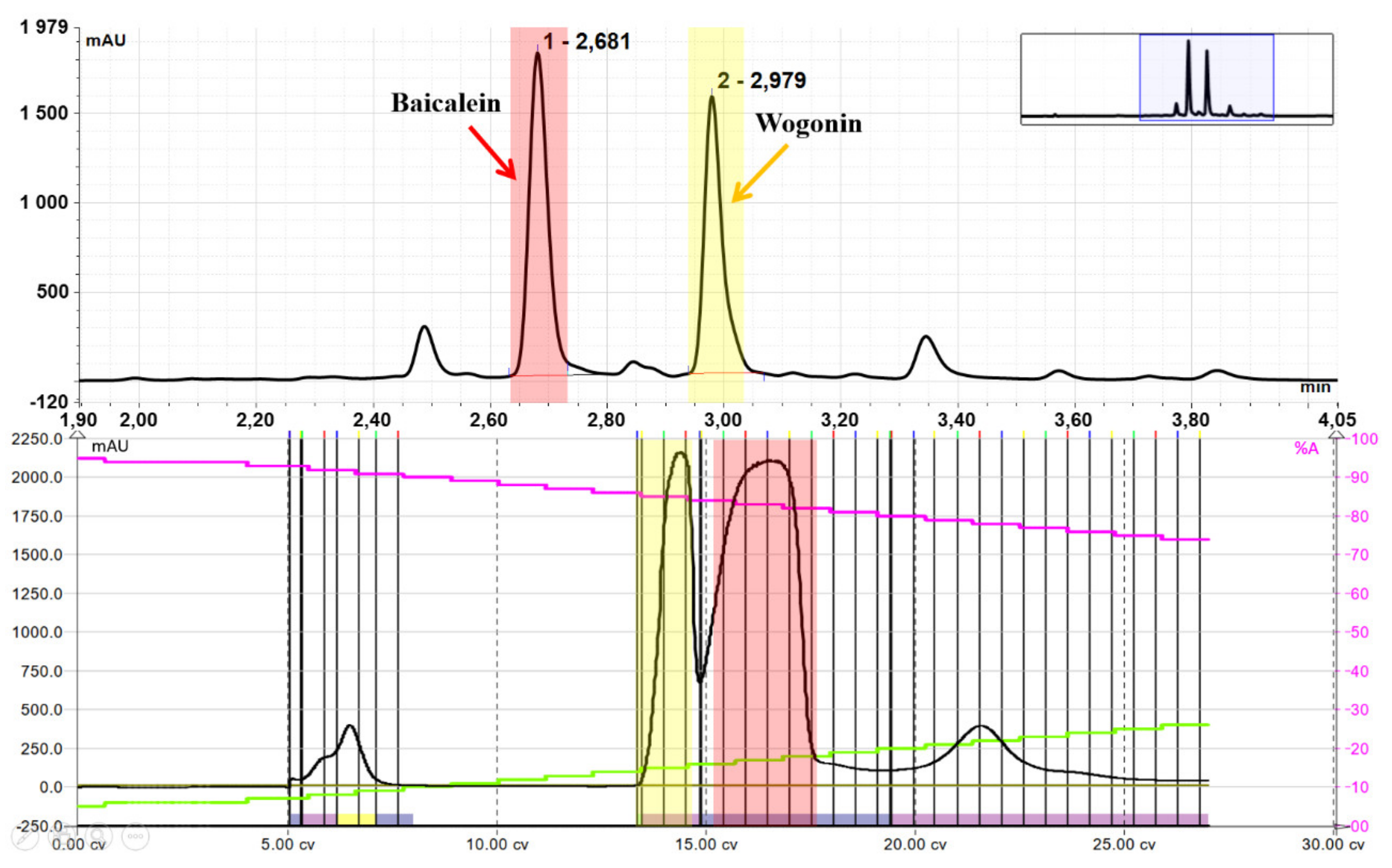
2.1. Hydrolysis of Dried, Powdered Baikal Skullcap Root
- Vessel: 100 mL round bottom flask; after achieving the reaction time, the mixture was poured to 100 mL of ice-cold water and stirred for 15 min.
- Vessel: 250 mL round bottom flask; after achieving the reaction time, the mixture was poured to 300 mL of ice-cold water and stirred for 15 min.
- Vessel: 1000 mL round bottom flask; after achieving the reaction time, the mixture was added to 900 mL of ice-cold water and stirred for 15 min.
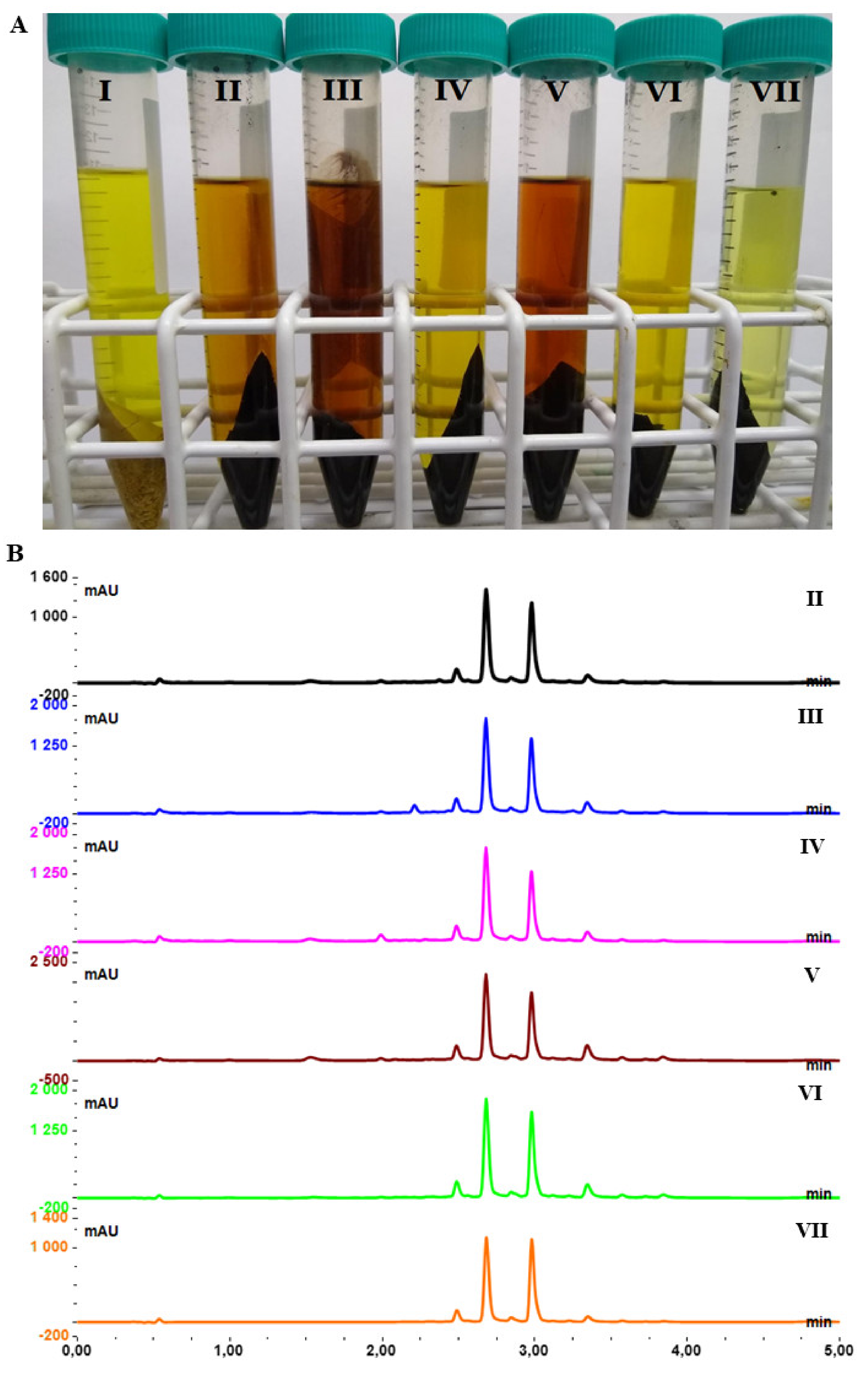
2.2. Selective Extraction of Dry Root Hydrolysate
2.3. Purification of Wogonin and Baicalein by Flash Chromatography
2.4. Biotransformation
2.4.1. Screening Tests
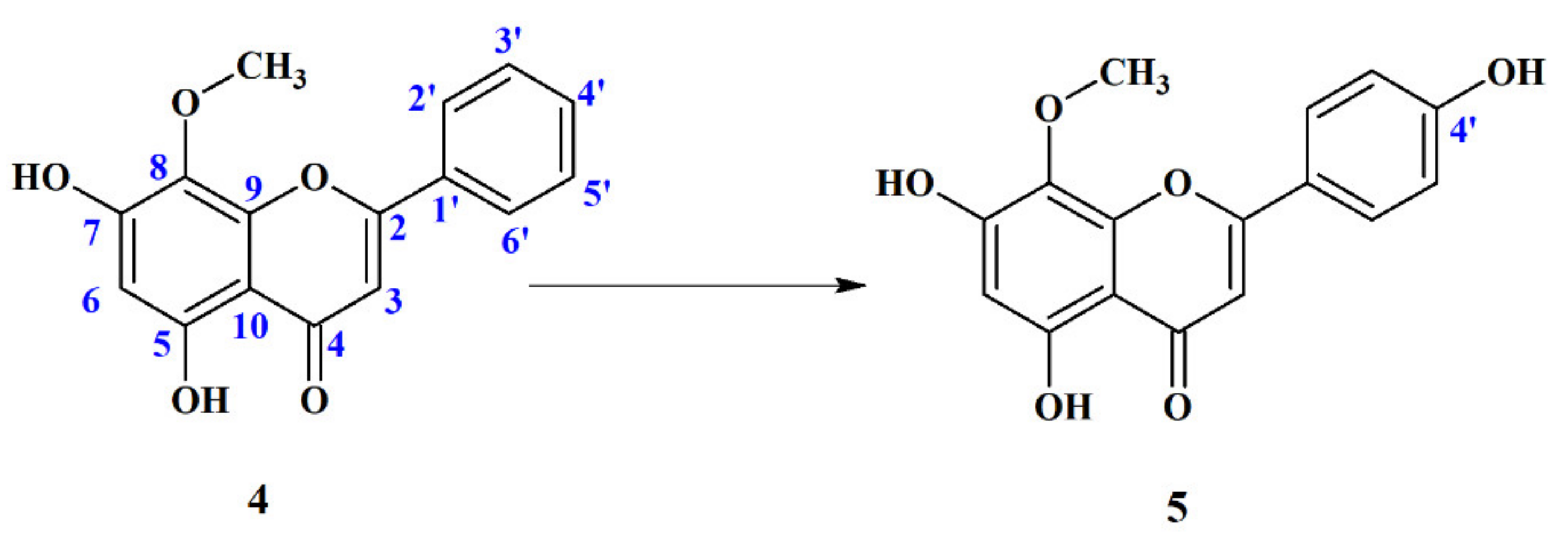
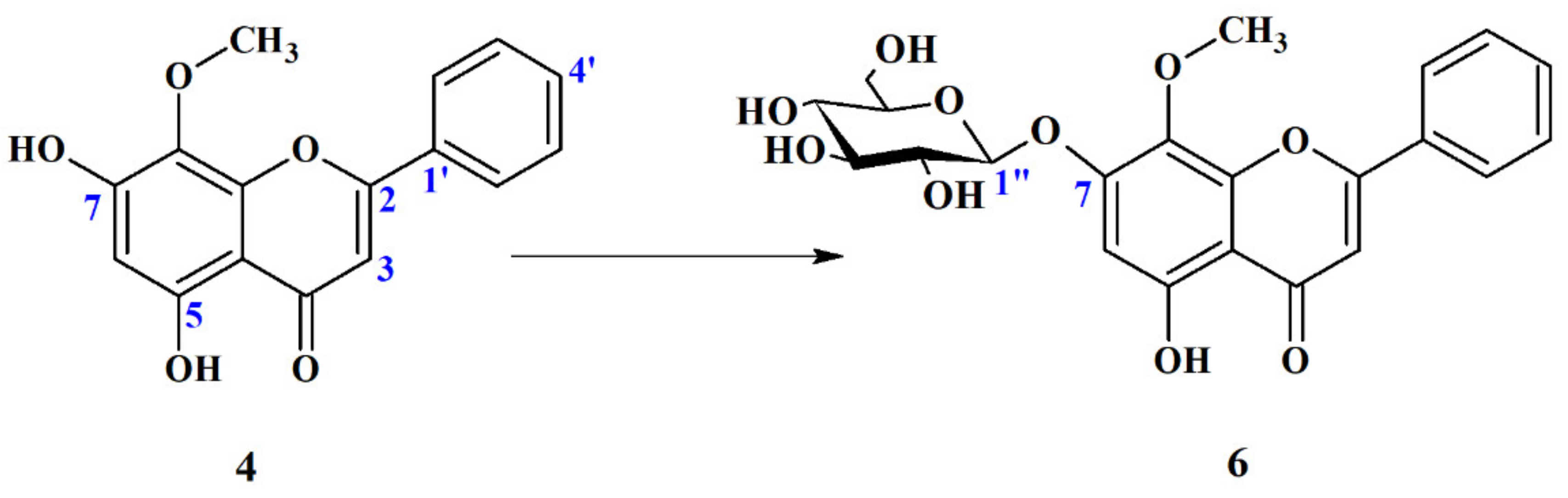
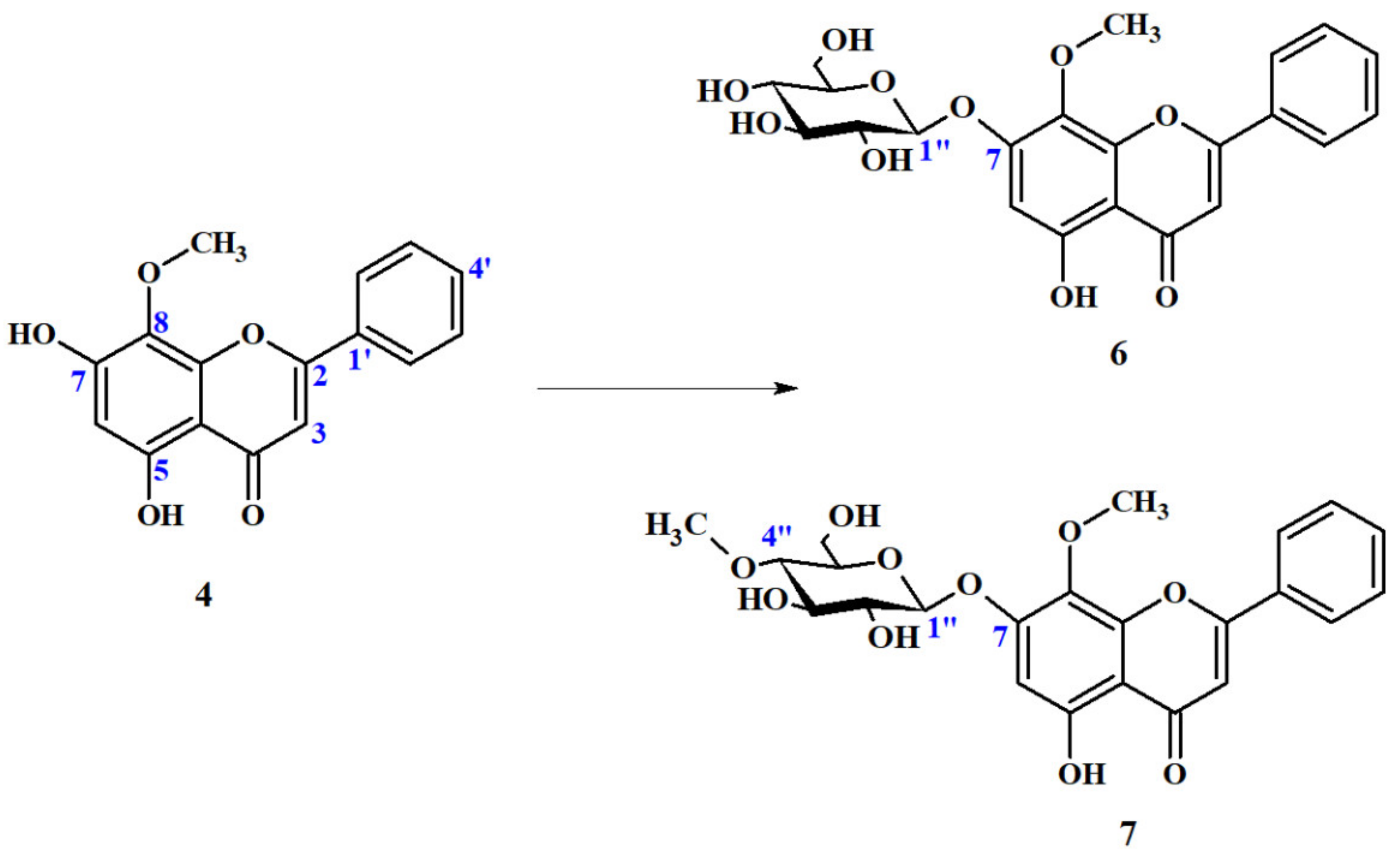
2.4.2. Biotransformation Products
4′-Hydroxywogonin (5,7,4′-Trihydroxy-8-Metoxyflavone) (5)
Wogonin 7-O-β-D-Glucopyranoside (6)
Wogonin 7-O-β-D-(4″-O-Methyl)-Glucopyranoside (7)
3. Material and Methods
3.1. General Experimental Methods
3.2. Materials
Biotransformation Products
3.3. Chemistry
Hydrolysis of Dry, Powdered Baikal Skullcap
3.4. Microorganisms
3.5. Conditions for Biotransformation
3.5.1. Screening Studies
3.5.2. Scale-Up Biotransformation
3.6. Products Isolation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhao, T.; Tang, H.; Xie, L.; Zheng, Y.; Ma, Z.; Sun, Q.; Li, X. Scutellaria baicalensis Georgi. (Lamiaceae): A review of its traditional uses, botany, phytochemistry, pharmacology and toxicology. J. Pharm. Pharmacol. 2019, 71, 1353–1369. [Google Scholar] [CrossRef] [Green Version]
- Błach, Z.; Lamer, E. Come back to root–therapeutic activities of Scutellaria baicalensis root in aspect of innate immunity regulation—Part I. Adv. Clin. Exp. Med. 2008, 17, 337–345. [Google Scholar]
- Makino, T.; Hishida, A.; Goda, Y.; Mizukami, H. Comparison of the major flavonoid content of S. baicalensis, S. lateriflora, and their commercial products. J. Nat. Med. 2008, 62, 294–299. [Google Scholar] [CrossRef]
- Wang, H.-Z.; Yu, C.-H.; Gao, J.; Zhao, G.-R. Effects of processing and extracting methods on active components in Radix Scutellariae by HPLC analysis. China J. Chin. Mater. Medica 2007, 32, 1637–1640. [Google Scholar]
- Shen, J.; Li, P.; He, C.-N.; Liu, H.-T.; Liu, Y.-Z.; Sun, X.-B.; Xu, R.; Xiao, P.-G. Simultaneous determination of 15 flavonoids from different parts of Scutellaria baicalensis and its chemometrics analysis. Chin. Herb. Med. 2018, 11, 20–27. [Google Scholar] [CrossRef]
- Gao, Z.; Huang, K.; Yang, X.; Xu, H. Free radical scavenging and antioxidant activities of flavonoids extracted from the radix of Scutellaria baicalensis Georgi. Biochim. Biophys. Acta BBA Gen. Subj. 1999, 1472, 643–650. [Google Scholar] [CrossRef]
- Chi, Y.S.; Lim, H.; Park, H.; Kim, H.P. Effects of wogonin, a plant flavone from Scutellaria radix, on skin inflammation: In vivo regulation of inflammation-associated gene expression. Biochem. Pharmacol. 2003, 66, 1271–1278. [Google Scholar] [CrossRef]
- Kimura, Y.; Yokoi, K.; Matsushita, N.; Okuda, H. Effects of Flavonoids Isolated from Scutellariae Radix on the Production of Tissue-type Plasminogen Activator and Plasminogen Activator Inhibitor-1 Induced by Thrombin and Thrombin Receptor Agonist Peptide in Cultured Human Umbilical Vein Endothelial Cells. J. Pharm. Pharmacol. 1997, 49, 816–822. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Tsang, S.-Y.; Yao, X.; Chen, Z.-Y. Biological properties of baicalein in cardiovascular system. Curr. Drug Targets Cardiovasc. Haematol. Disord. 2005, 5, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.A.; Attele, A.S.; Zhang, L.; Yuan, C.-S. Anti-HIV Activity of Medicinal Herbs: Usage and Potential Development. Am. J. Chin. Med. 2001, 29, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-K.; Xia, Y.; Yang, Z.-Y.; Natschke, S.L.M.; Lee, K.-H. Recent Advances in the Discovery and Development of Flavonoids and their Analogues as Antitumor and Anti-HIV Agents. Adv. Exp. Med. Biol. 1998, 439, 191–225. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.-C.; Du, J.; But, P.P.-H.; Deng, X.-L.; Zhang, Y.-W.; Ooi, V.E.-C.; Xu, H.-X.; Lee, S.H.-S.; Lee, S.F. Antiviral Chinese medicinal herbs against respiratory syncytial virus. J. Ethnopharmacol. 2001, 79, 205–211. [Google Scholar] [CrossRef]
- Huang, R.-L.; Chen, C.-C.; Huang, H.-L.; Chang, C.-G.; Chen, C.-F.; Chang, C.; Hsieh, M.-T. Anti-hepatitis B virus effects of wogonin isolated from Scutellaria baicalensis. Planta Med. 2000, 66, 694–698. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Zhao, L.; You, Q.; Yang, Y.; Gu, H.; Song, G.; Lu, N.; Xin, J. Anti-hepatitis B virus activity of wogonin in vitro and in vivo. Antivir. Res. 2007, 74, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-Z.; Mehendale, S.R.; Yuan, C.-S. Commonly Used Antioxidant Botanicals: Active Constituents and Their Potential Role in Cardiovascular Illness. Am. J. Chin. Med. 2007, 35, 543–558. [Google Scholar] [CrossRef]
- Li-Weber, M. New therapeutic aspects of flavones: The anticancer properties of Scutellaria and its main active constituents Wogonin, Baicalein and Baicalin. Cancer Treat. Rev. 2009, 35, 57–68. [Google Scholar] [CrossRef]
- Li, B.Q.; Fu, T.; Yan, Y.D.; Baylor, N.W.; Ruscetti, F.W.; Kung, H.F. Inhibition of HIV infection by baicalin—A flavonoid compound purified from Chinese herbal medicine. Cell. Mol. Boil. Res. 1993, 39, 119–124. [Google Scholar]
- Nagai, T.; Suzuki, Y.; Tomimori, T.; Yamada, H. Antiviral Activity of Plant Flavonoid, 5,7,4′-Trihydroxy-8-methoxyflavone, from the Roots of Scutellaria baicalensis against Influenza A (H3N2) and B Viruses. Biol. Pharm. Bull. 1995, 18, 295–299. [Google Scholar] [CrossRef] [Green Version]
- Coon, J.T.; Ernst, E. Complementary and alternative therapies in the treatment of chronic hepatitis C: A systematic review. J. Hepatol. 2004, 40, 491–500. [Google Scholar] [CrossRef]
- Li, Y.; Ooi, L.S.; Wang, H.; But, P.P.; Ooi, V.E. Antiviral activities of medicinal herbs traditionally used in southern mainland China. Phytother. Res. 2004, 18, 718–722. [Google Scholar] [CrossRef]
- Huang, Y.-F.; Bai, C.; He, F.; Xie, Y.; Zhou, H. Review on the potential action mechanisms of Chinese medicines in treating Coronavirus Disease 2019 (COVID-19). Pharmacol. Res. 2020, 158, 104939. [Google Scholar] [CrossRef]
- Tong, T.; Wu, Y.-Q.; Ni, W.-J.; Shen, A.-Z.; Liu, S. The potential insights of Traditional Chinese Medicine on treatment of COVID-19. Chin. Med. 2020, 15, 1–6. [Google Scholar] [CrossRef]
- Kong, Y.; Wu, H.-W.; Chen, Y.; Lai, S.; Yang, Z.-M.; Chen, J.-S. Mechanism of Tanreqing Injection on treatment of coronavirus disease 2019 based on network pharmacology and molecular docking. Chin. Trad. Herbal Drugs 2020, 7, 1785–1794. [Google Scholar]
- Shen, F.; Fu, Z.; Wu, Y.; Li, L.; Zhao, Y.; Xia, Y.; Kuang, G. Based on Network Pharmacology and High-Throughput Molecular Docking to Study the Potential Molecular Mechanism of Active Compounds That Bind SARS-Cov-2 Specific Target Protein in Jinhua Qingan Granules to Interfere with Covid-19. Modernization of Traditional Chinese Medicine and Materia Materia-World Science and Technology. 2020. Available online: https://kns.cnki.net/kcms/detail/11.5699.R.20200421.0949.004.html (accessed on 19 July 2021).
- Mao, Y.; Su, Y.; Xue, P.; Li, L.; Zhu, S. Discussion on the mechanism of Jinhua Qinggan Granule in the treatment of novel coronavirus pneumonia. J. Chin. Med. Mater. 2020, 43, 2843–2849. [Google Scholar]
- Huang, W.H.; Chien, P.Y.; Yang, C.H.; Lee, A.R. Novel synthesis of flavonoids of Scutellaria baicalensis Georgi. Chem. Pharm. Bull. 2003, 51, 339–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.-Y.; Cao, X.; Wang, X.; Guo, Q.-L.; You, Q.-D. Convenient Synthesis of Wogonin, A Flavonoid Natural Product with Extensive Pharmacological Activity. Org. Prep. Proced. Int. 2009, 41, 327–330. [Google Scholar] [CrossRef]
- Zhang, W.; Yi, D.; Gao, K.; Liu, M.; Yang, J.; Liao, X.; Yang, B. hydrolysis of Scutellarin and Related Glycosides to Scutellarein and the Corresponding Aglycones. J. Chem. Res. 2014, 38, 396–398. [Google Scholar] [CrossRef]
- Huszcza, E.; Bartmańska, A.; Tronina, T. Glycosylation of xanthohumol by fungi. Zeitschrift für Naturforschung C 2008, 63, 557–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartmańska, A.; Huszcza, E.; Tronina, T. Transformation of isoxanthohumol by fungi. J. Mol. Catal. B Enzym. 2009, 61, 221–224. [Google Scholar] [CrossRef]
- Bartmańska, A.; Tronina, T.; Huszcza, E. Transformation of 8-prenylnaringenin by Absidia coerulea and Beauveria bassiana. Bioorg. Med. Chem. Lett. 2012, 22, 6451–6453. [Google Scholar] [CrossRef] [PubMed]
- Tronina, T.; Bartmańska, A.; Milczarek, M.; Wietrzyk, J.; Popłoński, J.; Rój, E.; Huszcza, E. Antioxidant and antiproliferative activity of glycosides obtained by biotransformation of xanthohumol. Bioorg. Med. Chem. Lett. 2013, 23, 1957–1960. [Google Scholar] [CrossRef] [PubMed]
- Strugała, P.; Tronina, T.; Huszcza, E.; Gabrielska, J. Bioactivity In Vitro of Quercetin Glycoside Obtained in Beauveria bassiana Culture and Its Interaction with Liposome Membranes. Molecules 2017, 22, 1520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tronina, T.; Strugała, P.; Popłoński, J.; Włoch, A.; Sordon, S.; Bartmańska, A.; Huszcza, E. The Influence of Glycosylation of Natural and Synthetic Prenylated Flavonoids on Binding to Human Serum Albumin and Inhibition of Cyclooxygenases COX-1 and COX-2. Molecules 2017, 22, 1230. [Google Scholar] [CrossRef] [Green Version]
- Bartmańska, A.; Tronina, T.; Popłoński, J. Biotransformation of a major beer prenylflavonoid–isoxanthohumol. Z. Naturforsch. C. 2018, 74, 1–7. [Google Scholar] [CrossRef]
- Sordon, S.; Popłoński, J.; Tronina, T.; Huszcza, E. Microbial Glycosylation of Daidzein, Genistein and Biochanin A: Two New Glucosides of Biochanin A. Molecules 2017, 22, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sordon, S.; Popłoński, J.; Tronina, T.; Huszcza, E. Regioselective O-glycosylation of flavonoids by fungi Beauveria bassiana, Absidia coerulea and Absidia glauca. Bioorg. Chem. 2019, 93, 102750. [Google Scholar] [CrossRef]
- Łużny, M.; Tronina, T.; Kozłowska, E.; Dymarska, M.; Popłoński, J.; Łyczko, J.; Kostrzewa-Susłow, E.; Janeczko, T. Biotransformation of Methoxyflavones by Selected Entomopathogenic Filamentous Fungi. Int. J. Mol. Sci. 2020, 21, 6121. [Google Scholar] [CrossRef] [PubMed]
- Bartmańska, A.; Tronina, T.; Huszcza, E. Microbial sulfation of 8-prenylnaringenin. Z. Naturforsch. C. J. Biosci. 2013, 68, 231–235. [Google Scholar] [CrossRef]
- Łużny, M.; Tronina, T.; Kozłowska, E.; Kostrzewa-Susłow, E.; Janeczko, T. Biotransformation of 5,7-Methoxyflavones by Selected Entomopathogenic Filamentous Fungi. J. Agric. Food Chem. 2021, 69, 3879–3886. [Google Scholar] [CrossRef]
- Hollman, P.C.; Bijsman, M.N.; Van Gameren, Y.; Cnossen, E.P.; De Vries, J.H.; Katan, M.B. The sugar moiety is a major determinant of the absorption of dietary flavonoid glycosides in man. Free. Radic. Res. 1999, 31, 569–573. [Google Scholar] [CrossRef]
- Hanioka, N.; Isobe, T.; Tanaka-Kagawa, T.; Ohkawara, S. Wogonin glucuronidation in liver and intestinal microsomes of humans, monkeys, dogs, rats, and mice. Xenobiotica 2020, 50, 906–912. [Google Scholar] [CrossRef] [PubMed]
- Choi, R.-J.; Ha, I.-J.; Choi, J.-S.; Park, Y.-M.; Kim, Y.-S. Biotransformation of flavonoid-7-O-glucuronides by beta-glucuronidases. Nat. Prod. Sci. 2010, 16, 1–5. [Google Scholar] [CrossRef]
- Ku, S.; Zheng, H.; Park, M.S.; Ji, G.E. Optimization of β-glucuronidase activity from Lactobacillus delbrueckii Rh2 and and its use for biotransformation of baicalin and wogonoside. J. Appl. Biol. Chem. 2011, 54, 275–280. [Google Scholar] [CrossRef]
- Sakurama, H.; Kishino, S.; Uchibori, Y.; Yonejima, Y.; Ashida, H.; Kita, K.; Takahashi, S.; Ogawa, J. β-Glucuronidase from Lactobacillus brevis useful for baicalin hydrolysis belongs to glycoside hydrolase family 30. Appl. Microbiol. Biotechnol. 2013, 98, 4021–4032. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-S.; Kim, J.-Y.; Park, M.-S.; Zheng, H.; Ji, G.-E. Cloning and Expression of beta-Glucuronidase from Lactobacillus brevis in E. coli and Application in Bioconversion of Baicalin and Wogonoside. J. Microbiol. Biotechnol. 2009, 19, 1650–1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tronina, T.; Bartmańska, A.; Filip-Psurska, B.; Wietrzyk, J.; Popłoński, J.; Huszcza, E. Fungal metabolites of xanthohumol with potent antiproliferative activity on human cancer cell lines in vitro. Bioorg. Med. Chem. 2013, 21, 2001–2006. [Google Scholar] [CrossRef] [PubMed]
- Tronina, T.; Bartmańska, A.; Popłoński, J.; Huszcza, E. Transformation of xanthohumol by Aspergillus ochraceus. J. Basic Microbiol. 2013, 54, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Popłoński, J.; Sordon, S.; Tronina, T.; Bartmańska, A.; Huszcza, E. Fungal metabolism of naphthoflavones. J. Mol. Catal. B Enzym. 2015, 117, 1–6. [Google Scholar] [CrossRef]
- Bartmanska, A.; Tronina, T.; Popłoński, J.; Huszcza, E. Biotransformations of Prenylated Hop Flavonoids for Drug Discovery and Production. Curr. Drug Metab. 2013, 14, 1083–1097. [Google Scholar] [CrossRef] [PubMed]
- Sordon, S.; Madej, A.; Popłoński, J.; Bartmańska, A.; Tronina, T.; Brzezowska, E.; Juszczyk, P.; Huszcza, E. Regioselective ortho-Hydroxylations of Flavonoids by Yeast. J. Agric. Food Chem. 2016, 64, 5525–5530. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-L.; Ou, J.-C.; Chen, C.-F.; Kuo, Y.H. Flavonoids from the Roots of Scutellaria Luzonica Rolfe. J. Chin. Chem. Soc. 1991, 38, 619–623. [Google Scholar] [CrossRef]
- Yao, H.; Li, S.; Hu, J.; Chen, Y.; Huang, L.; Lin, J.; Li, G.; Lin, X. Chromatographic Fingerprint and Quantitative Analysis of Seven Bioactive Compounds of Scutellaria barbata. Planta Medica 2010, 77, 388–393. [Google Scholar] [CrossRef]
- Castillo, L.; Díaz, M.; González-Coloma, A.; Rossini, C. Differential activity against aphid settling of flavones obtained from Clytostoma callistegioides (Bignoniaceae). Ind. Crop. Prod. 2013, 44, 618–621. [Google Scholar] [CrossRef]
- Lin, F.-J.; Yen, F.-L.; Chen, P.-C.; Wang, M.-C.; Lin, C.-N.; Lee, C.-W.; Ko, H.-H. HPLC-Fingerprints and Antioxidant Constituents of Phyla nodiflora. Sci. World J. 2014, 2014, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sujatha, P.; Sreekanth, G.; Khasim, S.; Rao, B.V.A.; Kumar, B.R.; Rao, A.V.N.A. Flavonoids of Dikamali: A phytochemical reinvestigation. Nat. Prod. Res. 2013, 27, 1930–1932. [Google Scholar] [CrossRef]
- Sun, D.; Zhang, F.; Qian, J.; Shen, W.; Fan, H.; Tan, J.; Li, L.; Xu, C.; Yang, Y.; Cheng, H. 4′-hydroxywogonin inhibits colorectal cancer angiogenesis by disrupting PI3K/AKT signaling. Chem. Interactions 2018, 296, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Gu-Fang, Z.; Wu, L.-H.; Zhang, G.-F.; Xu, F.; Zhang, S.; Zhang, X.; Sun, L.; Yu, Y.; Zhang, Y.; Ye, R.D. 4′-Hydroxywogonin suppresses lipopolysaccharide-induced inflammatory responses in RAW 264.7 macrophages and acute lung injury mice. PLoS ONE 2017, 12, e0181191. [Google Scholar] [CrossRef] [Green Version]
- Li, P.F.; An, X.Z.; Ma, Y.M.; Kong, Q.L.; Bao, B.X.; Yu, J. [Effect of 4′-Hydroxywogonin on Proliferation and Apoptosis of Human Acute Lymphoblastic Leukemia Cells]. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2018, 26, 65–69. [Google Scholar] [CrossRef]
- Kostrzewa-Susłow, E.; Dmochowska-Gładysz, J.; Janeczko, T.; Sroda, K.; Michalak, K.; Palko, A. Microbial transformations of 6- and 7-methoxyflavones in Aspergillus niger and Penicillium chermesinum cultures. Z. Naturforsch. C. J. Biosci. 2012, 67, 411–417. [Google Scholar] [CrossRef]
- Kostrzewa-Susłow, E.; Janeczko, T. Microbial Transformations of 7-Hydroxyflavanone. Sci. World J. 2012, 2012, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Holland, H.L.; Morris, T.A.; Nava, P.J.; Zabic, M. A new paradigm for biohydroxylation by Beauveria bassiana ATCC 7159. Tetrahedron 1999, 55, 7441–7460. [Google Scholar] [CrossRef]
- Dymarska, M.; Janeczko, T.; Kostrzewa-Susłow, E. Glycosylation of Methoxylated Flavonoids in the Cultures of Isaria fumosorosea KCH J2. Molecules 2018, 23, 2578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dymarska, M.; Grzeszczuk, J.; Urbaniak, M.; Janeczko, T.; Pląskowska, E.; Stępień, Ł.; Kostrzewa-Susłow, E. Glycosylation of 6-methylflavone by the strain Isaria fumosorosea KCH J2. PLoS ONE 2017, 12, e0184885. [Google Scholar] [CrossRef]
- Dymarska, M.; Janeczko, T.; Kostrzewa-Susłow, E. Glycosylation of 3-Hydroxyflavone, 3-Methoxyflavone, Quercetin and Baicalein in Fungal Cultures of the Genus Isaria. Molecules 2018, 23, 2477. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mass of Dry Root [g] | Sulfuric Acid [mL] | Water [mL] | Reaction Time [min] | Wogonnoside Conversion * [%] | Baicalin Conversion* [%] | |
|---|---|---|---|---|---|---|
| 1 | 1.00 | 10 | 10 | 15 | 100 | 100 |
| 2 | 5.00 | 20 | 20 | 15 | 100 | 99 |
| 3 | 15.00 | 60 | 60 | 25 | 99 | 91 |
| Solvent | Extract Mass * [mg] | Wogonin [mg/g] ** | Baicalein [mg/g] ** |
|---|---|---|---|
| Ethanol | 82.3 ± 1.2 | 195.9 ± 2.7 | 310.4 ± 4.5 |
| Methanol | 88.2 ± 1.2 | 172.4 ± 2.3 | 295.6 ± 4.0 |
| 2-Propanol | 67.7 ± 1.0 | 206.4 ± 2.9 | 306.9 ± 4.3 |
| Acetone | 87.8 ± 1.4 | 227.0 ± 3.7 | 362.2 ± 5.9 |
| Ethyl acetate | 56.4 ± 0.8 | 328.8 ± 4.5 | 473.1 ± 6.5 |
| Diethyl Ether | 31.6 ± 0.4 | 368.2 ± 4.8 | 482.2 ± 5.1 |
| Microorganism | Ability * | Microorganism | Ability * |
|---|---|---|---|
| Absidia coerulea AM93 | + + + | Penicillium chermesinum AM113 | – |
| Absidia cylindrospora AM336 | + + + | Penicillium chrysogenum AM112 | – |
| Absidia glauca AM177 | + + + | Penicillium citrinum AM354 | – |
| Absidia glauca AM254 | + + + | Penicillium diversum AM388 | + + + |
| Aspergillus ochraceus AM370 | + + | Penicillium frequentans AM351 | + + + |
| Aspergillus ochraceus AM456 | – | Penicillium frequentas AM359 | + |
| Beauveria bassiana AM278 | + + + | Penicillium lilacinum AM111 | – |
| Beauveria bassiana AM737 | D | Penicillium purpurogenum AM80 | – |
| Cunninghamella japonica AM472 | + + + | Penicillium spinulosum AM114 | + |
| Fusarium avanaceum AM11 | – | Penicillium thomi AM91 | + + + |
| Fusarium culmorum AM196 | – | Penicillium urticae AM84 | – |
| Fusarium culmorum AM282 | – | Penicillium vermiculatum AM30 | – |
| Fusarium tricinctum AM16 | + | Penicillium vermiculatum AM81 | + + |
| Mortierella isabellina AM212 | + + + | Penicillium vinaceum AM110 | – |
| Mortierella vinacea AM149 | + + | Pezicula cinnamomea AM53 | – |
| Mucor hiemalis AM729 | + + | Rhizopus nigricans AM701 | + |
| Penicillium albidum AM79 | + + + | Rhodotorula marina AM77 | – |
| Penicillium camembertii AM83 | D | Trametes versicolor AM536 | + + + |
| Microorganism | Conversion [%] * |
|---|---|
| Absidia cylindrospora AM336 | 38.3 |
| Mortierella vinacae AM149 | 33.8 |
| Penicillium diversum AM388 | 33.2 |
| Mortierella isabellina AM212 | 22.4 |
| Penicillium thomi AM91 | 22.2 |
| Penicillium frequentans AM351 | 16.2 |
| Penicillium frequentas AM359 | 8.7 |
| Peniciilium spinulosum AM114 | 4.5 |
| Microorganism | Conversion [%] * |
|---|---|
| Cunninghamella japonica AM472 | 53.8 |
| Absidia coerulea AM93 | 26.0 |
| Beauveria bassiana AM278 | 25.3 |
| Absidia glauca AM177 | 19.8 |
| Mucor hiemalis AM729 | 19.5 |
| Absidia glauca AM254 | 6.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tronina, T.; Mrozowska, M.; Bartmańska, A.; Popłoński, J.; Sordon, S.; Huszcza, E. Simple and Rapid Method for Wogonin Preparation and Its Biotransformation. Int. J. Mol. Sci. 2021, 22, 8973. https://doi.org/10.3390/ijms22168973
Tronina T, Mrozowska M, Bartmańska A, Popłoński J, Sordon S, Huszcza E. Simple and Rapid Method for Wogonin Preparation and Its Biotransformation. International Journal of Molecular Sciences. 2021; 22(16):8973. https://doi.org/10.3390/ijms22168973
Chicago/Turabian StyleTronina, Tomasz, Monika Mrozowska, Agnieszka Bartmańska, Jarosław Popłoński, Sandra Sordon, and Ewa Huszcza. 2021. "Simple and Rapid Method for Wogonin Preparation and Its Biotransformation" International Journal of Molecular Sciences 22, no. 16: 8973. https://doi.org/10.3390/ijms22168973
APA StyleTronina, T., Mrozowska, M., Bartmańska, A., Popłoński, J., Sordon, S., & Huszcza, E. (2021). Simple and Rapid Method for Wogonin Preparation and Its Biotransformation. International Journal of Molecular Sciences, 22(16), 8973. https://doi.org/10.3390/ijms22168973