
Pyrimidine Biosynthetic Enzyme CAD: Its Function, Regulation, and Diagnostic Potential


Abstract
:1. Introduction
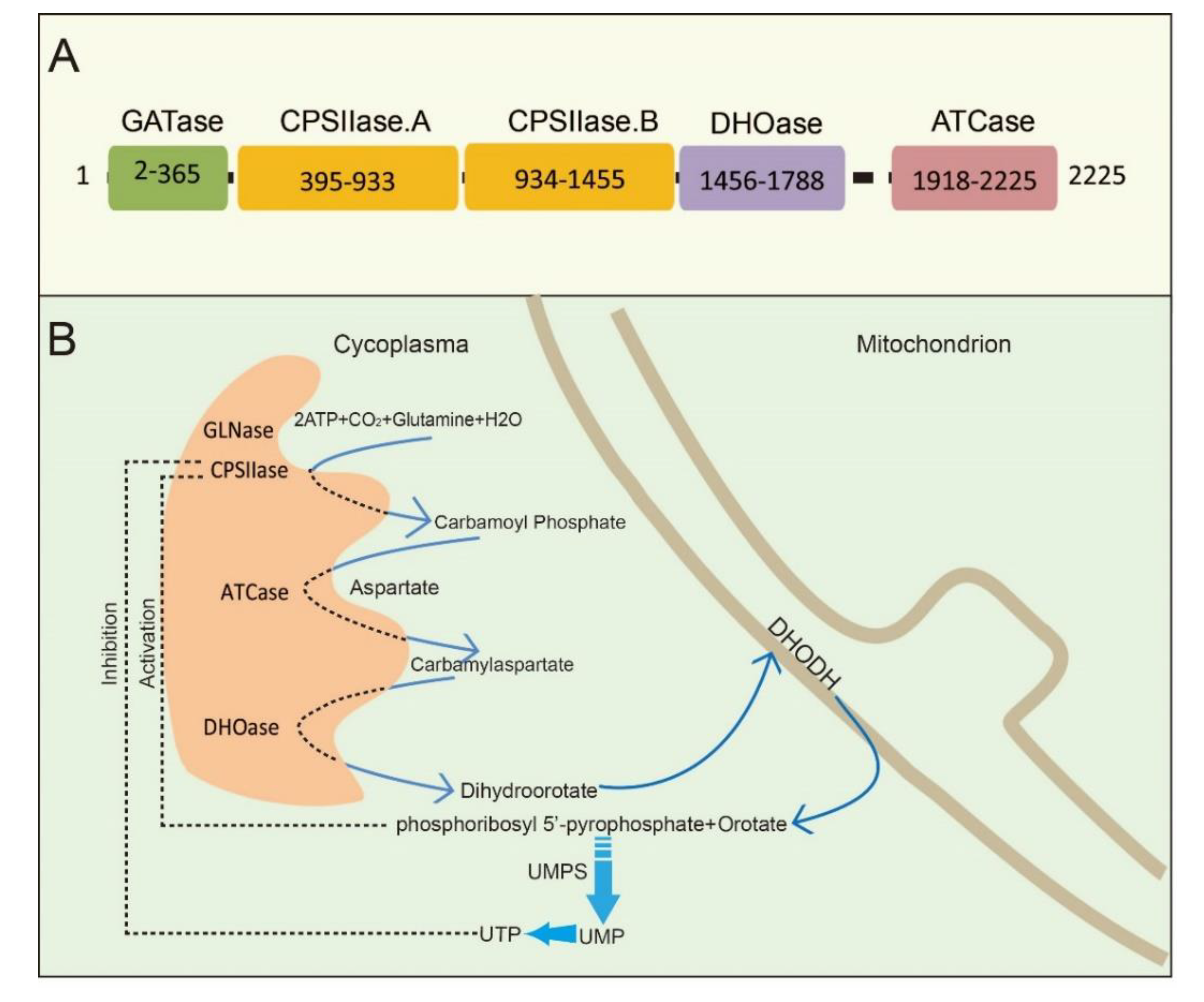
2. CAD Structure and Function
2.1. Overall Structure and Function of CAD
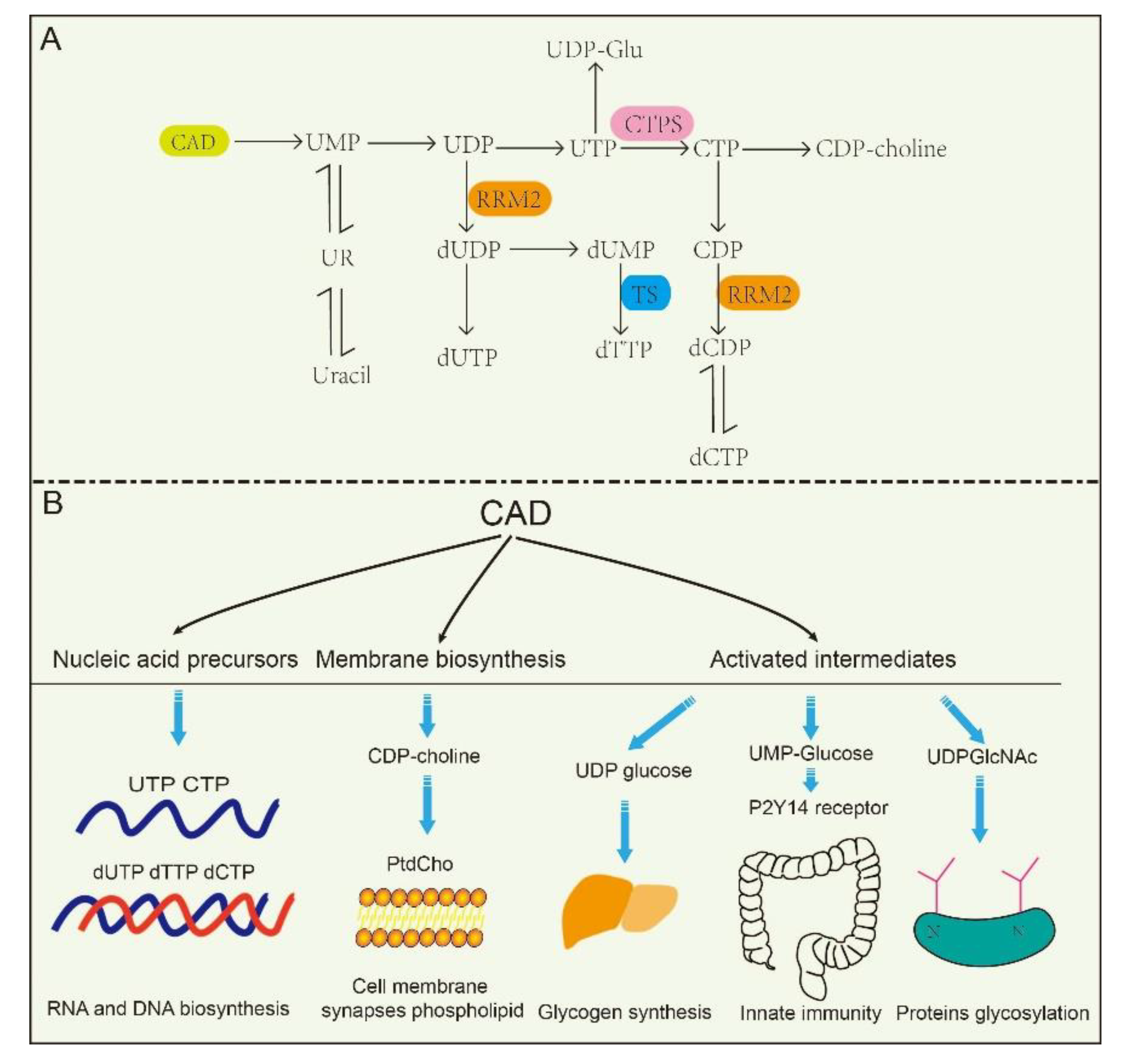
2.2. CAD Participates in Pyrimidine Nucleotide Biochemistry and Metabolism
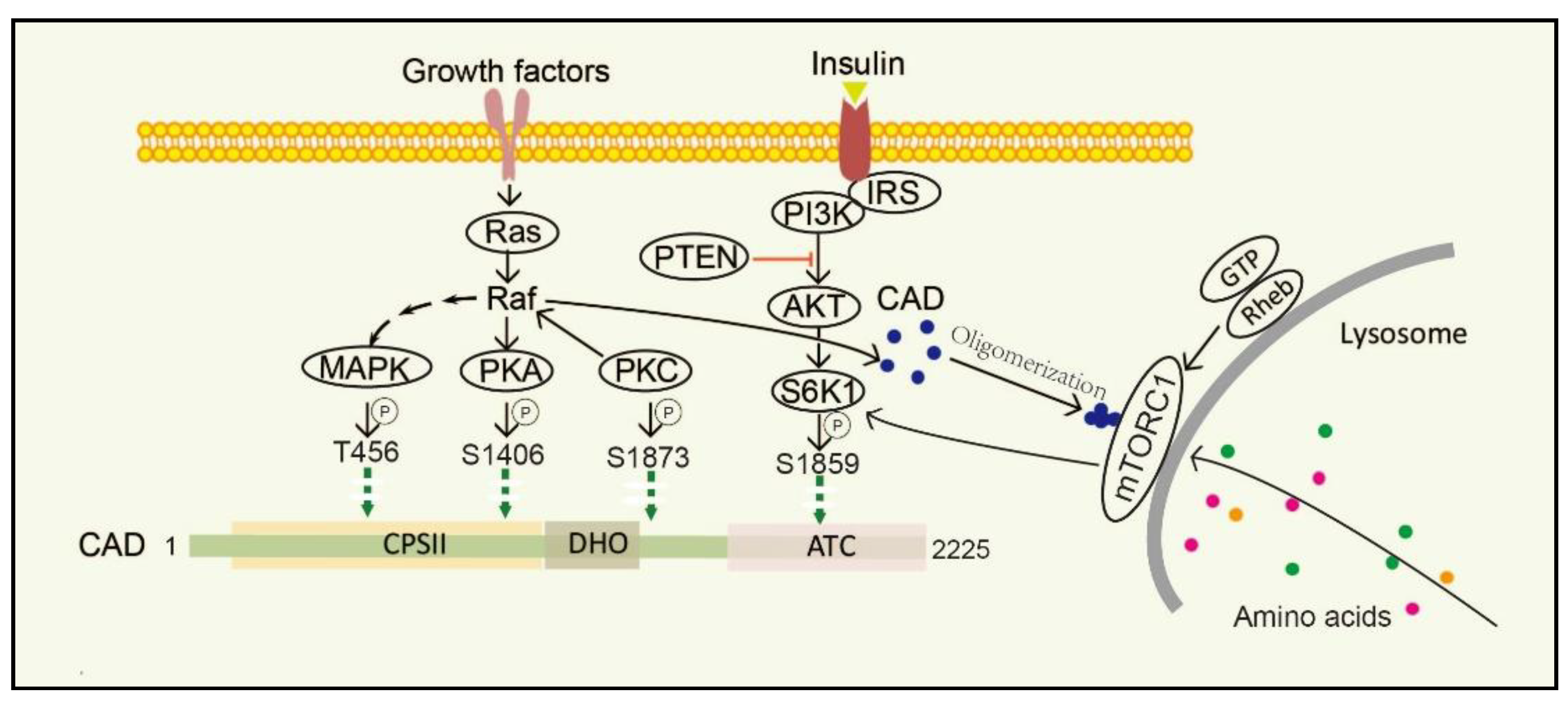
3. The Regulation of CAD
3.1. MAPK/cAMP-Dependent PKA/PKC Pathway
3.2. PI3K-AKT-mTORC1-S6K1 Pathway
4. Implications for Therapy of CAD-Related Diseases
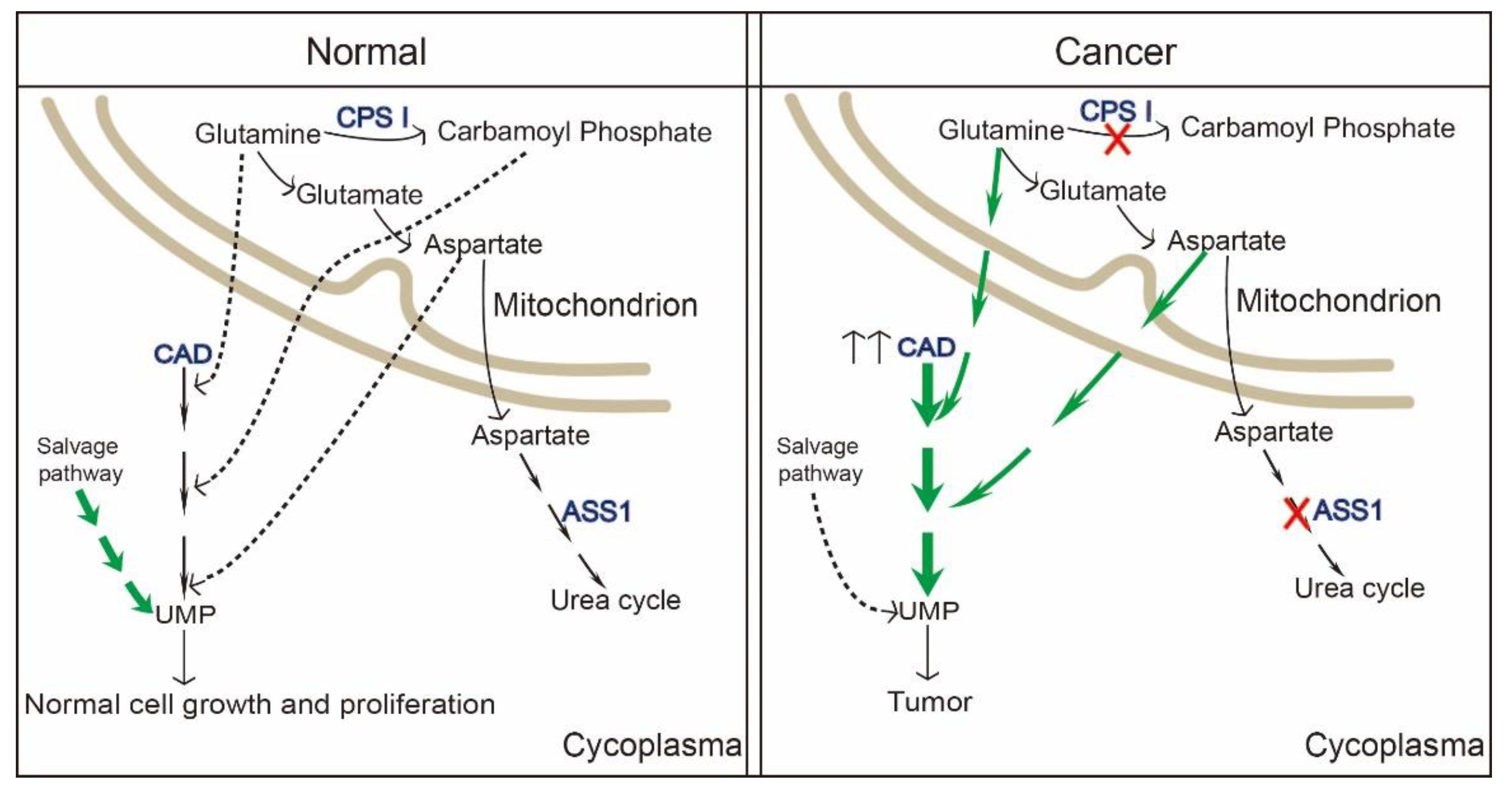
4.1. CAD and Tumors
4.2. CAD and Inherited Metabolic Diseases
4.3. CAD and Immunity
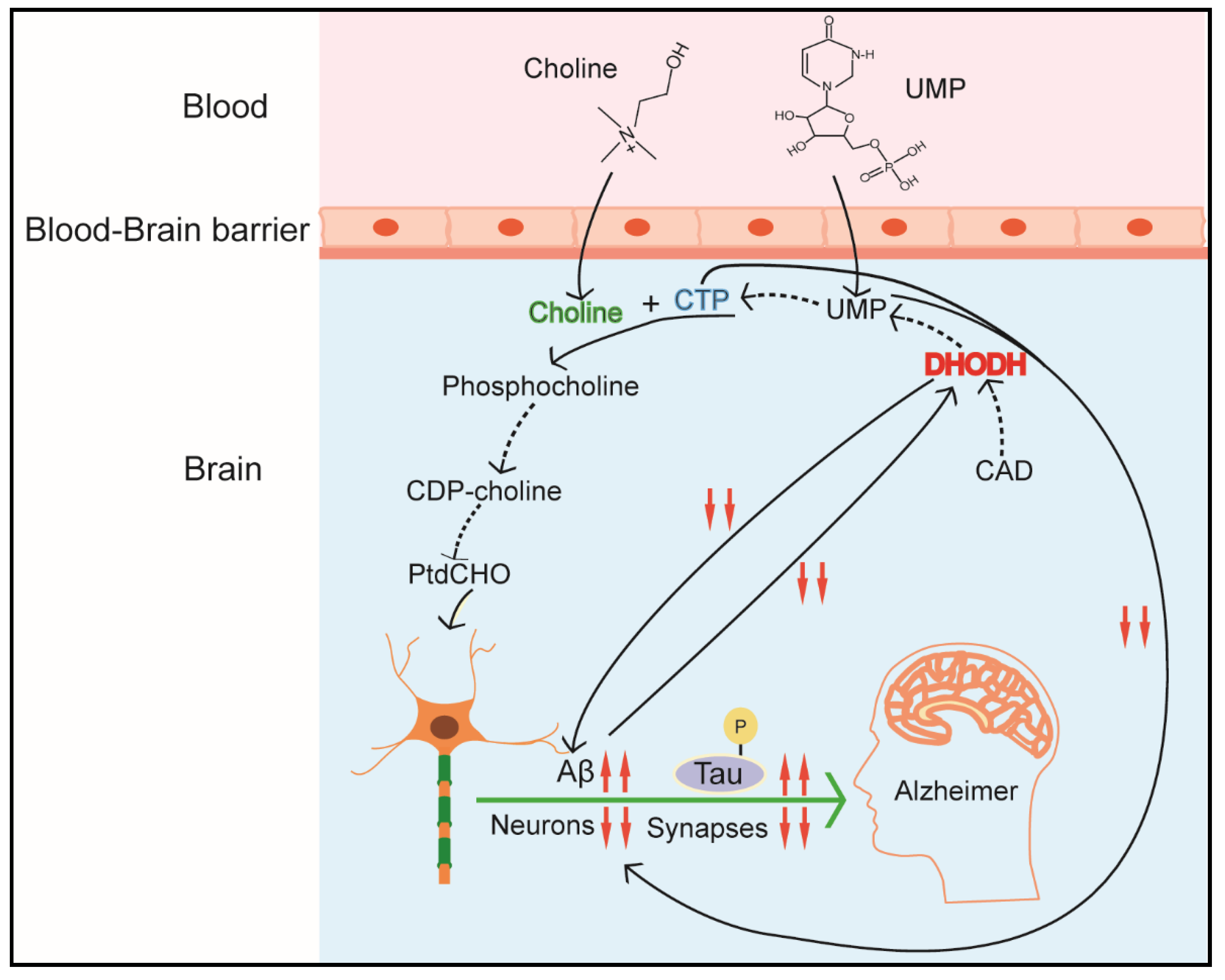
4.4. CAD and Neurological Disorders
5. Perspectives and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jones, M.E.; Spector, L.; Lipmann, F. CARBAMYL PHOSPHATE, THE CARBAMYL DONOR IN ENZYMATIC CITRULLINE SYNTHESIS1. J. Am. Chem. Soc. 1955, 77, 819–820. [Google Scholar] [CrossRef]
- Lacroute, F.; Pierard, A.; Grenson, M.; Wiame, J.M. The Biosynthesis of Carbamoyl Phosphate in Saccharomyces cerevisiae. J. Gen. Microbiol. 1965, 40, 127–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, M.E. Carbamyl Phosphate: Many forms of life use this molecule to synthesize arginine, uracil, and adenosine triphosphate. Science 1963, 140, 1373–1379. [Google Scholar] [CrossRef]
- Pierard, A.; Wiame, J.M. Regulation and mutation affecting a glutamine dependent formation of carbamyl phosphate in Escherichia coli. Biochem. Biophys. Res. Commun. 1964, 15, 76–81. [Google Scholar] [CrossRef]
- Issaly, I.M.; Issaly, A.S.; Reissig, J.L. Carbamyl phosphate biosynthesis in Bacillus subtilis. Biochim. Biophys. Acta Enzym. 1970, 198, 482–494. [Google Scholar] [CrossRef]
- Jones, M.E. Pyrimidine Nucleotide Biosynthesis in Animals: Genes, Enzymes, and Regulation of UMP Biosynthesis. Annu. Rev. Biochem. 1980, 49, 253–279. [Google Scholar] [CrossRef] [PubMed]
- Hoogenraad, N.J.; Levine, R.L.; Kretchmer, N. Copurification of carbamoyl phosphate synthetase and aspartate transcarbamoylase from mouse spleen. Biochem. Biophys. Res. Commun. 1971, 44, 981–988. [Google Scholar] [CrossRef]
- Shoaf, W.T.; Jones, M.E. Initial steps in pyrimidine synthesis in ehrlich ascites carcinoma. Biochem. Biophys. Res. Commun. 1971, 45, 796–802. [Google Scholar] [CrossRef]
- Novak, D.A.; Carver, J.D.; Barness, L.A. Dietary Nucleotides Affect Hepatic Growth and Composition in the Weanling Mouse. J. Parenter. Enter. Nutr. 1994, 18, 62–66. [Google Scholar] [CrossRef]
- Pluske, J.R.; Fenton, T.W.; Lorschy, M.L.; Pettigrew, J.E.; Sower, A.F.; Aherne, F.X. A modification to the isotope-dilution technique for estimating milk intake of pigs using pig serum. J. Anim. Sci. 1997, 75, 1279–1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- del Caño-Ochoa, F.; Moreno-Morcillo, M.; Ramón-Maiques, S. CAD, A Multienzymatic Protein at the Head of de Novo Pyrimidine Biosynthesis. Macromol. Protein Complexes II Struct. Funct. 2019, 93, 505–538. [Google Scholar] [CrossRef]
- Tu, H.-F.; Ko, C.-J.; Lee, C.-T.; Lee, C.-F.; Lan, S.-W.; Lin, H.-H.; Ku, C.-C.; Lee, D.-Y.; Chen, I.-C.; Chuang, Y.-H.; et al. Afatinib Exerts Immunomodulatory Effects by Targeting the Pyrimidine Biosynthesis Enzyme CAD. Cancer Res. 2021, 81, 3270–3282. [Google Scholar] [CrossRef] [PubMed]
- Sigoillot, F.D.; Kotsis, D.H.; Masko, E.M.; Bame, M.; Evans, D.R.; Evans, H.I. Protein kinase C modulates the up-regulation of the pyrimidine biosynthetic complex, CAD, by MAP kinase. Front. Biosci. 2007, 12, 3892–3898. [Google Scholar] [CrossRef] [Green Version]
- Koch, J.; Mayr, J.A.; Alhaddad, B.; Rauscher, C.; Bierau, J.; Kovacs-Nagy, R.; Coene, K.L.; Bader, I.; Holzhacker, M.; Prokisch, H.; et al. CAD mutations and uridine-responsive epileptic encephalopathy. Brain 2017, 140, 279–286. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Morcillo, M.; Grande-García, A.; Ruiz-Ramos, A.; del Caño-Ochoa, F.; Boskovic, J.; Ramón-Maiques, S. Structural Insight into the Core of CAD, the Multifunctional Protein Leading De Novo Pyrimidine Biosynthesis. Structure 2017, 25, 912–923.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sigoillot, F.D.; Berkowski, J.A.; Sigoillot, S.M.; Kotsis, D.H.; Guy, H.I. Cell Cycle-dependent Regulation of Pyrimidine Biosynthesis. J. Biol. Chem. 2003, 278, 3403–3409. [Google Scholar] [CrossRef] [Green Version]
- Hervé, G. Structural Insight into the Core of CAD. Structure 2017, 25, 819–820. [Google Scholar] [CrossRef] [Green Version]
- Lipscomb, W.N.; Kantrowitz, E.R. Structure and Mechanisms of Escherichia coli Aspartate Transcarbamoylase. Accounts Chem. Res. 2011, 45, 444–453. [Google Scholar] [CrossRef]
- Grande-García, A.; Lallous, N.; Díaz-Tejada, C.; Ramón-Maiques, S. Structure, Functional Characterization, and Evolution of the Dihydroorotase Domain of Human CAD. Structure 2014, 22, 185–198. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.; Chan, C.W.; Graham, S.C.; Christopherson, R.I.; Guss, J.M.; Maher, M.J. Structures of Ligand-free and Inhibitor Complexes of Dihydroorotase from Escherichia coli: Implications for Loop Movement in Inhibitor Design. J. Mol. Biol. 2007, 370, 812–825. [Google Scholar] [CrossRef]
- Löffler, M.; Carrey, E.A.; Knecht, W. The pathway to pyrimidines: The essential focus on dihydroorotate dehydrogenase, the mitochondrial enzyme coupled to the respiratory chain. Nucleosides Nucleotides Nucleic Acids 2020, 39, 1281–1305. [Google Scholar] [CrossRef]
- Okesli, A.; Khosla, C.; Bassik, M.C. Human pyrimidine nucleotide biosynthesis as a target for antiviral chemotherapy. Curr. Opin. Biotechnol. 2017, 48, 127–134. [Google Scholar] [CrossRef]
- Wittmann, J.G.; Heinrich, D.; Gasow, K.; Frey, A.; Diederichsen, U.; Rudolph, M.G. Structures of the Human Orotidine-5′-Monophosphate Decarboxylase Support a Covalent Mechanism and Provide a Framework for Drug Design. Structure 2008, 16, 82–92. [Google Scholar] [CrossRef] [Green Version]
- Deans, R.M.; Morgens, D.W.; Ökesli, A.; Pillay, S.; Horlbeck, M.A.; Kampmann, M.; Gilbert, L.; Li, A.; Mateo, R.; Smith, M.; et al. Parallel shRNA and CRISPR-Cas9 screens enable antiviral drug target identification. Nat. Chem. Biol. 2016, 12, 361–366. [Google Scholar] [CrossRef] [Green Version]
- Ladner, J.E.; Kitchell, J.P.; Honzatko, R.B.; Ke, H.M.; Volz, K.W.; Kalb, A.J.; Ladner, R.C.; Lipscomb, W.N. Gross quaternary changes in aspartate carbamoyltransferase are induced by the binding of N-(phosphonacetyl)-L-aspartate: A 3.5-A resolution study. Proc. Natl. Acad. Sci. USA 1982, 79, 3125–3128. [Google Scholar] [CrossRef] [Green Version]
- Thoden, J.B.; Holden, H.M.; Wesenberg, G.; Raushel, F.M.; Rayment, I. Structure of carbamoyl phosphate synthetase: A journey of 96 A from substrate to product. Biochemistry 1997, 36, 6305–6316. [Google Scholar] [CrossRef]
- Thoden, J.B.; Phillips, G.N.; Neal, T.M.; Raushel, F.M.; Holden, H.M. Molecular Structure of Dihydroorotase: A Paradigm for Catalysis through the Use of a Binuclear Metal Center. Biochemistry 2001, 40, 6989–6997. [Google Scholar] [CrossRef]
- Ruiz-Ramos, A.; Velazquez-Campoy, A.; Grande-García, A.; Moreno-Morcillo, M.; Ramón-Maiques, S. Structure and Functional Characterization of Human Aspartate Transcarbamoylase, the Target of the Anti-tumoral Drug PALA. Structure 2016, 24, 1081–1094. [Google Scholar] [CrossRef] [Green Version]
- Serre, V.; Penverne, B.; Souciet, J.L.; Potier, S.; Guy, H.; Evans, D.; Vicart, P.; Herve, G. Integrated allosteric regulation in the S. cerevisiae carbamylphosphate synthetase—Aspartate transcarbamylase multifunctional protein. BMC Biochem. 2004, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Carrey, E.A. Phosphorylation, allosteric effectors and inter-domain contacts in CAD; their role in regulation of early steps of pyrimidine biosynthesis. Biochem. Soc. Trans. 1993, 21, 191–195. [Google Scholar] [CrossRef] [Green Version]
- Coxam, B.; Neyt, C.; Grassini, D.R.; Le Guen, L.; Smith, K.A.; Schulte-Merker, S.; Hogan, B.M. carbamoyl-phosphate synthetase 2, aspartate transcarbamylase, and dihydroorotase (cad) regulates Notch signaling and vascular development in zebrafish. Dev. Dyn. 2015, 244, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Cammer, W.; Downing, M. Localization of the multifunctional protein CAD in astrocytes of rodent brain. J. Histochem. Cytochem. 1991, 39, 695–700. [Google Scholar] [CrossRef] [Green Version]
- Cotruvo, J.A.; Stubbe, J. Class I Ribonucleotide Reductases: Metallocofactor Assembly and Repair In Vitro and In Vivo. Annu. Rev. Biochem. 2011, 80, 733–767. [Google Scholar] [CrossRef] [Green Version]
- Martin, E.; Palmic, N.; Sanquer, S.; Lenoir, C.; Hauck, F.; Mongellaz, C.; Fabrega, S.; Nitschké, P.; Degli Esposti, M.; Schwartzentruber, J.; et al. CTP synthase 1 deficiency in humans reveals its central role in lymphocyte proliferation. Nature 2014, 510, 288–292. [Google Scholar] [CrossRef] [PubMed]
- Mollick, T.; Laín, S. Modulating pyrimidine ribonucleotide levels for the treatment of cancer. Cancer Metab. 2020, 8, 1–11. [Google Scholar] [CrossRef]
- Carreras, C.W.; Santi, D.V. The catalytic mechanism and structure of thymidylate synthase. Annu. Rev. Biochem. 1995, 64, 721–762. [Google Scholar] [CrossRef]
- Rd, S.G. Pyrimidine biosynthesis. Am. J. Clin. Nutr. 1979, 32, 1290. [Google Scholar]
- Huang, M.; Graves, L.M. De novo synthesis of pyrimidine nucleotides; emerging interfaces with signal transduction pathways. Experientia 2003, 60, 321–336. [Google Scholar] [CrossRef]
- Teoh, S.T.; Ogrodzinski, M.P.; Lunt, S.Y. UDP-glucose 6-dehydrogenase knockout impairs migration and decreases in vivo metastatic ability of breast cancer cells. Cancer Lett. 2020, 492, 21–30. [Google Scholar] [CrossRef]
- Wolfe, A.L.; Zhou, Q.; Toska, E.; Galeas, J.; Ku, A.A.; Koche, R.P.; Bandyopadhyay, S.; Scaltriti, M.; Lebrilla, C.B.; McCormick, F.; et al. UDP-glucose pyrophosphorylase 2, a regulator of glycogen synthesis and glycosylation, is critical for pancreatic cancer growth. Proc. Natl. Acad. Sci. USA 2021, 118, e2103592118. [Google Scholar] [CrossRef]
- Ng, B.G.; Wolfe, L.A.; Ichikawa, M.; Markello, T.; He, M.; Tifft, C.J.; Gahl, W.A.; Freeze, H.H. Biallelic mutations in CAD, impair de novo pyrimidine biosynthesis and decrease glycosylation precursors. Hum. Mol. Genet. 2015, 24, 3050–3057. [Google Scholar] [CrossRef] [Green Version]
- Kent, C.; Carman, G.M. Interactions among pathways for phosphatidylcholine metabolism, CTP synthesis and secretion through the Golgi apparatus. Trends Biochem. Sci. 1999, 24, 146–150. [Google Scholar] [CrossRef]
- Wang, L.; Pooler, A.M.; Albrecht, M.A.; Wurtman, R.J. Dietary Uridine-5’-Monophosphate Supplementation Increases Potassium-Evoked Dopamine Release and Promotes Neurite Outgrowth in Aged Rats. J. Mol. Neurosci. 2005, 27, 137–146. [Google Scholar] [CrossRef]
- Cansev, M.; Watkins, C.J.; van der Beek, E.M.; Wurtman, R.J. Oral uridine-5′-monophosphate (UMP) increases brain CDP-choline levels in gerbils. Brain Res. 2005, 1058, 101–108. [Google Scholar] [CrossRef]
- López-Coviella, I.; Agut, J.; Savci, V.; Ortiz, J.A.; Wurtman, R.J. Evidence that 5′-Cytidinediphosphocholine Can Affect Brain Phospholipid Composition by Increasing Choline and Cytidine Plasma Levels. J. Neurochem. 2002, 65, 889–894. [Google Scholar] [CrossRef] [PubMed]
- Bailey, C.J. Orotic aciduria and uridine monophosphate synthase: A reappraisal. J. Inherit. Metab. Dis. 2009, 32, 227–233. [Google Scholar] [CrossRef]
- Zhu, A.; Hopkins, K.M.; Friedman, R.A.; Bernstock, J.D.; Broustas, C.G.; Lieberman, H.B. DNMT1 and DNMT3B regulate tumorigenicity of human prostate cancer cells by controlling RAD9 expression through targeted methylation. Carcinogenesis 2020, 42, 220–231. [Google Scholar] [CrossRef]
- Panigrahi, S.K.; Broustas, C.G.; Cuiper, P.Q.; Virk, R.K.; Lieberman, H.B. FOXP1 and NDRG1 act differentially as downstream effectors of RAD9-mediated prostate cancer cell functions. Cell. Signal. 2021, 86, 110091. [Google Scholar] [CrossRef]
- Chen, K.-Y.; Chen, C.-C.; Chang, Y.-C.; Chang, M.-C. Resveratrol induced premature senescence and inhibited epithelial-mesenchymal transition of cancer cells via induction of tumor suppressor Rad9. PLoS ONE 2019, 14, e0219317. [Google Scholar] [CrossRef] [Green Version]
- Lindsey-Boltz, L.A.; Wauson, E.M.; Graves, L.M.; Sancar, A. The human Rad9 checkpoint protein stimulates the carbamoyl phosphate synthetase activity of the multifunctional protein CAD. Nucleic Acids Res. 2004, 32, 4524–4530. [Google Scholar] [CrossRef] [Green Version]
- Nakashima, A.; Kawanishi, I.; Eguchi, S.; Yu, E.H.; Eguchi, S.; Oshiro, N.; Yoshino, K.-I.; Kikkawa, U.; Yonezawa, K. Association of CAD, a multifunctional protein involved in pyrimidine synthesis, with mLST8, a component of the mTOR complexes. J. Biomed. Sci. 2013, 20, 24. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Akasu, H.; Shimono, W.; Matsu, C.; Fujiwara, Y.; Shibagaki, Y.; Heard, J.J.; Tamanoi, F.; Hattori, S. Rheb Protein Binds CAD (Carbamoyl-phosphate Synthetase 2, Aspartate Transcarbamoylase, and Dihydroorotase) Protein in a GTP- and Effector Domain-dependent Manner and Influences Its Cellular Localization and Carbamoyl-phosphate Synthetase (CPSase) Activity. J. Biol. Chem. 2015, 290, 1096–1105. [Google Scholar] [CrossRef] [Green Version]
- Ben-Sahra, I.; Howell, J.J.; Asara, J.M.; Manning, B.D. Stimulation of de Novo Pyrimidine Synthesis by Growth Signaling Through mTOR and S6K1. Science 2013, 339, 1323–1328. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Chen, S.; Yu, W.; Chi, Z.; Wang, Z.; Xu, T.; Zhang, J.; Jiang, D.; Guo, Y.; Fang, H.; et al. AKT controls NLRP3 inflammasome activation by inducing DDX3X phosphorylation. FEBS Lett. 2021. online ahead of print. [Google Scholar] [CrossRef]
- Hu, X.; Wang, D.; Sun, L.; Gao, Y.; Zhou, D.; Tong, X.; Li, J.; Lin, H.; Qing, Y.; Du, S.; et al. Disturbed mitochondrial acetylation in accordance with the availability of acetyl groups in hepatocellular carcinoma. Mitochondrion 2021, 60, 150–159. [Google Scholar] [CrossRef]
- Gama, R.R.; Arantes, L.M.R.B.; Sorroche, B.P.; De Marchi, P.; Melendez, M.E.; Carvalho, R.S.; de Lima, M.A.; Vettore, A.L.; Carvalho, A.L. Evaluation of acetylation and methylation in oral rinse of patients with head and neck cancer history exposed to valproic acid. Sci. Rep. 2021, 11, 16415. [Google Scholar] [CrossRef]
- Chen, Y.T.; Xiang, D.; Zhao, X.Y.; Chu, X.Y. Upregulation of lncRNA NIFK-AS1 in hepatocellular carcinoma by m6A methylation promotes disease progression and sorafenib resistance. Hum. Cell 2021, 1–12. [Google Scholar] [CrossRef]
- Cho, H.J.; Ryu, K.-J.; Baek, K.E.; Lim, J.; Kim, T.; Song, C.Y.; Yoo, J.; Lee, J.Y.A.H.G. Cullin 3/KCTD5 Promotes the Ubiqutination of Rho Guanine Nucleotide Dissociation Inhibitor 1 and Regulates Its Stability. J. Microbiol. Biotechnol. 2020, 30, 1488–1494. [Google Scholar] [CrossRef]
- Chen, H.; Venkat, S.; McGuire, P.; Gan, Q.; Fan, C. Recent Development of Genetic Code Expansion for Posttranslational Modification Studies. Molecules 2018, 23, 1662. [Google Scholar] [CrossRef] [Green Version]
- Van Damme, P.; Lasa, M.; Polevoda, B.; Gazquez, C.; Elosegui-Artola, A.; Kim, D.S.; De-Juan-Pardo, E.; Demeyer, K.; Hole, K.; Larrea, E.; et al. N-terminal acetylome analyses and functional insights of the N-terminal acetyltransferase NatB. Proc. Natl. Acad. Sci. USA 2012, 109, 12449–12454. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, C.; Kumar, C.; Gnad, F.; Nielsen, M.L.; Rehman, M.; Walther, T.C.; Olsen, J.V.; Mann, M. Lysine Acetylation Targets Protein Complexes and Co-Regulates Major Cellular Functions. Science 2009, 325, 834–840. [Google Scholar] [CrossRef] [Green Version]
- Bruserud, O.; Stapnes, C.; Tronstad, K.J.; Ryningen, A.; Ånensen, N.; Gjertsen, B.T. Protein lysine acetylation in normal and leukaemic haematopoiesis: HDACs as possible therapeutic targets in adult AML. Expert Opin. Ther. Targets 2006, 10, 51–68. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Ning, F.; Cai, Y.; Sheng, H.; Zheng, R.; Yin, X.; Lu, Z.; Su, L.; Chen, X.; Zeng, C.; et al. The EGFR-P38 MAPK axis up-regulates PD-L1 through miR-675-5p and down-regulates HLA-ABC via hexokinase-2 in hepatocellular carcinoma cells. Cancer Commun. 2021, 41, 62–78. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.M.; Xu, M.; Sargen, M.; Jang, H.; Zhang, M.; Zhang, T.; Zhu, B.; Jones, K.; Kim, J.; Mendoza, L.; et al. Novel MAPK/AKT-impairing germline NRAS variant identified in a melanoma-prone family. Fam. Cancer 2021, 1–9. [Google Scholar] [CrossRef]
- Sigoillot, F.D.; Kotsis, D.H.; Serre, V.; Sigoillot, S.M.; Evans, D.R.; Guy, H.I. Nuclear localization and mitogen-activated protein kinase phosphorylation of the multifunctional protein CAD. J. Biol. Chem. 2005, 280, 25611–25620. [Google Scholar] [CrossRef] [Green Version]
- Cobb, M.H.; Goldsmith, E.J. How MAP kinases are regulated. J. Biol. Chem. 1995, 270, 14843–14846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graves, L.M.; Guy, H.I.; Kozlowski, P.; Huang, M.; Lazarowski, E.; Pope, R.M.; Collins, M.A.; Dahlstrand, E.N.; Earp, H.S., 3rd; Evans, D.R. Regulation of carbamoyl phosphate synthetase by MAP kinase. Nature 2000, 403, 328–332. [Google Scholar] [CrossRef]
- Evans, D.R.; Guy, H.I. Mammalian Pyrimidine Biosynthesis: Fresh Insights into an Ancient Pathway. J. Biol. Chem. 2004, 279, 33035–33038. [Google Scholar] [CrossRef] [Green Version]
- Carrey, E.A.; Campbell, D.G.; Hardie, D.G. Phosphorylation and activation of hamster carbamyl phosphate synthetase II by cAMP-dependent protein kinase. A novel mechanism for regulation of pyrimidine nucleotide biosynthesis. EMBO J. 1985, 4, 3735–3742. [Google Scholar] [CrossRef]
- Kolch, W.; Heidecker, G.; Kochs, G.; Hummel, R.; Vahidi, H.; Mischak, H.; Finkenzeller, G.; Marme, D.; Rapp, U.R. Protein kinase C alpha activates RAF-1 by direct phosphorylation. Nature 1993, 364, 249–252. [Google Scholar] [CrossRef]
- Kotsis, D.H.; Masko, E.M.; Sigoillot, F.D.; Di Gregorio, R.; Guy-Evans, H.I.; Evans, D.R. Protein kinase A phosphorylation of the multifunctional protein CAD antagonizes activation by the MAP kinase cascade. Mol. Cell. Biochem. 2007, 301, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Saxton, R.A.; Sabatini, D.M. mTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linke, M.; Fritsch, S.D.; Sukhbaatar, N.; Hengstschläger, M.; Weichhart, T. mTORC1 and mTORC2 as regulators of cell metabolism in immunity. FEBS Lett. 2017, 591, 3089–3103. [Google Scholar] [CrossRef]
- Ardestani, A.; Lupse, B.; Kido, Y.; Leibowitz, G.; Maedler, K. mTORC1 Signaling: A Double-Edged Sword in Diabetic β Cells. Cell Metab. 2018, 27, 314–331. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, J.; Zhou, Q.; Liu, Y.; Chen, W.; Xu, H. mTORC1 signaling is essential for neurofibromatosis type I gene modulated osteogenic differentiation of BMSCs. J. Cell. Biochem. 2018, 120, 2886–2896. [Google Scholar] [CrossRef]
- Laplante, M.; Sabatini, D.M. mTOR Signaling in Growth Control and Disease. Cell 2012, 149, 274–293. [Google Scholar] [CrossRef] [Green Version]
- Wullschleger, S.; Loewith, R.; Hall, M.N. TOR Signaling in Growth and Metabolism. Cell 2006, 124, 471–484. [Google Scholar] [CrossRef] [Green Version]
- Avruch, J.; Hara, K.; Lin, Y.; Liu, M.; Long, X.; Ortizvega, S.; Yonezawa, K. Insulin and amino-acid regulation of mTOR signaling and kinase activity through the Rheb GTPase. Oncogene 2006, 25, 6361–6372. [Google Scholar] [CrossRef] [Green Version]
- Vetter, I.R.; Wittinghofer, A. The guanine nucleotide-binding switch in three dimensions. Science 2001, 294, 1299–1304. [Google Scholar] [CrossRef] [Green Version]
- Wyant, G.A.; Abu-Remaileh, M.; Wolfson, R.L.; Chen, W.; Freinkman, E.; Danai, L.V.; Heiden, M.G.V.; Sabatini, D.M. mTORC1 Activator SLC38A9 Is Required to Efflux Essential Amino Acids from Lysosomes and Use Protein as a Nutrient. Cell 2017, 171, 642–654.e12. [Google Scholar] [CrossRef]
- Robitaille, A.M.; Christen, S.; Shimobayashi, M.; Cornu, M.; Fava, L.L.; Moes, S.; Prescianotto-Baschong, C.; Sauer, U.; Jenoe, P.; Hall, M.N. Quantitative Phosphoproteomics Reveal mTORC1 Activates de Novo Pyrimidine Synthesis. Science 2013, 339, 1320–1323. [Google Scholar] [CrossRef]
- Wang, X.; Yang, K.; Wu, Q.; Kim, L.J.Y.; Morton, A.R.; Gimple, R.C.; Prager, B.C.; Shi, Y.; Zhou, W.; Bhargava, S.; et al. Targeting pyrimidine synthesis accentuates molecular therapy response in glioblastoma stem cells. Sci. Transl. Med. 2019, 11, eaau4972. [Google Scholar] [CrossRef]
- Zoncu, R.; Bar-Peled, L.; Efeyan, A.; Wang, S.; Sancak, Y.; Sabatini, D.M. mTORC1 Senses Lysosomal Amino Acids Through an Inside-Out Mechanism That Requires the Vacuolar H+-ATPase. Science 2011, 334, 678–683. [Google Scholar] [CrossRef] [Green Version]
- Chernova, O.B.; Chernov, M.V.; Ishizaka, Y.; Agarwal, M.L.; Stark, G.R. MYC abrogates p53-mediated cell cycle arrest in N-(phosphonacetyl)-L-aspartate-treated cells, permitting CAD gene amplification. Mol. Cell. Biol. 1998, 18, 536–545. [Google Scholar] [CrossRef] [Green Version]
- Abadjian, M.-C.Z.; Edwards, W.B.; Anderson, C.J. Imaging the Tumor Microenvironment. Adv. Exp. Med. Biol. 2017, 1036, 229–257. [Google Scholar] [CrossRef]
- Wang, H.; Wang, X.; Xu, L.; Zhang, J.; Cao, H. High expression levels of pyrimidine metabolic rate–limiting enzymes are adverse prognostic factors in lung adenocarcinoma: A study based on The Cancer Genome Atlas and Gene Expression Omnibus datasets. Purinergic Signal. 2020, 16, 347–366. [Google Scholar] [CrossRef]
- Ridder, D.A.; Schindeldecker, M.; Weinmann, A.; Berndt, K.; Urbansky, L.; Witzel, H.R.; Heinrich, S.; Roth, W.; Straub, B.K. Key Enzymes in Pyrimidine Synthesis, CAD and CPS1, Predict Prognosis in Hepatocellular Carcinoma. Cancers 2021, 13, 744. [Google Scholar] [CrossRef]
- Rabinovich, S.; Adler, L.; Yizhak, K.; Sarver, A.; Silberman, A.; Agron, S.; Stettner, N.; Sun, Q.; Brandis, A.; Helbling, D.; et al. Diversion of aspartate in ASS1-deficient tumours fosters de novo pyrimidine synthesis. Nature 2015, 527, 379–383. [Google Scholar] [CrossRef]
- Mac, S.M.; Farnham, P.J. CAD, a c-Myc target gene, is not deregulated in Burkitt’s lymphoma cell lines. Mol. Carcinog. 2000, 27, 84–96. [Google Scholar] [CrossRef]
- Palm, W.; Park, Y.; Wright, K.; Pavlova, N.N.; Tuveson, D.A.; Thompson, C.B. The Utilization of Extracellular Proteins as Nutrients Is Suppressed by mTORC1. Cell 2015, 162, 259–270. [Google Scholar] [CrossRef] [Green Version]
- Cox, J.A.; LaMora, A.; Johnson, S.L.; Voigt, M.M. Novel role for carbamoyl phosphate synthetase 2 in cranial sensory circuit formation. Int. J. Dev. Neurosci. 2013, 33, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Cameron, J.S.; Moro, F.; Simmonds, H.A. Gout, uric acid and purine metabolism in paediatric nephrology. Pediatr. Nephrol. 1993, 7, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Lafita-Navarro, M.C.; Venkateswaran, N.; Kilgore, J.A.; Kanji, S.; Han, J.; Barnes, S.; Williams, N.S.; Buszczak, M.; Burma, S.; Conacci-Sorrell, M. Inhibition of the de novo pyrimidine biosynthesis pathway limits ribosomal RNA transcription causing nucleolar stress in glioblastoma cells. PLoS Genet. 2020, 16, e1009117. [Google Scholar] [CrossRef]
- Fairbanks, L.D.; Bofill, M.; Ruckemann, K.; Simmonds, H.A. Importance of ribonucleotide availability to proliferating T-lymphocytes from healthy humans. Disproportionate expansion of pyrimidine pools and contrasting effects of de novo synthesis inhibitors. J. Biol. Chem. 1995, 270, 29682–29689. [Google Scholar] [CrossRef] [Green Version]
- Aoki, T.; Wéber, G. Carbamoyl phosphate synthetase (glutamine-hydrolyzing): Increased activity in cancer cells. Science 1981, 212, 463–465. [Google Scholar] [CrossRef]
- Sigoillot, F.D.; Sigoillot, S.M.; Guy, H.I. Breakdown of the regulatory control of pyrimidine biosynthesis in human breast cancer cells. Int. J. Cancer 2004, 109, 491–498. [Google Scholar] [CrossRef]
- Martinez-Reyes, I.; Cardona, L.R.; Kong, H.; Vasan, K.; McElroy, G.S.; Werner, M.; Kihshen, H.; Reczek, C.R.; Weinberg, S.E.; Gao, P.; et al. Mitochondrial ubiquinol oxidation is necessary for tumour growth. Nature 2020, 585, 288–292. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Stark, G.R.; Hoogenraad, N.J. Inhibition by N-(Phosphonacetyl)-l-Aspartate of Aspartate Transcarbamylase Activity and Drug-induced Cell Proliferation in Mice. J. Biol. Chem. 1974, 249, 6951–6955. [Google Scholar] [CrossRef]
- Loh, E.; Kufe, D.W. Synergistic effects with inhibitors of de novo pyrimidine synthesis, acivicin, and N-(phosphonacetyl)-L-aspartic acid. Cancer Res. 1981, 41, 3419–3423. [Google Scholar]
- Peters, G.J.; Sharma, S.L.; Laurensse, E.; Pinedo, H.M. Inhibition of pyrimidine de novo synthesis by DUP-785 (NSC 368390). Investig. New Drugs 1987, 5, 235–244. [Google Scholar] [CrossRef]
- Jin, L.; Li, Y.; Pu, F.; Wang, H.; Zhang, D.; Bai, J.; Shang, Y.; Ma, Z.; Ma, X.-X. Inhibiting pyrimidine biosynthesis impairs Peste des Petits Ruminants Virus replication through depletion of nucleoside pools and activation of cellular immunity. Vet. Microbiol. 2021, 260, 109186. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Luo, T.; Ding, X.; Chang, Y.; Liu, C.; Zhang, Y.; Hao, S.; Yin, Q.; Jiang, B. Inhibition of mitochondrial complex III induces differentiation in acute myeloid leukemia. Biochem. Biophys. Res. Commun. 2021, 547, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Zhang, Z.; Dai, Y.; Liu, S.; Fu, E. Brequinar inhibits enterovirus replication by targeting biosynthesis pathway of pyrimidines. Am. J. Transl. Res. 2020, 12, 8247–8255. [Google Scholar]
- Ladds, M.; van Leeuwen, I.M.M.; Drummond, C.J.; Chu, S.; Healy, A.R.; Popova, G.; Pastor Fernandez, A.; Mollick, T.; Darekar, S.; Sedimbi, S.K.; et al. A DHODH inhibitor increases p53 synthesis and enhances tumor cell killing by p53 degradation blockage. Nat. Commun. 2018, 9, 1107. [Google Scholar] [CrossRef] [Green Version]
- Del Cano-Ochoa, F.; Ramon-Maiques, S. Deciphering CAD: Structure and function of a mega-enzymatic pyrimidine factory in health and disease. Protein Sci. 2021, 30, 1995–2008. [Google Scholar] [CrossRef] [PubMed]
- Grohmann, K.; Lauffer, H.; Lauenstein, P.; Hoffmann, G.F.; Seidlitz, G. Hereditary Orotic Aciduria with Epilepsy and without Megaloblastic Anemia. Neuropediatrics 2015, 46, 123–125. [Google Scholar] [CrossRef] [PubMed]
- Harden, K.K.; Robinson, J.L. Deficiency of UMP synthase in dairy cattle: A model for hereditary orotic aciduria. J. Inherit. Metab. Dis. 1987, 10, 201–209. [Google Scholar] [CrossRef]
- Ng, S.B.; Buckingham, K.J.; Lee, C.; Bigham, A.W.; Tabor, H.K.; Dent, K.M.; Huff, C.D.; Shannon, P.T.; Jabs, E.; Nickerson, D.A.; et al. Exome sequencing identifies the cause of a mendelian disorder. Nat. Genet. 2009, 42, 30–35. [Google Scholar] [CrossRef] [Green Version]
- del Caño-Ochoa, F.; Ng, B.G.; Abedalthagafi, M.; Almannai, M.; Cohn, R.D.; Costain, G.; Elpeleg, O.; Houlden, H.; Karimiani, E.G.; Liu, P.; et al. Cell-based analysis of CAD variants identifies individuals likely to benefit from uridine therapy. Genet. Med. 2020, 22, 1598–1605. [Google Scholar] [CrossRef]
- Sesma, J.I.; Weitzer, C.D.; Livraghi-Butrico, A.; Dang, H.; Donaldson, S.; Alexis, N.E.; Jacobson, K.A.; Harden, T.K.; Lazarowski, E.R. UDP-glucose promotes neutrophil recruitment in the lung. Purinergic Signal. 2016, 12, 627–635. [Google Scholar] [CrossRef] [Green Version]
- Arase, T.; Uchida, H.; Kajitani, T.; Ono, M.; Tamaki, K.; Oda, H.; Nishikawa, S.; Kagami, M.; Nagashima, T.; Masuda, H.; et al. The UDP-Glucose Receptor P2RY14 Triggers Innate Mucosal Immunity in the Female Reproductive Tract by Inducing IL-8. J. Immunol. 2009, 182, 7074–7084. [Google Scholar] [CrossRef] [Green Version]
- Werts, C.; Girardin, S.E.; Philpott, D.J. TIR, CARD and PYRIN: Three domains for an antimicrobial triad. Cell Death Differ. 2006, 13, 798–815. [Google Scholar] [CrossRef]
- Abdelnaby, H.; Ndiaye, N.C.; D’Amico, F.; Fouad, A.M.; Hassan, S.; Elshafey, A.; Al Hashash, W.; Faisal, M.; Alshamali, Y.; Al-Taweel, T.; et al. NOD2/CARD15 polymorphisms (P268S, IVS8+158, G908R, L1007fs, R702W) among Kuwaiti patients with Crohn’s disease: A case-control study. Saudi J. Gastroenterol. 2021, 27, 249. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, J.C.; Damen, M.S.M.A.; Oosting, M.; De Jong, D.J.; Heinhuis, B.; Gomes, R.S.; Araújo, C.S.; Netea, M.G.; Ribeiro-Dias, F.; Joosten, L.A.B. The NOD2 receptor is crucial for immune responses towards New World Leishmania species. Sci. Rep. 2017, 7, 15219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Bruyn, M.; Vermeire, S. NOD2 and bacterial recognition as therapeutic targets for Crohn’s disease. Expert Opin. Ther. Targets 2017, 21, 1123–1139. [Google Scholar] [CrossRef] [PubMed]
- Rolhion, N.; Barnich, N.; Claret, L.; Darfeuille-Michaud, A. Strong decrease in invasive ability and outer membrane vesicle release in Crohn’s disease-associated adherent-invasive Escherichia coli strain LF82 with the yfgL gene deleted. J. Bacteriol. 2005, 187, 2286–2296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eitel, J.; Krull, M.; Hocke, A.C.; N’Guessan, P.D.; Zahlten, J.; Schmeck, B.; Slevogt, H.; Hippenstiel, S.; Suttorp, N.; Opitz, B. Beta-PIX and Rac1 GTPase mediate trafficking and negative regulation of NOD2. J. Immunol. 2008, 181, 2664–2671. [Google Scholar] [CrossRef] [Green Version]
- von Kampen, O.; Lipinski, S.J.; Till, A.; Martin, S.; Nietfeld, W.; Lehrach, H.; Schreiber, S.; Rosenstiel, P. Caspase Recruitment Domain-containing Protein 8 (CARD8) Negatively Regulates NOD2-mediated Signaling. J. Biol. Chem. 2010, 285, 19921–19926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pashenkov, M.V.; Dagil, Y.A.; Pinegin, B.V. NOD1 and NOD2: Molecular targets in prevention and treatment of infectious diseases. Int. Immunopharmacol. 2018, 54, 385–400. [Google Scholar] [CrossRef]
- Oda, F.; Murakami, M.; Hanakawa, Y.; Tohyama, M.; Nakano, N.; Nishikomori, R.; Sayama, K.; Kambe, N. A sporadic case of granulomatous disease negative for NOD2 mutations and mimicking Blau syndrome. Clin. Exp. Dermatol. 2017, 43, 57–58. [Google Scholar] [CrossRef]
- Sun, J.; Wang, X.; Zhang, X.; Zhang, W. Identification of a novel missense mutation in the NOD2 gene in a Chinese child with early-onset sarcoidosis. Indian J. Dermatol. Venereol. Leprol. 2018, 84, 645. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.Y.; Pang, J.C.; Lu, M.X.; Yang, X.L.; Zhu, H.P.; Ke, X.L.; Liu, Z.G.; Cao, J.M.; Wang, M. Molecular characterization, expression and functional analysis of NOD1, NOD2 and NLRC3 in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2018, 73, 207–219. [Google Scholar] [CrossRef]
- Richmond, A.L.; Kabi, A.; Homer, C.R.; Marina-Garcia, N.; Nickerson, K.P.; Nesvizhskii, A.I.; Sreekumar, A.; Chinnaiyan, A.M.; Nunez, G.; McDonald, C. The nucleotide synthesis enzyme CAD inhibits NOD2 antibacterial function in human intestinal epithelial cells. Gastroenterology 2012, 142, 1483–1492.e1486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jatana, S.; Homer, C.R.; Madajka, M.; Ponti, A.K.; Kabi, A.; Papay, F.; McDonald, C. Pyrimidine synthesis inhibition enhances cutaneous defenses against antibiotic resistant bacteria through activation of NOD2 signaling. Sci. Rep. 2018, 8, 8708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redzic, Z.B.; Malatiali, S.A.; Craik, J.D.; Rakic, M.L.; Isakovic, A.J. Blood–Brain Barrier Efflux Transport of Pyrimidine Nucleosides and Nucleobases in the Rat. Neurochem. Res. 2008, 34, 566–573. [Google Scholar] [CrossRef]
- Pesini, A.; Iglesias, E.; Bayona-Bafaluy, M.P.; Garrido-Pérez, N.; Meade, P.; Gaudó, P.; Jiménez-Salvador, I.; Benito, P.A.; Montoya, J.; Ferrer, I.; et al. Brain pyrimidine nucleotide synthesis and Alzheimer disease. Aging 2019, 11, 8433–8462. [Google Scholar] [CrossRef]
- Bourget, P.A.; Tremblay, G.C. Pyrimidine biosynthesis in rat brain. J. Neurochem. 1972, 19, 1617–1624. [Google Scholar] [CrossRef]
- Piccoli, F.; Camarda, R.; Bonavita, V. Purine and pyrimidine nucleotides in the brain of normal and convulsant rats. J. Neurochem. 1969, 16, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Moussavi, Z.; Koski, L.; Fitzgerald, P.B.; Millikin, C.; Lithgow, B.; Jafari-Jozani, M.; Wang, X. Repeated Transcranial Magnetic Stimulation for Improving Cognition in Alzheimer Disease: Protocol for an Interim Analysis of a Randomized Controlled Trial. JMIR Res. Protoc. 2021, 10, e31183. [Google Scholar] [CrossRef]
- Pozdnyakov, D.I.; Miroshnichenko, K.A.; Voronkov, A.V.; Kovaleva, T.G. The Administration of the New Pyrimidine Derivative-4-{2-[2-(3,4-Dimethoxyphenyl)-Vinyl]-6-Ethyl-4-Oxo-5-Phenyl-4H-Pyrimid ine-1-Il}Benzsulfamide Restores the Activity of Brain Cells in Experimental Chronic Traumatic Encephalopathy by Maintaining Mitochondrial Function. Medicina 2019, 55, 386. [Google Scholar]
- Singh, S.; Dhanawat, M.; Gupta, S.; Kumar, D.; Kakkar, S.; Nair, A.; Verma, I.; Sharma, P. Naturally Inspired Pyrimidines Analogues for Alzheimer’s Disease. Curr. Neuropharmacol. 2020, 19, 136–151. [Google Scholar] [CrossRef]
- Lai, M.K.P.; Tan, M.G.; Kirvell, S.; Hobbs, C.; Lee, J.; Esiri, M.M.; Chen, C.P.; Francis, P.T. Selective loss of P2Y2 nucleotide receptor immunoreactivity is associated with Alzheimer’s disease neuropathology. J. Neural Transm. 2008, 115, 1165–1172. [Google Scholar] [CrossRef] [PubMed]
- Pooler, A.M.; Guez, D.H.; Benedictus, R.; Wurtman, R.J. Uridine enhances neurite outgrowth in nerve growth factor-differentiated PC12 [corrected]. Neuroscience 2005, 134, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Wurtman, R.J.; Ulus, I.H.; Cansev, M.; Watkins, C.J.; Wang, L.; Marzloff, G. Synaptic proteins and phospholipids are increased in gerbil brain by administering uridine plus docosahexaenoic acid orally. Brain Res. 2006, 1088, 83–92. [Google Scholar] [CrossRef] [PubMed]
- O’Dwyer, P.J.; Manola, J.; Valone, F.H.; Ryan, L.M.; Hines, J.D.; Wadler, S.; Haller, D.G.; Arbuck, S.G.; Weiner, L.M.; Mayer, R.J.; et al. Fluorouracil modulation in colorectal cancer: Lack of improvement with N -phosphonoacetyl- l -aspartic acid or oral leucovorin or interferon, but enhanced therapeutic index with weekly 24-hour infusion schedule--an Eastern Cooperative Oncology Group/Cancer and Leukemia Group B Study. J. Clin. Oncol. 2001, 19, 2413–2421. [Google Scholar] [PubMed]
- Christopherson, R.I.; Lyons, S.D.; Wilson, P.K. Inhibitors of de Novo Nucleotide Biosynthesis as Drugs. Accounts Chem. Res. 2002, 35, 961–971. [Google Scholar] [CrossRef]
- Zheng, X.; Yang, X.; Lin, J.; Song, F.; Shao, Y. Low curcumin concentration enhances the anticancer effect of 5-fluorouracil against colorectal cancer. Phytomedicine 2021, 85, 153547. [Google Scholar] [CrossRef]
- Muggia, F.M.; Tsiatis, A.A.; O’Connell, M.J.; Glick, J.H.; Opfell, R.W.; Coren, A. Phase II trial of PALA in lymphoma: An Eastern Cooperative Oncology Group study. Cancer Treat. Rep. 1984, 68, 551–553. [Google Scholar]
- Kleeberg, U.R.; Mulder, J.H.; Rümke, P.; Thomas, D.; Rozencweig, M. N-(Phosphonacetyl)-l-aspartate (PALA) in advanced malignant melanoma: A phase II trial of the EORTC malignant melanoma cooperative group. Eur. J. Cancer Clin. Oncol. 1982, 18, 723–726. [Google Scholar] [CrossRef]
- Santana-Codina, N.; Roeth, A.A.; Zhang, Y.; Yang, A.; Mashadova, O.; Asara, J.M.; Wang, X.; Bronson, R.T.; Lyssiotis, C.A.; Ying, H.; et al. Oncogenic KRAS supports pancreatic cancer through regulation of nucleotide synthesis. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Koundinya, M.; Sudhalter, J.; Courjaud, A.; Lionne, B.; Touyer, G.; Bonnet, L.; Menguy, I.; Schreiber, I.; Perrault, C.; Vougier, S.; et al. Dependence on the Pyrimidine Biosynthetic Enzyme DHODH Is a Synthetic Lethal Vulnerability in Mutant KRAS-Driven Cancers. Cell Chem. Biol. 2018, 25, 705–717.e11. [Google Scholar] [CrossRef] [Green Version]
- Bardeleben, C.; Sharma, S.; Reeve, J.R.; Bassilian, S.; Frost, P.; Hoang, B.; Shi, Y.; Lichtenstein, A. Metabolomics Identifies Pyrimidine Starvation as the Mechanism of 5-Aminoimidazole-4-Carboxamide-1--Riboside-Induced Apoptosis in Multiple Myeloma Cells. Mol. Cancer Ther. 2013, 12, 1310–1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmermann, B.H.; Kemling, N.M.; Evans, D.R. Function of Conserved Histidine Residues in Mammalian Dihydroorotase. Biochemistry 1995, 34, 7038–7046. [Google Scholar] [CrossRef] [PubMed]
- Bindu, P.S.; Sonam, K.; Govindaraj, P.; Govindaraju, C.; Chiplunkar, S.; Nagappa, M.; Kumar, R.; Vekhande, C.C.; Arvinda, H.R.; Gayathri, N.; et al. Outcome of epilepsy in patients with mitochondrial disorders: Phenotype genotype and magnetic resonance imaging correlations. Clin. Neurol. Neurosurg. 2018, 164, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Xu, H.; Wang, T.; Wu, Y. A Patient With CAD Deficiency Responsive to Uridine and Literature Review. Front. Neurol. 2020, 11, 64. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tumor or Model Type | Inhibitors | Target | Clinical Traits (+) or Not (−) | References |
|---|---|---|---|---|
| Colorectal cancer | N-phosphonoacetyl-l-aspartic acid leucovorin | ATCase | − | [135,136,137] |
| Lymphoma Melanoma | N-(phosphonacetyl)-L-aspartate (PALA) | ATCase | +, terminated, Clinical phase II | [138,139] |
| Myeloid leukemia | ASLAN003 | DHODH | +, processing in clinical phase II | [35] |
| Myeloid malignancies | BAY 2402234 | DHODH | +, terminated, clinical phase II | [140,141] |
| Myeloma cell lines | AICAr | UMPS | − | [142] |
| central nervous system disorder (multiple sclerosis) | Teriflunomide | DHODH | + | [82] |
| Effect on Function of CAD or Related Clinical Phenotypes | Variations in CAD | Mechanism | Location | References |
|---|---|---|---|---|
| CAD loss catalytic activity | p.His1471Ala/p. His1473Ala | Active site zinc mutants, no zinc-binding | DHOase | [19,143] |
| p.His1590Asn/p. His1614Asn | Alter the coordination of Zn-βcoordinating residue | |||
| p.Cys1613Ser | Alter the coordination of Zn-γ coordinating residue | |||
| P.Thr1562Ala/p. Thr1563Ala | Disturb and make the structure of CAD unstable | |||
| p.Asp1686Asn | Affect the coordination of Zn-α | |||
| Activity of CAD decreased nearly 100-fold compared to wild-type, CAD catalytic activity↓ | p.Glu1637Thr | Alter the coordination of Zn-γ coordinating residue | DHOase | [19] |
| 11.5% of wild-type catalytic activity of DHOase, CAD catalytic activity↓ | p.His1642Asn | Bind similar amounts of zinc compared to wild type, but a 3-fold increase of Km | DHOase | [143] |
| 2.9% of wild-type catalytic activity of DHOase, CAD catalytic activity↓ | p.His1690Asn | A 9-fold increase of Km, pH dependence in both the degradative and the biosynthetic decreased | DHOase | [143] |
| CAD catalytic activity↓, PMA-induced Thr-456 phosphorylation↓ | p.Ser1873Ala | AlterPKC phosphorylation site | - | [13] |
| CAD mutations, neurometabolic disorder (Seizures, developmental delay, etc.) | c.98T > G | Main inducement of epileptic encephalopathy | GATase | [14,144,145] |
| c.1843-3C > T | Affect the splice acceptor site of intron 12 | CPSIIase.B | ||
| c.5365C > T | Polypeptide missed at the last 438 of 2225 amino acids of CAD protein | DHOase | ||
| Biallelic mutations in CAD, de novo pyrimidine biosynthesis↓, glycosylation precursors↓ | c.1843-1G > A | In-frame deletion of exon 13 | CPSIIase.B | [41] |
| c.6071G > A | Carbamoyl-phosphate binding↓ | DHOase |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, G.; Li, D.; Wang, T.; He, S. Pyrimidine Biosynthetic Enzyme CAD: Its Function, Regulation, and Diagnostic Potential. Int. J. Mol. Sci. 2021, 22, 10253. https://doi.org/10.3390/ijms221910253
Li G, Li D, Wang T, He S. Pyrimidine Biosynthetic Enzyme CAD: Its Function, Regulation, and Diagnostic Potential. International Journal of Molecular Sciences. 2021; 22(19):10253. https://doi.org/10.3390/ijms221910253
Chicago/Turabian StyleLi, Guanya, Dunhui Li, Tao Wang, and Shanping He. 2021. "Pyrimidine Biosynthetic Enzyme CAD: Its Function, Regulation, and Diagnostic Potential" International Journal of Molecular Sciences 22, no. 19: 10253. https://doi.org/10.3390/ijms221910253
APA StyleLi, G., Li, D., Wang, T., & He, S. (2021). Pyrimidine Biosynthetic Enzyme CAD: Its Function, Regulation, and Diagnostic Potential. International Journal of Molecular Sciences, 22(19), 10253. https://doi.org/10.3390/ijms221910253