
Molecular Mechanism of Arsenic-Induced Neurotoxicity including Neuronal Dysfunctions

 ,
, 
 and
and 
Abstract
:1. Introduction
2. Means of Human Arsenic Exposure
2.1. Drinking Water
2.2. Diet
2.3. Industrialized and Wood Preservatives
2.4. Smoking
2.5. Air
2.6. Cosmetics
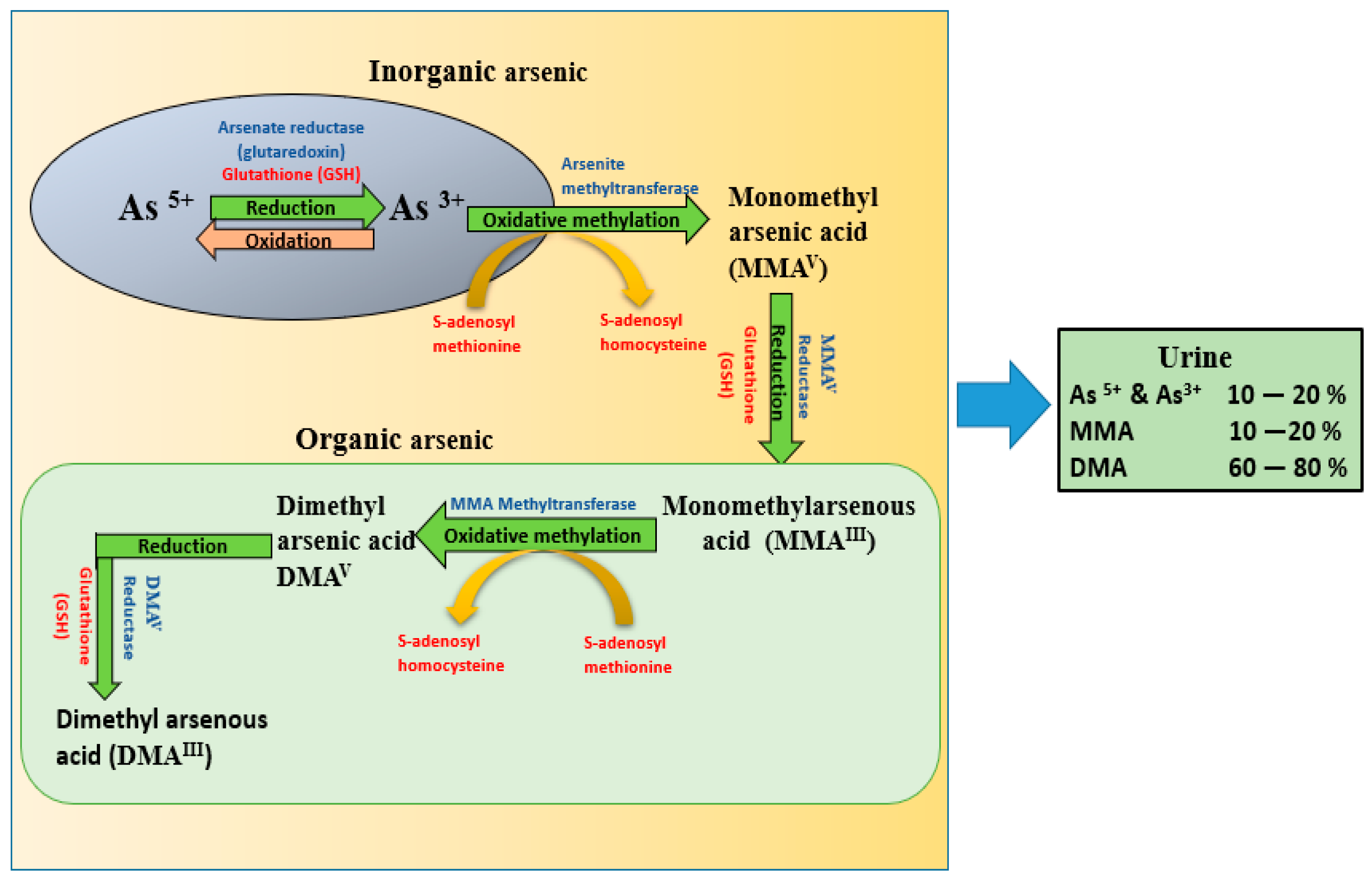
3. Metabolic Pathway of Arsenic
4. Neuronal Effects of Arsenic
4.1. Neurotransmitter Mediated Impacts of Arsenic
4.2. Neurodevelopmental Defects and the Effects of Aging
4.3. Neurobehavioral Effects of Aging in Animal Models
4.4. Neurobehavioral Effects of Aging in Humans
5. Toxicological Pathways
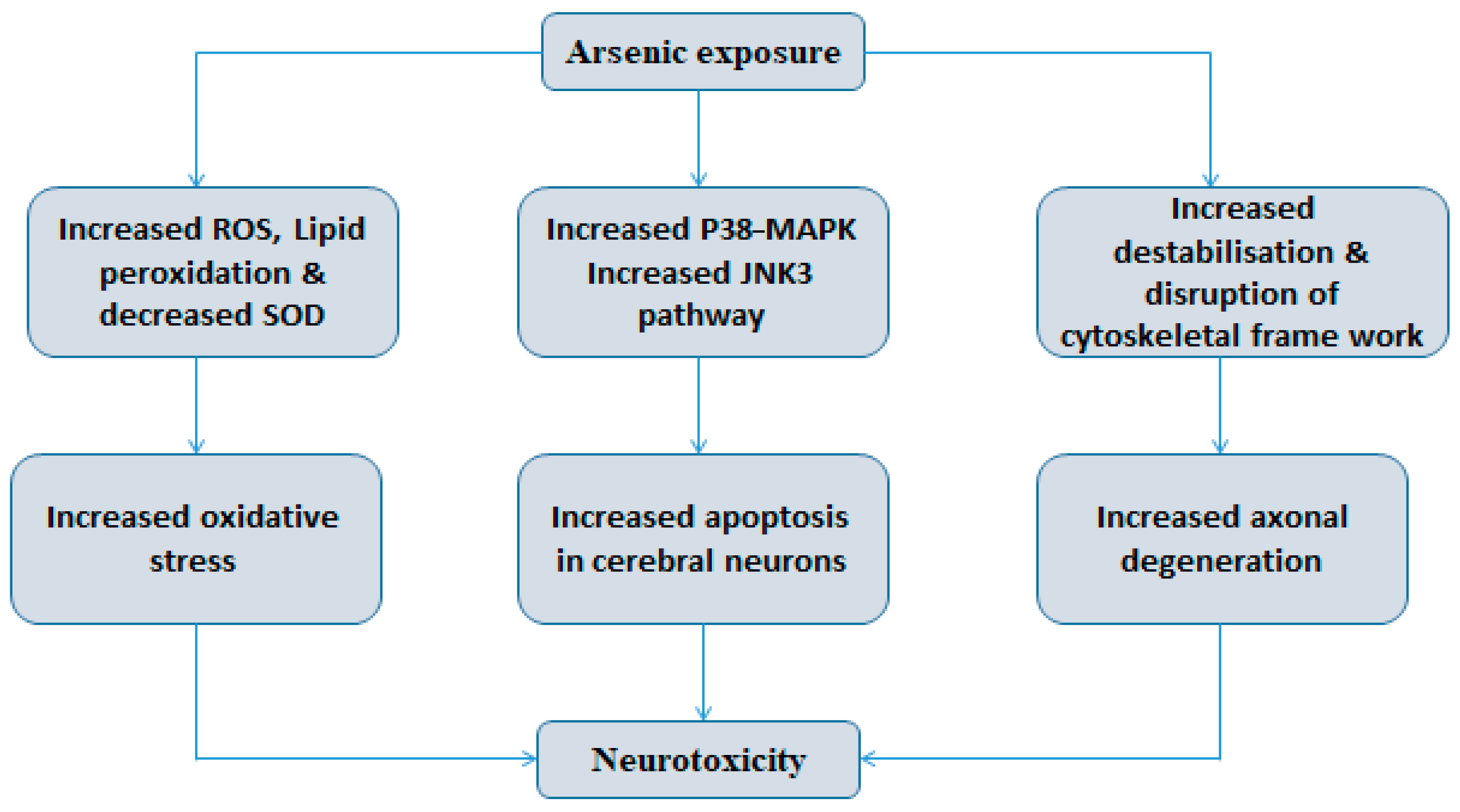
5.1. Molecular Mechanisms of Arsenic-Induced Oxidative Stress
- (i.)
- Arsenic is known to induce changes in mitochondria, the mitochondrial membrane integrity and reduce membrane potential. These morphological alterations are primary sites for the unregulated production of superoxide anion radicals causing a cascade of downstream processes resulting in the formation of free radicals. The further build-up of oxidative stress leads to failure of the oxidative defense system and results in toxic manifestations [150].
- (ii.)
- Mitochondrial complexes I and III produce O2− in the electron transport chain. Arsenic inhibits succinic dehydrogenase activity and promotes uncoupling of oxidative phosphorylation with the output of O2−, which leads to a buildup of oxidative stress [151].
- (iii.)
- Arsenic may also generate ROS through NAD(P)H oxidase assisted processes. NAD(P)H oxidase is a membrane-bound enzyme that produces superoxides by transferring electrons from NAD(P)H within the cell around the membrane and combining those to molecular oxygen to generate superoxide anions. It was demonstrated in mammalian endothelial cell culture that arsenic acts as an extracellular signal for the Ras proteins (cdc42), which activate NAD(P)H oxidase to generate ROS [96].
- (iv.)
- Arsenic can also generate ROS by affecting nitric oxide (NO) synthase enzyme system. Nitric oxide synthase iso-enzymes are coupled to produce NO from L-arginine and molecular oxygen without producing superoxides. Exposure to arsenic disrupts this coupling produces ROS [97].
- (v.)
- Metabolism of As(III) to As(V) in normal conditions results in the generation of H2O2 [98].
- (vi.)
- ROS are generated during the formation of intermediate arsine species such as dimethylarsenic peroxyl radicals-metabolic by-products of dimethylarsinic acid [99].
- (vii.)
- Methylated 3+ organic arsenicals react with sulfhydryl groups (-SH) in antioxidative proteins and inhibit their activity, which results in a build-up of oxidative stress [100].
5.2. Mitochondrial Dysfunctions
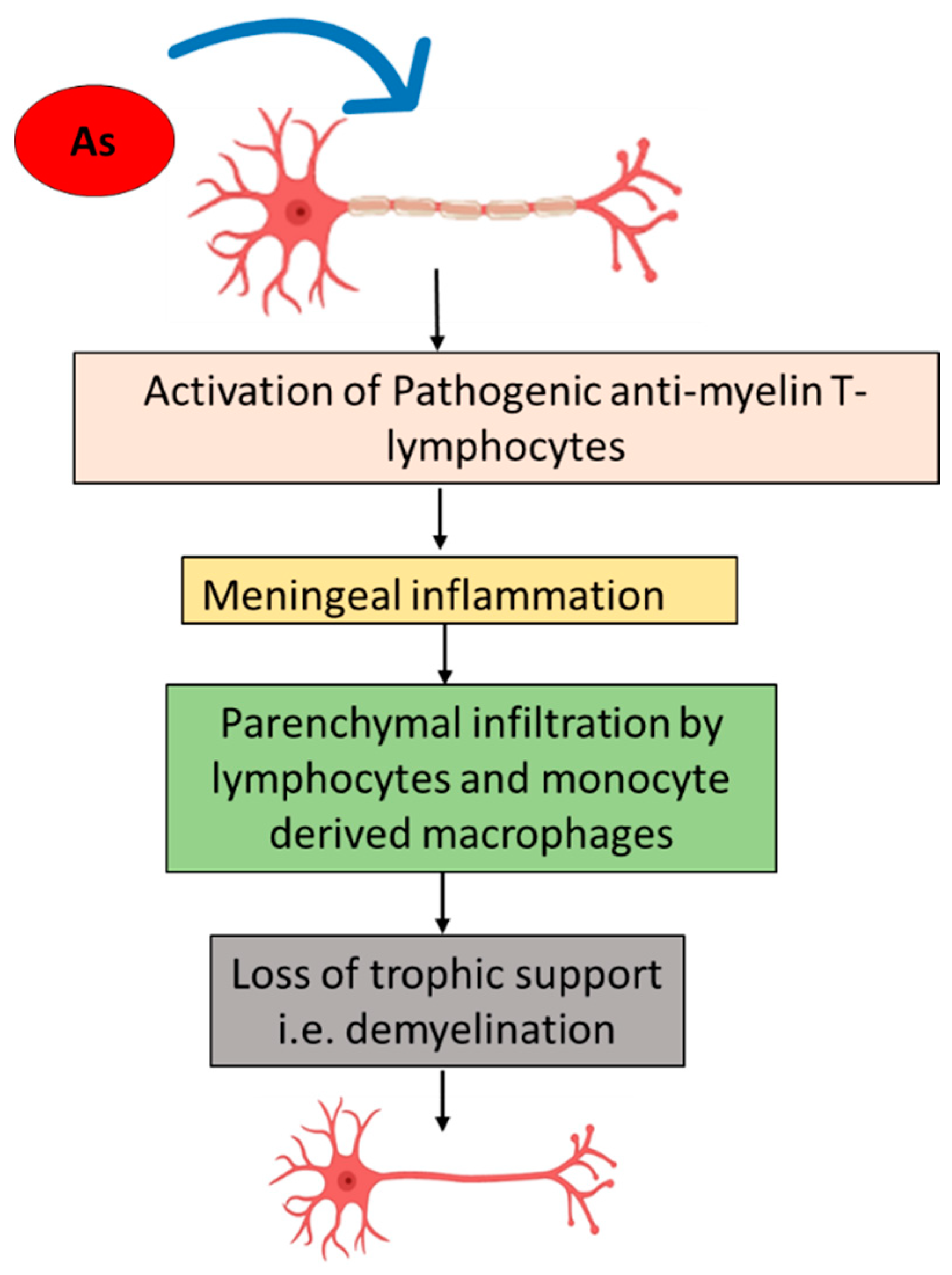
5.3. Demyelination and Myelination
5.4. Effects on Nerve Conduction
5.5. Overview of Epigenetical and Transgenerational Effects
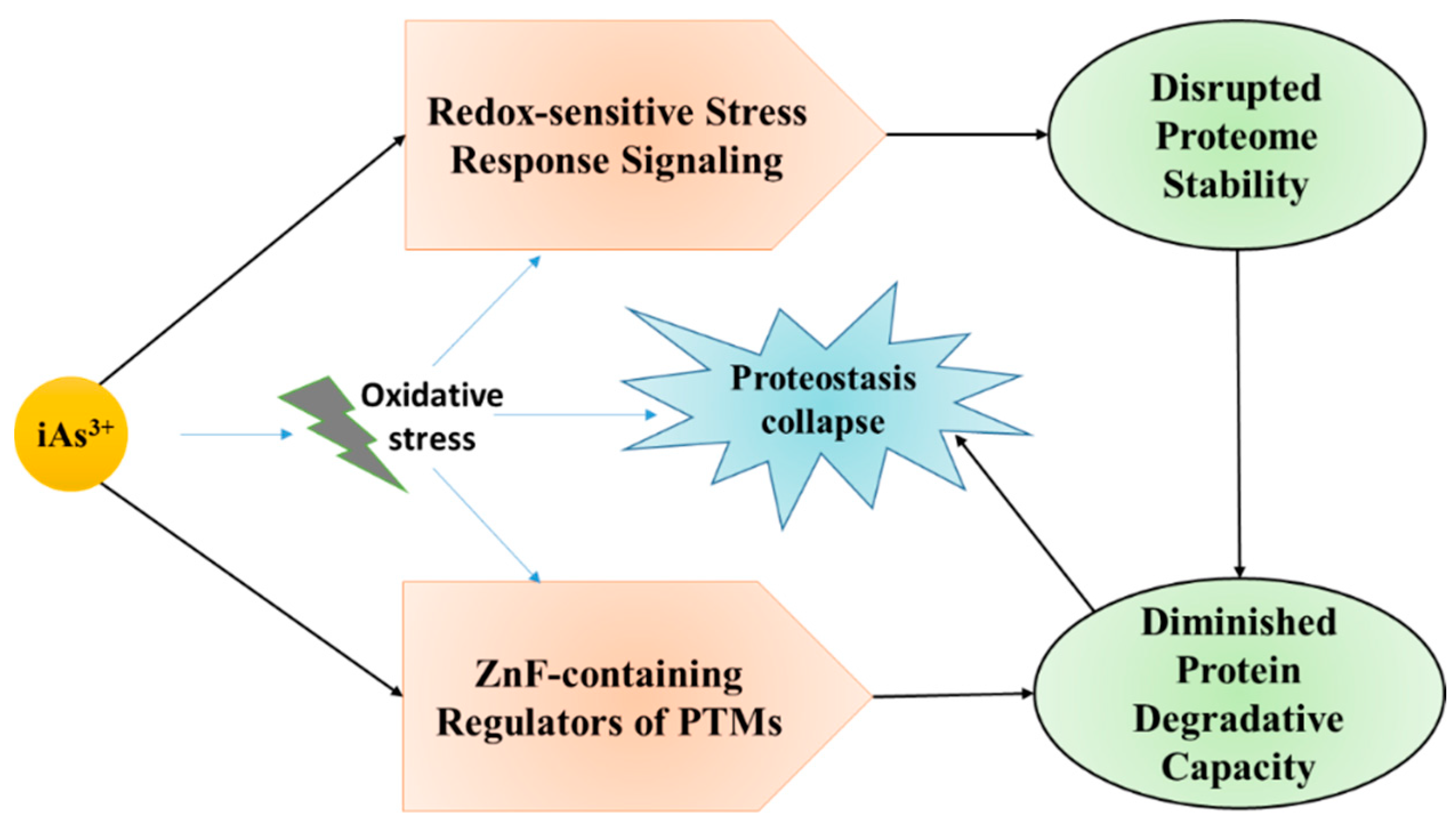
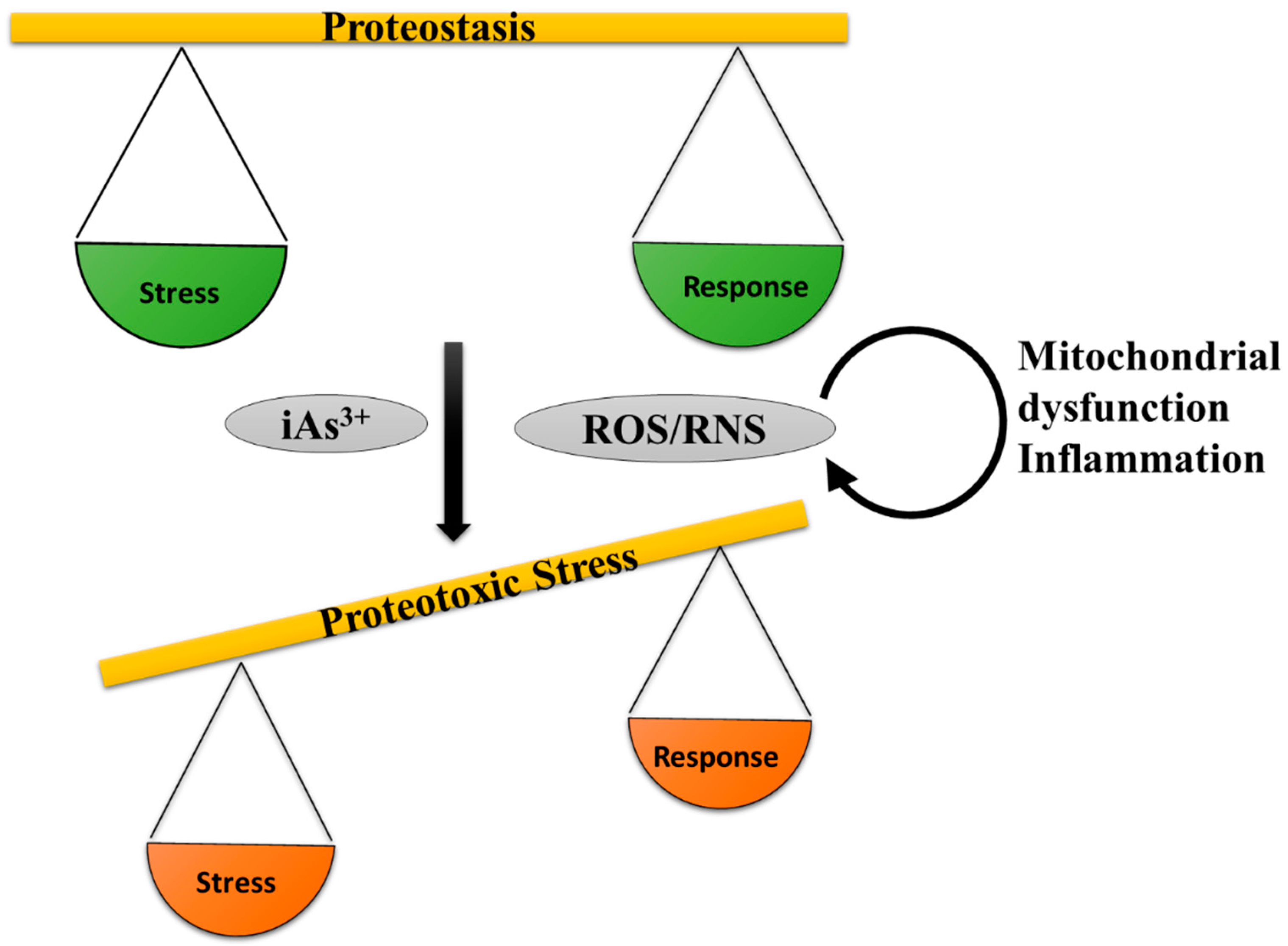
5.6. Proteinopathy and Arsenic Toxicity
6. Strategies for the Removal of Arsenic
7. Conclusions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s disease |
| APP | Amyloid precursor protein |
| As | Arsenic |
| BDNF | Brain-derived neurotrophic factor |
| CRFR1 | Corticotrophin-releasing factor |
| DA | Dopamine |
| EPA | Environmental protection agency |
| GSH | Glutathione |
| HPA | Hypothalamic-pituitary-adrenal |
| MMA | Monomethyl arsonate |
| NCS | Nerve conduction studies |
| NCV | Nerve conduction velocity |
| NO | Nitric oxide |
| PDH | Pyruvate dehydrogenase |
| ROS | Reactive oxygen species |
| RBC | Red blood cells |
| SOD1 | Superoxide dismutase |
| SOD2 | Superoxide dismutase |
| SDH | Succinate dehydrogenase |
| SAM | S-adenosylmethionine |
| -SH | Thiol |
| UPS | Ubiquitin-proteasome system |
| WBC | White blood cells |
| WHO | World health organization |
References
- Kurosawa, K.; Egashira, K.; Tani, M.; Jahiruddin, M.; Moslehuddin, A.Z.M.; Rahman, Z.M. Groundwater–soil–crop relationship with respect to arsenic contamination in farming villages of Bangladesh—A preliminary study. Environ. Pollut. 2008, 156, 563–565. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.; Juhasz, A.; Weber, J.; Naidu, R. Arsenic uptake and speciation in rice plants grown under greenhouse conditions with arsenic contaminated irrigation water. Sci. Total Environ. 2008, 392, 277–283. [Google Scholar] [CrossRef]
- Ayotte, J.D.; Montgomery, D.L.; Flanagan, S.M.; Robinson, K.W. Arsenic in Groundwater in Eastern New England: Occurrence, Controls, and Human Health Implications. Environ. Sci. Technol. 2003, 37, 2075–2083. [Google Scholar] [CrossRef]
- Karagas, M.R.; Stukel, T.; Tosteson, T.D. Assessment of cancer risk and environmental levels of arsenic in New Hampshire. Int. J. Hyg. Environ. Health 2002, 205, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Sengupta, M.K.; Hossain, M.A.; Ahamed, S.; Das, B.; Nayak, B.; Lodh, D.; Rahman, M.M.; Chakraborti, D. Arsenic contamination in groundwater: A global perspective with emphasis on the Asian scenario. J. Health Popul. Nutr. 2006, 24, 142–163. [Google Scholar] [PubMed]
- Tchounwou, P.B.; Centeno, J.A.; Patlolla, A.K. Arsenic toxicity, mutagenesis, and carcinogenesis—A health risk assessment and management approach. Mol. Cell. Biochem. 2004, 255, 47–55. [Google Scholar] [CrossRef] [PubMed]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. Some Drinking-Water Disinfectants and Con-taminants, including Arsenic, IARC Monographs on the Evaluation of Carcinogenic Risks to Humans. IARC Monogr. Eval. Carcinog. Risks Hum. 2004, 84, 1–477. [Google Scholar]
- National Research Council. Arsenic in Drinking Water; The National Academies Press: Washington, DC, USA, 1999. [Google Scholar] [CrossRef]
- Hong, H.; Wu, H.; Chen, J.; Wu, B.; Yu, H.; Yan, B.; Liang, Y. Cytotoxicity induced by iodinated haloacetamides via ROS accumulation and apoptosis in HepG-2 cells. Environ. Pollut. 2018, 242, 191–197. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, H.; Zhang, J.; Ndayambaje, E.; Lin, H.; Chen, J.; Hong, H. Chronic exposure to dichloroacetamide induces biochemical and histopathological changes in the gills of zebrafish. Environ. Toxicol. 2019, 34, 781–787. [Google Scholar] [CrossRef]
- Pedlar, R.; Ptashynski, M.; Wautier, K.; Evans, R.; Baron, C.; Klaverkamp, J. The accumulation, distribution, and toxicological effects of dietary arsenic exposure in lake whitefish (Coregonus clupeaformis) and lake trout (Salvelinus namaycush). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2002, 131, 73–91. [Google Scholar] [CrossRef]
- Datta, S.; Ghosh, D.; Saha, D.R.; Bhattacharaya, S.; Mazumder, S. Chronic exposure to low concentration of arsenic is immunotoxic to fish: Role of head kidney macrophages as biomarkers of arsenic toxicity to Clarias batrachus. Aquat. Toxicol. 2009, 92, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Dauphiné, D.C.; Smith, A.H.; Yuan, Y.; Balmes, J.R.; Bates, M.N.; Steinmaus, C. Case-Control Study of Arsenic in Drinking Water and Lung Cancer in California and Nevada. Int. J. Environ. Res. Public Health 2013, 10, 3310–3324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maimaitiyiming, Y.; Zhu, H.-H.; Yang, C.; Naranmandura, H. Biotransformation of arsenic trioxide by AS3MT favors eradication of acute promyelocytic leukemia: Revealing the hidden facts. Drug Metab. Rev. 2020, 52, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Torres, J.; Santos, P.; Ferrari, C.; Kremer, C.; Kremer, E. Solution Chemistry of Arsenic Anions in the Presence of Metal Cations. J. Solut. Chem. 2017, 46, 2231–2247. [Google Scholar] [CrossRef]
- Santra, S.C.; Samal, A.C.; Bhattacharya, P.; Banerjee, S.; Biswas, A.; Majumdar, J. Arsenic in Foodchain and Community Health Risk: A Study in Gangetic West Bengal. Procedia Environ. Sci. 2013, 18, 2–13. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Yang, Y.; Shao, H.; Sun, W.; Gu, M.; Wang, H.; Jiang, L.; Qu, L.; Sun, D.; Gao, Y. Sodium Arsenite-Induced Learning and Memory Impairment Is Associated with Endoplasmic Reticulum Stress-Mediated Apoptosis in Rat Hippocampus. Front. Mol. Neurosci. 2017, 10, 286. [Google Scholar] [CrossRef] [Green Version]
- Lewchalermvong, K.; Rangkadilok, N.; Nookabkaew, S.; Suriyo, T.; Satayavivad, J. Arsenic Speciation and Accumulation in Selected Organs after Oral Administration of Rice Extracts in Wistar Rats. J. Agric. Food Chem. 2018, 66, 3199–3209. [Google Scholar] [CrossRef]
- Chen, P.; Miah, M.R.; Aschner, M. Metals and Neurodegeneration. F1000Research 2016, 5, 366. [Google Scholar] [CrossRef]
- Uttara, B.; Singh, A.; Zamboni, P.; Mahajan, R. Oxidative Stress and Neurodegenerative Diseases: A Review of Upstream and Downstream Antioxidant Therapeutic Options. Curr. Neuropharmacol. 2009, 7, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Breijyeh, Z.; Karaman, R. Comprehensive Review on Alzheimer’s Disease: Causes and Treatment. Molecules 2020, 25, 5789. [Google Scholar] [CrossRef]
- Baker, B.A.; Cassano, V.A.; Murray, C. Arsenic Exposure, Assessment, Toxicity, Diagnosis, and Management: Guidance for Occupational and Environmental Physicians. J. Occup. Environ. Med. 2018, 60, e634–e639. [Google Scholar] [CrossRef]
- Valberg, P.; Beck, B.; Bowers, T.; Keating, J.; Bergstrom, P.; Boardman, P. Issues in Setting Health-Based Cleanup Levels for Arsenic in Soil. Regul. Toxicol. Pharmacol. 1997, 26, 219–229. [Google Scholar] [CrossRef]
- WHO. Arsenic—World Health Organizaiton. Fact Sheets. 2018. Available online: https://www.who.int/news-room/fact-sheets/detail/arsenic (accessed on 3 August 2021).
- Martinez, V.D.; Vucic, E.; Becker-Santos, D.D.; Gil, L.; Lam, W.L. Arsenic Exposure and the Induction of Human Cancers. J. Toxicol. 2011, 2011, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Xiao, T.; Zhang, A. Association between and risk of trace elements related to skin and liver damage induced by arsenic from c oal burning. Ecotoxicol. Environ. Saf. 2021, 208, 111719. [Google Scholar] [CrossRef]
- Bhattacharya, P.; Jacks, G.; Ahmed, K.M.; Routh, J.; Khan, A.A. Arsenic in Groundwater of the Bengal Delta Plain Aquifers in Bangladesh. Bull. Environ. Contam. Toxicol. 2002, 69, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, P.; Frisbie, S.; Smith, E.; Naidu, R.; Jacks, G. Arsenic in the Environment: A Global Perspective. In Handbook of Heavy Metals in the Environment; Sarker, B., Ed.; Marcell Dekker Inc.: New York, NY, USA, 2001. [Google Scholar]
- World Health Organization. Guidelines for Drinking-Water Quality; World Health Organization: Geneva, Switzerland, 1993. [Google Scholar]
- Gomez-Caminero, A.; Howe, P.D.; Hughes, M.; Kenyon, E.; Lewis, D.; Moore, M.; Aitio, A.; Becking, G.C.; Ng, J. Arsenic and Arsenic Compounds; World Health Organization: Geneva, Switzerland, 2001. [Google Scholar]
- Argos, M.; Ahsan, H.; Graziano, J.H. Arsenic and human health: Epidemiologic progress and public health implications. Rev. Environ. Health 2012, 27, 191–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhury, U.K.; Biswas, B.K.; Chowdhury, T.R.; Samanta, G.; Mandal, B.K.; Basu, G.C.; Chanda, C.R.; Lodh, D.; Saha, K.C.; Mukherjee, S.K. Groundwater arsenic contamination in Bangladesh and West Bengal, India. Environ. Health Perspect. 2000, 108, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Berg, M.; Tran, H.C.; Nguyen, T.C.; Pham, H.V.; Schertenleib, R.; Giger, W. Arsenic Contamination of Groundwater and Drinking Water in Vietnam: A Human Health Threat. Environ. Sci. Technol. 2001, 35, 2621–2626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nriagu, J.O. Arsenic in the environment (Part II, Human health and ecosystem effects). Adv. Environ. Sci. Technol. 1994, 164, 237–238. [Google Scholar]
- Chen, S.-L.; Dzeng, S.R.; Yang, M.-H.; Chiu, K.-H.; Shieh, G.-M.; Wai, C.M. Arsenic Species in Groundwaters of the Blackfoot Disease Area, Taiwan. Environ. Sci. Technol. 1994, 28, 877–881. [Google Scholar] [CrossRef]
- Smedley, P.; Kinniburgh, D.; Macdonald, D.; Nicolli, H.; Barros, A.; Tullio, J.; Pearce, J.; Alonso, M. Arsenic associations in sediments from the loess aquifer of La Pampa, Argentina. Appl. Geochem. 2005, 20, 989–1016. [Google Scholar] [CrossRef]
- Grantham, D.A.; Jones, J.F. Arsenic Contamination of Water Wells in Nova Scotia. J. Am. Water Work. Assoc. 1977, 69, 653–657. [Google Scholar] [CrossRef]
- Chakraborti, D.; Sengupta, M.K.; Rahman, M.M.; Ahamed, S.; Chowdhury, U.K.; Hossain, A.; Mukherjee, S.C.; Pati, S.; Saha, K.C.; Dutta, R.N.; et al. Groundwater arsenic contamination and its health effects in the Ganga-Meghna-Brahmaputra plain. J. Environ. Monit. 2004, 6, 74N–83N. [Google Scholar] [CrossRef]
- Al Rmalli, S.; Haris, P.; Harrington, C.; Ayub, M. A survey of arsenic in foodstuffs on sale in the United Kingdom and imported from Bangladesh. Sci. Total Environ. 2005, 337, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Le, X.C.; Cullen, W.R.; Reimer, K.J. Human urinary arsenic excretion after one-time ingestion of seaweed, crab, and shrimp. Clin. Chem. 1994, 40, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Lu, F. Blackfoot disease: Arsenic or humic acid? Lancet 1990, 336, 115–116. [Google Scholar] [CrossRef]
- Jones, A.S.; Marini, J.; Solo-Gabriele, H.M.; Robey, N.M.; Townsend, T.G. Arsenic, copper, and chromium from treated wood products in the US disposal sector. Waste Manag. 2019, 87, 731–740. [Google Scholar] [CrossRef]
- Morais, S.; Fonseca, H.; Oliveira, S.; Oliveira, H.; Gupta, V.; Sharma, B.; Pereira, M.D.L. Environmental and Health Hazards of Chromated Copper Arsenate-Treated Wood: A Review. Int. J. Environ. Res. Public Health 2021, 18, 5518. [Google Scholar] [CrossRef]
- Goodman, T. Human Health Aspects. In Toxic Cyanobacteria in Water, a Guide to Their Public Health Consequences, Monitoring and Management, 1999; Chorus, I., Bartram, J., Eds.; E & FN Spon: London, UK, 1999. [Google Scholar]
- Ball, V.E. Output, input, and productivity measurement in US agriculture 1948–1979. Am. J. Agric. Econ. 1985, 67, 475–486. [Google Scholar] [CrossRef] [Green Version]
- Bustaffa, E.; Gorini, F.; Bianchi, F.; Minichilli, F. Factors Affecting Arsenic Methylation in Contaminated Italian Areas. Int. J. Environ. Res. Public Health 2020, 17, 5226. [Google Scholar] [CrossRef]
- Tseng, C.-H. Blackfoot Disease and Arsenic: A Never-Ending Story. J. Environ. Sci. Health Part C 2005, 23, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Ferreccio, C.; González, C.; Milosavjlevic, V.; Marshall, G.; Sancha, A.M.; Smith, A.H. Lung Cancer and Arsenic Concentrations in Drinking Water in Chile. Epidemiology 2000, 11, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Morales, K.H.; Ryan, L.; Kuo, T.-L.; Wu, M.-M.; Chen, C.-J. Risk of internal cancers from arsenic in drinking water. Environ. Health Perspect. 2000, 108, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Steinmaus, C.; Yuan, Y.; Bates, M.N.; Smith, A.H. Case-Control Study of Bladder Cancer and Drinking Water Arsenic in the Western United States. Am. J. Epidemiol. 2003, 158, 1193–1201. [Google Scholar] [CrossRef]
- Jacobson-Kram, D.; Mushak, P.; Piscator, M.; Sivulka, D. Health Assessment Document for Inorganic Arsenic; Final Report; NTIS: Springfield, VA, USA, 1984; Volume 1984.
- Kuivenhoven, M.; Mason, K. Arsenic Toxicity. In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2021. [Google Scholar]
- Chung, J.-Y.; Yu, S.-D.; Hong, Y.-S. Environmental Source of Arsenic Exposure. J. Prev. Med. Public Health 2014, 47, 253–257. [Google Scholar] [CrossRef] [Green Version]
- Mandal, B.K.; Suzuki, K.T. Arsenic round the world: A review. Talanta 2002, 58, 201–235. [Google Scholar] [CrossRef]
- Yadav, M.K.; Saidulu, D.; Gupta, A.K.; Ghosal, P.S.; Mukherjee, A. Status and management of arsenic pollution in groundwater: A comprehensive appraisal of recent global scenario, human health impacts, sustainable field-scale treatment technologies. J. Environ. Chem. Eng. 2021, 9, 105203. [Google Scholar] [CrossRef]
- Sahu, R.; Saxena, P.; Johnson, S.; Mathur, H.; Agarwal, H. Heavy metals in cosmetics. Cent. Sci. Environ. 2014. Available online: https://cdn.cseindia.org/userfiles/Heavy_Metals_in_Cosmetics_Report.pdf (accessed on 3 August 2021).
- Hostynek, J. Toxic potential from metals absorbed through the skin. Cosmet. Toilet. 1998, 113, 33–42. [Google Scholar]
- Borowska, S.; Brzóska, M.M. Metals in cosmetics: Implications for human health. J. Appl. Toxicol. 2015, 35, 551–572. [Google Scholar] [CrossRef]
- Atz, V.L.; Pozebon, D. Graphite furnace atomic absorption spectrometry (GFAAS) methodology for trace element determination in eye shadow and lipstick. At. Spectrosc. 2009, 30, 82–91. [Google Scholar]
- Saint-Jacques, N.; Parker, L.; Brown, P.; Dummer, T.J. Arsenic in drinking water and urinary tract cancers: A systematic review of 30 years of epidemiological evidence. Environ. Health 2014, 13, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kharroubi, W.; Ahmed, S.H.; Nury, T.; Andreoletti, P.; Sakly, R.; Hammami, M.; Lizard, G. Mitochondrial dysfunction, oxidative stress and apoptotic induction in microglial BV-2 cells treated with sodium arsenate. J. Environ. Sci. 2017, 51, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Khairul, I.; Wang, Q.Q.; Jiang, Y.H.; Wang, C.; Naranmandura, H. Metabolism, toxicity and anticancer activities of arsenic compounds. Oncotarget 2017, 8, 23905–23926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gbaruko, B.C.; Ana, G.; Nwachukwu, J.K. Ecotoxicology of arsenic in the hydrosphere: Implications for public health. Afr. J. Biotechnol. 2008, 7, 4737–4742. [Google Scholar]
- Engström, K.S.; Broberg, K.; Concha, G.; Nermell, B.; Warholm, M.; Vahter, M. Genetic Polymorphisms Influencing Arsenic Metabolism: Evidence from Argentina. Environ. Health Perspect. 2007, 115, 599–605. [Google Scholar] [CrossRef] [Green Version]
- Pavlogeorgatos, G.; Vasilis, K. The importance of mercury determination and speciation to the health of the general population. Glob. NEST Int. J. 2003, 4, 107–125. [Google Scholar]
- Jha, A.; Noditi, M.; Nilsson, R.; Natarajan, A. Genotoxic effects of sodium arsenite on human cells. Mutat. Res. Mol. Mech. Mutagen. 1992, 284, 215–221. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, F.; Liao, Y.; Jin, Y.; Sun, G. Effects of arsenite in astrocytes on neuronal signaling transduction. Toxicology 2012, 303, 43–53. [Google Scholar] [CrossRef]
- Sinha, D.; Prasad, P. Health effects inflicted by chronic low-level arsenic contamination in groundwater: A global public health challenge. J. Appl. Toxicol. 2019, 40, 87–131. [Google Scholar] [CrossRef]
- Itoh, T.; Zhang, Y.F.; Murai, S.; Saito, H.; Nagahama, H.; Miyate, H.; Saito, Y.; Abe, E. The effect of arsenic trioxide on brain monoamine metabolism and locomotor activity of mice. Toxicol. Lett. 1990, 54, 345–353. [Google Scholar] [PubMed]
- Hong, Y.-S.; Song, K.-H.; Chung, J.-Y. Health Effects of Chronic Arsenic Exposure. J. Prev. Med. Public Health 2014, 47, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.D.; Rahman, M.M.; Mise, N.; Sikder, M.T.; Ichihara, G.; Khabiruddin, M.; Kurasaki, M.; Ichihara, S. Environmental arsenic exposure and its contribution to human disease, toxicity mechanism and management. Environ. Pollut. 2021, 289, 117940. [Google Scholar] [CrossRef] [PubMed]
- Tolins, M.; Ruchirawat, M.; Landrigan, P. The Developmental Neurotoxicity of Arsenic: Cognitive and Behavioral Consequences of Early Life Exposure. Ann. Glob. Health 2014, 80, 303–314. [Google Scholar] [CrossRef]
- Calderon, J.; Navarro, M.; Capdevillea, M.; Diaza, M.; Golden, A.; Leyvaa, I.; Aburtod, V.; Barriga, F.D. Exposure to Arsenic and Lead and Neuropsychological Development in Mexican Children. Environ. Res. 2001, 85, 69–76. [Google Scholar] [CrossRef]
- Li, Z.; Li, X.; Qian, Y.; Guo, C.; Wang, Z.; Wei, Y. The sustaining effects of e-waste-related metal exposure on hypothalmus-pituitary-adrenal axis reactivity and oxidative stress. Sci. Total Environ. 2020, 739, 139964. [Google Scholar] [CrossRef]
- Alao, M.E.; Perin, J.; Brooks, W.A.; Hossain, L.; Goswami, D.; Zaman, K.; Yunus, M.; Khan, A.F.; Jahan, Y.; Ahmed, D.; et al. Urinary arsenic is associated with wasting and underweight status in young children in rural Bangladesh. Environ. Res. 2020, 195, 110025. [Google Scholar] [CrossRef]
- Florea, A.-M.; Splettstoesser, F.; Büsselberg, D. Arsenic trioxide (As2O3) induced calcium signals and cytotoxicity in two human cell lines: SY-5Y neuroblastoma and 293 embryonic kidney (HEK). Toxicol. Appl. Pharmacol. 2007, 220, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Malavasi, F.; Deaglio, S.; Funaro, A.; Ferrero, E.; Horenstein, A.L.; Ortolan, E.; Vaisitti, T.; Aydin, S. Evolution and Function of the ADP Ribosyl Cyclase/CD38 Gene Family in Physiology and Pathology. Physiol. Rev. 2008, 88, 841–886. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.-W.; Liou, S.-H.; Hsueh, Y.-M.; Lyu, W.-S.; Liu, C.-S.; Liu, H.-J.; Chung, M.-C.; Hung, P.-H.; Chung, C.-J. Risk of Alzheimer’s disease with metal concentrations in whole blood and urine: A case–control study using propensity score matching. Toxicol. Appl. Pharmacol. 2018, 356, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Sharma, B.; Sharma, P. Arsenic toxicity induced endothelial dysfunction and dementia: Pharmacological interdiction by histone deacetylase and inducible nitric oxide synthase inhibitors. Toxicol. Appl. Pharmacol. 2013, 273, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Martinez, E.J.; Kolb, B.L.; Bell, A.; Savage, D.D.; Allan, A.M. Moderate perinatal arsenic exposure alters neuroendocrine markers associated with depression and increases depressive-like behaviors in adult mouse offspring. NeuroToxicology 2008, 29, 647–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vahidnia, A.; Romijn, F.; Tiller, M.; Van Der Voet, G.B.; De Wolff, F.A. Arsenic-induced toxicity: Effect on protein composition in sciatic nerve. Hum. Exp. Toxicol. 2006, 25, 667–674. [Google Scholar] [CrossRef]
- Barguilla, I.; Peremartí, J.; Bach, J.; Marcos, R.; Hernández, A. Role of As3mt and Mth1 in the genotoxic and carcinogenic effects induced by long-term exposures to arsenic in MEF cells. Toxicol. Appl. Pharmacol. 2020, 409, 115303. [Google Scholar] [CrossRef]
- Medda, N.; Patra, R.; Ghosh, T.K.; Maiti, S. Neurotoxic mechanism of arsenic: Synergistic effect of effect of mitochondrial instability, oxidative stress and hormonal - neurotransmitter impairment. Biol. Trace Elem. Res. 2020, 198, 8–15. [Google Scholar] [CrossRef]
- Niño, S.A.; Martel-Gallegos, G.; Castro-Zavala, A.; Ortega-Berlanga, B.; Delgado, J.M.; Hernández-Mendoza, H.; Romero-Guzmán, E.; Ríos-Lugo, J.; Rosales-Mendoza, S.; Jiménez-Capdeville, M.E.; et al. Chronic Arsenic Exposure Increases Aβ(1–42) Production and Receptor for Advanced Glycation End Products Expression in Rat Brain. Chem. Res. Toxicol. 2017, 31, 13–21. [Google Scholar] [CrossRef]
- Hardy, J.; Selkoe, D.J. The amyloid hypothesis of Alzheimer’s disease: Progress and problems on the road to therapeutics. Science 2002, 297, 353–356. [Google Scholar] [CrossRef] [Green Version]
- Gouras, G.K.; Olsson, T.T.; Hansson, O. β-Amyloid peptides and amyloid plaques in Alzheimer’s disease. Neurotherapeutics 2015, 12, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Virk, D.; Kumar, A.; Jaggi, A.S.; Singh, N. Ameliorative role of rolipram, PDE-4 inhibitor, against sodium arsenite–induced vascular dementia in rats. Environ. Sci. Pollut. Res. 2021. [Google Scholar] [CrossRef]
- Ashok, A.; Rai, N.K.; Tripathi, S.; Bandyopadhyay, S. Exposure to As-, Cd-, and Pb-Mixture Induces Aβ, Amyloidogenic APP Processing and Cognitive Impairments via Oxidative Stress-Dependent Neuroinflammation in Young Rats. Toxicol. Sci. 2014, 143, 64–80. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Li, J.; Zheng, J.; Qin, S. Reactive Astrocytes in Neurodegenerative Diseases. Aging Dis. 2019, 10, 664–675. [Google Scholar] [CrossRef] [Green Version]
- Shavali, S.; Sens, D.A. Synergistic neurotoxic effects of arsenic and dopamine in human dopaminergic neuroblastoma SH-SY5Y cells. Toxicol. Sci. 2008, 102, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Lu, C.; Wang, J.; Hu, W.; Cao, Z.; Sun, D.; Xia, H.; Ma, X. Developmental mechanisms of arsenite toxicity in zebrafish (Danio rerio) embryos. Aquat. Toxicol. 2009, 91, 229–237. [Google Scholar] [CrossRef]
- Niño, S.A.; Morales-Martínez, A.; Chi-Ahumada, E.; Carrizales, L.; Delgado, R.S.; Pérez-Severiano, F.; Díaz-Cintra, S.; Jiménez-Capdeville, M.E.; Zarazua, S. Arsenic Exposure Contributes to the Bioenergetic Damage in an Alzheimer’s Disease Model. ACS Chem. Neurosci. 2018, 10, 323–336. [Google Scholar] [CrossRef]
- Liu, X.; Piao, F.; Li, Y. Protective Effect of Taurine on the Decreased Biogenic Amine Neurotransmitter Levels in the Brain of Mice Exposed to Arsenic. Adv. Exp. Med. Biol. 2013, 776, 277–287. [Google Scholar] [CrossRef]
- Dipp, V.R.; Valles, S.; Ortiz-Kerbertt, H.; Suarez, J.V.; Bardullas, U. Neurobehavioral Alterations in Zebrafish Due to Long-Term Exposure to Low Doses of Inorganic Arsenic. Zebrafish 2018, 15, 575–585. [Google Scholar] [CrossRef]
- Ellinsworth, D.C. Arsenic, reactive oxygen, and endothelial dysfunction. J. Pharmacol. Exp. Ther. 2015, 353, 458–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pou, S.; Pou, W.; Bredt, D.; Snyder, S.; Rosen, G. Generation of superoxide by purified brain nitric oxide synthase. J. Biol. Chem. 1992, 267, 24173–24176. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Hayashi, H.; Hasegawa, A.; Yamanaka, K.; Okada, S. DNA damage induced in cultured human alveolar (L-132) cells by exposure to dimethylarsinic acid. Environ. Health Perspect. 1994, 102, 285–288. [Google Scholar] [PubMed]
- Xu, M.; Niu, Q.; Hu, Y.; Feng, G.; Wang, H.; Li, S. Proanthocyanidins Antagonize Arsenic-Induced Oxidative Damage and Promote Arsenic Methylation through Activation of the Nrf2 Signaling Pathway. Oxidative Med. Cell. Longev. 2019, 2019, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Yin, Y.; Zhang, J.; Yu, H.; Wang, X.; Wu, J.; Xue, Y. Hydroxyl radical generation and oxidative stress in Carassius auratus liver, exposed to pyrene. Ecotoxicol. Environ. Saf. 2008, 71, 446–453. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Kim, J.-M. Arsenic Toxicity in Male Reproduction and Development. Dev. Reprod. 2015, 19, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Liu, J.; Liu, B.; Liu, X.; Yu, X. Prenatal exposure to arsenic and neurobehavioral development of newborns in China. Environ. Int. 2018, 121, 421–427. [Google Scholar] [CrossRef]
- Sarkar, S.; Mukherjee, S.; Chattopadhyay, A.; Bhattacharya, S. Low dose of arsenic trioxide triggers oxidative stress in zebrafish brain: Expression of antioxidant genes. Ecotoxicol. Environ. Saf. 2014, 107, 1–8. [Google Scholar] [CrossRef]
- de Castro, M.R.; Lima, J.V.; de Freitas, D.P.S.; de Souza Valente, R.; Dummer, N.S.; de Aguiar, R.B.; dos Santos, L.C.; Marins, L.F.; Geracitano, L.A.; Monserrat, J.M. Behavioral and neurotoxic effects of arsenic exposure in zebrafish (Danio rerio, Teleostei: Cyprinidae). Comp. Biochem. Physiol. C Toxicol. 2009, 150, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Mejía, J.; Barriga, F.D.; Calderón, J.; Ríos, C.; Jiménez-Capdeville, M. Effects of Lead–Arsenic Combined Exposure on Central Monoaminergic Systems. Neurotoxicol. Teratol. 1997, 19, 489–497. [Google Scholar] [CrossRef]
- Yadav, R.S.; Shukla, R.K.; Sankhwar, M.L.; Patel, D.K.; Ansari, R.W.; Pant, A.B.; Islam, F.; Khanna, V.K. Neuroprotective effect of curcumin in arsenic-induced neurotoxicity in rats. NeuroToxicology 2010, 31, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Nagaraja, T.; Desiraju, T. Effects on Operant Learning and Brain Acetylcholine Esterase Activity in Rats following Chronic Inorganic Arsenic Intake. Hum. Exp. Toxicol. 1994, 13, 353–356. [Google Scholar] [CrossRef]
- Grozio, A.; Sociali, G.; Sturla, L.; Caffa, I.; Soncini, D.; Salis, A.; Raffaelli, N.; De Flora, A.; Nencioni, A.; Bruzzone, S. CD73 Protein as a Source of Extracellular Precursors for Sustained NAD+ Biosynthesis in FK866-treated Tumor Cells. J. Biol. Chem. 2013, 288, 25938–25949. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Su, J.; Yao, S.; Zhang, Y.; Cao, F.; Wang, F.; Wang, H.; Li, J.; Xi, S. Fluoride and Arsenic Exposure Impairs Learning and Memory and Decreases mGluR5 Expression in the Hippocampus and Cortex in Rats. PLoS ONE 2014, 9, e96041. [Google Scholar] [CrossRef] [Green Version]
- Kannan, G.M.; Tripathi, N.; Dube, S.N.; Gupta, M.; Flora, S.; Flora, S.J.S. Toxic Effects of Arsenic (III) on Some Hematopoietic and Central Nervous System Variables in Rats and Guinea Pigs. J. Toxicol. Clin. Toxicol. 2001, 39, 675–682. [Google Scholar] [CrossRef] [PubMed]
- Gresser, M. ADP-arsenate. Formation by submitochondrial particles under phosphorylating conditions. J. Biol. Chem. 1981, 256, 5981–5983. [Google Scholar] [CrossRef]
- Nagaraja, T.; Desiraju, T. Regional alterations in the levels of brain biogenic amines, glutamate, GABA, and GAD activity due to chronic consumption of inorganic arsenic in developing and adult rats. Bull. Environ. Contam. Toxicol. 1993, 50. [Google Scholar] [CrossRef] [PubMed]
- Baldissarelli, L.A.; Capiotti, K.M.; Bogo, M.R.; Ghisleni, G.; Bonan, C.D. Arsenic alters behavioral parameters and brain ectonucleotidases activities in zebrafish (Danio rerio). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2012, 155, 566–572. [Google Scholar] [CrossRef] [Green Version]
- Valles, S.; Hernández-Sánchez, J.; Dipp, V.R.; Huerta-González, D.; Olivares-Bañuelos, T.N.; Gonzalez-Fraga, J.A.; Bardullas, U. Exposure to low doses of inorganic arsenic induces transgenerational changes on behavioral and epigenetic markers in zebrafish (Danio rerio). Toxicol. Appl. Pharmacol. 2020, 396, 115002. [Google Scholar] [CrossRef]
- Chattopadhyay, S.; Bhaumik, S.; Chaudhury, A.N.; Das Gupta, S. Arsenic induced changes in growth development and apoptosis in neonatal and adult brain cells in vivo and in tissue culture. Toxicol. Lett. 2002, 128, 73–84. [Google Scholar] [CrossRef]
- Barros, D.M.; e Souza, T.M.; De David, T.; Choi, H.; Aguzzoli, A.; Madche, C.; Ardenghi, P.; Medina, J.H.; Izquierdo, I. Simultaneous modulation of retrieval by dopaminergic D1, β-noradrenergic, serotonergic-1A and cholinergic muscarinic receptors in cortical structures of the rat. Behav. Brain Res. 2001, 124, 1–7. [Google Scholar] [CrossRef]
- Izquierdo, I.; Bevilaqua, L.R.; Rossato, J.I.; Bonini, J.S.; Medina, J.H.; Cammarota, M. Different molecular cascades in different sites of the brain control memory consolidation. Trends Neurosci. 2006, 29, 496–505. [Google Scholar] [CrossRef]
- Tsai, S.-Y.; Chou, H.-Y.; The, H.-W.; Chen, C.-M.; Chen, C.-J. The Effects of Chronic Arsenic Exposure from Drinking Water on the Neurobehavioral Development in Adolescence. NeuroToxicology 2003, 24, 747–753. [Google Scholar] [CrossRef]
- Mukherjee, S.C.; Rahman, M.M.; Chowdhury, U.K.; Sengupta, M.K.; Lodh, D.; Chanda, C.R.; Saha, K.C.; Chakraborti, D. Neuropathy in Arsenic Toxicity from Groundwater Arsenic Contamination in West Bengal, India. J. Environ. Sci. Health Part A 2003, 38, 165–183. [Google Scholar] [CrossRef]
- Shokoohi, R.; Khazaei, M.; Karami, M.; Seidmohammadi, A.; Berijani, N.; Khotanlou, H.; Torkshavand, Z. The relationship between chronic exposure to arsenic through drinking water and hearing function in exposed population aged 10–49 years: A cross-sectional study. Ecotoxicol. Environ. Saf. 2021, 211, 111939. [Google Scholar] [CrossRef]
- Wasserman, G.A.; Liu, X.; Parvez, F.; Ahsan, H.; Factor-Litvak, P.; van Geen, A.; Slavkovich, V.; Lolacono, N.J.; Cheng, Z.; Hussain, I. Water arsenic exposure and children’s intellectual function in Araihazar, Bangladesh. Environ. Health Perspect. 2004, 112, 1329–1333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, C.; Inaoka, T.; Matsui, T.; Ishigaki, K.; Murayama, N.; Ohtsuka, R. Effects of arsenic on younger generations. J. Environ. Sci. Health Part A 2003, 38, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.-H.; Qiu, Z.-Q.; Shu, W.-Q.; Zhang, Y.-Y.; Zhang, L.; Chen, J.-A. Effects of arsenic exposure from drinking water on spatial memory, ultra-structures and NMDAR gene expression of hippocampus in rats. Toxicol. Lett. 2009, 184, 121–125. [Google Scholar] [CrossRef]
- Rodríguez, V.; Carrizales, L.; Mendoza, M.; Fajardo, O.; Giordano, M. Effects of sodium arsenite exposure on development and behavior in the rat. Neurotoxicol. Teratol. 2002, 24, 743–750. [Google Scholar] [CrossRef]
- Martinez-Finley, E.J.; Ali, A.-M.S.; Allan, A.M. Learning deficits in C57BL/6J mice following perinatal arsenic exposure: Consequence of lower corticosterone receptor levels? Pharmacol. Biochem. Behav. 2009, 94, 271–277. [Google Scholar] [CrossRef] [Green Version]
- Kozul-Horvath, C.D.; Zandbergen, F.; Jackson, B.P.; Enelow, R.I.; Hamilton, J.W. Effects of Low-Dose Drinking Water Arsenic on Mouse Fetal and Postnatal Growth and Development. PLoS ONE 2012, 7, e38249. [Google Scholar] [CrossRef]
- Dong, J.; Su, S.-Y. The Association between Arsenic and Children’s Intelligence: A Meta-analysis. Biol. Trace Elem. Res. 2008, 129, 88–93. [Google Scholar] [CrossRef]
- Liang, C.; Wu, X.; Huang, K.; Yan, S.; Li, Z.; Xia, X.; Pan, W.; Sheng, J.; Tao, R.; Tao, Y.; et al. Domain- and sex-specific effects of prenatal exposure to low levels of arsenic on children’s development at 6 months of age: Findings from the Ma’anshan birth cohort study in China. Environ. Int. 2019, 135, 105112. [Google Scholar] [CrossRef]
- von Ehrenstein, O.S.; Poddar, S.; Yuan, Y.; Mazumder, D.G.; Eskenazi, B.; Basu, A.; Hira-Smith, M.; Ghosh, N.; Lahiri, S.; Haque, R.; et al. Children’s Intellectual Function in Relation to Arsenic Exposure. Epidemiology 2007, 18, 44–51. [Google Scholar] [CrossRef]
- Rocha-Amador, D.; Navarro, M.E.; Carrizales, L.; Morales, R.; Calderon, J. Decreased intelligence in children and exposure to fluoride and arsenic in drinking water. Cad. Saúde Pública 2007, 23, S579–S587. [Google Scholar] [CrossRef]
- O’Bryant, S.E.; Edwards, M.; Menon, C.V.; Gong, G.; Barber, R. Long-Term Low-Level Arsenic Exposure Is Associated with Poorer Neuropsychological Functioning: A Project FRONTIER Study. Int. J. Environ. Res. Public Health 2011, 8, 861–874. [Google Scholar] [CrossRef] [PubMed]
- Wasserman, G.A.; Liu, X.; Parvez, F.; Ahsan, H.; Factor-Litvak, P.; Kline, J.; Van Geen, A.; Slavkovich, V.; LoIacono, N.J.; Levy, D.; et al. Water Arsenic Exposure and Intellectual Function in 6-Year-Old Children in Araihazar, Bangladesh. Environ. Health Perspect. 2007, 115, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Wright, R.O.; Amarasiriwardena, C.; Woolf, A.D.; Jim, R.; Bellinger, D.C. Neuropsychological correlates of hair arsenic, manganese, and cadmium levels in school-age children residing near a hazardous waste site. NeuroToxicology 2006, 27, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Tseng, C.-H. The potential biological mechanisms of arsenic-induced diabetes mellitus. Toxicol. Appl. Pharmacol. 2004, 197, 67–83. [Google Scholar] [CrossRef]
- Lu, T.-H.; Su, C.-C.; Chen, Y.-W.; Yang, C.-Y.; Wu, C.-C.; Hung, D.-Z.; Chen, C.-H.; Cheng, P.-W.; Liu, S.-H.; Huang, C.-F. Arsenic induces pancreatic β-cell apoptosis via the oxidative stress-regulated mitochondria-dependent and endoplasmic reticulum stress-triggered signaling pathways. Toxicol. Lett. 2011, 201, 15–26. [Google Scholar] [CrossRef]
- Jensen, A.; Bro-Rasmussen, F. Environmental Cadmium in Europe. Rev. Environ. Contam. Toxicol. 1992, 125, 101–181. [Google Scholar] [CrossRef]
- Butterfield, D.A.; Castegna, A.; Lauderback, C.M.; Drake, J. Evidence that amyloid beta-peptide-induced lipid peroxidation and its sequelae in Alzheimer’s disease brain contribute to neuronal death1. Neurobiol. Aging 2002, 23, 655–664. [Google Scholar] [CrossRef]
- Chappell, W.; Beck, B.; Brown, K.; Chaney, R.; Cothern, R.; Cothern, C.; Irgolic, K.; North, D.; Thornton, I.; Tsongas, T. Inorganic arsenic: A need and an opportunity to improve risk assessment. Environ. Health Perspect. 1997, 105, 1060–1067. [Google Scholar] [CrossRef]
- Mount, C.; Monje, M. Wrapped to Adapt: Experience-Dependent Myelination. Neuron 2017, 95, 743–756. [Google Scholar] [CrossRef]
- Bustaffa, E.; Stoccoro, A.; Bianchi, F.; Migliore, L. Genotoxic and epigenetic mechanisms in arsenic carcinogenicity. Arch. Toxicol. 2014, 88, 1043–1067. [Google Scholar] [CrossRef]
- Frost, G.; Li, Y.-M. The role of astrocytes in amyloid production and Alzheimer’s disease. Open Biol. 2017, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firdaus, F.; Zafeer, M.F.; Waseem, M.; Ullah, R.; Ahmad, M.; Afzal, M. Thymoquinone alleviates arsenic induced hippocampal toxicity and mitochondrial dysfunction by modulating mPTP in Wistar rats. Biomed. Pharmacother. 2018, 102, 1152–1160. [Google Scholar] [CrossRef] [PubMed]
- Mao, J.; Yang, J.; Zhang, Y.; Li, T.; Wang, C.; Xu, L.; Hu, Q.; Wang, X.; Jiang, S.; Nie, X. Arsenic trioxide mediates HAPI microglia inflammatory response and subsequent neuron apoptosis through p38/JNK MAPK/STAT3 pathway. Toxicol. Appl. Pharmacol. 2016, 303, 79–89. [Google Scholar] [CrossRef] [Green Version]
- Saha, S.; Sadhukhan, P.; Mahalanobish, S.; Dutta, S.; Sil, P.C. Ameliorative role of genistein against age-dependent chronic arsenic toxicity in murine brains via the regulation of oxidative stress and inflammatory signaling cascades. J. Nutr. Biochem. 2018, 55, 26–40. [Google Scholar] [CrossRef]
- Halliwell, B.; Cross, C.E. Oxygen-derived species: Their relation to human disease and environmental stress. Environ. Health Perspect. 1994, 102, 5–12. [Google Scholar]
- Olsen, L.F.; Issinger, O.-G.; Guerra, B. The Yin and Yang of redox regulation. Redox Rep. 2013, 18, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Forman, H.J. Redox signaling: An evolution from free radicals to aging. Free Radic. Biol. Med. 2016, 97, 398–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Culotta, V.C.; Yang, M.; O’Halloran, T.V. Activation of superoxide dismutases: Putting the metal to the pedal. Biochim. Biophys. Acta BBA-Bioenerg. 2006, 1763, 747–758. [Google Scholar] [CrossRef] [Green Version]
- Cohen, S.M.; Arnold, L.L.; Eldan, M.; Lewis, A.S.; Beck, B.D. Methylated Arsenicals: The Implications of Metabolism and Carcinogenicity Studies in Rodents to Human Risk Assessment. Crit. Rev. Toxicol. 2006, 36, 99–133. [Google Scholar] [CrossRef]
- Corsini, E.; Asti, L.; Viviani, B.; Marinovich, M.; Galli, C.L. Sodium Arsenate Induces Overproduction of Interleukin-1α in Murine Keratinocytes: Role of Mitochondria. J. Investig. Dermatol. 1999, 113, 760–765. [Google Scholar] [CrossRef] [Green Version]
- Pieczenik, S.R.; Neustadt, J. Mitochondrial dysfunction and molecular pathways of disease. Exp. Mol. Pathol. 2007, 83, 84–92. [Google Scholar] [CrossRef]
- Dwivedi, N.; Mehta, A.; Yadav, A.; Binukumar, B.; Gill, K.D.; Flora, S.J. MiADMSA reverses impaired mitochondrial energy metabolism and neuronal apoptotic cell death after arsenic exposure in rats. Toxicol. Appl. Pharmacol. 2011, 256, 241–248. [Google Scholar] [CrossRef]
- Flora, S.J. Arsenic-induced Oxidative Stress and Its Reversibility Following Combined Administration Of N-acetylcysteine and Meso 2,3-dimercaptosuccinic Acid in Rats. Clin. Exp. Pharmacol. Physiol. 1999, 26, 865–869. [Google Scholar] [CrossRef]
- Flora, S.J.S.; Bhadauria, S.; Kannan, G.M.; Singh, N. Arsenic induced oxidative stress and the role of antioxidant supplementation during chelation: A review. J. Environ. Biol. 2007, 28. [Google Scholar]
- Prakash, C.; Soni, M.; Kumar, V. Biochemical and Molecular Alterations Following Arsenic-Induced Oxidative Stress and Mitochondrial Dysfunction in Rat Brain. Biol. Trace Elem. Res. 2015, 167, 121–129. [Google Scholar] [CrossRef]
- Gopalkrishnan, A.; Rao, M.V. Amelioration by Vitamin A upon Arsenic Induced Metabolic and Neurotoxic Effects. J. Health Sci. 2006, 52, 568–577. [Google Scholar] [CrossRef] [Green Version]
- Hughes, M.F. Arsenic toxicity and potential mechanisms of action. Toxicol. Lett. 2002, 133, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Dungdung, S.R.; Chowdhury, S.T.; Mandal, A.K.; Sarkar, S.; Ghosh, D.; Das, N. Encapsulation of the flavonoid quercetin with an arsenic chelator into nanocapsules enables the simultaneous delivery of hydrophobic and hydrophilic drugs with a synergistic effect against chronic arsenic accumulation and oxidative stress. Free Radic. Biol. Med. 2011, 51, 1893–1902. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Piao, F.; Zhao, Y.; Li, S.; Wang, Y.; Liu, P. Subchronic exposure to arsenic decreased Sdha expression in the brain of mice. NeuroToxicology 2009, 30, 538–543. [Google Scholar] [CrossRef]
- Orrenius, S.; Zhivotovsky, B.; Nicotera, P. Regulation of cell death: The calcium–apoptosis link. Nat. Rev. Mol. Cell Biol. 2003, 4, 552–565. [Google Scholar] [CrossRef]
- Gibson, G.E.; Chen, H.-L.; Xu, H.; Qiu, L.; Xu, Z.; Denton, T.; Shi, Q. Deficits in the mitochondrial enzyme α-ketoglutarate dehydrogenase lead to Alzheimer’s disease-like calcium dysregulation. Neurobiol. Aging 2012, 33, 1121.e13–1121.e24. [Google Scholar] [CrossRef] [Green Version]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans; World Health Organization; International Agency for Research on Cancer. Some Drinking-Water Disinfectants and Contaminants, Including Arsenic; IARC: Geneva, Switzerland, 2004; Volume 84.
- Lebel, C.; Gee, M.; Camicioli, R.; Wieler, M.; Martin, W.W.; Beaulieu, C. Diffusion tensor imaging of white matter tract evolution over the lifespan. NeuroImage 2011, 60, 340–352. [Google Scholar] [CrossRef]
- McKenzie, I.A.; Ohayon, D.; Li, H.; de Faria, J.P.; Emery, B.; Tohyama, K.; Richardson, W.D. Motor skill learning requires active central myelination. Science 2014, 346, 318–322. [Google Scholar] [CrossRef] [PubMed]
- Pinto, M.M.S.C.; Reis, A.; Almeida, A.; Ordens, C.; Silva, M.M.V.G.; Freitas, S.; Simoes, M.; Moreira, P.; Dinis, P.; Diniz, M.L.; et al. Human predisposition to cognitive impairment and its relation with environmental exposure to potentially toxic elements. Environ. Geochem. Health 2017, 40, 1767–1784. [Google Scholar] [CrossRef]
- Aung, K.H.; Kurihara, R.; Nakashima, S.; Maekawa, F.; Nohara, K.; Kobayashi, T.; Tsukahara, S. Inhibition of neurite outgrowth and alteration of cytoskeletal gene expression by sodium arsenite. NeuroToxicology 2012, 34, 226–235. [Google Scholar] [CrossRef]
- Chandravanshi, L.P.; Gupta, R.; Shukla, R.K. Developmental Neurotoxicity of Arsenic: Involvement of Oxidative Stress and Mitochondrial Functions. Biol. Trace Elem. Res. 2018, 186, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Rumbeiha, W.K.; Oehme, F.W.; Ballantyne, B.; Marrs, T.; Turner, P. Veterinary Toxicology; Stockton Press: New York, NY, USA, 1993. [Google Scholar]
- Niño, S.A.; Chi-Ahumada, E.; Ortíz, J.; Zarazua, S.; Concha, L.; Jiménez-Capdeville, M.E. Demyelination associated with chronic arsenic exposure in Wistar rats. Toxicol. Appl. Pharmacol. 2020, 393, 114955. [Google Scholar] [CrossRef] [PubMed]
- Donofrio, P.D.; Wilbourn, A.J.; Albers, J.W.; Do, L.R.; Salanga, V.; Greenberg, H.S.; Rogers, D.L. Acute arsenic intoxication presenting as guillainbarréa-like syndrome. Muscle Nerve 1987, 10, 114–120. [Google Scholar] [CrossRef] [Green Version]
- Tseng, H.-P.; Wang, Y.-H.; Wu, M.-M.; The, H.-W.; Chiou, H.-Y.; Chen, C.-J. Association between chronic exposure to arsenic and slow nerve conduction velocity among adolescents in Taiwan. J. Health Popul. Nutr. 2006, 24, 182–189. [Google Scholar] [PubMed]
- Chhuttani, P.; Chawla, L.; Sharma, T. Arsenical neuropathy. Neurology 1967, 17, 269. [Google Scholar] [CrossRef]
- Goebel, H.H.; Schmidt, P.F.; Bohl, J.; Tettenborn, B.; Krämer, G.; Gutmann, L. Polyneuropathy Due to Acute Arsenic Intoxication: Biopsy Studies. J. Neuropathol. Exp. Neurol. 1990, 49, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Heyman, A.; Pfeiffer Jr, J.B.; Willett, R.W.; Taylor, H.M. Peripheral neuropathy caused by arsenical intoxication: A study of 41 cases with observations on the effects of BAL (2,3, dimercapto-propanol). N. Engl. J. Med. 1956, 254, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Reichard, J.F.; Puga, A. Effects of arsenic exposure on DNA methylation and epigenetic gene regulation. Epigenomics 2010, 2, 87–104. [Google Scholar] [CrossRef] [Green Version]
- Hamdi, M.; Yoshinaga, M.; Packianathan, C.; Qin, J.; Hallauer, J.; McDermott, J.R.; Yang, H.-C.; Tsai, K.-J.; Liu, Z. Identification of an S-adenosylmethionine (SAM) dependent arsenic methyltransferase in Danio rerio. Toxicol. Appl. Pharmacol. 2012, 262, 185–193. [Google Scholar] [CrossRef] [Green Version]
- Mirbahai, L.; Southam, A.D.; Sommer, U.; Williams, T.D.; Bignell, J.P.; Lyons, B.P.; Viant, M.R.; Chipman, J.K. Disruption of DNA Methylation via S-Adenosylhomocysteine Is a Key Process in High Incidence Liver Carcinogenesis in Fish. J. Proteome Res. 2013, 12, 2895–2904. [Google Scholar] [CrossRef]
- Samikkannu, T.; Chen, C.-H.; Yih, L.-H.; Wang, A.S.S.; Lin, S.-Y.; Chen, T.-C.; Jan, K.-Y. Reactive Oxygen Species Are Involved in Arsenic Trioxide Inhibition of Pyruvate Dehydrogenase Activity. Chem. Res. Toxicol. 2003, 16, 409–414. [Google Scholar] [CrossRef]
- Howe, C.; Gamble, M.V. Influence of Arsenic on Global Levels of Histone Posttranslational Modifications: A Review of the Literature and Challenges in the Field. Curr. Environ. Health Rep. 2016, 3, 225–237. [Google Scholar] [CrossRef] [Green Version]
- Maimaitiyiming, Y.; Wang, Q.Q.; Hsu, C.-H.; Naranmandura, H. Arsenic induced epigenetic changes and relevance to treatment of acute promyelocytic leukemia and beyond. Toxicol. Appl. Pharmacol. 2020, 406, 115212. [Google Scholar] [CrossRef]
- Lee, V.M.-Y.; Goedert, M.; Trojanowski, J.Q. Neurodegenerative Tauopathies. Annu. Rev. Neurosci. 2001, 24, 1121–1159. [Google Scholar] [CrossRef]
- Serrano-Pozo, A.; Frosch, M.P.; Masliah, E.; Hyman, B.T. Neuropathological Alterations in Alzheimer Disease. Cold Spring Harb. Perspect. Med. 2011, 1, a006189. [Google Scholar] [CrossRef]
- Hartl, F.U.; Bracher, A.; Hayer-Hartl, M. Molecular chaperones in protein folding and proteostasis. Nature 2011, 475, 324–332. [Google Scholar] [CrossRef]
- Jacobson, T.; Navarrete, C.; Sharma, S.K.; Sideri, T.C.; Ibstedt, S.; Priya, S.; Grant, C.; Christen, P.; Goloubinoff, P.; Tamás, M.J. Arsenite interferes with protein folding and triggers formation of protein aggregates in yeast. J. Cell Sci. 2012, 125, 5073–5083. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Zhao, P.; Guo, G.; Guo, Y.; Li, S.; He, Y.; Sun, X.; Chai, H.; Zhang, W.; Xing, M. Arsenic Trioxide Exposure Induces Heat Shock Protein Responses in Cock Livers. Biol. Trace Elem. Res. 2015, 170, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Del Razo, L.M.; Quintanilla-Vega, B.; Brambila-Colombres, E.; Aranda, E.S.C.; Manno, M.; Albores, A. Stress Proteins Induced by Arsenic. Toxicol. Appl. Pharmacol. 2001, 177, 132–148. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Xiao, Y.; Wang, Y. SILAC-Based Quantitative Proteomic Analysis Unveils Arsenite-Induced Perturbation of Multiple Pathways in Human Skin Fibroblast Cells. Chem. Res. Toxicol. 2017, 30, 1006–1014. [Google Scholar] [CrossRef] [Green Version]
- He, D.; Xiong, Y.; Wang, L.; Sun, W.; Liu, R.; Yue, T. Arsenic (III) Removal from a High-Concentration Arsenic (III) Solution by Forming Ferric Arsenite on Red Mud Surface. Minerals 2020, 10, 583. [Google Scholar] [CrossRef]
- Pettine, M.; Campanella, L.; Millero, F.J. Arsenite oxidation by H2O2 in aqueous solutions. Geochim. Cosmochim. Acta 1999, 63, 2727–2735. [Google Scholar] [CrossRef]
- Garelick, H.; Dybowska, A.; Valsami-Jones, E.; Priest, N. Remediation Technologies for Arsenic Contaminated Drinking Waters (9 pp). J. Soils Sediments 2005, 5, 182–190. [Google Scholar] [CrossRef]
- Saha, J.; Dikshit, K.; Bandyopadhyay, M. Comparative Studies for Selection of Technologies for Arsenic Removal from Drinking Water. In Proceedings of the BUET-UNU International Workshop on Technologies for Arsenic Removal from Drinking Water, Dhaka, Bangladesh, 5–7 May 2001. [Google Scholar]
- Sancha, A.M. Review of Coagulation Technology for Removal of Arsenic: Case of Chile. J. Health Popul. Nutr. 2006, 24, 267–272. [Google Scholar] [PubMed]
- Elazzouzi, M.; Haboubi, K.; Elyoubi, M. Enhancement of electrocoagulation-flotation process for urban wastewater treatment using Al and Fe electrodes: Techno-economic study. Mater. Today Proc. 2019, 13, 549–555. [Google Scholar] [CrossRef]
- Bazrafshan, E.; Mohammadi, L.; Ansari-Moghaddam, A.; Mahvi, A.H. Heavy metals removal from aqueous environments by electrocoagulation process–a systematic review. J. Environ. Health Sci. Eng. 2015, 13, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, A.E.; Boncukcuoğlu, R.; Kocakerim, M.M. A quantitative comparison between electrocoagulation and chemical coagulation for boron removal from boron-containing solution. J. Hazard. Mater. 2007, 149, 475–481. [Google Scholar] [CrossRef]
- Gupta, R.; Gupta, S.K.; Pathak, D.D. Selective adsorption of toxic heavy metal ions using guanine-functionalized mesoporous silica [SBA-16-g] from aqueous solution. Microporous Mesoporous Mater. 2019, 288, 109577. [Google Scholar] [CrossRef]
- McKay, G.; Porter, J.F.; Prasad, G.R. The Removal of Dye Colours from Aqueous Solutions by Adsorption on Low-cost Materials. Water Air Soil Pollut. 1999, 114, 423–438. [Google Scholar] [CrossRef]
- Faruqi, A.; Henderson, M.; Henderson, R.; Stuetz, R.; Gladman, B.; McDowall, B.; Zamyadi, A. Removal of algal taste and odour compounds by granular and biological activated carbon in full-scale water treatment plants. Water Supply 2018, 18, 1531–1544. [Google Scholar] [CrossRef]
- Samsami, S.; Mohamadizaniani, M.; Sarrafzadeh, M.-H.; Rene, E.R.; Firoozbahr, M. Recent advances in the treatment of dye-containing wastewater from textile industries: Overview and perspectives. Process. Saf. Environ. Prot. 2020, 143, 138–163. [Google Scholar] [CrossRef]
- Majumder, A.; Ramrakhiani, L.; Mukherjee, D.; Mishra, U.; Halder, A.; Mandal, A.K.; Ghosh, S. Green synthesis of iron oxide nanoparticles for arsenic remediation in water and sludge utilization. Clean Technol. Environ. Policy 2019, 21, 795–813. [Google Scholar] [CrossRef]
- Mishra, T.; Mahato, D.K. A comparative study on enhanced arsenic(V) and arsenic(III) removal by iron oxide and manganese oxide pillared clays from ground water. J. Environ. Chem. Eng. 2016, 4, 1224–1230. [Google Scholar] [CrossRef]
- Kundu, S.; Gupta, A. As(III) removal from aqueous medium in fixed bed using iron oxide-coated cement (IOCC): Experimental and modeling studies. Chem. Eng. J. 2007, 129, 123–131. [Google Scholar] [CrossRef]
- Ansari, R.; Sadegh, M. Application of Activated Carbon for Removal of Arsenic Ions from Aqueous Solutions. E-J. Chem. 2007, 4, 103–108. [Google Scholar] [CrossRef] [Green Version]
- Jain, C.; Singh, R. Technological options for the removal of arsenic with special reference to South East Asia. J. Environ. Manag. 2012, 107, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norton, D.M.; Rahman, M.; Shane, A.L.; Hossain, M.Z.; Kulick, R.M.; Bhuiyan, M.I.; Wahed, M.A.; Yunus, M.; Islam, M.S.; Breiman, R.F.; et al. Flocculant-disinfectant point-of-use water treatment for reducing arsenic exposure in rural Bangladesh. Int. J. Environ. Health Res. 2009, 19, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chen, A.S.; Lewis, G.M.; Sorg, T.J.; Supply, W. Arsenic and Uranium Removal from Drinking Water by Adsorptive Media: US EPA Demonstration Project at Upper Bodfish in Lake Isabella, CA: Final Performance Evaluation Report; National Risk Management Research Laboratory, Office of Research and Development, U.S. Environmental Protection Agency: Cincinnati, OH, USA, 2010.
- Bundschuh, J.; Litter, M.; Ciminelli, V.S.; Morgada, M.E.; Cornejo, L.; Hoyos, S.G.; Hoinkis, J.; Alarcón-Herrera, M.T.; Armienta, M.A.; Bhattacharya, P. Emerging mitigation needs and sustainable options for solving the arsenic problems of rural and isolated urban areas in Latin America – A critical analysis. Water Res. 2010, 44, 5828–5845. [Google Scholar] [CrossRef]
- Ahmed, M.F. An Overview of Arsenic Removal Technologies in Bangladesh and India. In Proceedings of the BUET-UNU International Workshop on Technologies for Arsenic Removal from Drinking Water, Dhaka, Bangladesh, 5–7 May 2001; pp. 5–7. [Google Scholar]
- Mondal, S.; Roy, A.; Mukherjee, R.; Mondal, M.; Karmakar, S.; Chatterjee, S.; Mukherjee, M.; Bhattacharjee, S.; De, S. A socio-economic study along with impact assessment for laterite based technology demonstration for arsenic mitigation. Sci. Total Environ. 2017, 583, 142–152. [Google Scholar] [CrossRef]
- Özeler, D.; Yetiş, Ü.; Demirer, G. Life cycle assesment of municipal solid waste management methods: Ankara case study. Environ. Int. 2006, 32, 405–411. [Google Scholar] [CrossRef]
- Garza-Lombo, C.; Pappa, A.; Panayiotidis, M.I.; Gonsebatt, M.E.; Franco, R. Arsenic-induced neurotoxicity: A mechanistic appraisal. JBIC J. Biol. Inorg. Chem. 2019, 24, 1305–1316. [Google Scholar] [CrossRef]
- Abdollahzade, N.; Babri, S.; Majidinia, M. Attenuation of chronic arsenic neurotoxicity via melatonin in male offspring of maternal rats exposed to arsenic during conception: Involvement of oxidative DNA damage and inflammatory signaling cascades. Life Sci. 2020, 266, 118876. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Arsenic | Species | Exposure Duration | Pathological Mechanisms | Toxic Outcome | Ref. |
|---|---|---|---|---|---|
| Sodium arsenite | Rat | 4 months | Increased APP (amyloid precursor protein) and RAGE 9, also increased enzymatic activity of BACE1 (β-secretase). | Neurodegeneration disorders associated with amyloid accumulation. | [2] |
| Sodium arsenite | Rat | 1 month | Increased lipid peroxidation and decrease in nerve conduction velocity. myelin thickness, area, and perimeter of axons. | Impaired central nervous system. | [3] |
| Sodium arsenite | Rat | 9 h | Absence of neurofilament and fibroblast Proteins. | Altered cytoskeletal composition | [4] |
| Sodium arsenite | Rat | 28-days | Reduction in superoxide dismutase-2 and Catalase action in hippocampus, striatum and cortex. | Altered locomotor activity and grip strength. | [5] |
| Sodium arsenite | Rat | 4 months | Elevated oxidative stress, lipid peroxidation and reduced glutathione levels in brain mitochondria. | Increased oxidative stress and mitochondrial damage. | [39] |
| Sodium arsenite | Rat | 28-days | Increased oxidative stress. Decrease in superoxide dismutase-2 activity. | Increased apoptosis in brain cells. | [7] |
| Sodium arsenite | Rat | 10-weeks | Decrease in antioxidative defense mechanisms (GPx, GST, MnSOD, CAT and GR), enhanced LPO observed in the mitochondria at cerebral cortex, cerebellum and hippocampus. | Significant impact on behavioral functions like total locomotor activity, open field behavior, exploratory behavior and grip strength. | [136] |
| Sodium arsenite | Rat | 28-days | Increased oxidative stress in frontal cortex and hippocampus. Increased levels of Nrf2 and HO-1 proteins. | Demise of myelin sheath in neurons and imprecise cristae in the mitochondria both hippocampal and frontal cortex regions. Cholinergic deficits detected. | [137] |
| Sodium arsenite | Rat | 3 months | Biochemical and molecular modifications via inducing oxidative stress and dysfunction of mitochondria. | Mitochondrial decreasing complexes activity and functional impairment. | [31] |
| Sodium arsenite | Rat | 3 months | Reduced NR2A expression in the hippocampus. | Impaired memory. | [10] |
| Sodium arsenite | Rat | 3 months | mGluR5 mRNA and protein expression in hippocampus and cortex. | Learning and memory ability declined. | [11] |
| Sodium arsenite | Rat | 30 days | Lowered expression of NMDAR NR2B subunit and EAAC1 in the brain (hippocampus). | Spatial memory impairment. | [12] |
| Arsenic trioxide | Mice | 45 days | Significant raise in lipid peroxidation, glycogen in cerebral hemisphere and cerebellum. | Neurotoxic effects. | [13] |
| Arsenic trioxide | Mice | 60 days | Reduction of Sdha expression and activity in brain, mitochondrial respiratory chain genes downregulation. | Neurodegeneration disorders. | [14] |
| Arsenic trioxide | T98G and A172 cells | 6, 8 and 24 h | Aggregated mitochondria and MMP dissipation. | Induced apoptosis. | [138] |
| Arsenic trioxide | SY-5Y cells | 24, 48 and 72 h | Elevated intracellular calcium ions. | Increased occurrence of apoptosis and DNA damage. | [139] |
| Sodium arsenite | Primary astrocytes | 24 h | Decreased mitochondrial membrane permeability and decreased protein expression of GLT-1, GS, and GLAST. | Inhibit glutamate metabolism leading to neurotoxicity. | [67] |
| Arsenic trioxide | Rat neuronal cells | 8 h | Increased expression of calpain 1, cdk5, p25 levels. | Induced neuronal cell apoptosis. | [69] |
| Arsenic trioxide | Neuro-2a cells | 24 h | Oxidative stress damage decreased Nrf2 and thioredoxin expression. Mitochondrial dysfunction, PARP activation and caspase cascades, caspase-3 activity. | Neuronal cell death. | [70] |
| Sodium arsenite | Bergmann glial cells | 24 h | Increased EAAT1/GLAST activity and decrease in GLU transport. | Neuronal damage. | [71] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thakur, M.; Rachamalla, M.; Niyogi, S.; Datusalia, A.K.; Flora, S.J.S. Molecular Mechanism of Arsenic-Induced Neurotoxicity including Neuronal Dysfunctions. Int. J. Mol. Sci. 2021, 22, 10077. https://doi.org/10.3390/ijms221810077
Thakur M, Rachamalla M, Niyogi S, Datusalia AK, Flora SJS. Molecular Mechanism of Arsenic-Induced Neurotoxicity including Neuronal Dysfunctions. International Journal of Molecular Sciences. 2021; 22(18):10077. https://doi.org/10.3390/ijms221810077
Chicago/Turabian StyleThakur, Manisha, Mahesh Rachamalla, Som Niyogi, Ashok Kumar Datusalia, and Swaran Jeet Singh Flora. 2021. "Molecular Mechanism of Arsenic-Induced Neurotoxicity including Neuronal Dysfunctions" International Journal of Molecular Sciences 22, no. 18: 10077. https://doi.org/10.3390/ijms221810077
APA StyleThakur, M., Rachamalla, M., Niyogi, S., Datusalia, A. K., & Flora, S. J. S. (2021). Molecular Mechanism of Arsenic-Induced Neurotoxicity including Neuronal Dysfunctions. International Journal of Molecular Sciences, 22(18), 10077. https://doi.org/10.3390/ijms221810077