
Anti-Cancer Effects of Cyclic Peptide ALOS4 in a Human Melanoma Mouse Model

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
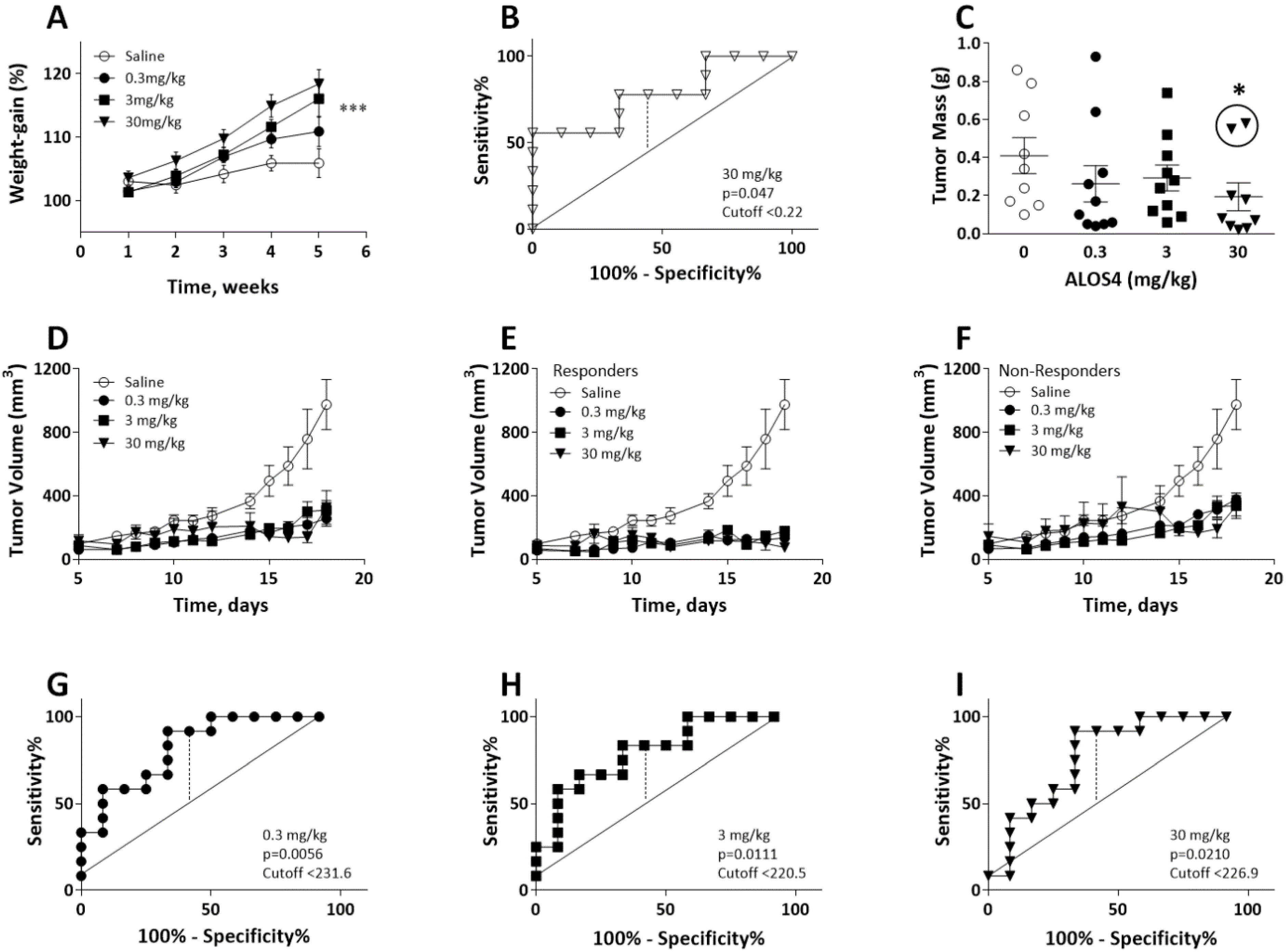
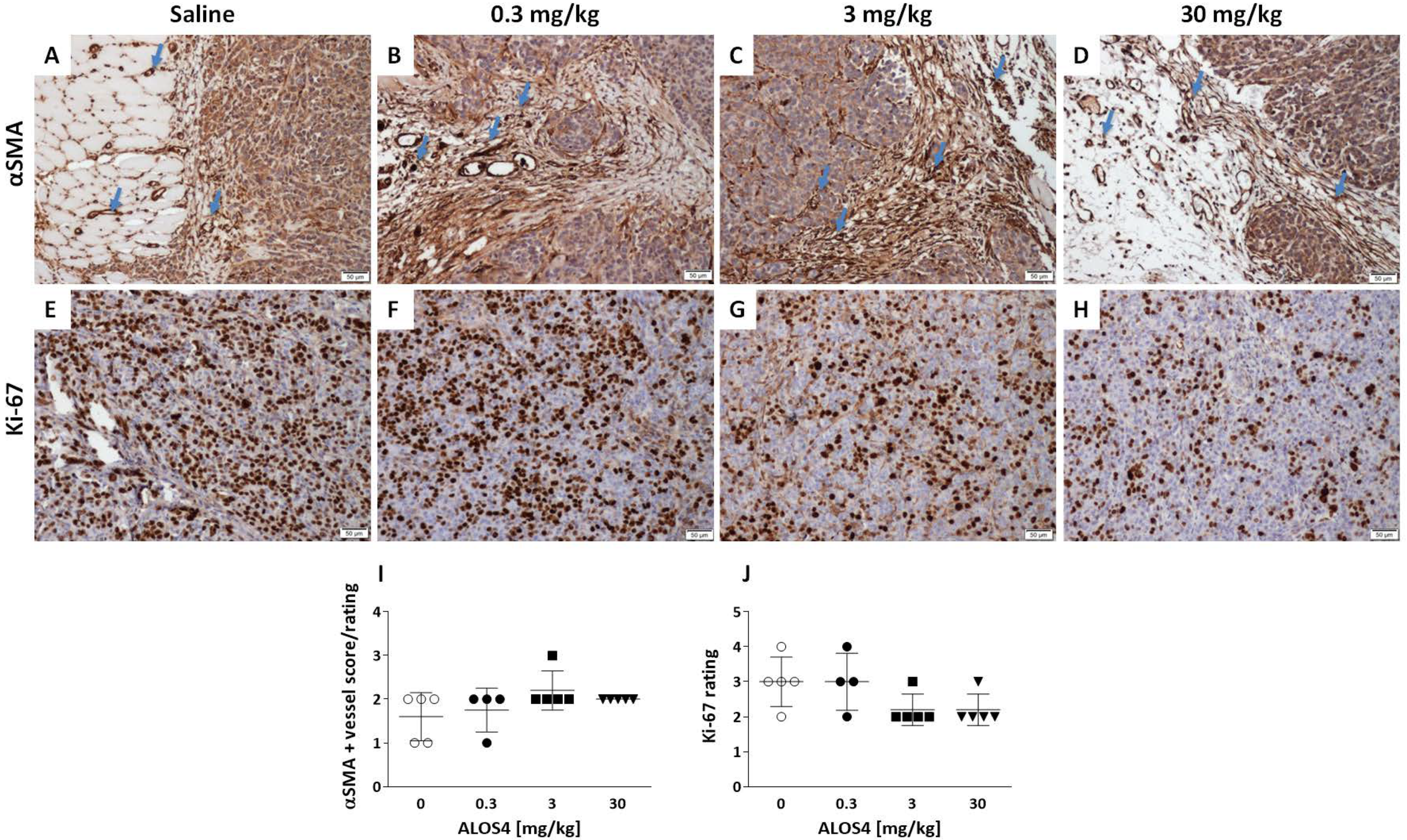
2.1. ALOS4 Selectively Affects Tumor Development in the A375 Xenograft Model
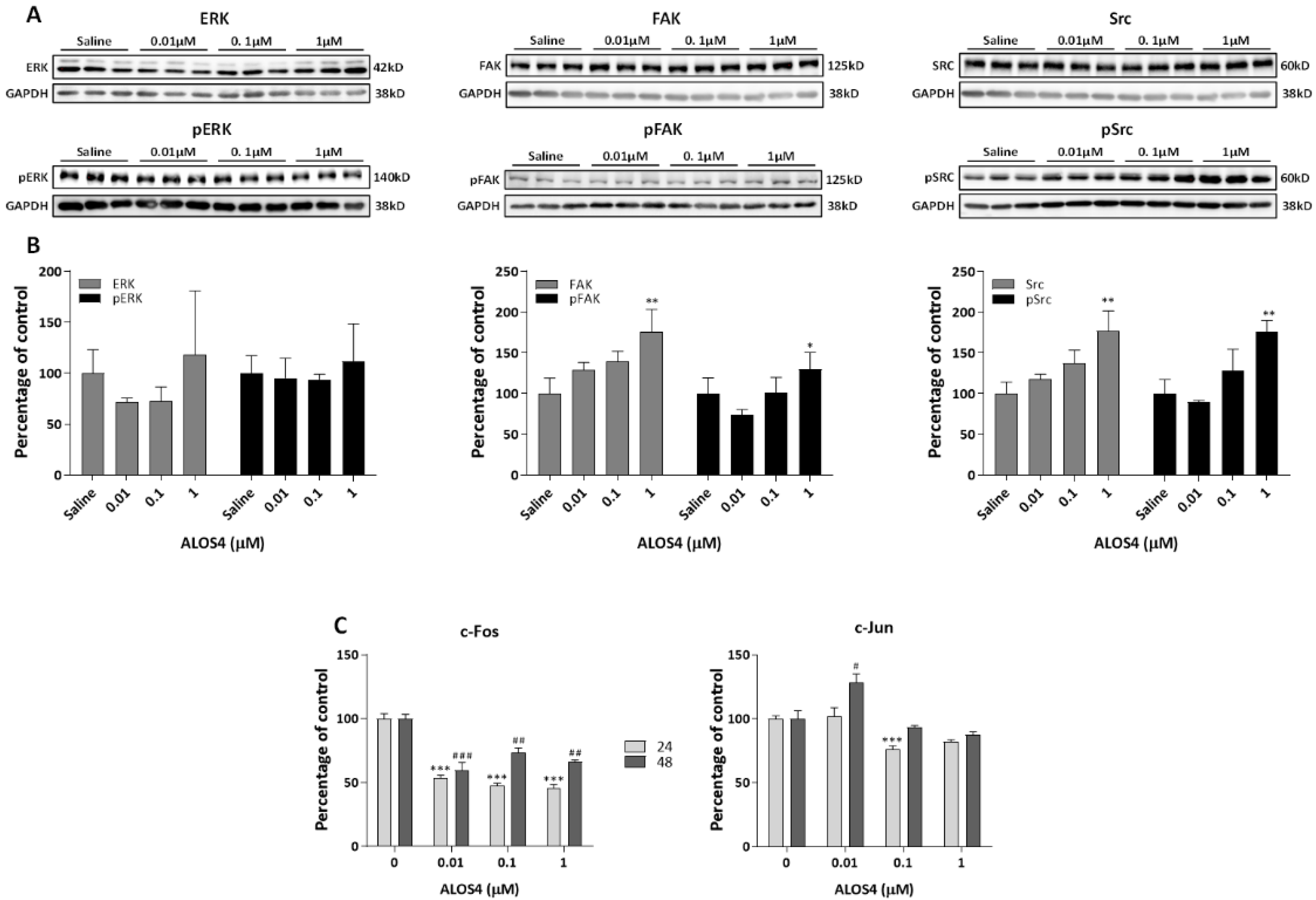
2.2. The Effect of ALOS4 on Integrin-Related Signal Transduction
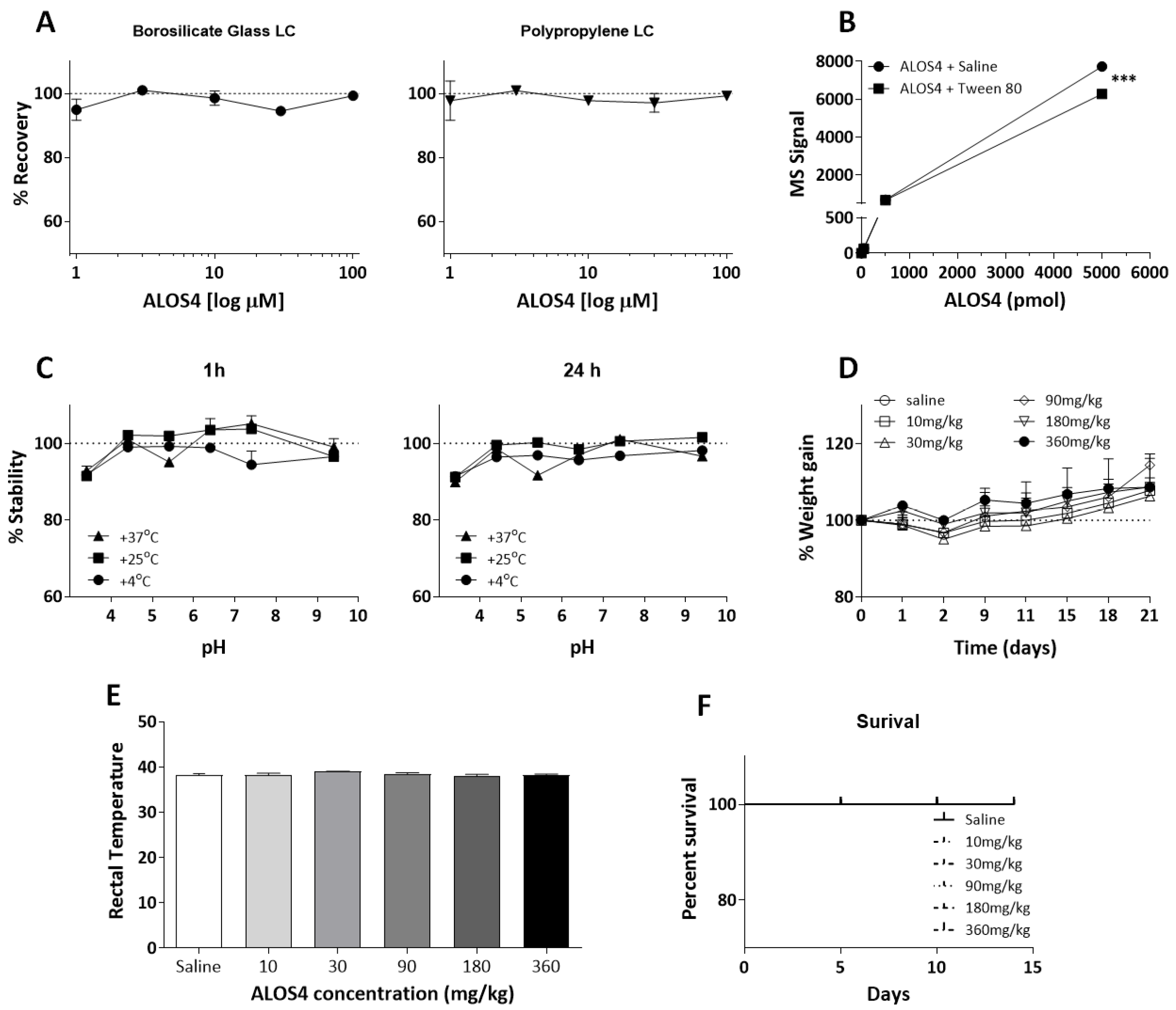
2.3. ALOS4 Does Not Adhere to Inert Materials and Is Stable over a Range of Acid/Base and Temperature Conditions
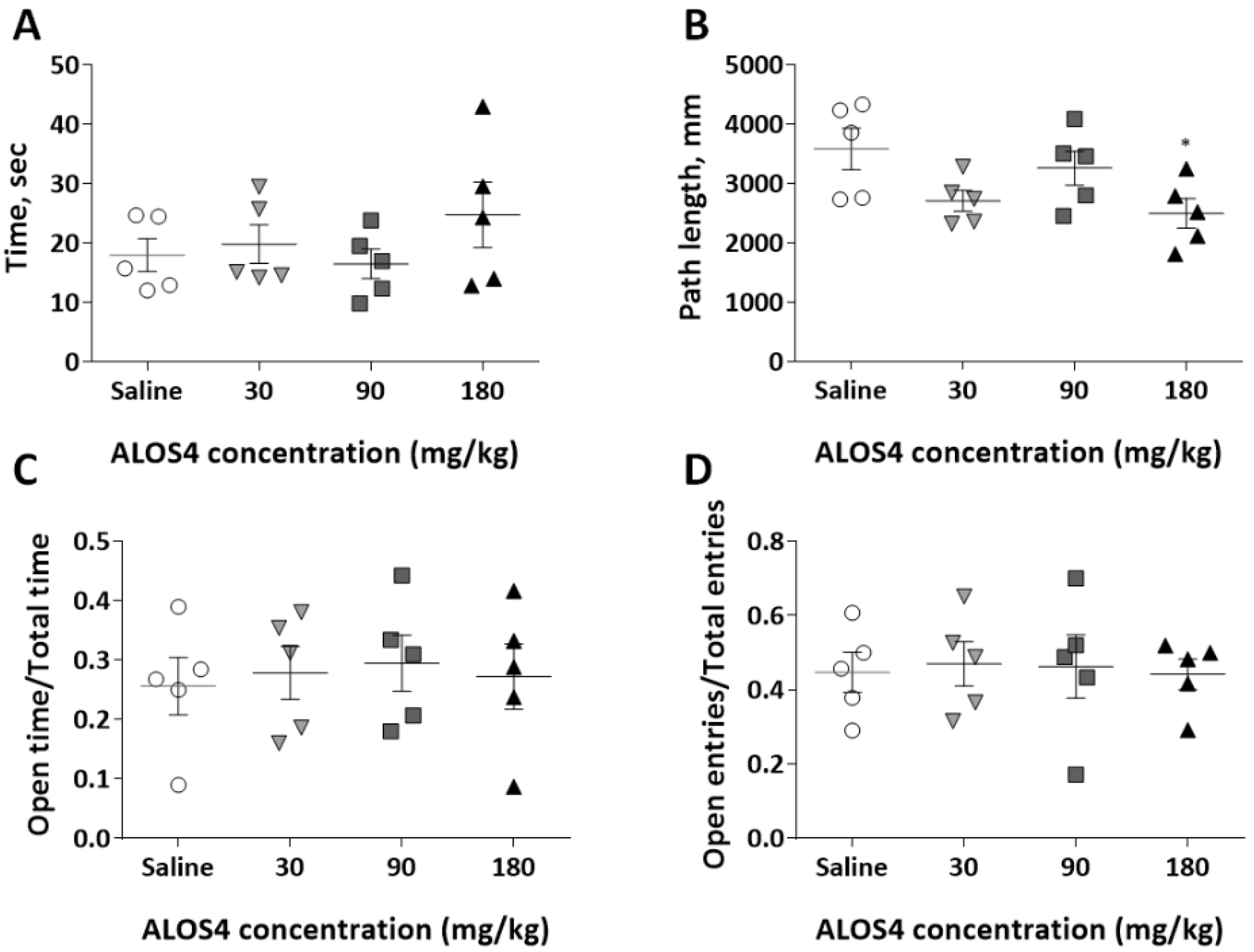
2.4. ALOS4 Shows High Safety and No Toxicity In Vivo
2.5. ALOS4 Does Not Affect Blood Cell Counts or Blood Chemistry
3. Discussion
4. Materials and Methods
4.1. ALOS4
4.2. Cell Cultures
4.3. Chemical Properties Assays
4.4. Animals
4.5. Behavioral Models
4.5.1. Open Field
4.5.2. Elevated plus Maze
4.5.3. Toxicity Assessment
4.5.4. Subcutaneous Model of Melanoma
4.5.5. Immunohistochemistry of A375 Tumor
4.5.6. RNA Extraction and qRT-PCR
4.5.7. Protein Extraction and Western Blot Analysis
4.5.8. Complete Blood Cell Count and Blood Chemistry
4.5.9. Statistical Analysis
5. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cooper, C.R.; Chay, C.H.; Pienta, K.J. The role of alpha(v)beta(3) in prostate cancer progression. Neoplasia 2002, 4, 191–194. [Google Scholar] [CrossRef] [Green Version]
- Takayama, S.; Ishii, S.; Ikeda, T.; Masamura, S.; Doi, M.; Kitajima, M. The relationship between bone metastasis from human breast cancer and integrin alpha(v)beta3 expression. Anticancer Res. 2005, 25, 79–83. [Google Scholar]
- Koistinen, P.; Heino, J. Integrins in Cancer Cell Invasion. In Madame Curie Bioscience Database [Internet]; Landes Bioscience: Austin, TX, USA, 2013. [Google Scholar]
- Attieh, Y.; Clark, A.G.; Grass, C.; Richon, S.; Pocard, M.; Mariani, P.; Elkhatib, N.; Betz, T.; Gurchenkov, B.; Vignjevic, D.M. Cancer-associated fibroblasts lead tumor invasion through integrin-beta3-dependent fibronectin assembly. J. Cell Biol. 2017, 216, 3509–3520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weis, S.M.; Cheresh, D.A. alphaV integrins in angiogenesis and cancer. Cold Spring Harb. Perspect. Med. 2011, 1, a006478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, J.P.; Stehle, T.; Zhang, R.; Joachimiak, A.; Frech, M.; Goodman, S.L.; Arnaout, M.A. Crystal structure of the extracellular segment of integrin alpha Vbeta3 in complex with an Arg-Gly-Asp ligand. Science 2002, 296, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Bellis, S.L. Advantages of RGD peptides for directing cell association with biomaterials. Biomaterials 2011, 32, 4205–4210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, L.A.; Learmonth, D.A.; Sousa, R.A.; Salgado, A.J. Alphavbeta3 and alpha5beta1 integrin-specific ligands: From tumor angiogenesis inhibitors to vascularization promoters in regenerative medicine? Biotechnol. Adv. 2018, 36, 208–227. [Google Scholar] [CrossRef]
- Sloan, E.K.; Pouliot, N.; Stanley, K.L.; Chia, J.; Moseley, J.M.; Hards, D.K.; Anderson, R.L. Tumor-specific expression of alphavbeta3 integrin promotes spontaneous metastasis of breast cancer to bone. Breast Cancer Res. BCR 2006, 8, R20. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Giancotti, F.G. Integrin signalling during tumour progression. Nat. Reviews. Mol. Cell Biol. 2004, 5, 816–826. [Google Scholar] [CrossRef]
- Rathinam, R.; Alahari, S.K. Important role of integrins in the cancer biology. Cancer Metastasis Rev. 2010, 29, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Bello, L.; Francolini, M.; Marthyn, P.; Zhang, J.; Carroll, R.S.; Nikas, D.C.; Strasser, J.F.; Villani, R.; Cheresh, D.A.; Black, P.M. Alpha(v)beta3 and alpha(v)beta5 integrin expression in glioma periphery. Neurosurgery 2001, 49, 380–389. [Google Scholar] [CrossRef]
- van den Hoogen, C.; van der Horst, G.; Cheung, H.; Buijs, J.T.; Pelger, R.C.; van der Pluijm, G. Integrin alphav expression is required for the acquisition of a metastatic stem/progenitor cell phenotype in human prostate cancer. Am. J. Pathol. 2011, 179, 2559–2568. [Google Scholar] [CrossRef]
- McCabe, N.P.; De, S.; Vasanji, A.; Brainard, J.; Byzova, T.V. Prostate cancer specific integrin alphavbeta3 modulates bone metastatic growth and tissue remodeling. Oncogene 2007, 26, 6238–6243. [Google Scholar] [CrossRef] [Green Version]
- Felding-Habermann, B.; O’Toole, T.E.; Smith, J.W.; Fransvea, E.; Ruggeri, Z.M.; Ginsberg, M.H.; Hughes, P.E.; Pampori, N.; Shattil, S.J.; Saven, A.; et al. Integrin activation controls metastasis in human breast cancer. Proc. Natl. Acad. Sci. USA 2001, 98, 1853–1858. [Google Scholar] [CrossRef] [Green Version]
- Huang, R.; Rofstad, E.K. Integrins as therapeutic targets in the organ-specific metastasis of human malignant melanoma. J. Exp. Clin. Cancer Res. CR 2018, 37, 92. [Google Scholar] [CrossRef]
- Heistein, J.B.; Acharya, U. Malignant Melanoma; StatPearls: Treasure Island, FL, USA, 2021. [Google Scholar]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA A Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [Green Version]
- Domingues, B.; Lopes, J.M.; Soares, P.; Populo, H. Melanoma treatment in review. ImmunoTargets Ther. 2018, 7, 35–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meerovitch, K.; Bergeron, F.; Leblond, L.; Grouix, B.; Poirier, C.; Bubenik, M.; Chan, L.; Gourdeau, H.; Bowlin, T.; Attardo, G. A novel RGD antagonist that targets both alphavbeta3 and alpha5beta1 induces apoptosis of angiogenic endothelial cells on type I collagen. Vasc. Pharmacol. 2003, 40, 77–89. [Google Scholar] [CrossRef]
- McNeel, D.G.; Eickhoff, J.; Lee, F.T.; King, D.M.; Alberti, D.; Thomas, J.P.; Friedl, A.; Kolesar, J.; Marnocha, R.; Volkman, J.; et al. Phase I trial of a monoclonal antibody specific for alphavbeta3 integrin (MEDI-522) in patients with advanced malignancies, including an assessment of effect on tumor perfusion. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2005, 11, 7851–7860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, M.A.; Paolillo, M.; Sanchez-Hernandez, Y.; Curti, D.; Ciusani, E.; Serra, M.; Colombo, L.; Schinelli, S. A small-molecule RGD-integrin antagonist inhibits cell adhesion, cell migration and induces anoikis in glioblastoma cells. Int. J. Oncol. 2013, 42, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Carter, A. Integrins as target: First phase III trial launches, but questions remain. J. Natl. Cancer Inst. 2010, 102, 675–677. [Google Scholar] [CrossRef] [PubMed]
- Hersey, P.; Sosman, J.; O’Day, S.; Richards, J.; Bedikian, A.; Gonzalez, R.; Sharfman, W.; Weber, R.; Logan, T.; Buzoianu, M.; et al. A randomized phase 2 study of etaracizumab, a monoclonal antibody against integrin alpha(v)beta(3), + or − dacarbazine in patients with stage IV metastatic melanoma. Cancer 2010, 116, 1526–1534. [Google Scholar] [CrossRef] [PubMed]
- Yacobovich, S.; Tuchinsky, L.; Kirby, M.; Kardash, T.; Agranyoni, O.; Nesher, E.; Redko, B.; Gellerman, G.; Tobi, D.; Gurova, K.; et al. Novel synthetic cyclic integrin alphavbeta3 binding peptide ALOS4: Antitumor activity in mouse melanoma models. Oncotarget 2016, 7, 63549–63560. [Google Scholar] [CrossRef] [Green Version]
- Redko, B.; Tuchinsky, H.; Segal, T.; Tobi, D.; Luboshits, G.; Ashur-Fabian, O.; Pinhasov, A.; Gerlitz, G.; Gellerman, G. Toward the development of a novel non-RGD cyclic peptide drug conjugate for treatment of human metastatic melanoma. Oncotarget 2017, 8, 757–768. [Google Scholar] [CrossRef] [Green Version]
- Harburger, D.S.; Calderwood, D.A. Integrin signalling at a glance. J. Cell Sci. 2009, 122, 159–163. [Google Scholar] [CrossRef] [Green Version]
- Vellon, L.; Menendez, J.A.; Lupu, R. A bidirectional “alpha(v)beta(3) integrin-ERK1/ERK2 MAPK” connection regulates the proliferation of breast cancer cells. Mol. Carcinog. 2006, 45, 795–804. [Google Scholar] [CrossRef]
- Dhillon, A.S.; Hagan, S.; Rath, O.; Kolch, W. MAP kinase signalling pathways in cancer. Oncogene 2007, 26, 3279–3290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serfilippi, L.M.; Pallman, D.R.; Russell, B. Serum clinical chemistry and hematology reference values in outbred stocks of albino mice from three commonly used vendors and two inbred strains of albino mice. Contemp. Top. Lab. Anim. Sci. 2003, 42, 46–52. [Google Scholar] [PubMed]
- Duffy, M.J.; McGowan, P.M.; Gallagher, W.M. Cancer invasion and metastasis: Changing views. J. Pathol. 2008, 214, 283–293. [Google Scholar] [CrossRef]
- Zuo, H. iRGD: A Promising Peptide for Cancer Imaging and a Potential Therapeutic Agent for Various Cancers. J. Oncol. 2019, 2019, 9367845. [Google Scholar] [CrossRef]
- Chinot, O.L. Cilengitide in glioblastoma: When did it fail? Lancet. Oncol. 2014, 15, 1044–1045. [Google Scholar] [CrossRef]
- Alberts, S.R.; Fishkin, P.A.; Burgart, L.J.; Cera, P.J.; Mahoney, M.R.; Morton, R.F.; Johnson, P.A.; Nair, S.; Goldberg, R.M.; North Central Cancer Treatment, G. CPT-11 for bile-duct and gallbladder carcinoma: A phase II North Central Cancer Treatment Group (NCCTG) study. Int. J. Gastrointest. Cancer 2002, 32, 107–114. [Google Scholar] [CrossRef]
- Zhang, J.; Mueller, S.T. A note on ROC analysis and non-parametric estimate of sensitivity. Psychometrika 2005, 70, 203–212. [Google Scholar] [CrossRef] [Green Version]
- Vasan, N.; Baselga, J.; Hyman, D.M. A view on drug resistance in cancer. Nature 2019, 575, 299–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balmativola, D.; Marchio, C.; Maule, M.; Chiusa, L.; Annaratone, L.; Maletta, F.; Montemurro, F.; Kulka, J.; Figueiredo, P.; Varga, Z.; et al. Pathological non-response to chemotherapy in a neoadjuvant setting of breast cancer: An inter-institutional study. Breast Cancer Res. Treat. 2014, 148, 511–523. [Google Scholar] [CrossRef] [Green Version]
- Gopalakrishnan, V.; Spencer, C.N.; Nezi, L.; Reuben, A.; Andrews, M.C.; Karpinets, T.V.; Prieto, P.A.; Vicente, D.; Hoffman, K.; Wei, S.C.; et al. Gut microbiome modulates response to anti-PD-1 immunotherapy in melanoma patients. Science 2018, 359, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Sherlach, K.S.; Roepe, P.D. Drug resistance associated membrane proteins. Front. Physiol. 2014, 5, 108. [Google Scholar] [CrossRef] [Green Version]
- Dhanapal, R.; Saraswathi, T.; Govind, R.N. Cancer cachexia. J. Oral Maxillofac. Pathol. JOMFP 2011, 15, 257–260. [Google Scholar] [CrossRef]
- Argiles, J.M.; Lopez-Soriano, F.J.; Stemmler, B.; Busquets, S. Therapeutic strategies against cancer cachexia. Eur. J. Transl. Myol. 2019, 29, 7960. [Google Scholar] [CrossRef] [Green Version]
- Fox, K.M.; Brooks, J.M.; Gandra, S.R.; Markus, R.; Chiou, C.F. Estimation of Cachexia among Cancer Patients Based on Four Definitions. J. Oncol. 2009, 2009, 693458. [Google Scholar] [CrossRef] [Green Version]
- Tisdale, M.J. Cachexia in cancer patients. Nat. Reviews. Cancer 2002, 2, 862–871. [Google Scholar] [CrossRef] [PubMed]
- Bolos, V.; Gasent, J.M.; Lopez-Tarruella, S.; Grande, E. The dual kinase complex FAK-Src as a promising therapeutic target in cancer. OncoTargets Ther. 2010, 3, 83–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawai, H.; Okada, Y.; Funahashi, H.; Matsuo, Y.; Takahashi, H.; Takeyama, H.; Manabe, T. Activation of focal adhesion kinase enhances the adhesion and invasion of pancreatic cancer cells via extracellular signal-regulated kinase-1/2 signaling pathway activation. Mol. Cancer 2005, 4, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.J.; Pan, W.W.; Liu, S.B.; Shen, Z.F.; Xu, Y.; Hu, L.L. ERK/MAPK signalling pathway and tumorigenesis. Exp. Ther. Med. 2020, 19, 1997–2007. [Google Scholar] [CrossRef] [Green Version]
- Oliva, I.B.; Coelho, R.M.; Barcellos, G.G.; Saldanha-Gama, R.; Wermelinger, L.S.; Marcinkiewicz, C.; Benedeta Zingali, R.; Barja-Fidalgo, C. Effect of RGD-disintegrins on melanoma cell growth and metastasis: Involvement of the actin cytoskeleton, FAK and c-Fos. Toxicon Off. J. Int. Soc. Toxinology 2007, 50, 1053–1063. [Google Scholar] [CrossRef] [PubMed]
- Wortzel, I.; Seger, R. The ERK Cascade: Distinct Functions within Various Subcellular Organelles. Genes Cancer 2011, 2, 195–209. [Google Scholar] [CrossRef]
- Yee, K.L.; Weaver, V.M.; Hammer, D.A. Integrin-mediated signalling through the MAP-kinase pathway. IET Syst. Biol. 2008, 2, 8–15. [Google Scholar] [CrossRef]
- Sundaram, M.V. Canonical RTK-Ras-ERK signaling and related alternative pathways. In WormBook: The Online Review of C. elegans Biology [Internet]; WormBook: Pasadena, CA, USA, 2018. [Google Scholar] [CrossRef]
- Goebel-Stengel, M.; Stengel, A.; Tache, Y.; Reeve, J.R., Jr. The importance of using the optimal plasticware and glassware in studies involving peptides. Anal. Biochem. 2011, 414, 38–46. [Google Scholar] [CrossRef] [Green Version]
- Furman, J.L.; Chiu, M.; Hunter, M.J. Early engineering approaches to improve peptide developability and manufacturability. AAPS J. 2015, 17, 111–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bottger, R.; Hoffmann, R.; Knappe, D. Differential stability of therapeutic peptides with different proteolytic cleavage sites in blood, plasma and serum. PLoS ONE 2017, 12, e0178943. [Google Scholar] [CrossRef] [PubMed]
- Gad, S.C. Preclinical Development Handbook: ADME and Biopharmaceutical Properties; Wiley: Hoboken, NJ, USA, 2008; p. 1352. [Google Scholar]
- Allen, D.D.; Caviedes, R.; Cardenas, A.M.; Shimahara, T.; Segura-Aguilar, J.; Caviedes, P.A. Cell lines as in vitro models for drug screening and toxicity studies. Drug Dev. Ind. Pharm. 2005, 31, 757–768. [Google Scholar] [CrossRef] [PubMed]
- Dolgos, H.; Freisleben, A.; Wimmer, E.; Scheible, H.; Kratzer, F.; Yamagata, T.; Gallemann, D.; Fluck, M. In vitro and in vivo drug disposition of cilengitide in animals and human. Pharmacol. Res. Perspect. 2016, 4, e00217. [Google Scholar] [CrossRef] [PubMed]
- Yassin, S.; Hu, J.; Xu, H.; Li, C.; Setrerrahmane, S. In vitro and in vivo activities of an antitumor peptide HM-3: A special dose-efficacy relationship on an HCT116 xenograft model in nude mice. Oncol. Rep. 2016, 36, 2951–2959. [Google Scholar] [CrossRef]
- Nurgali, K.; Jagoe, R.T.; Abalo, R. Editorial: Adverse Effects of Cancer Chemotherapy: Anything New to Improve Tolerance and Reduce Sequelae? Front. Pharmacol. 2018, 9, 245. [Google Scholar] [CrossRef] [PubMed]
- Gross, M.; Sheinin, A.; Nesher, E.; Tikhonov, T.; Baranes, D.; Pinhasov, A.; Michaelevski, I. Early onset of cognitive impairment is associated with altered synaptic plasticity and enhanced hippocampal GluA1 expression in a mouse model of depression. Neurobiol. Aging 2015, 36, 1938–1952. [Google Scholar] [CrossRef]
- Nesher, E.; Gross, M.; Lisson, S.; Tikhonov, T.; Yadid, G.; Pinhasov, A. Differential responses to distinct psychotropic agents of selectively bred dominant and submissive animals. Behav. Brain Res. 2013, 236, 225–235. [Google Scholar] [CrossRef]
- Gibson-Corley, K.N.; Olivier, A.K.; Meyerholz, D.K. Principles for valid histopathologic scoring in research. Vet. Pathol. 2013, 50, 1007–1015. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Normal Range | Saline | ALOS4 30 mg/kg | ALOS4 90 mg/kg | ALOS4 180 mg/kg | |
|---|---|---|---|---|---|
| WBC 103/µL | 6.5–24.5 | 5.49 ± 1.8 | 4.51 ± 1.3 | 6.07 ± 1.2 | 3.2 ± 0.06 |
| RBC 106/µL | 7.31–10.03 | 9.21 ± 0.8 | 9.16 ± 0.3 | 9.78 ± 0.3 | 8.02 ± 0.6 |
| HGB g/dL | 13.1–16.2 | 14.68 ± 1.02 | 14.3 ± 0.6 | 15.42 ± 0.5 | 13.2 ± 1.04 |
| Hematocrit % | 36.8–48.7 | 44.8 ± 3.2 | 43.96 ± 1.9 | 47.88 ± 1.1 | 40.12 ± 3.15 |
| MCV fL | 46.0–50.9 | 48.94 ± 1.2 | 47.96 ± 0.6 | 48.96 ± 0.24 | 49.96 ± 0.32 |
| MCV pg | 15–18 | 16.06 ± 0.3 | 15.6 ± 0.2 | 15.76 ± 0.15 | 16.42 ± 0.2 |
| MCHC g/dL | 33.7–36.4 | 32.82 ± 0.5 | 32.54 ± 0.23 | 32.2 ± 0.4 | 32.88 ± 0.4 |
| Platelets 103/µL | 674–1675 | 535.4 ± 139.7 | 582.4 ± 132 | 770.2 ± 189 | 661.8 ± 160.5 |
| Normal Range | Saline | ALOS4 30 mg/kg | ALOS4 90 mg/kg | ALOS4 180 mg/kg | |
|---|---|---|---|---|---|
| Creatinine mg/dL | 0.2–0.4 | 0.31 ± 0.03 | 0.26 ± 0.014 | 0.25 ± 0.06 | 0.27 ± 0.02 |
| Calcium mg/dL | 9.8–10.8 | 11.34 ± 0.25 | 10.29 ± 0.11 | 10.38 ± 0.29 | 10.42 ± 0.1 |
| Phosphorus mg/dL | 6.4–11.3 | 10.43 ± 0.9 | 8.6 ± 0.56 | 9.53 ± 0.4 | 8.44 ± 0.75 |
| Glucose mg/dL | 169–282 | 176.2 ± 7.4 | 169.8 ± 6.9 | 170.75 ± 10.7 | 183.2 ± 14.3 |
| Urea mg/dL | 39–62 | 55.48 ± 2.9 | 46.08 ± 1.7 | 48.23 ± 3.2 | 46.02 ± 3.9 |
| Cholesterol mg/dL | 56–133 | 140.4 ± 7.9 | 108.2 ± 9.5 | 142.25 ± 13.7 | 141.8 ± 9.1 |
| TP g/dL | 4.7–5.8 | 6.23 ± 0.11 | 6.17 ± 0.18 | 6.32 ± 0.13 | 6.28 ± 0.09 |
| Alb g/dL | 3.3–4.0 | 4.3 ± 0.09 | 4.3 ± 0.14 | 4.3 ± 0.06 | 4.36 ± 0.024 |
| Globulin g/dL | 1.4–2.0 | 1.93 ± 0.12 | 1.87 ± 0.07 | 2.02 ± 0.07 | 1.59 ± 0.4 |
| Total Bilirubin mg/dL | 0.16–0.32 | 0.1 ± 0.02 | 0.11 ± 0.02 | 0.14 ± 0.02 | 0.11 ± 0.03 |
| Alkaline Phos IU/L | 43–125 | 0.31 ± 0.03 | 0.26 ± 0.014 | 0.25 ± 0.06 | 0.27 ± 0.02 |
| SGOT IU/L | 69–191 | 11.34 ± 0.25 | 10.29 ± 0.11 | 10.38 ± 0.29 | 10.42 ± 0.1 |
| SGTP IU/L | 26–120 | 10.43 ± 0.9 | 8.6 ± 0.56 | 9.53 ± 0.4 | 8.44 ± 0.75 |
| Sodium mmol/L | 151–156 | 176.2 ± 7.4 | 169.8 ± 6.9 | 170.75 ± 10.7 | 183.2 ± 14.3 |
| Potassium mmol/L | 7.3–10.2 | 55.48 ± 2.9 | 46.08 ± 1.7 | 48.23 ± 3.2 | 46.02 ± 3.9 |
| Chloride mmol/L | 110–119 | 140.4 ± 7.9 | 108.2 ± 9.5 | 142.25 ± 13.7 | 141.8 ± 9.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Levi, B.; Yacobovich, S.; Kirby, M.; Becker, M.; Agranyoni, O.; Redko, B.; Gellerman, G.; Pinhasov, A.; Koman, I.; Nesher, E. Anti-Cancer Effects of Cyclic Peptide ALOS4 in a Human Melanoma Mouse Model. Int. J. Mol. Sci. 2021, 22, 9579. https://doi.org/10.3390/ijms22179579
Levi B, Yacobovich S, Kirby M, Becker M, Agranyoni O, Redko B, Gellerman G, Pinhasov A, Koman I, Nesher E. Anti-Cancer Effects of Cyclic Peptide ALOS4 in a Human Melanoma Mouse Model. International Journal of Molecular Sciences. 2021; 22(17):9579. https://doi.org/10.3390/ijms22179579
Chicago/Turabian StyleLevi, Bar, Shiri Yacobovich, Michael Kirby, Maria Becker, Oryan Agranyoni, Boris Redko, Gary Gellerman, Albert Pinhasov, Igor Koman, and Elimelech Nesher. 2021. "Anti-Cancer Effects of Cyclic Peptide ALOS4 in a Human Melanoma Mouse Model" International Journal of Molecular Sciences 22, no. 17: 9579. https://doi.org/10.3390/ijms22179579
APA StyleLevi, B., Yacobovich, S., Kirby, M., Becker, M., Agranyoni, O., Redko, B., Gellerman, G., Pinhasov, A., Koman, I., & Nesher, E. (2021). Anti-Cancer Effects of Cyclic Peptide ALOS4 in a Human Melanoma Mouse Model. International Journal of Molecular Sciences, 22(17), 9579. https://doi.org/10.3390/ijms22179579