
Glycolytic Reprogramming in Silica-Induced Lung Macrophages and Silicosis Reversed by Ac-SDKP Treatment

 , , and
, , and 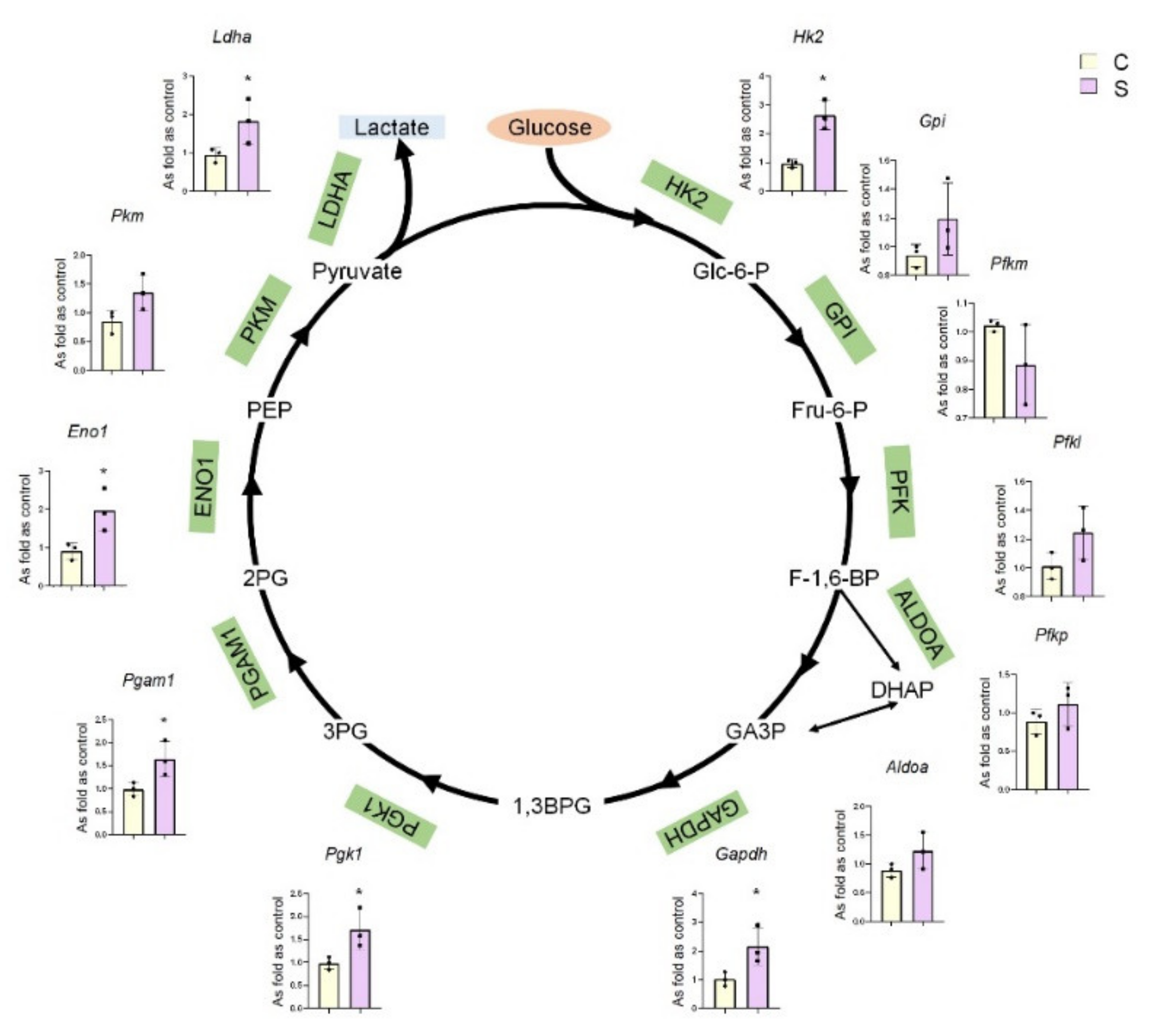{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Differential mRNA Expression Profiles of Key Glycolytic Enzymes in Silicotic Rat Lungs
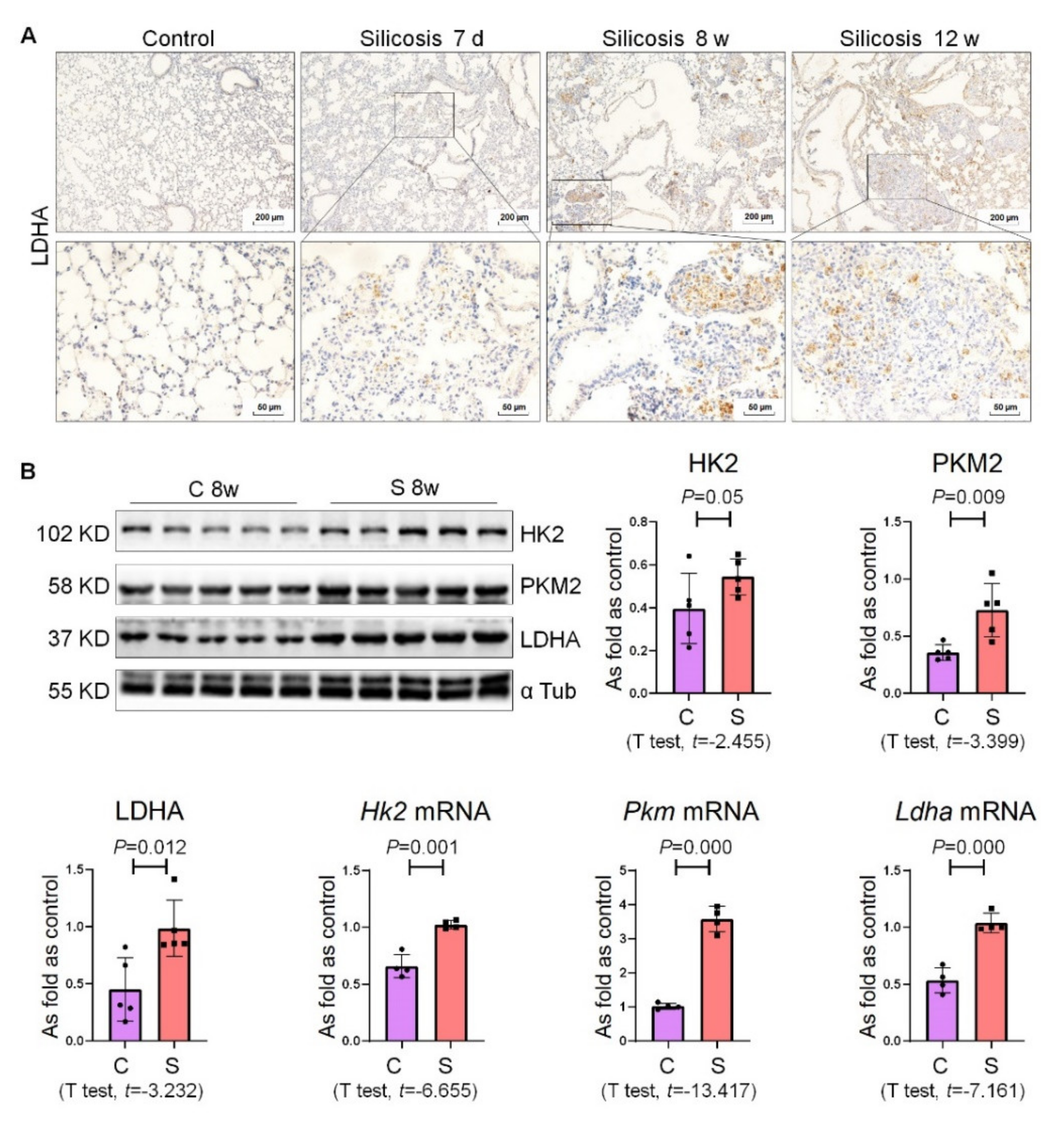
2.2. High Expression of Glycolysis in Rats Exposed to Silica
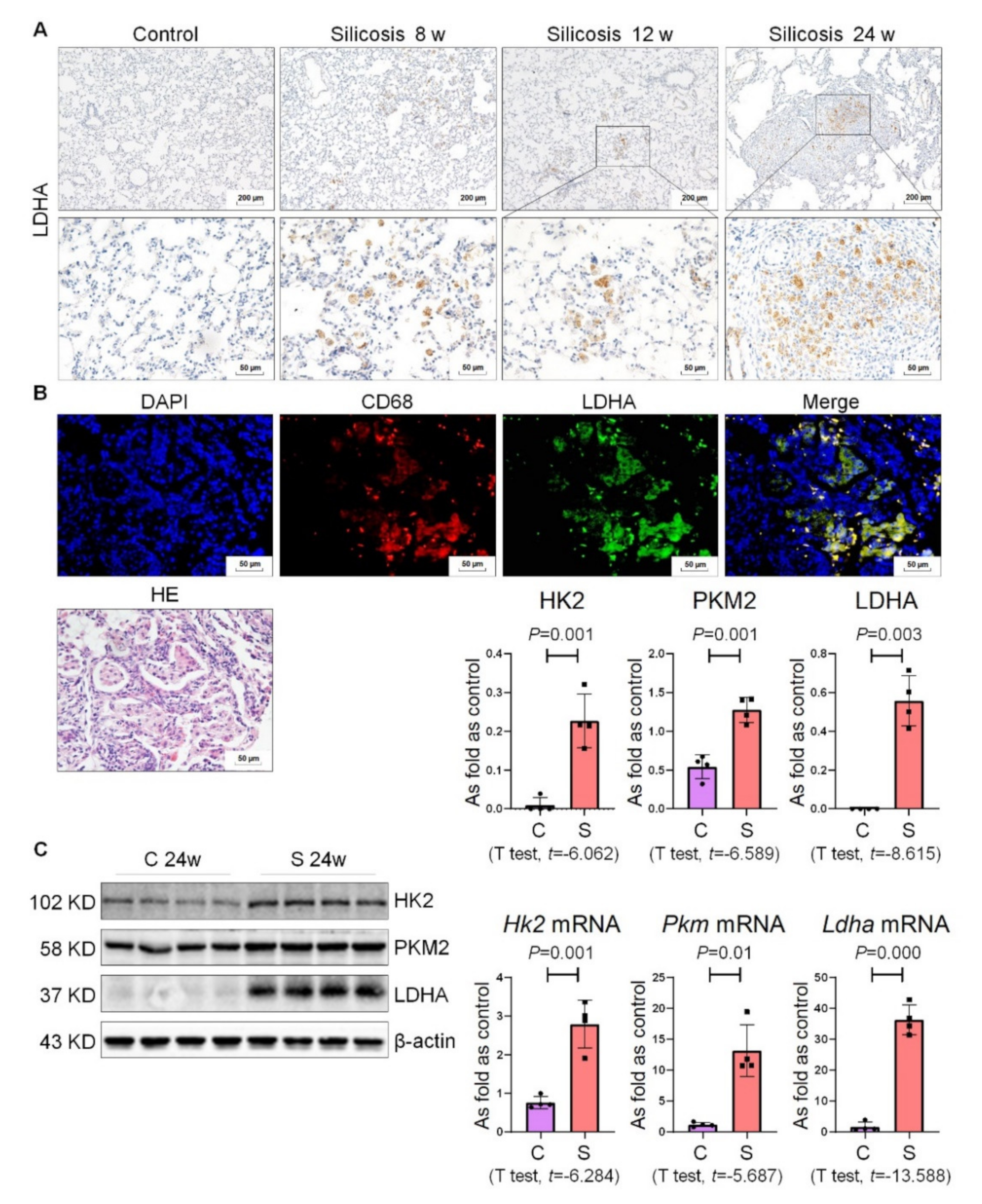
2.3. Enhanced Glycolysis Was the Crucial Metabolic Characteristic of Silicotic Mice
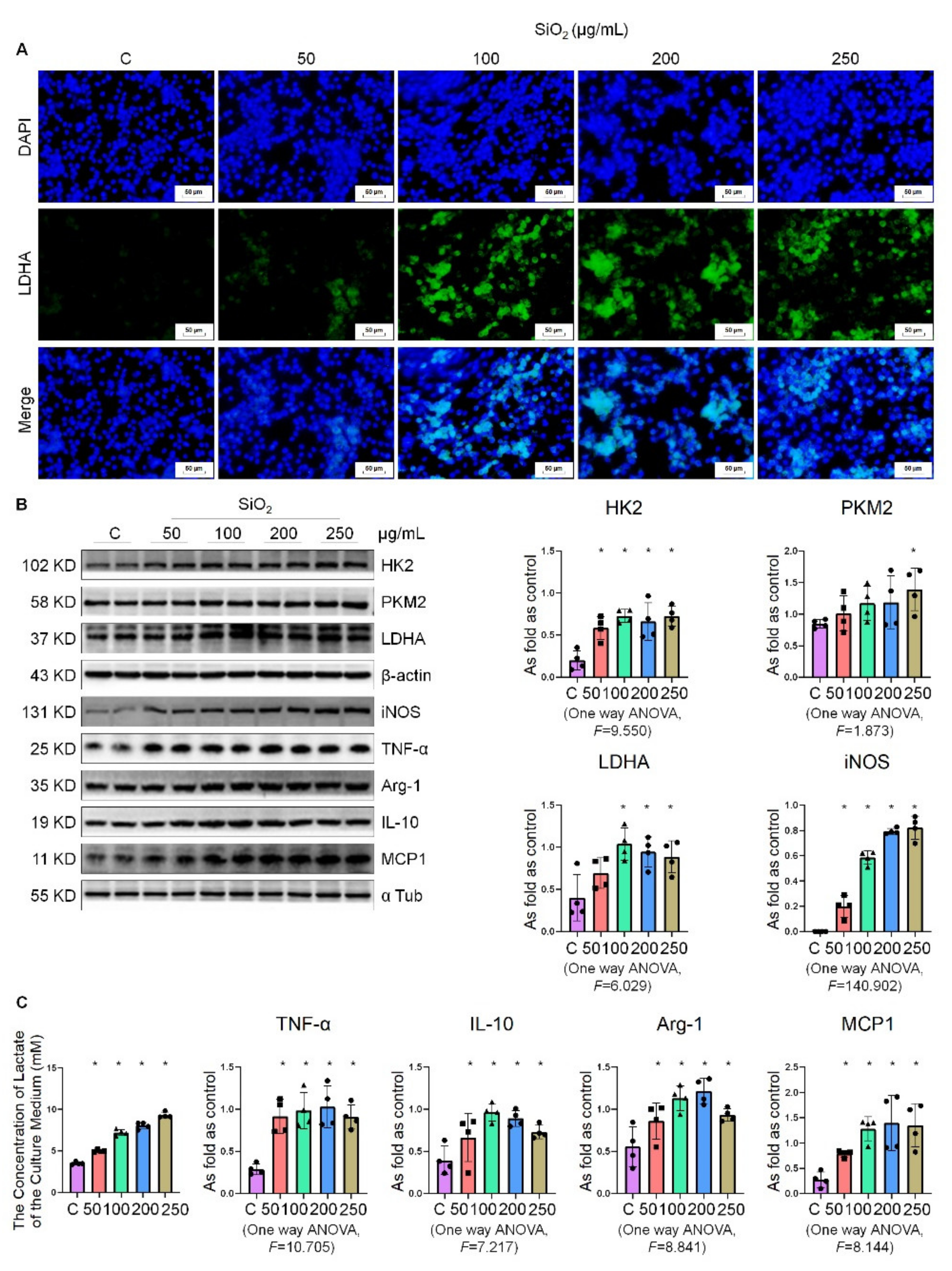
2.4. Silica Treatment Increased the Level of Glycolysis in Macrophages
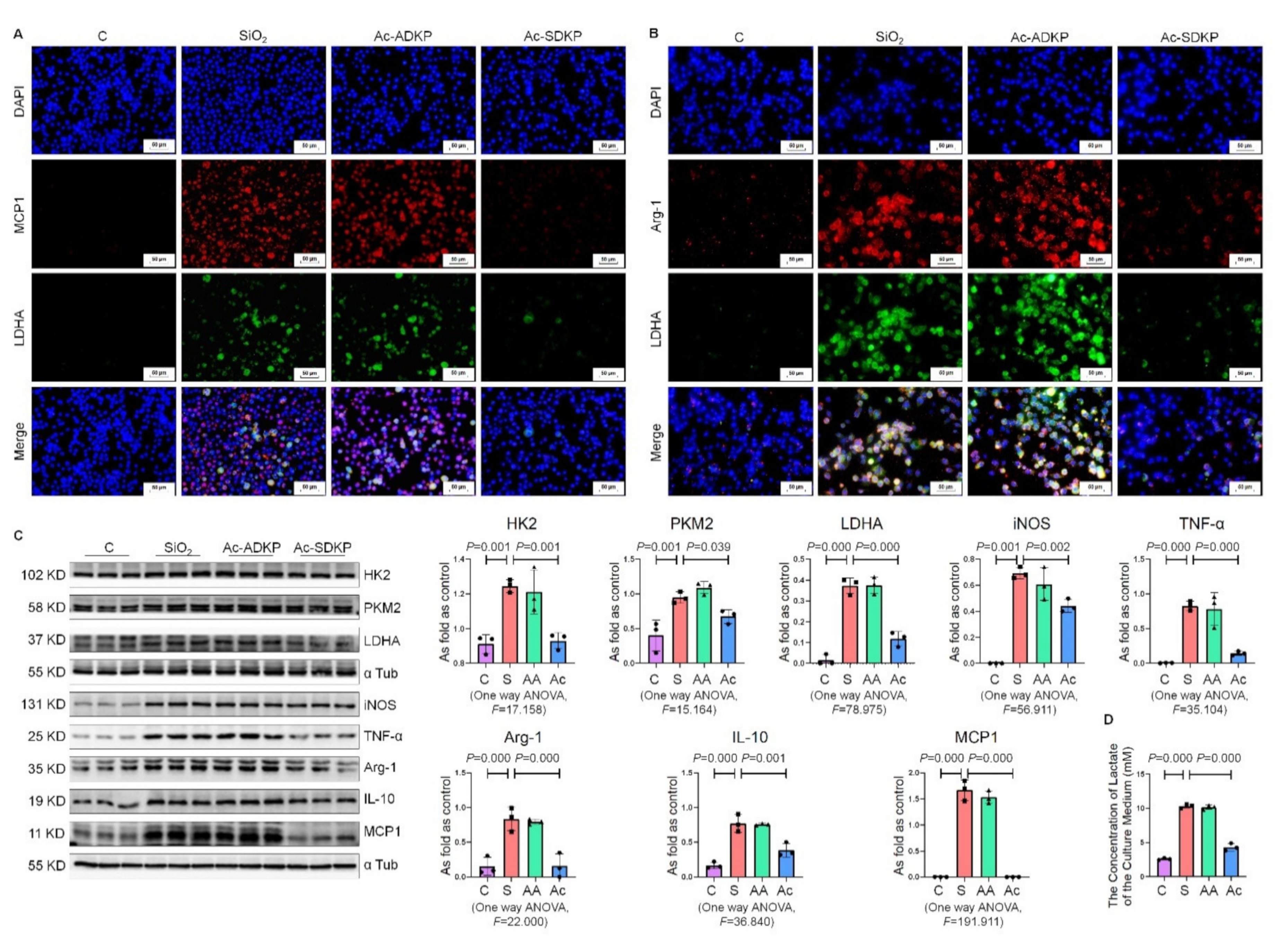
2.5. Ac-SDKP Attenuated the Enhancement of Glycolysis in Macrophages Treated with Silica
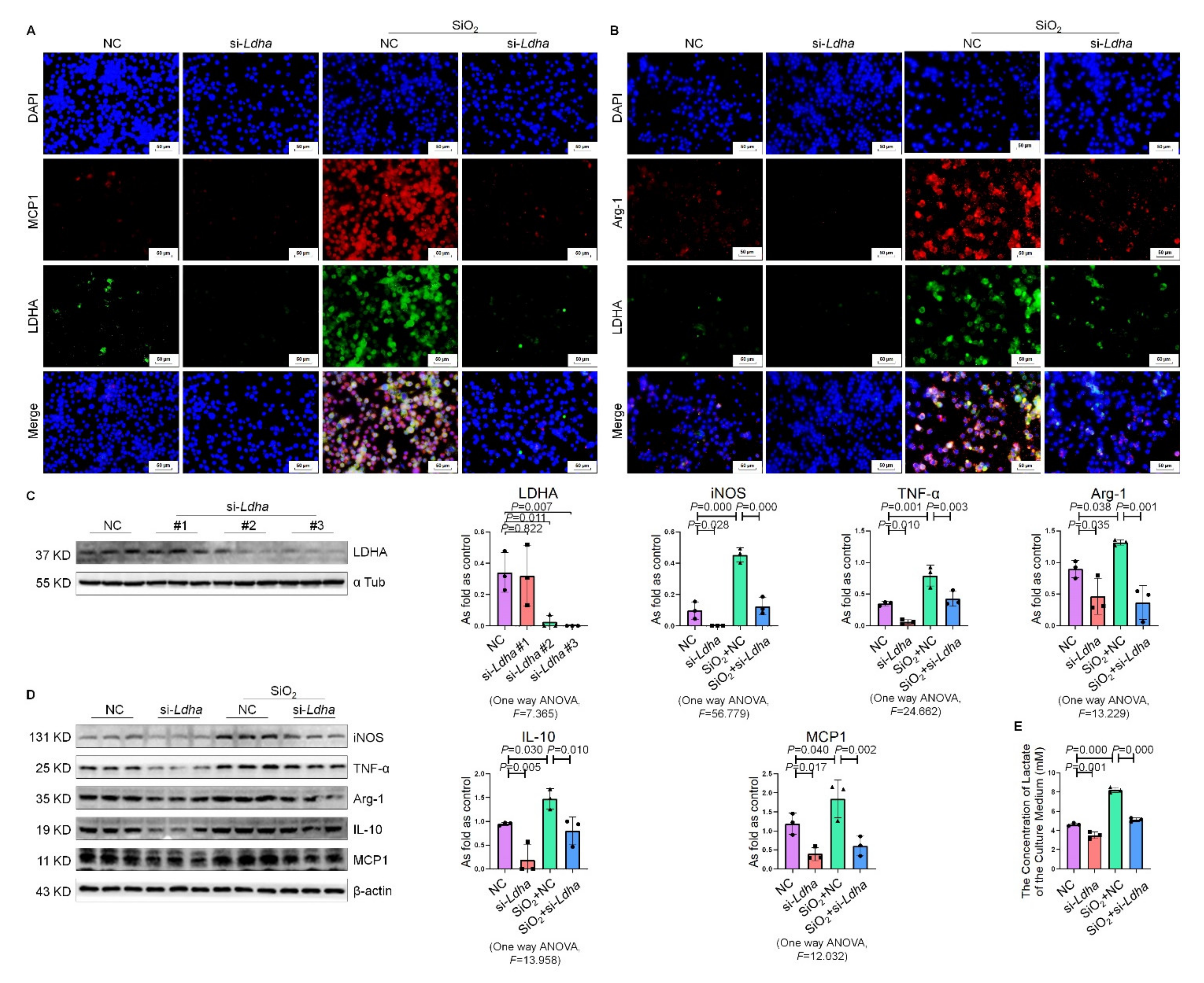
2.6. Knockdown of Ldha Inhibited Silica-Induced Macrophage Activation
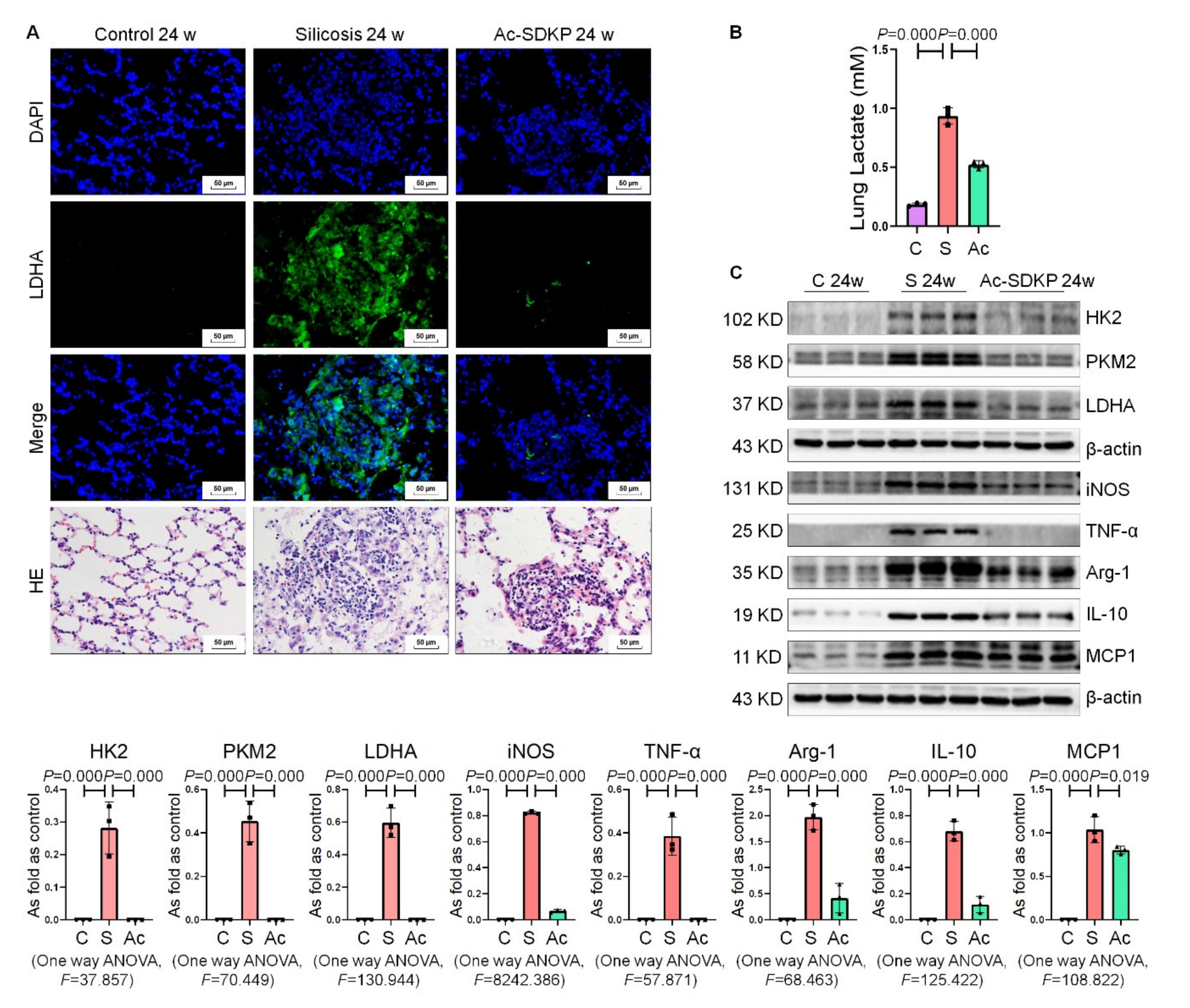
2.7. Ac-SDKP Attenuated High Glycolytic Activity and Macrophage Activation in Rats Exposed to Silica
3. Discussion
4. Materials and Methods
4.1. Animal Experiments
4.2. Cell Culture and Treatments
4.3. Cell Transfection
4.4. Immunohistochemistry and Immunofluorescence Staining
4.5. Western Blotting
4.6. Quantitative Real-Time PCR (qRT-PCR)
4.7. Lactate Production Measurement
4.8. Detection of ROS
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gao, X.; Xu, D.; Li, S.; Wei, Z.; Li, S.; Cai, W.; Mao, N.; Jin, F.; Li, Y.; Yi, X.; et al. Pulmonary Silicosis Alters MicroRNA Expression in Rat Lung and miR-411-3p Exerts Anti-fibrotic Effects by Inhibiting MRTF-A/SRF Signaling. Mol. Ther. Nucleic Acids 2020, 20, 851–865. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xu, D.; Yao, J.; Wei, Z.; Li, S.; Gao, X.; Cai, W.; Mao, N.; Jin, F.; Li, Y.; et al. Inhibition of miR-155-5p Exerts Anti-Fibrotic Effects in Silicotic Mice by Regulating Meprin alpha. Mol. Ther. Nucleic Acids 2020, 19, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Jin, F.; Geng, F.; Xu, D.; Li, Y.; Li, T.; Yang, X.; Liu, S.; Zhang, H.; Wei, Z.; Li, S.; et al. Ac-SDKP Attenuates Activation of Lung Macrophages and Bone Osteoclasts in Rats Exposed to Silica by Inhibition of TLR4 and RANKL Signaling Pathways. J. Inflamm. Res. 2021, 14, 1647–1660. [Google Scholar] [CrossRef]
- Soto-Heredero, G.; Gomez de Las Heras, M.M.; Gabande-Rodriguez, E.; Oller, J.; Mittelbrunn, M. Glycolysis—a key player in the inflammatory response. FEBS J. 2020, 287, 3350–3369. [Google Scholar] [CrossRef] [Green Version]
- Batista-Gonzalez, A.; Vidal, R.; Criollo, A.; Carreno, L.J. New Insights on the Role of Lipid Metabolism in the Metabolic Reprogramming of Macrophages. Front. Immunol. 2019, 10, 2993. [Google Scholar] [CrossRef]
- Xie, N.; Cui, H.; Ge, J.; Banerjee, S.; Guo, S.; Dubey, S.; Abraham, E.; Liu, R.M.; Liu, G. Metabolic characterization and RNA profiling reveal glycolytic dependence of profibrotic phenotype of alveolar macrophages in lung fibrosis. Am. J. Physiol. Lung Cell. Mol. Physiol. 2017, 313, L834–L844. [Google Scholar] [CrossRef] [PubMed]
- Xie, N.; Tan, Z.; Banerjee, S.; Cui, H.; Ge, J.; Liu, R.M.; Bernard, K.; Thannickal, V.J.; Liu, G. Glycolytic Reprogramming in Myofibroblast Differentiation and Lung Fibrosis. Am. J. Respir. Crit. Care Med. 2015, 192, 1462–1474. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Cheng, D.; Li, G.; Liu, Y.; Li, P.; Sun, W.; Ma, D.; Ni, C. CircHIPK3 regulates pulmonary fibrosis by facilitating glycolysis in miR-30a-3p/FOXK2-dependent manner. Int. J. Biol. Sci. 2021, 17, 2294–2307. [Google Scholar] [CrossRef]
- Yang, M.; Wang, D.; Gan, S.; Wang, B.; Yu, L.; Xie, Y.; Fan, L.; Ma, J.; Chen, W. Triiodothyronine ameliorates silica-induced pulmonary inflammation and fibrosis in mice. Sci. Total Environ. 2021, 790, 148041. [Google Scholar] [CrossRef]
- Cai, W.; Zhang, B.; Li, T.; Jin, F.; Li, Y.; Xu, H.; Yang, F. Transcriptomic analysis identifies upregulation of secreted phosphoprotein 1 in silicotic rats. Exp. Ther. Med. 2021, 21, 579. [Google Scholar] [CrossRef]
- Zhu, Y.; Yao, J.; Duan, Y.; Xu, H.; Cheng, Q.; Gao, X.; Li, S.; Yang, F.; Liu, H.; Yuan, J. Protein Expression Profile in Rat Silicosis Model Reveals Upregulation of PTPN2 and Its Inhibitory Effect on Epithelial-Mesenchymal Transition by Dephosphorylation of STAT3. Int. J. Mol. Sci. 2020, 21, 1189. [Google Scholar] [CrossRef] [Green Version]
- Kottmann, R.M.; Kulkarni, A.A.; Smolnycki, K.A.; Lyda, E.; Dahanayake, T.; Salibi, R.; Honnons, S.; Jones, C.; Isern, N.G.; Hu, J.Z.; et al. Lactic acid is elevated in idiopathic pulmonary fibrosis and induces myofibroblast differentiation via pH-dependent activation of transforming growth factor-β. Am. J. Respir. Crit. Care Med. 2012, 186, 740–751. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Xu, H.; Zhang, B.; Tao, T.; Liu, Y.; Xu, D.; Cai, W.; Wei, Z.; Li, S.; Zhang, H.; et al. Interaction of N-acetyl-seryl-aspartyl-lysyl-proline with the angiotensin-converting enzyme 2-angiotensin-(1-7)-Mas axis attenuates pulmonary fibrosis in silicotic rats. Exp. Physiol. 2019, 104, 1562–1574. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, Y.; Zhang, Y.; Li, S.; Zhang, M.; Jin, F.; Wei, Z.; Yang, Y.; Gao, X.; Mao, N.; et al. N-Acetyl-Seryl-Asparyl-Lysyl-Proline regulates lung renin angiotensin system to inhibit epithelial-mesenchymal transition in silicotic mice. Toxicol. Appl. Pharmacol. 2020, 408, 115255. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, D.; Li, Q.; Yang, Y.; Xu, H.; Wei, Z.; Wang, R.; Zhang, W.; Liu, Y.; Geng, Y.; et al. N-acetyl-seryl-aspartyl-lysyl-proline (Ac-SDKP) attenuates silicotic fibrosis by suppressing apoptosis of alveolar type II epithelial cells via mediation of endoplasmic reticulum stress. Toxicol. Appl. Pharmacol. 2018, 350, 1–10. [Google Scholar] [CrossRef]
- Shifeng, L.; Hong, X.; Xue, Y.; Siyu, N.; Qiaodan, Z.; Dingjie, X.; Lijuan, Z.; Zhongqiu, W.; Xuemin, G.; Wenchen, C.; et al. Ac-SDKP increases alpha-TAT 1 and promotes the apoptosis in lung fibroblasts and epithelial cells double-stimulated with TGF-beta1 and silica. Toxicol. Appl. Pharmacol. 2019, 369, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Yang, F.; Sun, Y.; Yuan, Y.; Cheng, H.; Wei, Z.; Li, S.; Cheng, T.; Brann, D.; Wang, R. A new antifibrotic target of Ac-SDKP: Inhibition of myofibroblast differentiation in rat lung with silicosis. PLoS ONE 2012, 7, e40301. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, S.P.; Goodwin, J.E.; Kanasaki, K.; Koya, D. Metabolic reprogramming by N-acetyl-seryl-aspartyl-lysyl-proline protects against diabetic kidney disease. Br. J. Pharmacol. 2020, 177, 3691–3711. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, J.; Dou, P.; Zhang, X.; Ran, X.; Liu, L.; Dou, D. The Ameliorative Effects of Arctiin and Arctigenin on the Oxidative Injury of Lung Induced by Silica via TLR-4/NLRP3/TGF-beta Signaling Pathway. Oxidative Med. Cell. Longev. 2021, 2021, 5598980. [Google Scholar] [CrossRef] [PubMed]
- Arra, M.; Swarnkar, G.; Ke, K.; Otero, J.E.; Ying, J.; Duan, X.; Maruyama, T.; Rai, M.F.; O’Keefe, R.J.; Mbalaviele, G.; et al. LDHA-mediated ROS generation in chondrocytes is a potential therapeutic target for osteoarthritis. Nat. Commun. 2020, 11, 3427. [Google Scholar] [CrossRef]
- Viola, A.; Munari, F.; Sanchez-Rodriguez, R.; Scolaro, T.; Castegna, A. The Metabolic Signature of Macrophage Responses. Front. Immunol. 2019, 10, 1462. [Google Scholar] [CrossRef] [Green Version]
- Woods, P.S.; Kimmig, L.M.; Meliton, A.Y.; Sun, K.A.; Tian, Y.; O’Leary, E.M.; Gokalp, G.A.; Hamanaka, R.B.; Mutlu, G.M. Tissue-Resident Alveolar Macrophages Do Not Rely on Glycolysis for LPS-induced Inflammation. Am. J. Respir. Cell Mol. Biol. 2020, 62, 243–255. [Google Scholar] [CrossRef]
- Khaing, P.; Summer, R. Maxed Out on Glycolysis: Alveolar Macrophages Rely on Oxidative Phosphorylation for Cytokine Production. Am. J. Respir. Cell Mol. Biol. 2020, 62, 139–140. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, R.; Gu, H.; Zhang, E.; Qu, J.; Cao, W.; Huang, X.; Yan, H.; He, J.; Cai, Z. Metabolic reprogramming in macrophage responses. Biomark. Res. 2021, 9, 1. [Google Scholar] [CrossRef]
- Judge, J.L.; Nagel, D.J.; Owens, K.M.; Rackow, A.; Phipps, R.P.; Sime, P.J.; Kottmann, R.M. Prevention and treatment of bleomycin-induced pulmonary fibrosis with the lactate dehydrogenase inhibitor gossypol. PLoS ONE 2018, 13, e0197936. [Google Scholar] [CrossRef]
- Judge, J.L.; Owens, K.M.; Pollock, S.J.; Woeller, C.F.; Thatcher, T.H.; Williams, J.P.; Phipps, R.P.; Sime, P.J.; Kottmann, R.M. Ionizing radiation induces myofibroblast differentiation via lactate dehydrogenase. Am. J. Physiol. Lung Cell. Mol. Physiol. 2015, 309, L879–L887. [Google Scholar] [CrossRef] [Green Version]
- Judge, J.L.; Lacy, S.H.; Ku, W.Y.; Owens, K.M.; Hernady, E.; Thatcher, T.H.; Williams, J.P.; Phipps, R.P.; Sime, P.J.; Kottmann, R.M. The Lactate Dehydrogenase Inhibitor Gossypol Inhibits Radiation-Induced Pulmonary Fibrosis. Radiat. Res. 2017, 188, 35–43. [Google Scholar] [CrossRef]
- Massip-Copiz, M.M.; Valdivieso, A.G.; Clauzure, M.; Mori, C.; Asensio, C.J.A.; Aguilar, M.A.; Santa-Coloma, T. EGFR activity upregulates lactate dehydrogenase A (LDHA) expression, LDH activity and lactate secretion in cultured IB3-1 cystic fibrosis lung epithelial cells. Biochem. Cell Biol. (Biochim. Et Biol. Cell.) 2021, 99, 476–487. [Google Scholar] [CrossRef]
- Cantero-Navarro, E.; Fernandez-Fernandez, B.; Ramos, A.M.; Rayego-Mateos, S.; Rodrigues-Diez, R.R.; Sanchez-Nino, M.D.; Sanz, A.B.; Ruiz-Ortega, M.; Ortiz, A. Renin-angiotensin system and inflammation update. Mol. Cell. Endocrinol. 2021, 529, 111254. [Google Scholar] [CrossRef]
- Ma, X.; Yuan, Y.; Zhang, Z.; Zhang, Y.; Li, M. An analog of Ac-SDKP improves heart functions after myocardial infarction by suppressing alternative activation (M2) of macrophages. Int. J. Cardiol. 2014, 175, 376–378. [Google Scholar] [CrossRef]
- Nakagawa, P.; Romero, C.A.; Jiang, X.; D’Ambrosio, M.; Bordcoch, G.; Peterson, E.L.; Harding, P.; Yang, X.-P.; Carretero, O.A. Ac-SDKP decreases mortality and cardiac rupture after acute myocardial infarction. PLoS ONE 2018, 13, e0190300. [Google Scholar] [CrossRef] [Green Version]
- Sharma, U.; Rhaleb, N.-E.; Pokharel, S.; Harding, P.; Rasoul, S.; Peng, H.; Carretero, O.A. Novel anti-inflammatory mechanisms of N-Acetyl-Ser-Asp-Lys-Pro in hypertension-induced target organ damage. Am. J. Physiol. Heart Circ. Physiol. 2008, 294, H1226–H1232. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wei, Z.; Li, G.; Zhang, Q.; Niu, S.; Xu, D.; Mao, N.; Chen, S.; Gao, X.; Cai, W.; et al. Silica Perturbs Primary Cilia and Causes Myofibroblast Differentiation during Silicosis by Reduction of the KIF3A-Repressor GLI3 Complex. Theranostics 2020, 10, 1719–1732. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.; Xu, H.; Zhang, B.; Gao, X.; Li, S.; Wei, Z.; Li, S.; Mao, N.; Jin, F.; Li, Y.; et al. Differential expression of lncRNAs during silicosis and the role of LOC103691771 in myofibroblast differentiation induced by TGF-beta1. Biomed. Pharmacother. 2020, 125, 109980. [Google Scholar] [CrossRef]
- Gao, X.; Xu, H.; Xu, D.; Li, S.; Wei, Z.; Li, S.; Cai, W.; Mao, N.; Jin, F.; Li, Y.; et al. MiR-411-3p alleviates Silica-induced pulmonary fibrosis by regulating Smurf2/TGF-beta signaling. Exp. Cell Res. 2020, 388, 111878. [Google Scholar] [CrossRef] [PubMed]
- Na, M.; Hong, X.; Fuyu, J.; Dingjie, X.; Sales, D.; Hui, Z.; Zhongqiu, W.; Shifeng, L.; Xuemin, G.; Wenchen, C.; et al. Proteomic profile of TGF-beta1 treated lung fibroblasts identifies novel markers of activated fibroblasts in the silica exposed rat lung. Exp. Cell Res. 2019, 375, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Xu, H.; Zhang, Y.; Yi, X.; Yang, X.; Chen, Y.; Mao, N.; Li, S.; Xu, D.; Li, S.; et al. Rho GDP dissociation inhibitor alpha silencing attenuates silicosis by inhibiting RhoA/Rho kinase signalling. Exp. Cell Res. 2019, 380, 131–140. [Google Scholar] [CrossRef]
- Hui, Z.; Dingjie, X.; Yuan, Y.; Zhongqiu, W.; Na, M.; Mingjian, B.; Yu, G.; Guangyuan, L.; Xuemin, G.; Shifeng, L.; et al. Silicosis decreases bone mineral density in rats. Toxicol. Appl. Pharmacol. 2018, 348, 117–122. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mao, N.; Yang, H.; Yin, J.; Li, Y.; Jin, F.; Li, T.; Yang, X.; Sun, Y.; Liu, H.; Xu, H.; et al. Glycolytic Reprogramming in Silica-Induced Lung Macrophages and Silicosis Reversed by Ac-SDKP Treatment. Int. J. Mol. Sci. 2021, 22, 10063. https://doi.org/10.3390/ijms221810063
Mao N, Yang H, Yin J, Li Y, Jin F, Li T, Yang X, Sun Y, Liu H, Xu H, et al. Glycolytic Reprogramming in Silica-Induced Lung Macrophages and Silicosis Reversed by Ac-SDKP Treatment. International Journal of Molecular Sciences. 2021; 22(18):10063. https://doi.org/10.3390/ijms221810063
Chicago/Turabian StyleMao, Na, Honghao Yang, Jie Yin, Yaqian Li, Fuyu Jin, Tian Li, Xinyu Yang, Ying Sun, Heliang Liu, Hong Xu, and et al. 2021. "Glycolytic Reprogramming in Silica-Induced Lung Macrophages and Silicosis Reversed by Ac-SDKP Treatment" International Journal of Molecular Sciences 22, no. 18: 10063. https://doi.org/10.3390/ijms221810063
APA StyleMao, N., Yang, H., Yin, J., Li, Y., Jin, F., Li, T., Yang, X., Sun, Y., Liu, H., Xu, H., & Yang, F. (2021). Glycolytic Reprogramming in Silica-Induced Lung Macrophages and Silicosis Reversed by Ac-SDKP Treatment. International Journal of Molecular Sciences, 22(18), 10063. https://doi.org/10.3390/ijms221810063