
Physiological and Molecular Analysis Reveals the Differences of Photosynthesis between Colored and Green Leaf Poplars

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Changes in Chl, Carotenoid and Anthocyanin Contents between ‘L2025’ and ‘ZHP’ Leaves
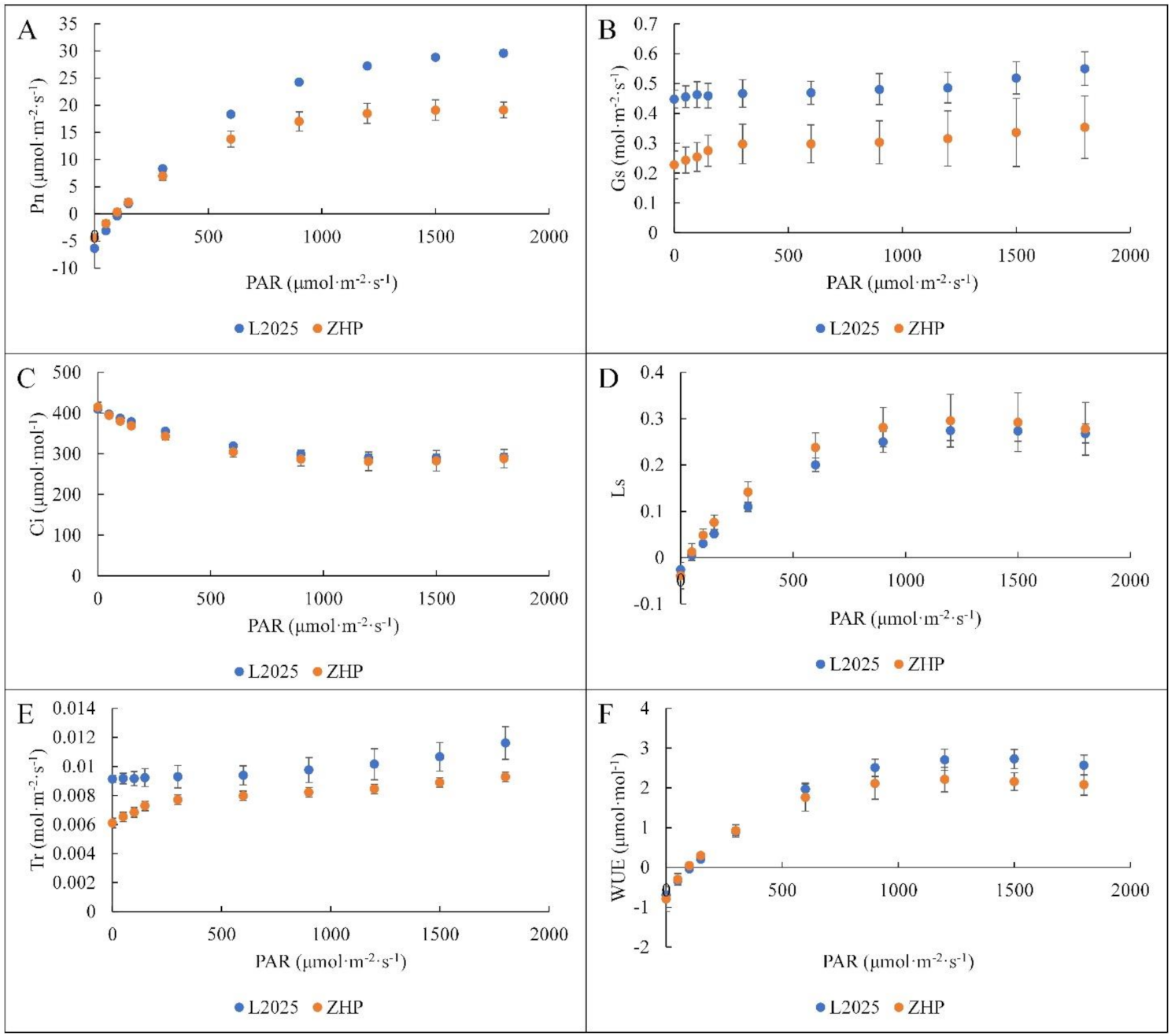
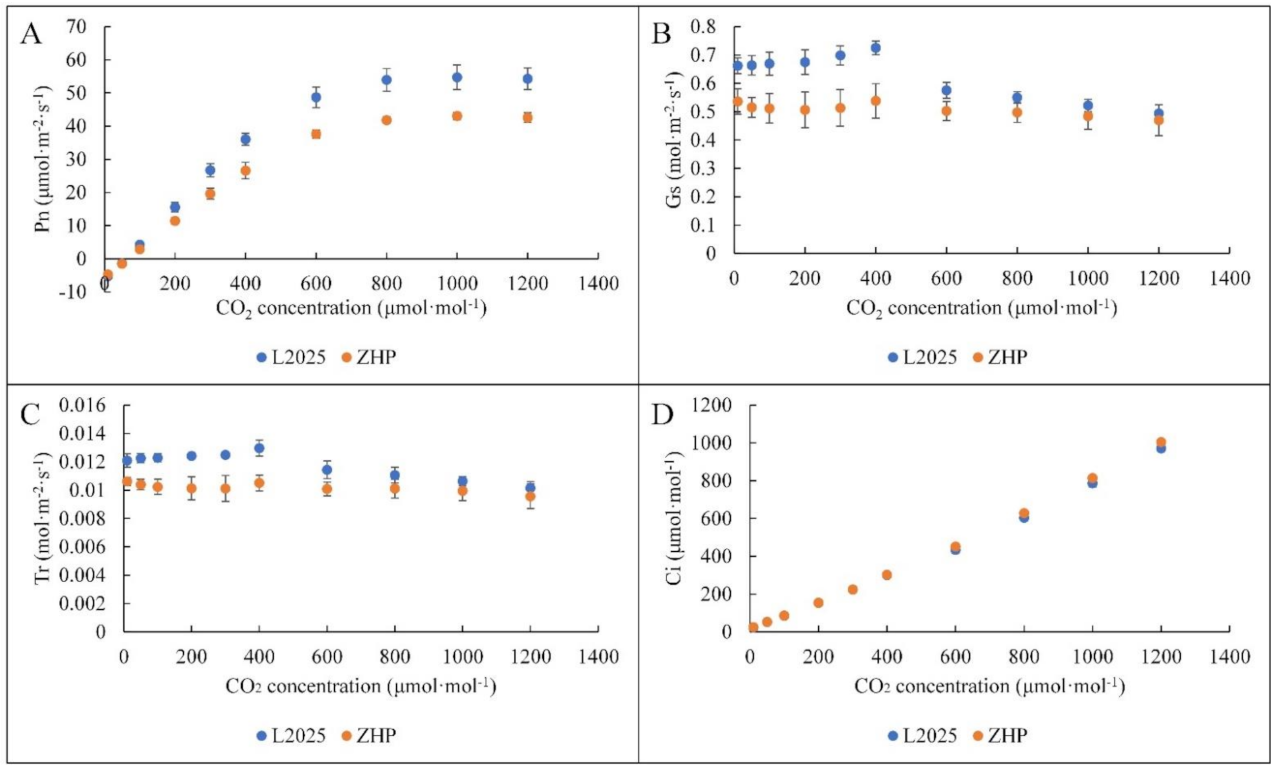
2.2. Changes in Light Response Curves and CO2 Response Curves between ‘L2025’ and ‘ZHP’ Leaves
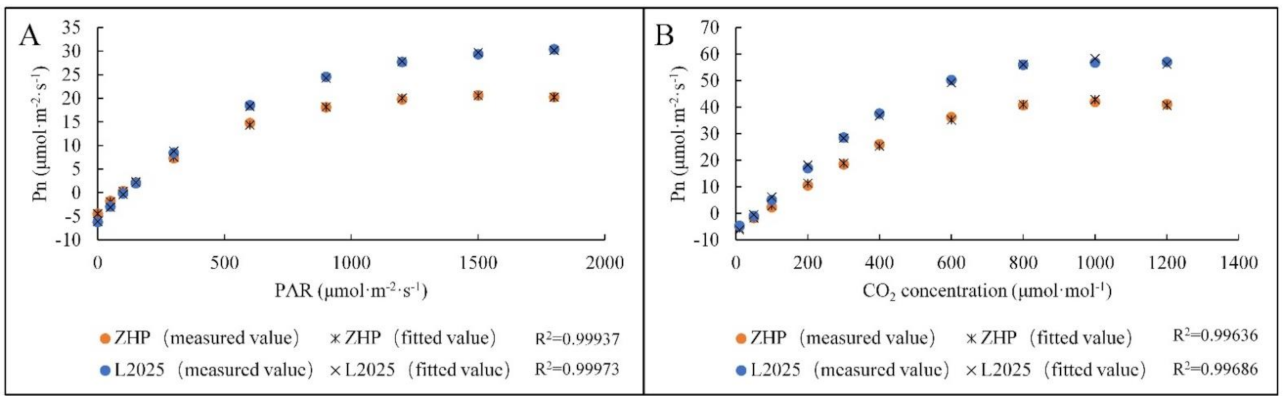
2.3. Model Fitting of Pn-PAR Response Curves and Pn-CO2 Concentration Response Curves in ‘L2025’ and ‘ZHP’ Seedling Leaves
2.4. Changes in Photosynthetic Characteristic Parameters between ‘L2025’ and ‘ZHP’ Leaves
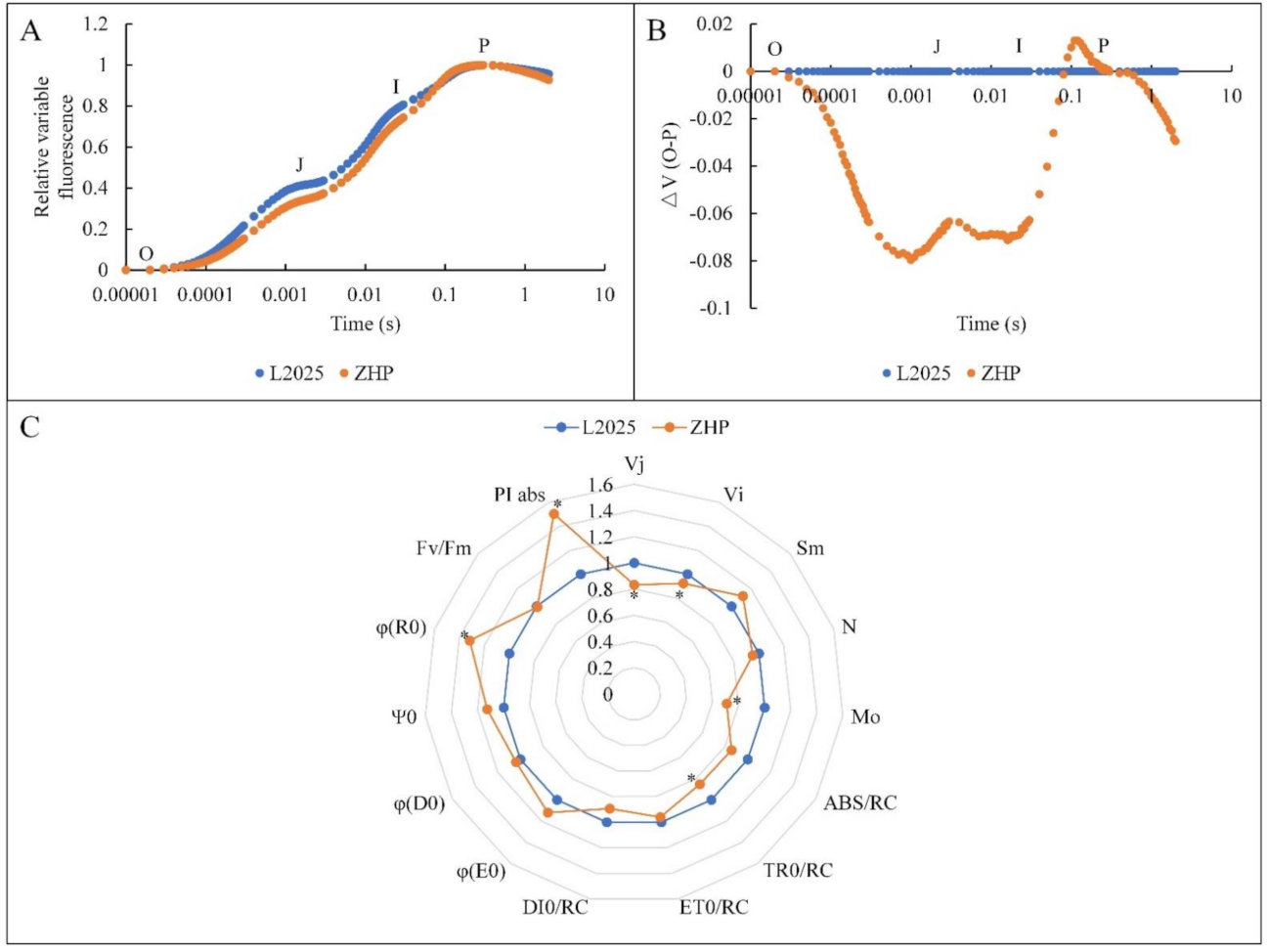
2.5. Changes in OJIP Curves between ‘L2025’ and ‘ZHP’ Leaves
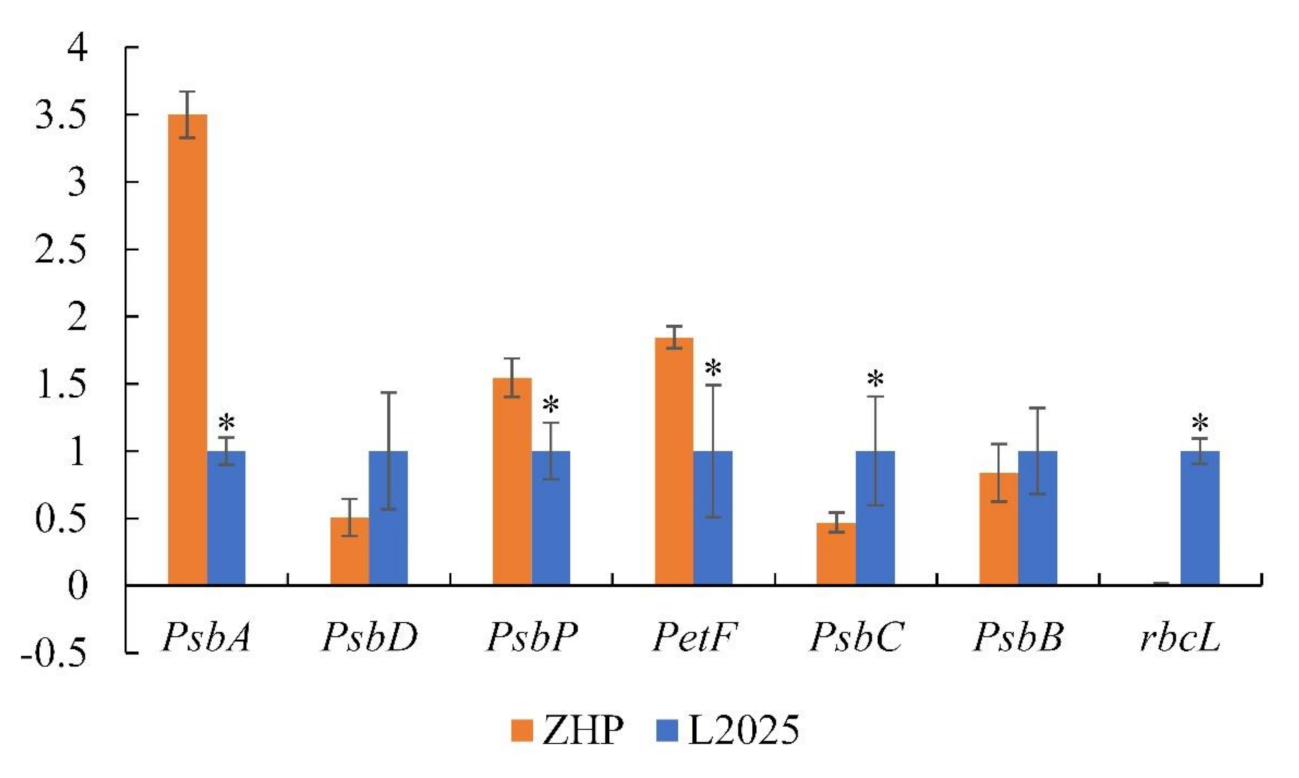
2.6. Changes in the Expression of Photosynthesis-Related Genes between ‘L2025’ and ‘ZHP’ Leaves
3. Materials and Methods
3.1. Plant Material and Growth Conditions
3.2. Measurement of Chl, Carotenoid and Anthocyanin Contents
3.3. Measurements of Light Response Curves and CO2 Response Curves
3.4. Measurement of Chl Fluorescence
3.5. Measurement of qPCR
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zhuang, W.; Wang, H.; Liu, T.; Wang, T.; Zhang, F.; Shu, X.; Zhai, H.; Wang, Z. Integrated physiological and genomic analysis reveals structural variations and expression patterns of candidate genes for colored-and green-leaf poplar. Sci. Rep. 2019, 9, 11150. [Google Scholar] [CrossRef]
- Zhang, K.; Wang, X.; Cui, J.; Ogweno, J.; Shi, K.; Zhou, Y.; Yu, J. Characteristics of gas exchange and chlorophyll fluorescence in red and green leaves of Begonia semperflorens. Biol. Plant. 2011, 55, 361–364. [Google Scholar] [CrossRef]
- Gao, M.Q.; Zhao, Y.Q.; Zong, Y.Z.; Wang, W.F. Photosynthetic Traits and Ecological Adaptability of Poplar 2025 and Its 3 Bud Sporting Color-leafed Cultivars. J. Northwest Forest. Univ. 2020, 35, 102–107. [Google Scholar]
- Zhang, F.; Wan, X.; Zheng, Y.; Sun, L.; Chen, Q.; Guo, Y.; Zhu, X.; Liu, M. Physiological and related anthocyanin biosynthesis genes responses induced by cadmium stress in a new colored-leaf plant “Quanhong Poplar”. Agroforest. Syst. 2014, 88, 343–355. [Google Scholar] [CrossRef]
- Song, Y.; Chen, Q.; Ci, D.; Shao, X.; Zhang, D. Effects of high temperature on photosynthesis and related gene expression in poplar. BMC Plant Biol. 2014, 14, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, G.; Zhang, S.; Guo, J.; Yang, R.; Li, H.; Fang, X.; Zhang, G. The effects of para-hydroxybenzoic acid treatment on photosynthetic parameters of Populus × euramericana “Neva”. Photosynthetica 2018, 56, 505–511. [Google Scholar] [CrossRef]
- Raveh, E.; Wang, N.; Nobel, P.S. Gas exchange and metabolite fluctuations in green and yellow bands of variegated leaves of the monocotyledonous CAM species Agave americana. Physiol. Plant. 1998, 103, 99–106. [Google Scholar] [CrossRef]
- Deng, X.; Zhang, H.; Wang, Y.; He, F.; Liu, J.; Xiao, X.; Shu, Z.; Li, W.; Wang, G.; Wang, G. Mapped clone and functional analysis of leaf-color gene Ygl7 in a rice hybrid (Oryza sativa L. ssp. indica). PLoS ONE 2014, 9, e99564. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Tang, S.; Zhang, S.; Shan, J.; Tang, C.; Chen, Q.; Jia, G.; Han, Y.; Zhi, H.; Diao, X. Gene mapping and functional analysis of the novel leaf color gene SiYGL1 in foxtail millet [Setaria italica (L.) P. Beauv]. Physiol. Plant. 2016, 157, 24–37. [Google Scholar] [CrossRef]
- Wu, H. Effect of different light qualities on growth, pigment content, chlorophyll fluorescence, and antioxidant enzyme activity in the red alga Pyropia haitanensis (Bangiales, Rhodophyta). BioMed Res. Int. 2016, 2016, 7383918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sytar, O.; Zivcak, M.; Neugart, S.; Toutounchi, P.; Brestic, M. Precultivation of young seedlings under different color shades modifies the accumulation of phenolic compounds in Cichorium leaves in later growth phases. Environ. Exp. Bot. 2019, 165, 30–38. [Google Scholar] [CrossRef]
- Torre, S.; Tattini, M.; Brunetti, C.; Guidi, L.; Gori, A.; Marzano, C.; Landi, M.; Sebastiani, F. De novo assembly and comparative transcriptome analyses of red and green morphs of sweet basil grown in full sunlight. PLoS ONE 2016, 11, e0160370. [Google Scholar] [CrossRef]
- Malik, T.; Wright, D.; Virk, D. Inheritance of net photosynthesis and transpiration efficiency in spring wheat, Triticum aestivum L., under drought. Plant Breed. 1999, 118, 93–95. [Google Scholar] [CrossRef]
- Rea, G.; Polticelli, F.; Antonacci, A.; Scognamiglio, V.; Katiyar, P.; Kulkarni, S.; Johanningmeier, U.; Giardi, M. Structure-based design of novel Chlamydomonas reinhardtii D1-D2 photosynthetic proteins for herbicide monitoring. Protein Sci. 2009, 18, 2139–2151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Wang, S.; Wang, P.; Gao, L.; Yang, R.; Li, Y. Label-free comparative proteomic and physiological analysis provides insight into leaf color variation of the golden-yellow leaf mutant of Lagerstroemia indica. J. Proteom. 2020, 228, 103942. [Google Scholar] [CrossRef]
- Zhong, X.; Sun, S.; Li, F.; Wang, J.; Shi, Z. Photosynthesis of a yellow-green mutant line in maize. Photosynthetica 2015, 53, 499–505. [Google Scholar] [CrossRef]
- Liu, M.; Lu, Y.; Wang, S.; Wu, F.; Li, J.; Wang, Y.; Zhao, J.; Shen, S. Characterization of the leaf color mutant hy and identification of the mutated gene in Chinese cabbage. J. Am. Soc. Hortic. Sci. 2018, 143, 363–369. [Google Scholar] [CrossRef] [Green Version]
- Sang, X.; Fang, L.; Vanichpakorn, Y.; Ling, Y.; Du, P.; Zhao, F.; Yang, Z.; He, G. Physiological character and molecular mapping of leaf-color mutant wyv1 in rice (Oryza sativa L.). Genes Genom. 2010, 32, 123–128. [Google Scholar] [CrossRef]
- Gould, K.; McKelvie, J.; Markham, K. Do anthocyanins function as antioxidants in leaves? Imaging of H2O2 in red and green leaves after mechanical injury. Plant Cell Environ. 2002, 25, 1261–1269. [Google Scholar] [CrossRef]
- Steyn, W.J.; Wand, S.; Holcroft, D.; Jacobs, G. Anthocyanins in vegetative tissues: A proposed unified function in photoprotection. New Phytol. 2002, 155, 349–361. [Google Scholar] [CrossRef]
- Gould, K.; Jay-Allemand, C.; Logan, B.; Baissac, Y.; Bidel, L. When are foliar anthocyanins useful to plants? Re-evaluation of the photoprotection hypothesis using Arabidopsis thaliana mutants that differ in anthocyanin accumulation. Environ. Exp. Bot. 2018, 154, 11–22. [Google Scholar] [CrossRef]
- Stirbet, A. On the relation between the Kautsky effect (chlorophyll a fluorescence induction) and photosystem II: Basics and applications of the OJIP fluorescence transient. J. Photochem. Photobiol. B 2011, 104, 236–257. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Zhang, D.; Zhou, L.; Zhang, X.; Liao, J.; Duan, Y.; Wen, B.; Ma, Y.; Wang, Y.; Fang, W. Transcriptomic and metabolomic profiling of Camellia sinensis L. cv. ‘Suchazao’ exposed to temperature stresses reveals modification in protein synthesis and photosynthetic and anthocyanin biosynthetic pathways. Tree Physiol. 2019, 39, 1583–1599. [Google Scholar] [CrossRef]
- Wang, T.; Luo, S.; Ma, Y.; Li, L.; Xie, Y.; Zhang, W. Chlorophyll a Fluorescence Transient and 2-Dimensional Electrophoresis Analyses Reveal Response Characteristics of Photosynthesis to Heat Stress in Malus.‘Prairifire’. Plants 2020, 9, 1040. [Google Scholar] [CrossRef]
- Kós, P.B.; Deák, Z.; Cheregi, O.; Vass, I. Differential regulation of psbA and psbD gene expression, and the role of the different D1 protein copies in the cyanobacterium Thermosynechococcus elongatus BP-1. BBA Bioenerg. 2008, 1777, 74–83. [Google Scholar] [CrossRef] [Green Version]
- Jain, A.; Cao, A.; Karthikeyan, A.; Baldwin, J.; Raghothama, K. Phosphate deficiency suppresses expression of light-regulated psbO and psbP genes encoding extrinsic proteins of oxygen-evolving complex of PSII. Curr. Sci. India 2005, 89, 1592–1596. [Google Scholar]
- Molina-Heredia, F.P.; Hervás, M.; Navarro, J.A.; Miguel, A. Cloning and Correct Expression in Escherichia coli of the petE and petJ Genes Respectively Encoding Plastocyanin and Cytochrome c6 from the Cyanobacterium Anabaena sp. PCC 7119. Biochem. Bioph. Res. Commun. 1998, 243, 302–306. [Google Scholar] [CrossRef] [PubMed]
- Okada, K. PetH is rate-controlling in the interaction between PetH, a component of the supramolecular complex with photosystem II, and PetF, a light-dependent electron transfer protein. Bioph. Res. Commun. 2009, 389, 394–398. [Google Scholar] [CrossRef]
- Floß, B.; Igloi, G.L.; Cassier-Chauvat, C.; Mühlenhoff, U. Molecular characterization and overexpression of the petF gene from Synechococcus elongatus: Evidence for a second site of electrostatic interaction between ferredoxin and the PSI-D subunit. Photosynth. Res. 1997, 54, 63–71. [Google Scholar] [CrossRef]
- Chase, M.W.; Soltis, D.E.; Olmstead, R.G.; Morgan, D.; Les, D.H.; Mishler, B.D.; Duvall, M.R.; Price, R.A.; Hills, H.G.; Qiu, Y. Phylogenetics of seed plants: An analysis of nucleotide sequences from the plastid gene rbcL. Ann. Mo. Bot. Gard. 1993, 80, 528–580. [Google Scholar] [CrossRef] [Green Version]
- Taylor, G. Populus: Arabidopsis for forestry. Do we need a model tree? Ann. Bot-Lond. 2002, 90, 681–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, S.Z.; Zhang, Z.Y.; Zhang, Q.; Lin, Y.Z. Progress in the study of molecular genetic improvements of poplar in China. J. Integr. Plant Biol. 2006, 48, 1001–1007. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, L.; Chen, B.; Qin, Z.; Xiao, Y.; Zhang, Y.; Yao, R.; Liu, H.; Yang, H. Progress in understanding the physiological and molecular responses of Populus to salt stress. Int. J. Mol. Sci. 2019, 20, 1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Cheng, X. A new poplar red foliar variety ‘Zhonghong’. Sci. Silvae Sin. 2008, 44, 173–174. [Google Scholar]
- Sheng, Q.; Zhu, Z. Photosynthetic Capacity, Stomatal Behavior and Chloroplast Ultrastructure in Leaves of the Endangered Plant Carpinus putoensis WC Cheng during Gaseous NO2 Exposure and after Recovery. Forests 2018, 9, 561. [Google Scholar] [CrossRef] [Green Version]
- Nisar, N.; Li, L.; Lu, S.; Khin, N.C.; Pogson, B.J. Carotenoid metabolism in plants. Mol. Plant 2015, 8, 68–82. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Jiang, W.; Yu, H. Effects of exogenous epibrassinolide on photosynthetic characteristics in tomato (Lycopersicon esculentum Mill) seedlings under weak light stress. J. Agr. Food Chem. 2010, 58, 3642–3645. [Google Scholar] [CrossRef]
- Lo Piccolo, E.; Landi, M.; Pellegrini, E.; Agati, G.; Giordano, C.; Giordani, T.; Lorenzini, G.; Malorgio, F.; Massai, R.; Nali, C. Multiple consequences induced by epidermally-located anthocyanins in young, mature and senescent leaves of Prunus. Front. Plant Sci. 2018, 9, 917. [Google Scholar] [CrossRef] [PubMed]
- Yan, N.; Xu, X.F.; Wang, Z.D.; Huang, J.Z.; Guo, D.P. Interactive effects of temperature and light intensity on photosynthesis and antioxidant enzyme activity in Zizania latifolia Turcz. plants. Photosynthetica 2013, 51, 127–138. [Google Scholar] [CrossRef]
- Feng, L.; Raza, M.A.; Li, Z.; Chen, Y.; Khalid, M.H.B.; Du, J.; Liu, W.; Wu, X.; Song, C.; Yu, L. The influence of light intensity and leaf movement on photosynthesis characteristics and carbon balance of soybean. Front. Plant Sci. 2019, 9, 1952. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.; Tran, T.; Nguyen, Q. Effects of light intensity on the growth, photosynthesis and leaf microstructure of hydroponic cultivated spinach (Spinacia oleracea L.) under a combination of red and blue LEDs in house. Int. J. Agric. Tech. 2019, 15, 75–90. [Google Scholar]
- Farquhar, G.D.; Sharkey, T.D. Stomatal conductance and photosynthesis. Annu. Rev. Plant Physiol. 1982, 33, 317–345. [Google Scholar] [CrossRef]
- Vialet-Chabrand, S.R.; Matthews, J.S.; McAusland, L.; Blatt, M.R.; Griffiths, H.; Lawson, T. Temporal dynamics of stomatal behavior: Modeling and implications for photosynthesis and water use. Plant Physiol. 2017, 174, 603–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sade, D.; Sade, N.; Shriki, O.; Lerner, S.; Gebremedhin, A.; Karavani, A.; Brotman, Y.; Osorio, S.; Fernie, A.R.; Willmitzer, L. Water balance, hormone homeostasis, and sugar signaling are all involved in tomato resistance to tomato yellow leaf curl virus. Plant Physiol. 2014, 165, 1684–1697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Andersen, M.N.; Jacobsen, S.E.; Jensen, C.R. Stomatal control and water use efficiency of soybean (Glycine max L. Merr.) during progressive soil drying. Environ. Exp. Bot. 2005, 54, 33–40. [Google Scholar] [CrossRef]
- Liu, E.; Mei, X.; Yan, C.; Gong, D.; Zhang, Y. Effects of water stress on photosynthetic characteristics, dry matter translocation and WUE in two winter wheat genotypes. Agr. Water Manag. 2016, 167, 75–85. [Google Scholar] [CrossRef]
- Leegood, R.C. C4 photosynthesis: Principles of CO2 concentration and prospects for its introduction into C3 plants. J. Exp. Bot. 2002, 53, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Leakey, A.D.; Uribelarrea, M.; Ainsworth, E.A.; Naidu, S.L.; Rogers, A.; Ort, D.R.; Long, S.P. Photosynthesis, productivity, and yield of maize are not affected by open-air elevation of CO2 concentration in the absence of drought. Plant Physiol. 2006, 140, 779–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dusenge, M.E.; Duarte, A.G.; Way, D.A. Plant carbon metabolism and climate change: Elevated CO2 and temperature impacts on photosynthesis, photorespiration and respiration. New Phytol. 2019, 221, 32–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharkey, T.D.; Stitt, M.; Heineke, D.; Gerhardt, R.; Raschke, K.; Heldt, H.W. Limitation of photosynthesis by carbon metabolism: II. O2-insensitive CO2 uptake results from limitation of triose phosphate utilization. Plant Physiol. 1986, 81, 1123–1129. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Rogers, A. The response of photosynthesis and stomatal conductance to rising [CO2]: Mechanisms and environmental interactions. Plant Cell Environ. 2007, 30, 258–270. [Google Scholar] [CrossRef]
- Fatichi, S.; Leuzinger, S.; Paschalis, A.; Langley, J.A.; Barraclough, A.D.; Hovenden, M.J. Partitioning direct and indirect effects reveals the response of water-limited ecosystems to elevated CO2. Proc. Natl. Acad. Sci. USA 2016, 113, 12757–12762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swann, A.L.; Hoffman, F.M.; Koven, C.D.; Randerson, J.T. Plant responses to increasing CO2 reduce estimates of climate impacts on drought severity. Proc. Natl. Acad. Sci. USA 2016, 113, 10019–10024. [Google Scholar] [CrossRef] [Green Version]
- Ye, Z.P.; Suggett, D.J.; Robakowski, P.; Kang, H.J. A mechanistic model for the photosynthesis–light response based on the photosynthetic electron transport of photosystem II in C3 and C4 species. New Phytol. 2013, 199, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.P.; Yu, Q. Comparison of a new model of light response of photosynthesis with traditional models. J. Shenyang Agr. Univ. 2007, 38, 771–775. [Google Scholar]
- Chen, Z.Y.; Peng, Z.; Yang, J.; Chen, W.Y.; Ou-Yang, Z.M. A mathematical model for describing light-response curves in Nicotiana tabacum L. Photosynthetica 2011, 49, 467–471. [Google Scholar] [CrossRef]
- Lang, Y.; Wang, M.; Zhang, G.; Zhao, Q. Experimental and simulated light responses of photosynthesis in leaves of three tree species under different soil water conditions. Photosynthetica 2013, 51, 370–378. [Google Scholar] [CrossRef]
- Fang, L.; Zhang, S.; Zhang, G.; Liu, X.; Xia, X.; Zhang, S.; Xing, W.; Fang, X. Application of five light-response models in the photosynthesis of Populus × euramericana cv. ‘Zhonglin46′ leaves. Appl. Biochem. Biotechnol. 2015, 176, 86–100. [Google Scholar] [CrossRef]
- Ye, Z.; Yu, Q. A comparison of response curves of winter wheat photosynthesis to flag leaf inte rcellular and air CO2 concentrations. Chin. J. Ecol. 2009, 28, 2233–2238. [Google Scholar]
- Ye, Z.; Zhao, Z. A modified rectangular hyperbola to describe the light-response curve of photosynthesis of Bidens pilosa L. grown under low and high light conditions. Front. Agr. China 2010, 4, 50–55. [Google Scholar] [CrossRef]
- Li, Y.; Liu, X.; Hao, K.; Yang, Q.; Yang, X.; Zhang, W.; Cong, Y. Light-response curve of photosynthesis and model fitting in leaves of Mangifera indica under different soil water conditions. Photosynthetica 2019, 57, 796–803. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.; Wu, L.; Chen, J.; Dong, L. Morphological plasticity, photosynthesis and chlorophyll fluorescence of Athyrium pachyphlebium at different shade levels. Photosynthetica 2011, 49, 611–618. [Google Scholar] [CrossRef]
- Wang, T.; Yang, W.; Xie, Y.; Shi, D.; Ma, Y.; Sun, X. Effects of exogenous nitric oxide on the photosynthetic characteristics of bamboo (Indocalamus barbatus McClure) seedlings under acid rain stress. Plant Growth Regul. 2017, 82, 69–78. [Google Scholar] [CrossRef]
- Chen, S.; Yang, J.; Zhang, M.; Strasser, R.J.; Qiang, S. Classification and characteristics of heat tolerance in Ageratina adenophora populations using fast chlorophyll a fluorescence rise OJIP. Environ. Exp. Bot. 2016, 122, 126–140. [Google Scholar] [CrossRef]
- Zeng, X.; Chow, W.; Su, L.; Peng, X.; Peng, C. Protective effect of supplemental anthocyanins on Arabidopsis leaves under high light. Physiol. Plant. 2010, 138, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Li, P.M.; Gao, H.Y.; Strasser, R.J. Application of the Fast Chlorophyll Fluorescence Induction Dynamics Analysis in Photosynthesis Study. J. Plant Physiol. Mol. Biol. 2005, 31, 559–566. [Google Scholar]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the chlorophyll a fluorescence transient. In Chlorophyll a Fluorescence; Springer: Berlin/Heidelberg, Germany, 2004; pp. 321–362. [Google Scholar]
- Faseela, P.; Sinisha, A.; Brestič, M.; Puthur, J. Chlorophyll a fluorescence parameters as indicators of a particular abiotic stress in rice. Photosynthetica 2019, 57, 108–115. [Google Scholar]
- Demetriou, G.; Neonaki, C.; Navakoudis, E.; Kotzabasis, K. Salt stress impact on the molecular structure and function of the photosynthetic apparatus—The protective role of polyamines. BBA Biochim. Biophys. Acta Bioenerg. 2007, 1767, 272–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfannschmidt, T. Chloroplast redox signals: How photosynthesis controls its own genes. Trends Plant Sci. 2003, 8, 33–41. [Google Scholar] [CrossRef] [Green Version]
- Pospíšil, P. Molecular mechanisms of production and scavenging of reactive oxygen species by photosystem II. BBA Biochim. Biophys. Acta Bioenerg. 2012, 1817, 218–231. [Google Scholar] [CrossRef] [Green Version]
- Jansen, M.A.; Gaba, V.; Greenberg, B.M.; Mattoo, A.K.; Edelman, M. Low threshold levels of ultraviolet-B in a background of photosynthetically active radiation trigger rapid degradation of the D2 protein of photosystem-II. Plant J. 1996, 9, 693–699. [Google Scholar] [CrossRef]
- Bi, A.; Fan, J.; Hu, Z.; Wang, G.; Amombo, E.; Fu, J.; Hu, T. Differential acclimation of enzymatic antioxidant metabolism and photosystem II photochemistry in tall fescue under drought and heat and the combined stresses. Front. Plant Sci. 2016, 7, 453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshioka, M.; Uchida, S.; Mori, H.; Komayama, K.; Ohira, S.; Morita, N.; Nakanishi, T.; Yamamoto, Y. Quality control of photosystem II: Cleavage of reaction center D1 protein in spinach thylakoids by FtsH protease under moderate heat stress. J. Biol. Chem. 2006, 281, 21660–21669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, F.; Chen, H.; Li, X.J.; Yang, M.F.; Liu, G.S.; Shen, S.H. A comparative proteomic analysis of rice seedlings under various high-temperature stresses. BBA Proteins Proteom. 2009, 1794, 1625–1634. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chl a (mg/g FW) | Chl b (mg/g FW) | Chl a/Chl b | Total Chl (mg/g FW) | Carotenoid (mg/g FW) | Anthocyanin (mg/g FW) | |
|---|---|---|---|---|---|---|
| ZHP | 1.622 ± 0.104 | 0.428 ± 0.037 * | 3.799 ± 0.167 * | 2.049 ± 0.137 | 0.389 ± 0.032 | 0.268 ± 0.030 * |
| 2025 | 1.604 ± 0.047 | 0.375 ± 0.013 | 4.275 ± 0.103 | 1.980 ± 0.058 | 0.371 ± 0.011 | 0.016 ± 0.002 |
| AQE | Rd (μmol·m−2·s−1) | Amax (μmol·m−2·s−1) | LCP (μmol·m−2·s−1) | LSP (μmol·m−2·s−1) | |
|---|---|---|---|---|---|
| ZHP | 0.052 ± 0.001 * | 4.377 ± 0.590 * | 19.287 ± 1.705 * | 92.833 ± 13.324 | 1579.150 ± 145.846 |
| L2025 | 0.062 ± 0.001 | 6.262 ± 0.239 | 29.508 ± 0.577 | 109.719 ± 5.970 | 1743.047 ± 70.205 |
| CE (μmol·m−2·s−1) | Rp (μmol·m−2·s−1) | CCP (μmol·mol−1) | CSP (μmol·mol −1) | |
|---|---|---|---|---|
| ZHP | 0.106 ± 0.013 * | 6.598 ± 0.509 * | 65.046 ± 3.825 | 995.966 ± 20.032 |
| L2025 | 0.142 ± 0.006 | 8.067 ± 0.291 | 59.064 ± 4.637 | 983.085 ± 16.474 |
| Gene Name | Accession | Description | Forward Primer (5′to 3′) | Reverse Primer (5′ to 3′) |
|---|---|---|---|---|
| PsbA | Potri.013G143200 | photosystem II P680 reaction center D1 protein | CTTAGTTTCCGTCTGG GTAT | GCTCAGCCTGGAATA CAATC |
| PsbD | Potri.008G208600 | photosystem II P680 reaction center D2 protein | ATTAGGTGGCTTGTG GACTT | GAGCCAACCAAACTG ACCT |
| PsbP | Potri.010G210000 | photosystem II oxygen-evolving enhancer protein 2 | TCATTGAGTTGGGCTT CC | ATTGAAGGTTGCCTCT GC |
| PetF | Potri.001G470700 | ferredoxin | GCTGGTGCTTGCTCTT CAT | CAGCCTCTATCTGGTC TTC |
| PsbC | Potri.010G032700 | photosystem II CP43 chlorophyll apoprotein | TATTCCCTGAGGAGTT TCTAC | ATAAGTTCATTGCTCC GACCC |
| PsbB | Potri.011G113900 | photosystem II CP47 chlorophyll apoprotein | TGTTGAGTTCTATGGT GGTG | ATCGGATTTCAAAGT AGCAC |
| rbcL | Potri.012G062600 | ribulose-1,5-bisphosphate carboxylase/oxygenase | GAACTTGTAGCCTCA TCCG | TACGGAATCATCTCC AAAG |
| Actin2 | Potri.019G010400.1 | Actin | GCCATCTCTCATCGG AATGGAA | AGGGCAGTGATTTCC TTGCTCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, T.; Li, L.; Cheng, G.; Shu, X.; Wang, N.; Zhang, F.; Zhuang, W.; Wang, Z. Physiological and Molecular Analysis Reveals the Differences of Photosynthesis between Colored and Green Leaf Poplars. Int. J. Mol. Sci. 2021, 22, 8982. https://doi.org/10.3390/ijms22168982
Wang T, Li L, Cheng G, Shu X, Wang N, Zhang F, Zhuang W, Wang Z. Physiological and Molecular Analysis Reveals the Differences of Photosynthesis between Colored and Green Leaf Poplars. International Journal of Molecular Sciences. 2021; 22(16):8982. https://doi.org/10.3390/ijms22168982
Chicago/Turabian StyleWang, Tao, Lingyu Li, Guanghao Cheng, Xiaochun Shu, Ning Wang, Fengjiao Zhang, Weibing Zhuang, and Zhong Wang. 2021. "Physiological and Molecular Analysis Reveals the Differences of Photosynthesis between Colored and Green Leaf Poplars" International Journal of Molecular Sciences 22, no. 16: 8982. https://doi.org/10.3390/ijms22168982
APA StyleWang, T., Li, L., Cheng, G., Shu, X., Wang, N., Zhang, F., Zhuang, W., & Wang, Z. (2021). Physiological and Molecular Analysis Reveals the Differences of Photosynthesis between Colored and Green Leaf Poplars. International Journal of Molecular Sciences, 22(16), 8982. https://doi.org/10.3390/ijms22168982