
Cannabinoids—A New Perspective in Adjuvant Therapy for Pulmonary Hypertension


Abstract
:1. Introduction
2. Cannabinoids in the Cardiopulmonary System
3. Effects of Cannabinoids on Systemic Vessels
4. The Systemic Versus Pulmonary Circulation
5. Cannabinoids Affect Pulmonary Circulation
6. Endothelium-Dependent Mechanisms of Pulmonary Vasorelaxation
6.1. Arachidonic-Acid-Derived Pathway
6.2. Vasorelaxation’s Dependence on Calcium-Dependent Potassium Channels
6.3. Regulation of Pulmonary Vascular Tension by NO
7. Receptor-Mediated Vasodilatation
7.1. Mechanism Dependent on CB1-Rs and CB2-Rs
7.2. Other G-Protein-Dependent Receptors
7.3. Other G-Protein-Independent Receptors
8. Cannabinoids in PH—In Vivo and In Vitro Studies
9. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| 2-AGE | noladin ether |
| Abn-CBD | abnormal-cannabidiol |
| APCA | arachidonylcyclopropylamide |
| ARA-S | arachidonoyl-L-serine |
| CB1,2-R | cannabinoid receptor types 1 and 2 |
| CBD | cannabidiol |
| CBRs | cannabinoid receptors |
| DAGL-α,β | diacylglycerol lipases α,β |
| DHEA | docosahexaenoyl ethanolamid |
| DEA | docosatetraenoyl ethanolamide |
| ECS | endocannabinoid system |
| EDH | endothelium-dependent hyperpolarization |
| EPEA | eicosapentaenoyl ethanolamide |
| ERAs | endothelin receptor antagonists |
| HEA | homo-γ-linolenyl ethanolamide |
| hPAs | human pulmonary arteries |
| LEA | inolenoyl ethanolamide |
| 2-LG | linoleoylglycerol |
| LPI L | alpha-Lysophosphatidylinositol |
| MCT | monocrotaline |
| MethAEA | methanandamide |
| NAGly N | arachidonoyl glycine |
| ODA | oleamide |
| OEA | oleoyl ethanolamide |
| PAH | pulmonary arterial hypertension |
| POEA | palmitoleoyl ethanolamide |
| PAP | pulmonary arterial pressure |
| PASMCs | pulmonary artery smooth muscle cells |
| PEA | ethanolamide |
| PH | pulmonary hypertension |
| PPAR-γ | peroxisome proliferator-activated receptor-γ |
| PVR | pulmonary vascular resistance |
| rPA | rat pulmonary artery |
| RVSP | right ventricular systolic pressure |
| SEA | stearoyl ethanolamide |
| U46619 | analogue of thromboxane A2 |
| VIR | virodhamine |
References
- Mandras, S.A.; Mehta, H.S.; Vaidya, A. Pulmonary Hypertension: A Brief Guide for Clinicians. Mayo Clin. Proc. 2020, 95, 1978–1988. [Google Scholar] [CrossRef]
- Oliveira, A.C.; Richards, E.M.; Raizada, M.K. Pulmonary hypertension: Pathophysiology beyond the lung. Pharmacol. Res. 2020, 151, 104518. [Google Scholar] [CrossRef]
- Dodson, M.W.; Brown, L.M.; Elliott, C.G. Pulmonary Arterial Hypertension. Heart Fail. Clin. 2018, 14, 255–269. [Google Scholar] [CrossRef]
- Lan, N.S.H.; Massam, B.D.; Kulkarni, S.S.; Lang, C.C. Pulmonary Arterial Hypertension: Pathophysiology and Treatment. Diseases 2018, 6, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humbert, M.; Guignabert, C.; Bonnet, S.; Dorfmüller, P.; Klinger, J.R.; Nicolls, M.R.; Olschewski, A.J.; Pullamsetti, S.S.; Schermuly, R.T.; Stenmark, K.R.; et al. Pathology and pathobiology of pulmonary hypertension: State of the art and research perspectives. Eur. Respir. J. 2019, 53, 1801887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huertas, A.; Tu, L.; Humbert, M.; Guignabert, C. Chronic inflammation within the vascular wall in pulmonary arterial hypertension: More than a spectator. Cardiovasc. Res. 2020, 116, 885–893. [Google Scholar] [CrossRef]
- Coons, J.C.; Pogue, K.; Kolodziej, A.R.; Hirsch, G.A.; George, M.P. Pulmonary Arterial Hypertension: A Pharmacotherapeutic Update. Curr. Cardiol. Rep. 2019, 22, 141. [Google Scholar] [CrossRef]
- Mayeux, J.D.; Pan, I.Z.; Dechand, J.; Jacobs, J.A.; Jones, T.L.; McKellar, S.H.; Beck, E.; Hatton, N.D.; Ryan, J.J. Management of Pulmonary Arterial Hypertension. Curr. Cardiovasc. Risk Rep. 2021, 15, 2. [Google Scholar] [CrossRef]
- Kozłowska, H.; Baranowska, M.; Schlicker, E.; Kozłowski, M.; Laudański, J.; Malinowska, B. Identification of the vasodilatory endothelial cannabinoid receptor in the human pulmonary artery. J. Hypertens. 2007, 25, 2240–2248. [Google Scholar] [CrossRef] [PubMed]
- Hornig, B. Endothelial vasodilatory cannabinoid receptor in the human pulmonary artery: A future option in the therapy of pulmonary hypertension? J. Hypertens. 2007, 25, 2202–2203. [Google Scholar] [CrossRef]
- ElSohly, M.A.; Radwan, M.M.; Gul, W.; Chandra, S.; Galal, A. Phytochemistry of Cannabis sativa L. Prog. Chem. Org. Nat. Prod. 2017, 103, 1–36. [Google Scholar] [CrossRef]
- Malinowska, B.; Toczek, M.; Pędzińska-Betiuk, A.; Schlicker, E. Cannabinoids in arterial, pulmonary and portal hypertension-mechanisms of action and potential therapeutic significance. Br. J. Pharmacol. 2019, 176, 1395–1411. [Google Scholar] [CrossRef]
- Laezza, C.; Pagano, C.; Navarra, G.; Pastorino, O.; Proto, M.C.; Fiore, D.; Piscopo, C.; Gazzerro, P.; Bifulco, M. The Endocannabinoid System: A Target for Cancer Treatment. Int. J. Mol. Sci. 2020, 21, 747. [Google Scholar] [CrossRef] [Green Version]
- Stanley, C.P.; O’Sullivan, S.E. Cyclooxygenase metabolism mediates vasorelaxation to 2-arachidonoylglycerol (2-AG) in human mesenteric arteries. Pharmacol. Res. 2014, 81, 74–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bondarenko, A.I. Endothelial atypical cannabinoid receptor: Do we have enough evidence? Br. J. Pharmacol. 2014, 171, 5573–5588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacher, P.; Kogan, N.M.; Mechoulam, R. Beyond THC and Endocannabinoids. Annu. Rev. Pharmacol. Toxicol. 2020, 60, 637–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, S.P.H.; Christopoulos, A.; Davenport, A.P.; Kelly, E.; Mathie, A.; Peters, J.A.; Veale, E.L.; Armstrong, J.F.; Faccenda, E.; Harding, S.D.; et al. The Concise Guide to Pharmacology 2019/20: G protein-coupled receptors. Br. J. Pharmacol. 2019, 176, S21–S141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahn, H.; Wolf, J.; Kram, F.; Frantz, S.; Wagner, J.A. The endocannabinoid arachidonyl ethanolamide (anandamide) increases pulmonary arterial pressure via cyclooxygenase-2 products in isolated rabbit lungs. Am. J. Physiol. Heart Circ. Physiol. 2005, 289, H2491–H2496. [Google Scholar] [CrossRef] [Green Version]
- Sadowska, O.; Baranowska-Kuczko, M.; Gromotowicz-Popławska, A.; Biernacki, M.; Kicman, A.; Malinowska, B.; Kasacka, I.; Krzyżewska, A.; Kozłowska, H. Cannabidiol Ameliorates Monocrotaline-Induced Pulmonary Hypertension in Rats. Int. J. Mol. Sci. 2020, 21, 7077. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, D.; Matthey, M.; Bindila, L.; Lerner, R.; Lutz, B.; Zimmer, A.; Fleischmann, B.K. Endocannabinoid anandamide mediates hypoxic pulmonary vasoconstriction. Proc. Natl. Acad. Sci. USA 2013, 110, 18710–18715. [Google Scholar] [CrossRef] [Green Version]
- Karpińska, O.; Baranowska-Kuczko, M.; Kloza, M.; Ambrożewicz, E.; Kozłowski, T.; Kasacka, I.; Malinowska, B.; Kozłowska, H. Activation of CB1 receptors by 2-arachidonoylglycerol attenuates vasoconstriction induced by U46619 and angiotensin II in human and rat pulmonary arteries. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017, 312, R883–R893. [Google Scholar] [CrossRef] [Green Version]
- Baranowska-Kuczko, M.; Kozłowska, H.; Kloza, M.; Sadowska, O.; Kozłowski, M.; Kusaczuk, M.; Kasacka, I.; Malinowska, B. Vasodilatory effects of cannabidiol in human pulmonary and rat small mesenteric arteries: Modification by hypertension and the potential pharmacological opportunities. J. Hypertens. 2020, 38, 896–911. [Google Scholar] [CrossRef]
- Karpińska, O.; Baranowska-Kuczko, M.; Malinowska, B.; Kloza, M.; Kusaczuk, M.; Gęgotek, A.; Golec, P.; Kasacka, I.; Kozłowska, H. Mechanisms of l-alpha-lysophosphatidylinositol-induced relaxation in human pulmonary arteries. Life Sci. 2018, 192, 38–45. [Google Scholar] [CrossRef]
- Baranowska-Kuczko, M.; Kozłowska, H.; Kozłowski, M.; Schlicker, E.; Kloza, M.; Surażyński, A.; Grzęda, E.; Malinowska, B. Mechanisms of endothelium-dependent relaxation evoked by anandamide in isolated human pulmonary arteries. Naunyn Schmiedebergs Arch. Pharmacol. 2014, 387, 477–486. [Google Scholar] [CrossRef] [Green Version]
- Zou, S.; Kumar, U. Cannabinoid Receptors and the Endocannabinoid System: Signaling and Function in the Central Nervous System. Int. J. Mol. Sci. 2018, 19, 833. [Google Scholar] [CrossRef] [Green Version]
- Irrera, N.; D’Ascola, A.; Pallio, G.; Bitto, A.; Mannino, F.; Arcoraci, V.; Rottura, M.; Ieni, A.; Minutoli, L.; Metro, D.; et al. β-Caryophyllene Inhibits Cell Proliferation through a Direct Modulation of CB2 Receptors in Glioblastoma Cells. Cancers 2020, 12, 1038. [Google Scholar] [CrossRef] [Green Version]
- Picciolo, G.; Pallio, G.; Altavilla, D.; Vaccaro, M.; Oteri, G.; Irrera, N.; Squadrito, F. β-Caryophyllene Reduces the Inflammatory Phenotype of Periodontal Cells by Targeting CB2 Receptors. Biomedicines 2020, 8, 164. [Google Scholar] [CrossRef]
- Irrera, N.; D’Ascola, A.; Pallio, G.; Bitto, A.; Mazzon, E.; Mannino, F.; Squadrito, V.; Arcoraci, V.; Minutoli, L.; Campo, G.M.; et al. β-Caryophyllene Mitigates Collagen Antibody Induced Arthritis (CAIA) in Mice Through a Cross-Talk between CB2 and PPAR-γ Receptors. Biomolecules 2019, 9, 326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kicman, A.; Toczek, M. The Effects of Cannabidiol, a Non-Intoxicating Compound of Cannabis, on the Cardiovascular System in Health and Disease. Int. J. Mol. Sci. 2020, 21, 6740. [Google Scholar] [CrossRef] [PubMed]
- Toczek, M.; Malinowska, B. Enhanced endocannabinoid tone as a potential target of pharmacotherapy. Life Sci. 2018, 204, 20–45. [Google Scholar] [CrossRef] [PubMed]
- Di Marzo, V.; Ligresti, A.; Cristino, L. The endocannabinoid system as a link between homoeostatic and hedonic pathways involved in energy balance regulation. Int. J. Obes. 2009, 2, S18–S24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meccariello, R.; Santoro, A.; D’Angelo, S.; Morrone, R.; Fasano, S.; Viggiano, A.; Pierantoni, R. The Epigenetics of the Endocannabinoid System. Int. J. Mol. Sci. 2020, 21, 1113. [Google Scholar] [CrossRef]
- Calder, P.C. Eicosanoids. Essays Biochem. 2020, 64, 423–441. [Google Scholar] [CrossRef] [PubMed]
- Malinowska, B.; Baranowska-Kuczko, M.; Schlicker, E. Triphasic blood pressure responses to cannabinoids: Do we understand the mechanism? Br. J. Pharmacol. 2012, 165, 2073–2088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, E.N.; Kelly, M.E.; Cringle, S.J.; Yu, D.Y. Role of Endothelium in Abnormal Cannabidiol-Induced Vasoactivity in Retinal Arterioles. Investig. Ophthalmol. Vis. Sci. 2015, 56, 4029–4037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, W.S.; Zheng, X.; Zhang, D.X. Role of endothelial TRPV4 channels in vascular actions of the endocannabinoid, 2-arachidonoylglycerol. Br. J. Pharmacol. 2015, 172, 5251–5264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanley, C.P.; Hind, W.H.; Tufarelli, C.; O’Sullivan, S.E. Cannabidiol causes endothelium-dependent vasorelaxation of human mesenteric arteries via CB1 activation. Cardiovasc. Res. 2015, 107, 568–578. [Google Scholar] [CrossRef] [Green Version]
- Stanley, C.P.; Hind, W.H.; Tufarelli, C.; O’Sullivan, S.E. The endocannabinoid anandamide causes endothelium-dependent vasorelaxation in human mesenteric arteries. Pharmacol. Res. 2016, 113, 356–363. [Google Scholar] [CrossRef]
- Suleimani, Y.M.; Al Mahruqi, A.S. The endogenous lipid N-arachidonoyl glycine is hypotensive and nitric oxide-cGMP-dependent vasorelaxant. Eur. J. Pharmacol. 2017, 794, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Zong, Y.; Zhou, X.; Cheng, J.; Yu, J.; Wu, J.; Jiang, C. Cannabinoids Regulate the Diameter of Pericyte-Containing Retinal Capillaries in Rats. Cell. Physiol. Biochem. 2017, 43, 2088–2101. [Google Scholar] [CrossRef]
- López-Dyck, E.; Andrade-Urzúa, F.; Elizalde, A.; Ferrer-Villada, T.; Dagnino-Acosta, A.; Huerta, M.; Osuna-Calleros, Z.; Rangel-Sandoval, C.; Sánchez-Pastor, E. ACPA and JWH-133 modulate the vascular tone of superior mesenteric arteries through cannabinoid receptors, BKCa channels, and nitric oxide dependent mechanisms. Pharmacol. Rep. 2017, 69, 1131–1139. [Google Scholar] [CrossRef]
- Baranowska-Kuczko, M.; Kozłowska, H.; Kloza, M.; Harasim-Symbor, E.; Biernacki, M.; Kasacka, I.; Malinowska, B. Beneficial Changes in Rat Vascular Endocannabinoid System in Primary Hypertension and under Treatment with Chronic Inhibition of Fatty Acid Amide Hydrolase by URB597. Int. J. Mol. Sci. 2021, 22, 4833. [Google Scholar] [CrossRef]
- Sánchez-Pastor, E.; Andrade, F.; Sánchez-Pastor, J.M.; Elizalde, A.; Huerta, M.; Virgen-Ortiz, A.; Trujillo, X.; Rodríguez-Hernández, A. Cannabinoid receptor type 1 activation by arachidonylcyclopropylamide in rat aortic rings causes vasorelaxation involving calcium-activated potassium channel subunit alpha-1 and calcium channel, voltage-dependent, L type, alpha 1C subunit. Eur. J. Pharmacol. 2014, 729, 100–106. [Google Scholar] [CrossRef]
- MacIntyre, J.; Dong, A.; Straiker, A.; Zhu, J.; Howlett, S.E.; Bagher, A.; Denovan-Wright, E.; Yu, D.Y.; Kelly, M.E. Cannabinoid and lipid-mediated vasorelaxation in retinal microvasculature. Eur. J. Pharmacol. 2014, 735, 105–114. [Google Scholar] [CrossRef]
- Wheal, A.J.; Cipriano, M.; Fowler, C.J.; Randall, M.D.; O’Sullivan, S.E. Cannabidiol improves vasorelaxation in Zucker diabetic fatty rats through cyclooxygenase activation. J. Pharmacol. Exp. Ther. 2014, 351, 457–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheal, A.J.; Jadoon, K.; Randall, M.D.; O’Sullivan, S.E. In Vivo Cannabidiol Treatment Improves Endothelium-Dependent Vasorelaxation in Mesenteric Arteries of Zucker Diabetic Fatty Rats. Front. Pharmacol. 2017, 8, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baranowska-Kuczko, M.; Kozłowska, H.; Kloza, M.; Karpińska, O.; Toczek, M.; Harasim, E.; Kasacka, I.; Malinowska, B. Protective role of cannabinoid CB1 receptors and vascular effects of chronic administration of FAAH inhibitor URB597 in DOCA-salt hypertensive rats. Life Sci. 2016, 151, 288–299. [Google Scholar] [CrossRef] [PubMed]
- Karpińska, O.; Baranowska-Kuczko, M.; Kloza, M.; Kozłowska, H. Endocannabinoids modulate Gq/11 protein-coupled receptor agonist-induced vasoconstriction via a negative feedback mechanism. J. Pharm. Pharmacol. 2018, 70, 214–222. [Google Scholar] [CrossRef] [Green Version]
- Clark, A.; Tawhai, M. Pulmonary Vascular Dynamics. Compr. Physiol. 2019, 9, 1081–1100. [Google Scholar] [CrossRef] [PubMed]
- Mark Evans, A.; Ward, J.P. Hypoxic pulmonary vasoconstriction—Invited article. Adv. Exp. Med. Biol. 2009, 648, 351–360. [Google Scholar] [CrossRef]
- Suresh, K.; Shimoda, L.A. Lung Circulation. Compr. Physiol. 2016, 6, 897–943. [Google Scholar] [CrossRef] [PubMed]
- Su, J.Y.; Vo, A.C. 2-Arachidonylglyceryl ether and abnormal cannabidiol-induced vascular smooth muscle relaxation in rabbit pulmonary arteries via receptor-pertussis toxin sensitive G proteins-ERK1/2 signaling. Eur. J. Pharmacol. 2007, 559, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Kozłowska, H.; Baranowska, M.; Schlicker, E.; Kozłowski, M.; Laudański, J.; Malinowska, B. Virodhamine relaxes the human pulmonary artery through the endothelial cannabinoid receptor and indirectly through a COX product. Br. J. Pharmacol. 2008, 155, 1034–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baranowska-Kuczko, M.; MacLean, M.R.; Kozłowska, H.; Malinowska, B. Endothelium-dependent mechanisms of the vasodilatory effect of the endocannabinoid, anandamide, in the rat pulmonary artery. Pharmacol. Res. 2012, 66, 251–259. [Google Scholar] [CrossRef]
- Hopps, J.J.; Dunn, W.R.; Randall, M.D. Enhanced vasorelaxant effects of the endocannabinoid-like mediator, oleamide, in hypertension. Eur. J. Pharmacol. 2012, 684, 102–107. [Google Scholar] [CrossRef] [Green Version]
- Milman, G.; Maor, Y.; Abu-Lafi, S.; Horowitz, M.; Gallily, R.; Batkai, S.; Mo, F.M.; Offertaler, L.; Pacher, P.; Kunos, G. N-arachidonoyl L-serine, an endocannabinoid-like brain constituent with vasodilatory properties. Proc. Natl. Acad. Sci. USA 2006, 103, 2428–2433. [Google Scholar] [CrossRef] [Green Version]
- Ren, S.Y.; Wang, Z.Z.; Zhang, Y.; Chen, N.H. Potential application of endocannabinoid system agents in neuropsychiatric and neurodegenerative diseases-focusing on FAAH/MAGL inhibitors. Acta Pharmacol. Sin. 2020, 41, 1263–1271. [Google Scholar] [CrossRef]
- Kozak, K.R.; Marnett, L.J. Oxidative metabolism of endocannabinoids. Prostagland. Leukot. Essent. Fat. Acids 2002, 66, 211–220. [Google Scholar] [CrossRef]
- Herradón, E.; Martín, M.I.; López-Miranda, V. Characterization of the vasorelaxant mechanisms of the endocannabinoid anandamide in rat aorta. Br. J. Pharmacol. 2007, 152, 699–708. [Google Scholar] [CrossRef] [Green Version]
- Vachiery, J.L.; Huez, S.; Gillies, H.; Layton, G.; Hayashi, N.; Gao, X.; Naeije, R. Safety, tolerability and pharmacokinetics of an intravenous bolus of sildenafil in patients with pulmonary arterial hypertension. Br. J. Pharmacol. 2011, 71, 289–292. [Google Scholar] [CrossRef] [Green Version]
- Merkus, D.; de Beer, V.J.; Houweling, B.; Duncker, D.J. Control of pulmonary vascular tone during exercise in health and pulmonary hypertension. Pharmacol. Ther. 2008, 119, 242–263. [Google Scholar] [CrossRef] [PubMed]
- Kloza, M.; Baranowska-Kuczko, M.; Toczek, M.; Kusaczuk, M.; Sadowska, O.; Kasacka, I.; Kozłowska, H. Modulation of Cardiovascular Function in Primary Hypertension in Rat by SKA-31, an Activator of KCa2.x and KCa3.1 Channels. Int. J. Mol. Sci. 2019, 20, 4118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murtaza, G.; Mermer, P.; Goldenberg, A.; Pfeil, U.; Paddenberg, R.; Weissmann, N.; Lochnit, G.; Kummer, W. TASK-1 potassium channel is not critically involved in mediating hypoxic pulmonary vasoconstriction of murine intra-pulmonary arteries. PLoS ONE 2017, 12, e0174071. [Google Scholar] [CrossRef] [PubMed]
- Hwang, M.H.; Lim, J.H.; Yun, H.I.; Rhee, M.H.; Cho, J.Y.; Hsu, W.H.; Park, S.C. Surfactin C inhibits the lipopolysaccharide-induced transcription of interleukin-1beta and inducible nitric oxide synthase and nitric oxide production in murine RAW 264.7 cells. Biotechnol. Lett. 2005, 27, 1605–1608. [Google Scholar] [CrossRef] [PubMed]
- Zoratti, C.; Kipmen-Korgun, D.; Osibow, K.; Malli, R.; Graier, W.F. Anandamide initiates Ca(2+) signaling via CB2 receptor linked to phospholipase C in calf pulmonary endothelial cells. Br. J. Pharmacol. 2003, 140, 1351–1362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bondarenko, A.I. Cannabinoids and Cardiovascular System. Adv. Exp. Med. Biol. 2019, 1162, 63–87. [Google Scholar] [CrossRef]
- Parmar, N.; Ho, W.S. N-arachidonoyl glycine, an endogenous lipid that acts as a vasorelaxant via nitric oxide and large conductance calcium-activated potassium channels. Br. J. Pharmacol. 2010, 160, 594–603. [Google Scholar] [CrossRef] [Green Version]
- Bondarenko, A.I.; Drachuk, K.; Panasiuk, O.; Sagach, V.; Deak, A.T.; Malli, R. N-arachidonoyl glycine suppresses Na+/Ca2+ exchanger-mediated Ca2+ entry into endothelial cells and activates BK channels independently of G-protein coupled receptors. Br. J. Pharmacol. 2013, 169, 933–948. [Google Scholar] [CrossRef] [Green Version]
- Godlewski, G.; Offertáler, L.; Osei-Hyiaman, D.; Mo, F.M.; Harvey-White, J.; Liu, J.; Davis, M.I.; Zhang, L.; Razdan, R.K.; Milman, G.; et al. The endogenous brain constituent N-arachidonoyl L-serine is an activator of large conductance Ca2+-activated K+ channels. J. Pharmacol. Exp. Ther. 2009, 328, 351–361. [Google Scholar] [CrossRef]
- McHugh, D.; Page, J.; Dunn, E.; Bradshaw, H.B. Δ(9)-Tetrahydrocannabinol and N-arachidonyl glycine are full agonists at GPR18 receptors and induce migration in human endometrial HEC-1B cells. Br. J. Pharmacol. 2012, 165, 2414–2424. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Ye, L.; Huang, Y.; Ding, X.; Wang, L. The potential role of TRPV1 in pulmonary hypertension: Angel or demon? Channels 2019, 13, 235–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Gu, Q.; Qu, C. Capsaicin pretreatment reversed pulmonary arterial hypertension by alleviating inflammation via p38MAPK pathway. Exp. Lung Res. 2017, 43, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Addison, M.P.; Singh, T.U.; Parida, S.; Choudhury, S.; Kasa, J.K.; Sukumaran, S.V.; Darzi, S.A.; Kandasamy, K.; Singh, V.; Kumar, D.; et al. NO synthase inhibition attenuates EDHF-mediated relaxation induced by TRPV4 channel agonist GSK1016790A in the rat pulmonary artery: Role of TxA2. Pharmacol. Rep. 2016, 68, 620–626. [Google Scholar] [CrossRef] [PubMed]
- Dryn, D.; Melnyk, M.; Kizub, I.; Hu, H.; Soloviev, A.; Zholos, A. The Role of TRPV4 Cation Channels in the Regulation of Phenylephrine-Induced Contraction of Rat Pulmonary Arter. Fiziol. Zh. 2016, 62, 79–86. [Google Scholar] [CrossRef]
- Randhawa, P.K.; Jaggi, A.S. TRPV4 channels: Physiological and pathological role in cardiovascular system. Basic Res. Cardiol. 2015, 110, 54. [Google Scholar] [CrossRef] [PubMed]
- Kozłowska, H.; Baranowska-Kuczko, M.; Schlicker, E.; Kozłowski, M.; Kloza, M.; Malinowska, B. Relaxation of human pulmonary arteries by PPARγ agonists. Naunyn Schmiedebergs Arch. Pharmacol. 2013, 386, 445–453. [Google Scholar] [CrossRef] [Green Version]
- Hansmann, G.; Wagner, R.A.; Schellong, S.; Perez, V.A.; Urashima, T.; Wang, L.; Sheikh, A.Y.; Suen, R.S.; Stewart, D.J.; Rabinovitch, M. Pulmonary arterial hypertension is linked to insulin resistance and reversed by peroxisome proliferator-activated receptor-gamma activation. Circulation 2007, 115, 1275–1284. [Google Scholar] [CrossRef]
- Guignabert, C.; Alvira, C.M.; Alastalo, T.P.; Sawada, H.; Hansmann, G.; Zhao, M.; Wang, L.; El-Bizri, N.; Rabinovitch, M. Tie2-mediated loss of peroxisome proliferator-activated receptor-gamma in mice causes PDGF receptor-beta-dependent pulmonary arterial muscularization. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 297, L1082–L1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabinovitch, M. PPARgamma and the pathobiology of pulmonary arterial hypertension. Adv. Exp. Med. Biol. 2010, 661, 447–458. [Google Scholar] [CrossRef]
- Ameshima, S.; Golpon, H.; Cool, C.D.; Chan, D.; Vandivier, R.W.; Gardai, S.J.; Wick, M.; Nemenoff, R.A.; Geraci, M.W.; Voelkel, N.F. Peroxisome proliferator-activated receptor gamma (PPARgamma) expression is decreased in pulmonary hypertension and affects endothelial cell growth. Circ. Res. 2003, 92, 1162–1169. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Murphy, T.C.; Nanes, M.S.; Hart, C.M. PPAR{gamma} regulates hypoxia-induced Nox4 expression in human pulmonary artery smooth muscle cells through NF-{kappa}B. Am. J. Physiol. Lung Cell. Mol. Physiol. 2010, 299, L559–L566. [Google Scholar] [CrossRef] [Green Version]
- Nisbet, R.E.; Bland, J.M.; Kleinhenz, D.J.; Mitchell, P.O.; Walp, E.R.; Sutliff, R.L.; Hart, C.M. Rosiglitazone attenuates chronic hypoxia-induced pulmonary hypertension in a mouse model. Am. J. Respir. Cell Mol. Biol. 2010, 42, 482–490. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, S.E.; Sun, Y.; Bennett, A.J.; Randall, M.D.; Kendall, D.A. Time-dependent vascular actions of cannabidiol in the rat aorta. Eur. J. Pharmacol. 2009, 612, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.Y.; Shih, H.J.; Huang, I.T.; Tsai, P.S.; Chen, K.J.; Huang, H.J. Magnesium sulfate mitigates the progression of monocrotaline pulmonary hypertension in rats. Int. J. Mol. Sci. 2019, 20, 4622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, N.S.; Gillespie, M.N.; McMurtry, I.F. Fifty years of monocrotaline-induced pulmonary hypertension: What has it meant to the field? Chest 2017, 152, 1106–1108. [Google Scholar] [CrossRef] [Green Version]
- Remiszewski, P.; Jarocka-Karpowicz, I.; Biernacki, M.; Jastrząb, A.; Schlicker, E.; Toczek, M.; Harasim-Symbor, E.; Pędzińska-Betiuk, A.; Malinowska, B. Chronic cannabidiol administration fails to diminish blood pressure in rats with primary and secondary hypertension despite its effects on cardiac and plasma endocannabinoid system, oxidative stress and lipid metabolism. Int. J. Mol. Sci. 2020, 21, 1295. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Zhang, J.; Liu, H.; Ma, W.; Yu, L.; Tan, X.; Wang, S.; Ren, F.; Li, X.; Li, X. Cannabidiol attenuates pulmonary arterial hypertension by improving vascular smooth muscle cells mitochondrial function. Theranostics 2021, 11, 5267–5278. [Google Scholar] [CrossRef]
- Duerr, G.D.; Feißt, A.; Halbach, K.; Verfuerth, L.; Gestrich, C.; Wenzel, D.; Zimmer, A.; Breuer, J.; Dewald, O. CB2-deficiency is associated with a stronger hypertrophy and remodeling of the right ventricle in a murine model of left pulmonary artery occlusion. Life Sci. 2018, 215, 96–105. [Google Scholar] [CrossRef]



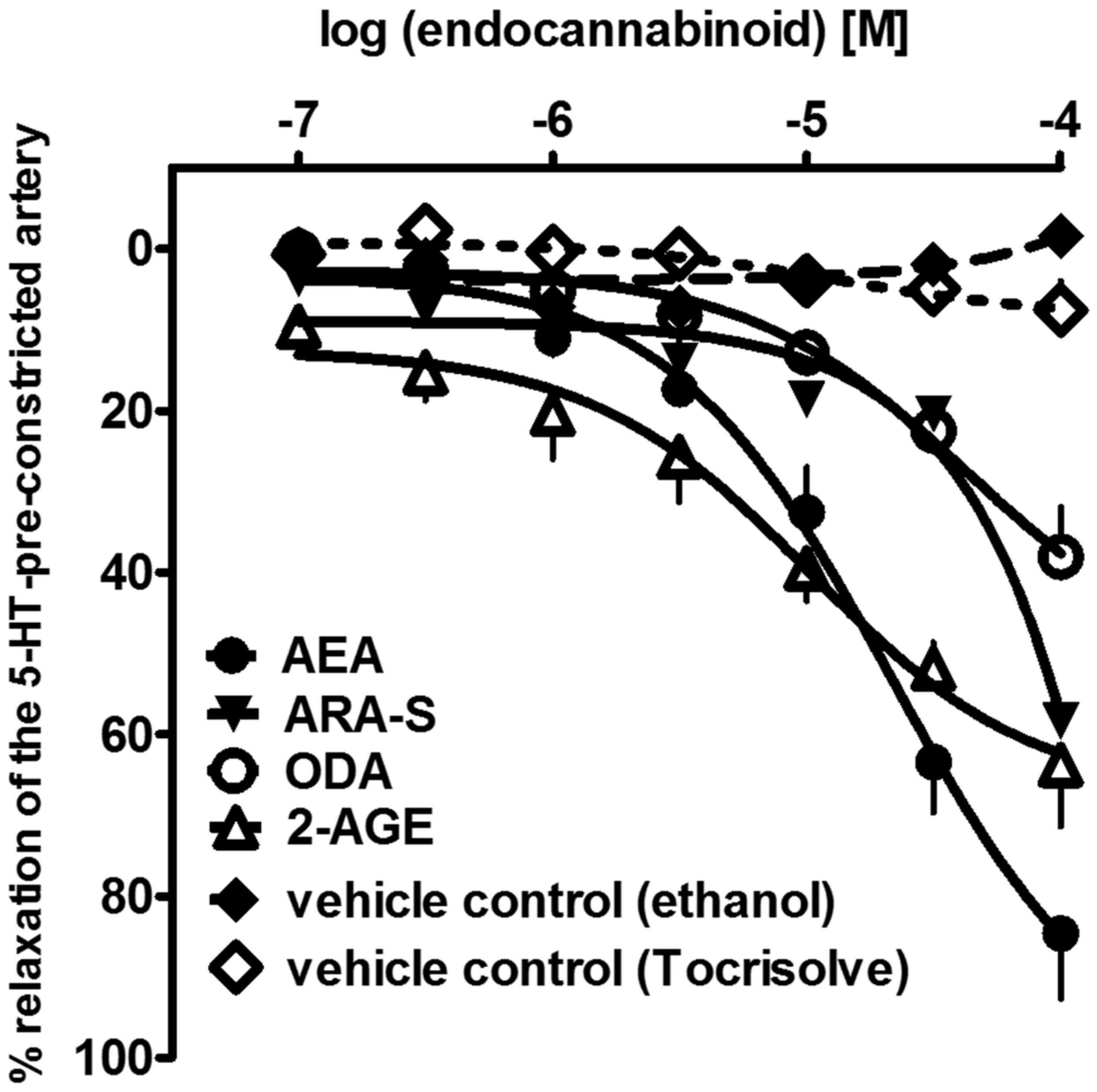{kind=link}
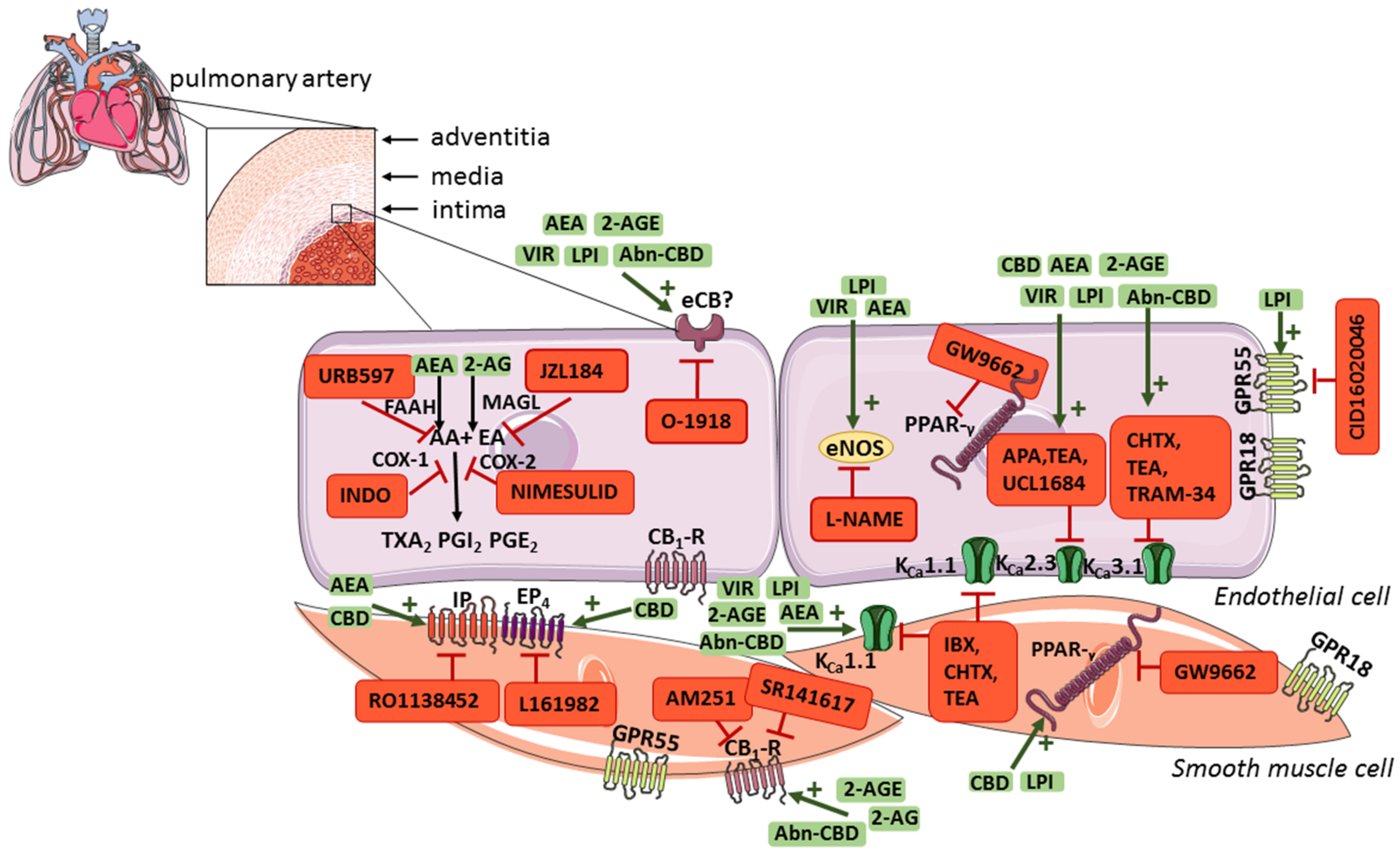{kind=link}
| Endocannabinoid System Components | Material | Species | Methods | Expression | References | |||
|---|---|---|---|---|---|---|---|---|
| Endothelium | Whole Vascular Wall | Whole Lung | ||||||
| ligands | 2-AG | lung cellular extracts | rabbit | LC/MS | + | [18] | ||
| lung | rat | + | [19] | |||||
| AEA | lung cellular extracts | rabbit | LC/MS | + | [18] | |||
| lung | rat | LC/MS | + | [19] | ||||
| mouse | LC/MRM | + | [20] | |||||
| receptors | CB1-R | pulmonary arteries | rat | IHC | + | [21] | ||
| WB | + | |||||||
| human | WB | + | ||||||
| IHC | + | |||||||
| + | [22] | |||||||
| CB2-R | pulmonary arteries | human | IHC | + | [22] | |||
| WB | + | [21] | ||||||
| rat | + | |||||||
| TRPV1 | pulmonary arteries | human | IHC | + | [22] | |||
| GPR18 | pulmonary arteries | human | IHC | + | [22] | |||
| GPR55 | pulmonary arteries | human | WB | + | [23] | |||
| IHC | + | |||||||
| enzymes | FAAH | pulmonary arteries | human | WB | + | [24] | ||
| lung | human | WB | + | |||||
| mouse | + | [20] | ||||||
| rabbit | RT-PCR | + | [18] | |||||
| Ligand | Blood Vessel | pEC50 | Mechanisms | References | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Endo | eNOS | COX | KCa | CB1-R | CB2-R | eCB | Other | ||||
| AEA | hMA | 5.7 | ↓ | ↓ | No | - | ↓ | No | ↓ | [38] | |
| rRet | 5.2 | - | - | - | - | - | - | - | [40] | ||
| 2-AG | rRet | 5.0 | - | - | - | - | - | - | - | [40] | |
| rMA | 5.9 * | ↓ | - | No | ↓ | No | No | - | TRPV4 | [36] | |
| 2-AGE | rMA | 5.6 * | No | - | - | - | - | - | - | [36] | |
| NAGLy | rMA | - | ↓ | ↓ | No | No | No | No | ↓ | [39] | |
| CBD | hMA | 5.1 | ↓ | ↓ | No | ↓ | ↓ | No | No | TRPV1 | [37] |
| rFA 1 | - | No | ↓ | ↓ | - | No | ↓ | No | SOD, EP4 | [45] | |
| rFA, rA 1 | - | - | ↓ | ↓ | - | - | - | - | [46] | ||
| rMA 1 | - | - | No | No | - | - | - | - | [46] | ||
| rMA 2 | 6.0 | No | - | - | - | No | No | - | [22] | ||
| rMA 3 | 5.5 | No | - | - | - | No | No | - | [22] | ||
| rMA 4 | 5.9 | ↓ | - | - | - | ↓ | ↓ | - | [22] | ||
| rMA 5 | 5.6 | No | - | - | - | ↓ | No | - | [22] | ||
| Abn-CBD | rRet | 4.5 | ↓ | No | - | ↓ | No | No | - | [44] | |
| pRet | - | ↓ | - | - | - | ↓ | - | ↓ | [35] | ||
| WIN 55,212-2 | rRet | 5.0 | ↓ | ↓ | No | - | ↓ | No | No | [40] | |
| JHW-133 | rMA | - | - | ↓ | - | - | - | ↓ | - | [41] | |
| MethAEA | rA 6 | 6.1 | - | - | - | - | - | - | - | [47] | |
| rMA 6 | 4.9 | - | - | - | - | No | - | - | TRPV1 | [47] | |
| rMA 4 | 5.6 | - | - | - | - | ↓ | - | - | TRPV1 | [47] | |
| rMA 5 | 5.6 | - | - | - | - | ↓ | - | - | [42] | ||
| rMA 2 | 6.1 | - | - | - | - | No | - | - | |||
| ACPA | rA | 4.3 | No | - | - | ↓ | No | - | - | Cav 1.2 | [43] |
| rMA | - | ↓ | ↓ | - | ↓ | ↓ | - | - | [41] | ||
| Species | Ligands | Vasoconstrictor | pEC50 | Concentration [μmol/L] | Endothelium | Inhibitors | KCa Inhibitors | Antagonists | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| eNOS | FAAH | COX-1 COX-2 | COX-2 | MAGL | KCl [60/120 mM] | KCa1.1 KCa3.1 | KCa1.1 | KCa2.3 | KCa3.1 | CB1-R | CB2-R | eCB | IP | EP4 | TRPV1 | PPAR-γ | References | ||||||
| L-NAME | URB597 | INDO | NIMES | JZL184 | CHTX | IBTX | UCL164/ APA * | TRAM-34 | AM251/ SR141716 * | AM630/ SR144528 * | O-1918 | RO1138452 | L161982 | CAPS | GW9662 | ||||||||
| human | AEA 1 | 5-HT | 5.2 | 0.1–100 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | [9] |
| AEA | U46619 | 5.0 | 0.1–100 | ↓ | ↓ | ↓ | ↓ | ↓ | - | ↓ | - | ↓ | - | - | No | No * | ↓ | ↓ | - | No | - | [24] | |
| VIR | 5-HT | 5.1 | 0.1–100 | ↓ | ↓3 | ↓ | ↓ | - | - | ↓ | ↓ | - | ↓ * | - | No | No * | ↓ 6.3 2 | - | - | No | - | [53] | |
| 2-AG | U46619 | 5.4 | 0.01–30 | ↓ | - | No | - | - | ↑ | - | - | - | - | - | ↓ 6.9 2 | - | - | - | - | - | - | [21] | |
| LPI | Phe | 6.4 | 0.01–3 | ↓ | ↓ | - | No | - | ↓ | - | ↓ | ↓ | ↓ | No | - | ↓ 5.8 2 | - | - | - | ↓ | [23] | ||
| CBD | U46619 | 5.0 | 0.1–30 | ↓ | No | - | ↓ | ↓ | - | ↓ | - | ↓ | ↓ | ↓ | No | No | No | ↓ 5.8 2 | ↓ 6.6 2 | ↓ | ↓ | [22] | |
| Abn-CBD | 5-HT | 4.8 | 0.1–100 | ↓ | No | - | No | - | - | ↓ | ↓ | - | ↓ * | - | - | - | ↓ 5.1 2 | - | - | - | - | [9] | |
| rabbit | 2-AGE | pCa 6.3 | - | 0.1–3 | ↓ | - | - | - | - | - | - | - | - | - | - | ↓/ ↓ * | - | ↓ | - | - | - | - | [52] |
| Abn-CBD | pCa 6.3 | - | 0.01–0.3 | ↓ | - | - | - | - | - | - | - | - | - | - | ↓/ ↓ * | - | ↓ | - | - | - | - | [52] | |
| rat | AEA | U46619 | 5.0 | 0.1–100 | ↓ | ↓ | ↓ | ↓ | - | - | ↓ | ↓ | - | ↓ * | - | No | No | ↓ 6.0 2 | ↓ 6.2 2 | - | No | - | [54] |
| Abn-CBD | U46619 | 4.6 | 0.1–100 | ↓ | - | - | - | - | - | ↓ | - | - | - | - | No | No | ↓ 5.4 2 | - | - | No | - | [54] | |
| Species | Model | Cannabinoid | Dose/Concentration/Route of Administration | Effect | References |
|---|---|---|---|---|---|
| rabbit | isolated, ventilated, and buffer-perfused lung | AEA | 0.5–5 μM | ↑ pulmonary arterial pressure | [18] |
| 2-AG | 0.2–0.4 μM | ||||
| rat | MCT-induced PH (60 mg/kg) | CBD | 10 mg/kg for 21 days, preventive model, i.p. | ↓ RVSP ↓ pulmonary arterial hypertrophy No right ventricular hypertrophy ↑ blood oxygen saturation ↑ concentration of endogenous cannabinoids in lung tissue: AEA, 2-LG, LEA, POEA, EPEA and NAGly ↓ the plasma concentrations of PAI-1 and t-PA | [19] |
| ↓ RVSP ↓ pulmonary arterial hypertrophy ↓ right ventricular hypertrophy | [87] | ||||
| mouse | sugen-hypoxia-induced PH | CBD | 10 mg/kg for 21 days, preventive model, i.g. | ↓ RVSP ↓ pulmonary arterial hypertrophy ↓ right ventricular hypertrophy ↓ mRNA levels of IL-6 and TNF-α in lung tissue | [87] |
| 10 mg/kg for 14 days after PH induction, therapeutic model i.g. | ↓ RVSP ↓ pulmonary arterial hypertrophy ↓ right ventricular hypertrophy | ||||
| PH-PASMC | 10 μM for 2 h | ↓ hyperproliferation ↓ mRNA levels of chemokine CCL2 and CXCL10 ↓ oxidative stress in mitochondria | |||
| human | hypoxia induced HPASMC cell culture | CBD | 10 μM for 2 h and 12 h | recover the dysfunctional mitochondria in hypoxia condition: ↓ oxidative stress ↓ excessive glycolysis | [87] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krzyżewska, A.; Baranowska-Kuczko, M.; Mińczuk, K.; Kozłowska, H. Cannabinoids—A New Perspective in Adjuvant Therapy for Pulmonary Hypertension. Int. J. Mol. Sci. 2021, 22, 10048. https://doi.org/10.3390/ijms221810048
Krzyżewska A, Baranowska-Kuczko M, Mińczuk K, Kozłowska H. Cannabinoids—A New Perspective in Adjuvant Therapy for Pulmonary Hypertension. International Journal of Molecular Sciences. 2021; 22(18):10048. https://doi.org/10.3390/ijms221810048
Chicago/Turabian StyleKrzyżewska, Anna, Marta Baranowska-Kuczko, Krzysztof Mińczuk, and Hanna Kozłowska. 2021. "Cannabinoids—A New Perspective in Adjuvant Therapy for Pulmonary Hypertension" International Journal of Molecular Sciences 22, no. 18: 10048. https://doi.org/10.3390/ijms221810048
APA StyleKrzyżewska, A., Baranowska-Kuczko, M., Mińczuk, K., & Kozłowska, H. (2021). Cannabinoids—A New Perspective in Adjuvant Therapy for Pulmonary Hypertension. International Journal of Molecular Sciences, 22(18), 10048. https://doi.org/10.3390/ijms221810048