
Comparative Evaluation of Sucrosomial Iron and Iron Oxide Nanoparticles as Oral Supplements in Iron Deficiency Anemia in Piglets

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
2.1. Piglets Supplemented with Sucrosomial Iron® (SI) but Not Iron Oxide Nanoparticles (IONPs) Show No Evidence of IDA
2.2. Piglets Supplemented with SI but Not IONPs Show Increased Blood Plasma Iron Level Compared to Anemic Animals although Iron Supplementation Does Not Increase Hepatic Iron Content
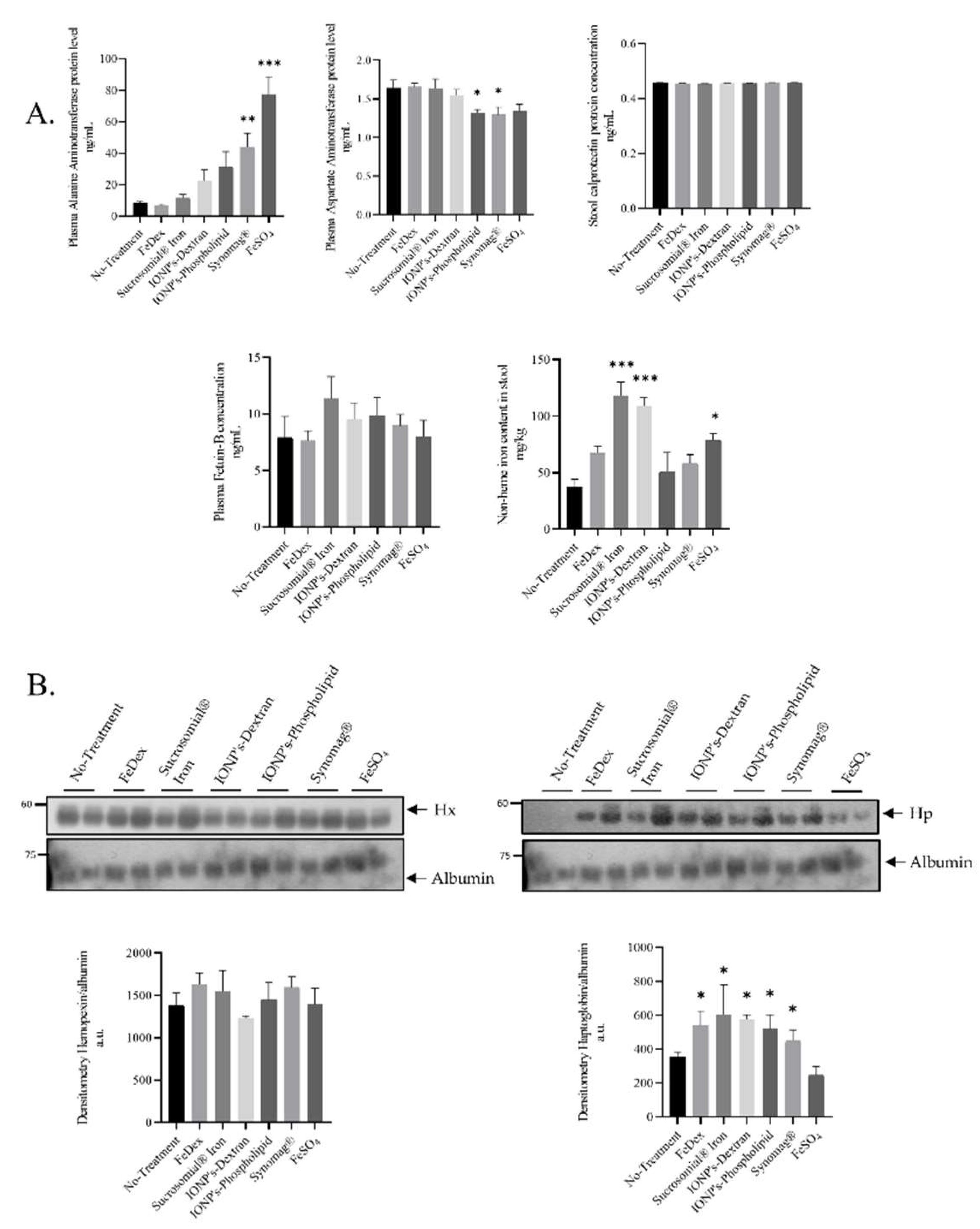
2.3. Toxicological Markers Are Not Altered Neither in SI- Nor IONPs-Supplemented Piglets Compared to Control Animals
2.4. Barely Detectable Concentration of Hepcidin-25 in the Blood Plasma of Piglets Orally Supplemented with SI and IONPs
2.5. FeSO4 but Not Other Oral Iron Supplements Increases Duodenal Hepcidin mRNA Abundance
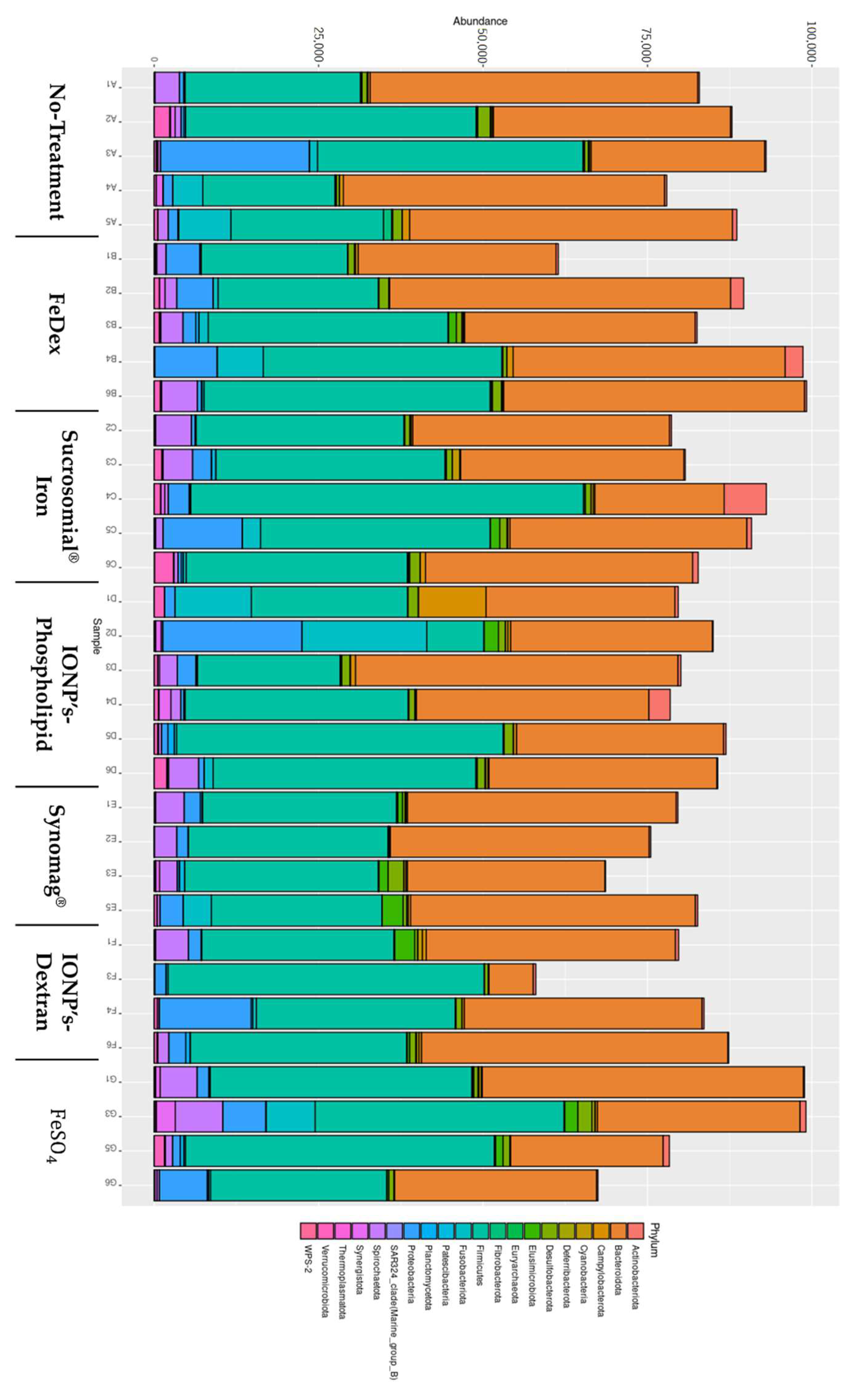
2.6. The Role of Oral Iron Supplementation in Modulation of Piglets Microbiome
3. Discussion
4. Materials and Methods
4.1. Piglets, Experimental Design and Biological Sample Collection
4.2. Measurement of Red Blood Cell Indices and Plasma Iron Parameters
4.3. Measurement of Calprotectin in Feces and Plasma Aspartate Transaminase, Alanine Transaminase and Fetuin-B Levels
4.4. Measurement of Iron Content in Tissues
4.5. Blood Plasma Hepcidin-25 Quantification
4.6. Real-Time Quantitative RT-PCR
4.7. Protein Extract Preparation and Western Blotting
4.8. Statistical Analysis
4.9. Metagenomics Analysis
Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Svoboda, M.; Drabek, J. Iron deficiency in suckling piglets: Etiology, clinical aspects and diagnosis. Folia Vet. 2005, 49, 104–111. [Google Scholar]
- Kim, J.C.; Wilcock, P.; Bedford, M.R. Iron status of piglets and impact of phytase superdosing on iron physiology: A review. Anim. Feed Sci. Technol. 2018, 235, 8–14. [Google Scholar] [CrossRef]
- Szudzik, M.; Starzyński, R.R.; Jończy, A.; Mazgaj, R.; Lenartowicz, M.; Lipiński, P. Iron supplementation in suckling piglets: An ostensibly easy therapy of neonatal iron deficiency anemia. Pharmaceuticals 2018, 11, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egeli, A.K.; Framstad, T. An evaluation of iron-dextran supplementation in piglets administered by injection on the first, third or fourth day after birth. Res. Vet. Sci. 1999, 66, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Ueberschär, S. Sudden death in suckling piglets following administration of iron-dextran-preparation. Dtsch. Tierarztl. Wochenschr. 1966, 73, 145–150. [Google Scholar] [PubMed]
- Bloor, S.R.; Schutte, R.; Hobson, A.R. Oral Iron Supplementation—Gastrointestinal Side Effects and the Impact on the Gut Microbiota. Microbiol. Res. 2021, 12, 33. [Google Scholar] [CrossRef]
- Yilmaz, B.; Li, H. Gut Microbiota and Iron: The Crucial Actors in Health and Disease. Pharmaceuticals 2018, 11, 98. [Google Scholar] [CrossRef] [Green Version]
- Szabo, P.; Bilkei, G. Iron deficiency in outdoor pig production. J. Vet. Med. Ser. A Physiol. Pathol. Clin. Med. 2002, 49, 390–391. [Google Scholar] [CrossRef]
- Svoboda, M.; Píšťková, K. Oral iron administration in suckling piglets—A review. Acta Vet. Brno 2018, 87, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Maes, D.; Steyaert, M.; Vanderhaeghe, C.; López Rodríguez, A.; De Jong, E.; Del Pozo Sacristán, R.; Vangroenweghe, F.; Dewulf, J. Comparison of oral versus parenteral iron supplementation on the health and productivity of piglets. Vet. Rec. 2011, 168, 188. [Google Scholar] [CrossRef]
- López, M.A.A.; Martos, F.C. Iron availability: An updated review. Int. J. Food Sci. Nutr. 2004, 55, 597–606. [Google Scholar] [CrossRef]
- Lipiński, P.; Starzyński, R.R.; Canonne-Hergaux, F.; Tudek, B.; Oliński, R.; Kowalczyk, P.; Dziaman, T.; Thibaudeau, O.; Gralak, M.A.; Smuda, E.; et al. Benefits and Risks of Iron Supplementation in Anemic Neonatal Pigs. Am. J. Pathol. 2010, 177, 1233–1243. [Google Scholar] [CrossRef]
- Krishnan, K.M. Biomedical Nanomagnetics: A Spin Through Possibilities in Imaging, Diagnostics, and Therapy. IEEE Trans. Magn. 2010, 46, 2523–2558. [Google Scholar] [CrossRef] [Green Version]
- Köhler, J.M.; Kluitmann, J.; Knauer, A. Metal Nano Networks by Potential-Controlled In Situ Assembling of Gold/Silver Nanoparticles. Chem. Open 2019, 8, 1369–1374. [Google Scholar]
- Cochran, D.B.; Wattamwar, P.P.; Wydra, R.; Hilt, J.Z.; Anderson, K.W.; Eitel, R.E.; Dziubla, T.D. Suppressing iron oxide nanoparticle toxicity by vascular targeted antioxidant polymer nanoparticles. Biomaterials 2013, 34, 9615–9622. [Google Scholar] [CrossRef] [PubMed]
- Baratli, Y.; Charlesr, L.; Smiri, A.-L.; Wolff, V.; Ben Taha, L.; Bouitbir, J.; Zoll, J.; Piquard, F.; Tebourbi, O.; Sakly, M.; et al. Impact of iron oxide nanoparticles on brain, heart, lung, liver and kidneys mitochondrial respiratory chain complexes activities and coupling. Toxicol. In Vitro 2013, 27, 2142–2148. [Google Scholar] [CrossRef]
- Chamorro, S.; Gutiérrez, L.; Vaquero, M.P.; Verdoy, D.; Salas, G.; Luengo, Y.; Brenes, A.; José Teran, F. Safety assessment of chronic oral exposure to iron oxide nanoparticles. Nanotechnology 2015, 26, 205101. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Ramírez, S.; Brilli, E.; Tarantino, G.; Muñoz, M. Sucrosomial® iron: A new generation iron for improving oral supplementation. Pharmaceuticals 2018, 11, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarantino, G.; Brilli, E.; Giordano, G.; Torelli, A.; Equitani, F. Innovative Oral Iron Supplement (Sucrosomial Iron®) Is Able to Downregulate Hepcidin Release during Inflammation: In Vitro Study. Blood 2015, 126, 4563. [Google Scholar] [CrossRef]
- Szudzik, M.; Lipiński, P.; Jończy, A.; Mazgaj, R.; Pieszka, M.; Kamyczek, M.; Smuda, E.; Starzyński, R.R. Long-term Effect of Split Iron Dextran/Hemoglobin Supplementation on Erythrocyte and Iron Status, Growth Performance, Carcass Parameters, and Meat Quality of Polish Large White and 990 Line Pigs. Biol. Trace Elem. Res. 2020, 196, 472–480. [Google Scholar] [CrossRef] [Green Version]
- Mazgaj, R.; Szudzik, M.; Lipiński, P.; Jończy, A.; Smuda, E.; Kamyczek, M.; Cieślak, B.; Swinkels, D.; Lenartowicz, M.; Starzyński, R.R. Effect of Oral Supplementation of Healthy Pregnant Sows with Sucrosomial Ferric Pyrophosphate on Maternal Iron Status and Hepatic Iron Stores in Newborn Piglets. Animals 2020, 10, 1113. [Google Scholar] [CrossRef] [PubMed]
- Parisi, F.; Berti, C.; Mandò, C.; Martinelli, A.; Mazzali, C.; Cetin, I. Effects of different regimens of iron prophylaxis on maternal iron status and pregnancy outcome: A randomized control trial. J. Matern. Neonatal Med. 2017, 30, 1787–1792. [Google Scholar] [CrossRef] [PubMed]
- Bassols, A.; Costa, C.; Eckersall, P.D.; Osada, J.; Sabrià, J.; Tibau, J. The pig as an animal model for human pathologies: A proteomics perspective. Proteomics Clin. Appl. 2014, 8, 715–731. [Google Scholar] [CrossRef] [PubMed]
- Egeli, A.K.; Framstad, T.; Morberg, H. Clinical Biochemistry, Haematology and Body Weight in Piglets. Acta Vet. Scand. 1998, 39, 381–393. [Google Scholar] [CrossRef] [PubMed]
- Ventrella, D.; Dondi, F.; Barone, F.; Serafini, F.; Elmi, A.; Giunti, M.; Romagnoli, N.; Forni, M.; Bacci, M.L. The biomedical piglet: Establishing reference intervals for haematology and clinical chemistry parameters of two age groups with and without iron supplementation. BMC Vet. Res. 2017, 13, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjarnason, I. The Use of Fecal Calprotectin in Inflammatory Bowel Disease. Gastroenterol. Hepatol. 2017, 13, 53–56. [Google Scholar]
- Bunn, S.K.; Bisset, W.M.; Golden, B.E. Faecal Calprotectin—An Objective Measure of Disease Activity in Childhood Inflammatory Bowel Disease. J. Pediatr. Gastroenterol. Nutr. 1999, 28, 549. [Google Scholar] [CrossRef]
- Olivier, E.; Soury, E.; Ruminy, P.; Husson, A.; Parmentier, F.; Daveau, M.; Salier, J.-P. Fetuin-B, a second member of the fetuin family in mammals. Biochem. J. 2000, 350, 589. [Google Scholar] [CrossRef]
- Giboney, P.T. Mildly elevated liver transaminase levels in the asymptomatic patient. Am. Fam. Physician 2005, 71, 1105–1110. [Google Scholar]
- Higashisaka, K.; Yoshioka, Y.; Yamashita, K.; Morishita, Y.; Fujimura, M.; Nabeshi, H.; Nagano, K.; Abe, Y.; Kamada, H.; Tsunoda, S.; et al. Acute phase proteins as biomarkers for predicting the exposure and toxicity of nanomaterials. Biomaterials 2011, 32, 3–9. [Google Scholar] [CrossRef]
- Nagano, T.; Yoshioka, Y.; Higashisaka, K.; Kunieda, A.; Hata, K.; Nagano, K.; Abe, Y.; Kamada, H.; Tsunoda, S.; Nabeshi, H.; et al. Potential of acute-phase proteins as biomarkers for sub-nano platinum exposure. Pharmazie 2012, 67, 958–959. [Google Scholar]
- Muckenthaler, M.U.; Rivella, S.; Hentze, M.W.; Galy, B. A Red Carpet for Iron Metabolism. Cell 2017, 168, 344–361. [Google Scholar] [CrossRef] [Green Version]
- Lipiński, P.; Styś, A.; Starzyński, R.R. Molecular insights into the regulation of iron metabolism during the prenatal and early postnatal periods. Cell. Mol. Life Sci. 2013, 70, 23–38. [Google Scholar] [CrossRef] [Green Version]
- Staroń, R.; Van Swelm, R.P.L.; Lipiński, P.; Gajowiak, A.; Lenartowicz, M.; Bednarz, A.; Gajewska, M.; Pieszka, M.; Laarakkers, C.M.M.; Swinkels, D.W.; et al. Urinary hepcidin levels in iron-deficient and iron-supplemented piglets correlate with hepcidin hepatic mRNA and serum levels and with body iron status. PLoS ONE 2015, 10, 1–12. [Google Scholar]
- Staron, R.; Lipinski, P.; Lenartowicz, M.; Bednarz, A.; Gajowiak, A.; Smuda, E.; Krzeptowski, W.; Pieszka, M.; Korolonek, T.; Hamza, I.; et al. Dietary hemoglobin rescues young piglets from severe iron deficiency anemia: Duodenal expression profile of genes involved in heme iron absorption. PLoS ONE 2017, 12, 1–22. [Google Scholar] [CrossRef]
- Bessman, N.J.; Mathieu, J.R.R.; Renassia, C.; Zhou, L.; Fung, T.C.; Fernandez, K.C.; Austin, C.; Moeller, J.B.; Zumerle, S.; Louis, S.; et al. Dendritic cell–derived hepcidin sequesters iron from the microbiota to promote mucosal healing. Science 2020, 368, 186–189. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.C.; Mahan, D.C. Effects of neonatal iron status, iron injections at birth, and weaning in young pigs from sows fed either organic or inorganic trace minerals. J. Anim. Sci. 2008, 86, 2261–2269. [Google Scholar] [CrossRef]
- Gulec, S.; Anderson, G.J.; Collins, J.F. Mechanistic and regulatory aspects of intestinal iron absorption. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 307, G397–G409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Y.; Tang, H.; Radosz, M.; Van Kirk, E.; Murdoch, W.J. pH-Responsive Nanoparticles for Cancer Drug Delivery. In Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2008; Volume 437, pp. 183–216. ISBN 9781588298911. [Google Scholar]
- Neuberger, T.; Schöpf, B.; Hofmann, H.; Hofmann, M.; von Rechenberg, B. Superparamagnetic nanoparticles for biomedical applications: Possibilities and limitations of a new drug delivery system. J. Magn. Magn. Mater. 2005, 293, 483–496. [Google Scholar] [CrossRef]
- Thakkar, H.; Patel, B.; Thakkar, S. A review on techniques for oral bioavailability enhancement of drugs. Int. J. Pharm. Sci. Rev. Res. 2010, 4, 203–223. [Google Scholar]
- Hilty, F.M.; Arnold, M.; Hilbe, M.; Teleki, A.; Knijnenburg, J.T.N.; Ehrensperger, F.; Hurrell, R.F.; Pratsinis, S.E.; Langhans, W.; Zimmermann, M.B. Iron from nanocompounds containing iron and zinc is highly bioavailable in rats without tissue accumulation. Nat. Nanotechnol. 2010, 5, 374–380. [Google Scholar] [CrossRef]
- Alavi, M.; Karimi, N.; Safaei, M. Application of Various Types of Liposomes in Drug Delivery Systems. Adv. Pharm. Bull. 2017, 7, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M.B. The potential of encapsulated iron compounds in food fortification: A review. Int. J. Vitam. Nutr. Res. 2004, 74, 453–461. [Google Scholar] [CrossRef]
- Pereira, D.I.A.; Aslam, M.F.; Frazer, D.M.; Schmidt, A.; Walton, G.E.; Mccartney, A.L.; Gibson, G.R.; Anderson, G.J.; Powell, J.J. Dietary iron depletion at weaning imprints low microbiome diversity and this is not recovered with oral nano Fe(III). Microbiologyopen 2015, 4, 12–27. [Google Scholar] [CrossRef] [PubMed]
- Churio, O.; Durán, E.; Guzmán-Pino, S.A.; Valenzuela, C. Use of encapsulation technology to improve the efficiency of an iron oral supplement to prevent anemia in suckling pigs. Animals 2019, 9, 1. [Google Scholar] [CrossRef] [Green Version]
- Wegmüller, R.; Zimmermann, M.B.; Moretti, D.; Arnold, M.; Langhans, W.; Hurrell, R.F. Particle Size Reduction and Encapsulation Affect the Bioavailability of Ferric Pyrophosphate in Rats. J. Nutr. 2004, 134, 3301–3304. [Google Scholar] [CrossRef] [Green Version]
- Shafie, E.; Keshavarz, S.; Kefayati, M.; Taheri, F.; Sarbakhsh, P.; Vafa, M. The effects of nanoparticles containing iron on blood and inflammatory markers in comparison to ferrous sulfate in anemic rats. Int. J. Prev. Med. 2016, 7, 117. [Google Scholar]
- Elsayed, H.H.; Al-Sherbini, A.S.A.M.; Abd-Elhady, E.E.; Ahmed, K.A.E.A. Treatment of Anemia Progression via Magnetite and Folate Nanoparticles In Vivo. ISRN Nanotechnol. 2014, 2014, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Hosseindoust, A.; Kim, M.; Kim, K.; Choi, Y.; Moturi, J.; Song, C.; Lee, S.; Cho, H.; Chae, B. Effects of hot melt extrusion processed nano-iron on growth performance, blood composition, and iron bioavailability in weanling pigs. J. Anim. Sci. Technol. 2019, 61, 216–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geisser, P.; Baer, M.; Schaub, E. Structure/histotoxicity relationship of parenteral iron preparations. Arzneimittelforschung 1992, 42, 1439–1452. [Google Scholar] [PubMed]
- Rausa, M.; Pagani, A.; Nai, A.; Campanella, A.; Gilberti, M.E.; Apostoli, P.; Camaschella, C.; Silvestri, L. Bmp6 Expression in Murine Liver Non Parenchymal Cells: A Mechanism to Control their High Iron Exporter Activity and Protect Hepatocytes from Iron Overload? PLoS ONE 2015, 10, e0122696. [Google Scholar] [CrossRef] [PubMed]
- Pigeon, C.; Ilyin, G.; Courselaud, B.; Leroyer, P.; Turlin, B.; Brissot, P.; Loréal, O. A New Mouse Liver-specific Gene, Encoding a Protein Homologous to Human Antimicrobial Peptide Hepcidin, Is Overexpressed during Iron Overload. J. Biol. Chem. 2001, 276, 7811–7819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, L.; Valore, E.V.; Nemeth, E.; Goodnough, J.B.; Gabayan, V.; Ganz, T. Iron transferrin regulates hepcidin synthesis in primary hepatocyte culture through hemojuvelin and BMP2/4. Blood 2007, 110, 2182–2189. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Wang, J.; Liu, J.; Wang, H.; Hillyer, C.D.; Blanc, L.; An, X.; Mohandas, N. Dynamic changes in murine erythropoiesis from birth to adulthood: Implications for the study of murine models of anemia. Blood Adv. 2021, 5, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Papanikolaou, G.; Pantopoulos, K. Systemic iron homeostasis and erythropoiesis. IUBMB Life 2017, 69, 399–413. [Google Scholar] [CrossRef] [Green Version]
- Qian, Q.; Nath, K.A.; Wu, Y.; Daoud, T.M.; Sethi, S. Hemolysis and acute kidney failure. Am. J. Kidney Dis. 2010, 56, 780–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geppert, M.; Hohnholt, M.C.; Nürnberger, S.; Dringen, R. Ferritin up-regulation and transient ROS production in cultured brain astrocytes after loading with iron oxide nanoparticles. Acta Biomater. 2012, 8, 3832–3839. [Google Scholar] [CrossRef] [PubMed]
- Levy, M.; Luciani, N.; Alloyeau, D.; Elgrabli, D.; Deveaux, V.; Pechoux, C.; Chat, S.; Wang, G.; Vats, N.; Gendron, F.; et al. Long term in vivo biotransformation of iron oxide nanoparticles. Biomaterials 2011, 32, 3988–3999. [Google Scholar] [CrossRef]
- Ebert, T.; Linder, N.; Schaudinn, A.; Busse, H.; Berger, J.; Lichtinghagen, R.; Keim, V.; Wiegand, J.; Karlas, T. Association of fetuin B with markers of liver fibrosis in nonalcoholic fatty liver disease. Endocrine 2017, 58, 246–252. [Google Scholar] [CrossRef]
- Park, E.J.; Kim, H.; Kim, Y.; Yi, J.; Choi, K.; Park, K. Inflammatory responses may be induced by a single intratracheal instillation of iron nanoparticles in mice. Toxicology 2010, 275, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Tolosano, E.; Altruda, F. Hemopexin: Structure, Function, and Regulation. DNA Cell Biol. 2002, 21, 297–306. [Google Scholar] [CrossRef]
- Fabiano, A.; Brilli, E.; Mattii, L.; Testai, L.; Moscato, S.; Citi, V.; Tarantino, G.; Zambito, Y. Ex vivo and in vivo study of sucrosomial® iron intestinal absorption and bioavailability. Int. J. Mol. Sci. 2018, 19, 2722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaeggi, T.; Kortman, G.A.M.; Moretti, D.; Chassard, C.; Holding, P.; Dostal, A.; Boekhorst, J.; Timmerman, H.M.; Swinkels, D.W.; Tjalsma, H.; et al. Iron fortification adversely affects the gut microbiome, increases pathogen abundance and induces intestinal inflammation in Kenyan infants. Gut 2015, 64, 731–742. [Google Scholar] [CrossRef] [PubMed]
- Mabbott, N.A.; Donaldson, D.S.; Ohno, H.; Williams, I.R.; Mahajan, A. Microfold (M) cells: Important immunosurveillance posts in the intestinal epithelium. Mucosal Immunol. 2013, 6, 666–677. [Google Scholar] [CrossRef] [Green Version]
- Medina-Reyes, E.I.; Rodríguez-Ibarra, C.; Déciga-Alcaraz, A.; Díaz-Urbina, D.; Chirino, Y.I.; Pedraza-Chaverri, J. Food additives containing nanoparticles induce gastrotoxicity, hepatotoxicity and alterations in animal behavior: The unknown role of oxidative stress. Food Chem. Toxicol. 2020, 146, 111814. [Google Scholar] [CrossRef]
- Perez-Muñoz, M.E.; Arrieta, M.-C.; Ramer-Tait, A.E.; Walter, J. A critical assessment of the “sterile womb” and “in utero colonization” hypotheses: Implications for research on the pioneer infant microbiome. Microbiome 2017, 5, 48. [Google Scholar] [CrossRef] [PubMed]
- Frese, S.A.; Parker, K.; Calvert, C.C.; Mills, D.A. Diet shapes the gut microbiome of pigs during nursing and weaning. Microbiome 2015, 3, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mach, N.; Berri, M.; Estellé, J.; Levenez, F.; Lemonnier, G.; Denis, C.; Leplat, J.-J.; Chevaleyre, C.; Billon, Y.; Doré, J.; et al. Early-life establishment of the swine gut microbiome and impact on host phenotypes. Environ. Microbiol. Rep. 2015, 7, 554–569. [Google Scholar] [CrossRef]
- Han, G.G.; Lee, J.-Y.; Jin, G.-D.; Park, J.; Choi, Y.H.; Kang, S.-K.; Chae, B.J.; Kim, E.B.; Choi, Y.-J. Tracing of the fecal microbiota of commercial pigs at five growth stages from birth to shipment. Sci. Rep. 2018, 8, 6012. [Google Scholar] [CrossRef] [Green Version]
- Mariat, D.; Firmesse, O.; Levenez, F.; Guimarăes, V.; Sokol, H.; Doré, J.; Corthier, G.; Furet, J.-P. The Firmicutes/Bacteroidetes ratio of the human microbiota changes with age. BMC Microbiol. 2009, 9, 123. [Google Scholar] [CrossRef]
- Macphail, A.P.; Charlton, R.W.; Bothwell, T.H.; Torrance, J.D. The Relationship between Maternal and Infant Iron Status. Scand. J. Haematol. 1981, 25, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Starzyński, R.R.; Laarakkers, C.M.M.; Tjalsma, H.; Swinkels, D.W.; Pieszka, M.; Styś, A.; Mickiewicz, M.; Lipiński, P. Iron Supplementation in Suckling Piglets: How to Correct Iron Deficiency Anemia without Affecting Plasma Hepcidin Levels. PLoS ONE 2013, 8, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laarakkers, C.M.M.; Wiegerinck, E.T.; Klaver, S.; Kolodziejczyk, M.; Gille, H.; Hohlbaum, A.M.; Tjalsma, H.; Swinkels, D.W. Improved Mass Spectrometry Assay For Plasma Hepcidin: Detection and Characterization of a Novel Hepcidin Isoform. PLoS ONE 2013, 8, e75518. [Google Scholar]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of Real-Time Quantitative Reverse Transcription-PCR Data: A Model-Based Variance Estimation Approach to Identify Genes Suited for Normalization, Applied to Bladder and Colon Cancer Data Sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [Green Version]
- Canonne-Hergaux, F.; Gruenheid, S.; Ponka, P.; Gros, P. Cellular and Subcellular Localization of the Nramp2 Iron Transporter in the Intestinal Brush Border and Regulation by Dietary Iron. Blood 1999, 93, 4406–4417. [Google Scholar] [CrossRef] [PubMed]
- Canonne-Hergaux, F.; Fleming, M.D.; Levy, J.E.; Gauthier, S.; Ralph, T.; Picard, V.; Andrews, N.C.; Gros, P. The Nramp2/DMT1 iron transporter is induced in the duodenum of microcytic anemia mk mice but is not properly targeted to the intestinal brush border. Blood 2000, 96, 3964–3970. [Google Scholar]
- Draper, H.H.; Squires, E.J.; Mahmoodi, H.; Wu, J.; Agarwal, S.; Hadley, M. A comparative evaluation of thiobarbituric acid methods for the determination of malondialdehyde in biological materials. Free Radic. Biol. Med. 1993, 15, 353–363. [Google Scholar] [CrossRef]
- Rstudio T. RStudio: Integrated Development for R. Rstudio Team, PBC: Boston, MA, USA, 2020. Available online: http//www.rstudio.com (accessed on 11 August 2021).
- CoreTeam, R. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Volume 2, ISBN 3900051070. [Google Scholar]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Ma, S.; Shungin, D.; Mallick, H.; Schirmer, M.; Nguyen, L.H.; Kolde, R.; Franzosa, E.; Vlamakis, H.; Xavier, R.; Huttenhower, C. Population Structure Discovery in Meta-Analyzed Microbial Communities and Inflammatory Bowel Disease. bioRxiv 2020. [Google Scholar] [CrossRef]
- Willis, A.D.; Martin, B.D. Estimating diversity in networked ecological communities. Biostatistics 2020, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Willis, A.; Bunge, J.; Whitman, T. Improved detection of changes in species richness in high diversity microbial communities. J. R. Stat. Soc. Ser. C 2017, 66, 963–977. [Google Scholar] [CrossRef] [Green Version]
- Willis, A.; Bunge, J. Estimating diversity via frequency ratios. Biometrics 2015, 71, 1042–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martino, C.; Morton, J.T.; Marotz, C.A.; Thompson, L.R.; Tripathi, A.; Knight, R.; Zengler, K. A Novel Sparse Compositional Technique Reveals Microbial Perturbations. mSystems 2019, 4, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Mcglinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. vegan: Community Ecology Package. R package version 2.4-2. Community Ecol. Package 2019, 2, 5–6. [Google Scholar]
- Martinez Arbizu, P. pairwiseAdonis: Pairwise multilevel comparison using adonis. R Package Version 0.0 2017. Available online: https://github.com/pmartinezarbizu/pairwiseAdonis (accessed on 11 August 2021).
- De Vries, A.; Ripley, B.D. Package “ggdendro”: Create Dendrograms and Tree Diagrams Using “ggplot2”. R Package Version 0.1-20 2016. Available online: https://cran.r-project.org/web/packages/ggdendro/index.html (accessed on 11 August 2021).
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Weiss, S.; Xu, Z.Z.; Peddada, S.; Amir, A.; Bittinger, K.; Gonzalez, A.; Lozupone, C.; Zaneveld, J.R.; Vázquez-Baeza, Y.; Birmingham, A.; et al. Normalization and microbial differential abundance strategies depend upon data characteristics. Microbiome 2017, 5, 27. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Groups | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| No Treatment | FeDex | Sucrosomial® Iron | IONP’s Phospholipid | Synomag® | IONP’s Dextran | F | p | ||
| RBC (×106/µL) | 3.96 | 6.35 *** | 6.06 ** | 5.84 ** | 5.31 | 4.86 | 5.51 * | 4.20 | 0.0028 |
| RBC-He (pg) | 11.2 | 14.3 ** | 13.0 | 11.0 | 11.4 | 10.9 | 11.9 | 5.66 | <0.0001 |
| HGB (g/dL) | 4.40 | 9.15 *** | 7.88 *** | 6.42 | 6.07 | 5.37 | 6.58 * | 8.23 | <0.0001 |
| HCT (%) | 12.7 | 29.2 *** | 24.8 *** | 20.4 * | 19.5 | 16.6 | 21.4 * | 8.10 | <0.0001 |
| MCV (fL) | 32.0 | 45.8 *** | 41.1 * | 34.9 | 36.7 | 34.2 | 38.6 | 5.87 | 0.0003 |
| RDW (%) | 46.6 | 35.7 * | 37.3 * | 46.2 | 45.4 | 45.2 | 41.1 | 3.76 | 0.0056 |
| MCH (pg) | 11.1 | 14.4 *** | 13.1 * | 11.0 | 11.4 | 11.0 | 12.1 | 7.49 | <0.001 |
| RET (K/uL) | 18.2 | 56.9 | 115 ** | 8.97 | 18.7 | 10.9 | 36.4 | 5.59 | <0.001 |
| RET-He (pg) | 15.6 | 14.6 | 16.1 | 18.6 | 17.4 | 13.9 | 13.4 | 1.24 | 0.310 |
| Tissue | Groups | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Parameter | No Treatment | FeDex | Sucrosomial® Iron | IONP’s Phospholipid | Synomag® | IONP’s Dextran | F | p | ||
| Non-heme iron level (mg Fe/kg of wet tissue) | liver | 35.6 | 112 *** | 32.4 | 22.9 | 25.0 | 29.0 | 33.6 | 15.2 | < 0.0001 |
| spleen | 94.8 | 46.4 * | 51.9 * | 65.9 | 57.8 | 57.1 | 52.1 | 2.01 | 0.0927 | |
| Plasma iron level (ug/dL) | 18.3 | 35.1 * | 31.1 * | 20.1 | 14.9 | 12.9 | 20.9 | 1.61 | 0.1779 | |
| TIBC (ug/dL) | 403 | 350 | 365 | 354 | 363 | 448 | 338 | 0.867 | 0.5291 | |
| TSAT % | 19.2 | 34.4 | 23.5 | 27.2 | 27.5 | 20.8 | 30.4 | 1.20 | 0.3297 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazgaj, R.; Lipiński, P.; Szudzik, M.; Jończy, A.; Kopeć, Z.; Stankiewicz, A.M.; Kamyczek, M.; Swinkels, D.; Żelazowska, B.; Starzyński, R.R. Comparative Evaluation of Sucrosomial Iron and Iron Oxide Nanoparticles as Oral Supplements in Iron Deficiency Anemia in Piglets. Int. J. Mol. Sci. 2021, 22, 9930. https://doi.org/10.3390/ijms22189930
Mazgaj R, Lipiński P, Szudzik M, Jończy A, Kopeć Z, Stankiewicz AM, Kamyczek M, Swinkels D, Żelazowska B, Starzyński RR. Comparative Evaluation of Sucrosomial Iron and Iron Oxide Nanoparticles as Oral Supplements in Iron Deficiency Anemia in Piglets. International Journal of Molecular Sciences. 2021; 22(18):9930. https://doi.org/10.3390/ijms22189930
Chicago/Turabian StyleMazgaj, Rafał, Paweł Lipiński, Mateusz Szudzik, Aneta Jończy, Zuzanna Kopeć, Adrian M. Stankiewicz, Marian Kamyczek, Dorine Swinkels, Beata Żelazowska, and Rafał R. Starzyński. 2021. "Comparative Evaluation of Sucrosomial Iron and Iron Oxide Nanoparticles as Oral Supplements in Iron Deficiency Anemia in Piglets" International Journal of Molecular Sciences 22, no. 18: 9930. https://doi.org/10.3390/ijms22189930
APA StyleMazgaj, R., Lipiński, P., Szudzik, M., Jończy, A., Kopeć, Z., Stankiewicz, A. M., Kamyczek, M., Swinkels, D., Żelazowska, B., & Starzyński, R. R. (2021). Comparative Evaluation of Sucrosomial Iron and Iron Oxide Nanoparticles as Oral Supplements in Iron Deficiency Anemia in Piglets. International Journal of Molecular Sciences, 22(18), 9930. https://doi.org/10.3390/ijms22189930